Introduction
Thyroid cancer (THCA) is one of the most common
types of endocrine cancer worldwide (1,2).
Accumulating evidence has indicated that numerous biomarkers are
involved in THCA prognosis, proliferation and radiation resistance.
For example, Huang et al (3)
observed that downregulation of microRNA (miR)-125a-5p promoted
glucose metabolism by upregulating CD147 in THCA. Furthermore, Choi
et al (4) demonstrated that
prospero homeobox 1 activates Wnt/β-catenin signaling, and induces
the expression of serpin family A member 1 and fatty acid binding
protein 4 in papillary THCA. However, the mechanisms underlying
THCA progression have not been fully elucidated.
Long noncoding RNAs (lncRNAs) are a type of RNA
transcript that are >200 bp in length, but which have no protein
coding potential (5). Previous
studies have revealed that lncRNAs are dysregulated in a number of
human diseases, and are involved in regulating various biological
processes, including RNA stabilization (6,7), X
chromosome inactivation (8,9), transcription and protein translation
(10). Several reports have
described how lncRNAs serve important roles in THCA. For example, a
recent study demonstrated that nuclear-enriched abundant transcript
1_2 is able to ‘sponge’ (or competitively inhibit) miR-106b-5p to
promote the expression of ATPase family AAA domain containing 2 in
papillary THCA (11). The lncRNA
CNALPTC1 acts as a competing endogenous RNA (ceRNA) by sponging the
miR-30 family in THCA (12). Several
studies have demonstrated that lncRNA expression is associated with
the prognosis of patients with THCA (13–15). Li
et al (16) identified 111
lncRNAs that were differentially expressed between tumor and normal
samples in THCA. Furthermore, the lncRNA growth arrest specific 5
was significantly associated with Tumor-Node-Metastasis (TNM)
staging, lymph node metastasis, disease-free survival (DFS) and
overall survival in THCA (17).
Small nucleolar RNA host gene 7 (SNHG7) is a novel
lncRNA involved in the progression of different types of human
cancer. lncRNA SNHG7 was first reported to be an oncogenic gene in
lung cancer, increasing the expression of Fas apoptotic inhibitory
molecule 2 (18). Wang et al
(19) demonstrated that lncRNA SNHG7
promotes the proliferation and inhibits apoptosis of gastric cancer
cells by repressing P15 and P16 expression. However, the clinical
value and functional role of SNHG7 in THCA has not been fully
elucidated. The present study investigated the expression pattern
and molecular functions of SNHG7 in THCA.
Materials and methods
Analysis of The Cancer Genome Atlas
(TCGA) and Gene Expression Omnibus (GEO) datasets
The public TCGA (cancergenome.nih.gov) database was used to analyze the
expression levels of SNHG7 in THCA. The clinical information of the
THCA TCGA dataset was generated from cBioPortal (www.cbioportal.org) (20,21). The
2009 TNM classification of the American Joint Committee on
Cancer/International Union Against Cancer was used to stage all the
patients (22). The survival
analysis, comparing patients with THCA with a high expression level
of SNHG7 (SNHG7-high) and those with a low expression level of
SNHG7 (SNHG7-low), was performed. In the present study, the web
application Cutoff Finder 2012 (molpath.charite.de/cutoff) was used to identify the
cut-off value (median expression value) to divide the THCA samples
into SNHG7-high and SNHG7-low groups. In order to validate TCGA
analysis, two GEO datasets, GSE50901, samples obtained from
adjacent healthy tissues from patients with THCA (23), and GSE33630, obtained from healthy
controls (24,25), were analyzed. The normalized data
were downloaded from the GEO repository (https://www.ncbi.nlm.nih.gov/gds). The normal samples
in the TCGA and GSE50901 datasets were adjacent normal tissue
samples taken from patients with thyroid cancer. The normal samples
in GSE33630 were taken from control patients.
Construction of SNHG7-mediated ceRNA
networks
In the present study, SNHG7- targeting microRNAs and
mRNAs were predicted using Starbase version 3.0 (http://starbase.sysu.edu.cn/index.php).
Subsequently, co-expression analysis was performed for SNHG7 in the
TCGA THCA dataset using the cBioPortal online software. The
SNHG7-target pairs with co-expression coefficient >0.3 were
selected for the construction of SNHG7-associated ceRNA networks.
The Cytoscape software version 3.4.0 (www.cytoscape.org) was used for visualization of the
co-expression networks.
Co-expression network construction and
analysis
According to the expression value of the Pearson's
correlation coefficient of SNHG7-gene pairs, SNHG7 co-expressed
genes were identified in the present study. The co-expressed
SNHG7-gene pairs with an absolute value of Pearson's correlation
coefficient ≥0.3 were selected, which included the top 1,000 genes
that were co-expressed with SNHG7 in thyroid cancer samples, and
the co-expression network was established by using the Cytoscape
software.
Protein-protein interaction (PPI)
network construction and analysis
By using the Search Tool for the Retrieval of
Interacting Genes/Proteins database version 11.0 (STRING #11.0)
(26), a PPI network mediated by
SNHG7 was constructed using its co-expressing mRNAs (combined
score, >0.4). Subsequently, a module analysis of the network was
performed using the Mcode plugin (27). The PPI network with the cut-offs of
degree ≥2 and of nodes ≥2-core were selected as hub networks.
Gene Ontology (GO) and Kyoto
Encyclopedia of Genes and Genomes (KEGG) pathway analysis
In the present study, GO and KEGG pathway analyses
were performed by using the Molecule Annotation System version 3.0
(bioinfo.capitalbio.com/mas3).
P<0.05 was considered to indicate a statistically significant
difference.
Cell culture and cell
transfection
The thyroid gland undifferentiated (anaplastic)
carcinoma CAL62 and the thyroid gland squamous SW579 carcinoma cell
lines were purchased from the American Type Culture Collection. The
cells were incubated in RPMI-1640 medium (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% fetal bovine serum (ExCell
Bio) in an incubator containing 5% CO2 at 37°C. The
small interfering (siRNA) for SNHG7 (5′-UUAGCAGAGUAAUUUGCACUU-3′)
and siRNA for the negative control (siNC;
5′-UAGCGACUAAACACAUCAA-3′) were purchased from Guangzhou RiboBio
Co., Ltd. The cells were seeded at a density of 3×105
cells/well in 6-well plates. Following 12 h of incubation in an
incubator containing 5% CO2 at 37°C, the cells were
transfected with 50 nM siNC or siSNHG7 using FuGENE® 6
transfection reagent (Promega Corporation). The transfected cells
were then used to perform the cell proliferation assay and cell
cycle assay.
Cell proliferation assay
Cell proliferation of the transfected cells was
detected using the Cell Counting Kit-8 (CCK-8) assay. In brief, the
cells were seeded at a density of 1,000 cells/well in the medium in
a 96-well plate and cell proliferation was measured at 0, 24, 48
and 72 h. The cells were incubated in an incubator containing 5%
CO2 at 37°C with 10 µl of CCK-8 solution (Dojindo
Molecular Technologies, Inc.) in each well for 2 h. The absorbance
at 450 nm was subsequently measured using a microplate reader.
Cell cycle assay
Transfected cells were collected 48 h after
transfection, and then the cells were fixed with pre-cooled 70%
ethanol, and incubated overnight at 4°C. Subsequently, the cells
were washed once with 1 ml PBS, followed by incubation in the dark
with 500 µl PBS containing 50 µg/ml propidium iodide, 0.2% Triton
X-100 and 100 µg/ml RNase A for 30 min at 4°C. The cell cycle was
analyzed using a flow cytometer and the Modfit LT version 4.1
software (Verity Software House, Inc.).
Reverse transcription-quantitative
(RT-q)PCR assay
Total RNA was extracted using the
E.Z.N.A.® Total RNA kit (Omega Bio-Tek, Inc.) 48 h
following transfection of the SW579 and CAL62 cell lines. RT was
performed using the PrimeScrip RT reagent Kit (Takara Biotechnology
Co., Ltd.) according to the manufacturer's instructions.
Temperature conditions for the RT reaction were: 37°C for 15 min;
and for reverse transcriptase inactivation reaction, 85°C for 5
sec. RT-qPCR was performed using the Hieff qPCR SYBR-Green Master
mix (YeaSen) according to the manufacturer's instructions.
Temperature protocol for pre-denaturation was: 95°C for 5 min; for
PCR reaction (35 cycles), 95°C for 5 sec and 60°C for 30 sec.
β-actin was used as the internal control. The relative expression
was calculated by the 2−ΔΔCq method (28) and every experiment was performed in
triplicates. The primer sequences were as follows: SNHG7 forward,
5′-CGATACCATTGAACACGCTGC-3′ andreverse, 5′-GGTTGAGGGTCCCAGTG-3′;
β-actin forward, 5′-CTCCATCCTGGCCTCGCTGT-3′ and reverse,
5′-GCTGTCACCTTCACCGTTCC-3′.
Statistical analysis
Statistical analyses and graphical representations
were performed using SPSS version 17.0 (SPSS, Inc.) and GraphPad
Prism version 6.0 (GraphPad Software, Inc.) Statistical comparisons
between two groups were performed using the t-test or the
Mann-Whitney U test. Statistical comparisons between two paired
groups were performed by the paired t-test. For analysis of >2
groups, one-way analysis of variance followed by Newman-Keuls post
hoc test was used. Survival curves were plotted using the
Kaplan-Meier method and the differences were examined using the
log-rank test. Experiments were repeated at least three times.
P<0.05 was considered to indicate a significantly significant
difference.
Results
lncRNA SNHG7 is upregulated in
THCA
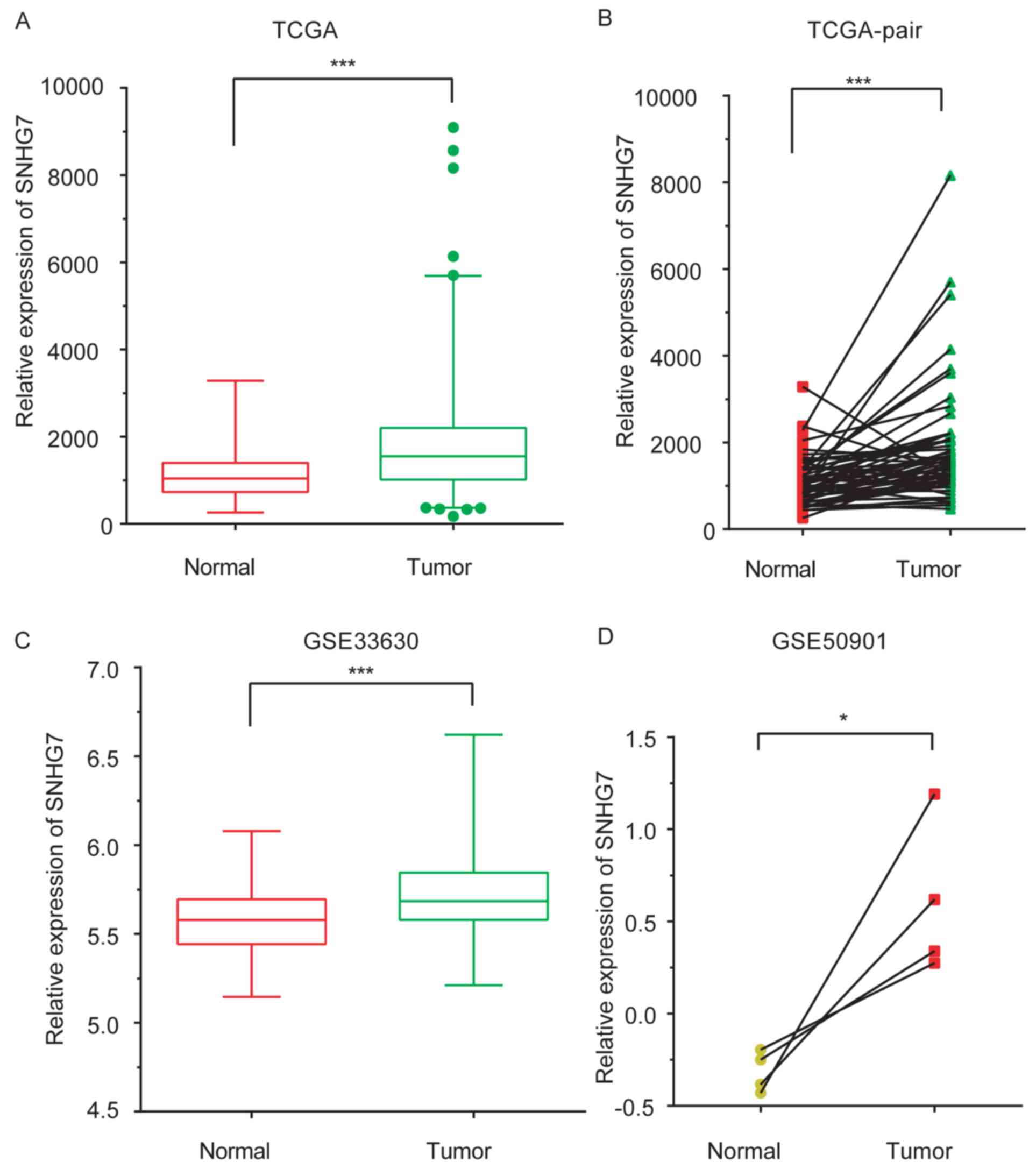
In the present study, SNHG7 expression levels in
samples from patients with THCA were compared with those in normal
samples by analyzing the THCA TCGA dataset. A total of 56 normal
samples and 578 tumor samples were included in the TCGA dataset. As
presented in Fig. 1A and B, SNHG7
was observed to be markedly upregulated in tumor samples compared
with control samples (P<0.001). In order to validate TCGA
analysis, two GEO datasets, GSE50901 (samples obtained from
adjacent healthy tissues from patients with THCA) (23), and GSE33630 (obtained from healthy
controls) (24,25), were analyzed. SNHG7 expression levels
were increased in THCA samples compared with normal samples
(P<0.001 and P<0.05; Fig. 1C and
D).
High expression of SNHG7 is associated
with progression of thyroid cancer
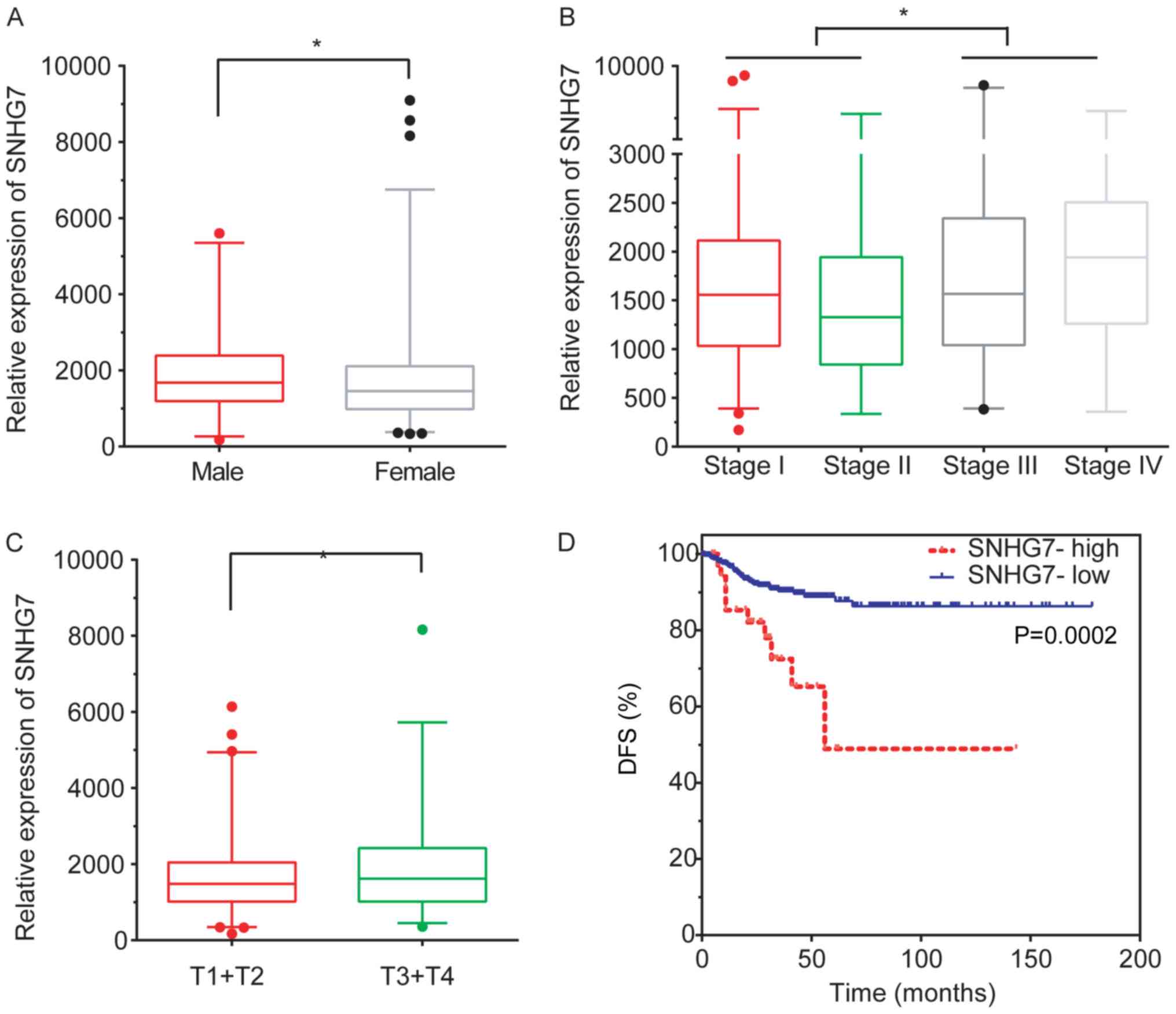
Subsequently, the association between SNHG7
expression levels and clinicopathological features, including age,
sex, grade, T and N stage and the recurrence status, were
evaluated. The analysis revealed that SNHG7 was downregulated in
female patients with THCA compared with male patients with THCA
(P<0.05; Fig. 2A). Furthermore,
SNHG7 expression was increased in patients with stage III and stage
IV compared with patients with stage I (P<0.05) and stage II
(P<0.01) THCA (P<0.05; Fig.
2B).
SNHG7 expression was revealed to be significantly
upregulated in stage T3 and T4 patients compared with stage T1 and
T2 patients (P<0.05; Fig. 2C).
This analysis demonstrated that an increased lncRNA SNHG7
expression level was associated with advanced pathology stages in
THCA.
lncRNA SNHG7 is associated with poor
prognosis in THCA
To further investigate the clinical significance of
lncRNA SNHG7 in THCA, SNHG7 expression and survival time in
patients with THCA was subsequently analyzed. The results obtained
demonstrated that a higher expression level of SNHG7 was closely
associated with shorter DFS time in thyroid cancer (P<0.001;
median DFS time, 56.11 and 67.22 months for the high and
low-expression groups, respectively; Fig. 2D), suggesting that SNHG7 may serve as
a biomarker in THCA.
Construction of SNHG7 mediated PPI
networks in THCA
By using the ‘guilt-by-association’ approach, the
potential roles of SNHG7 should be similar with its downstream
targets. Thus, we predicted potential roles of SNHG7 using
SNHG7-regulating mRNAs. Co-expression analysis for SNHG7 in THCA
was subsequently performed. The absolute values of Pearson's
correlation coefficient ≥0.3 were selected as the cut-off to
identify reliable SNHG7-mRNA pairs.
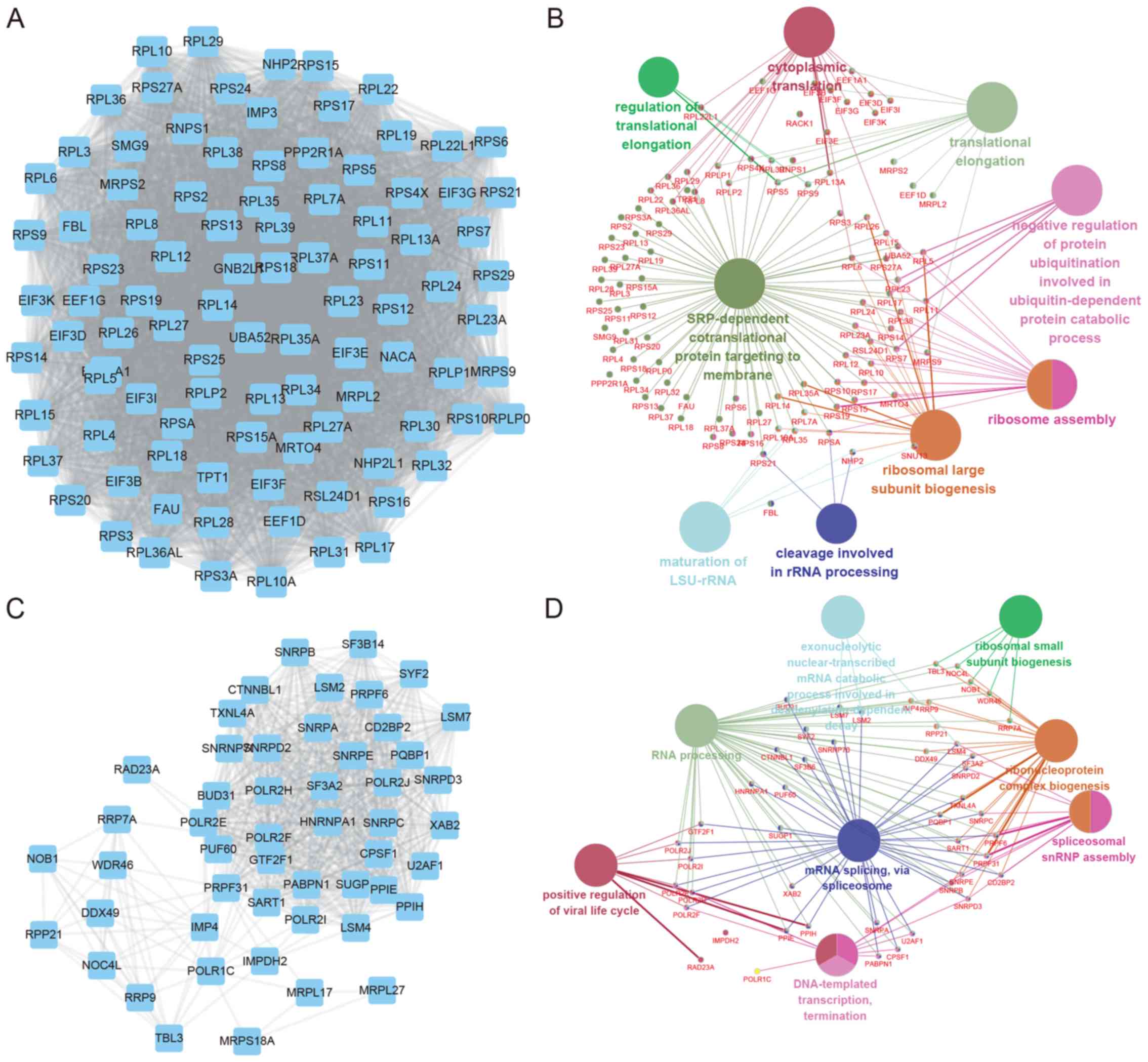
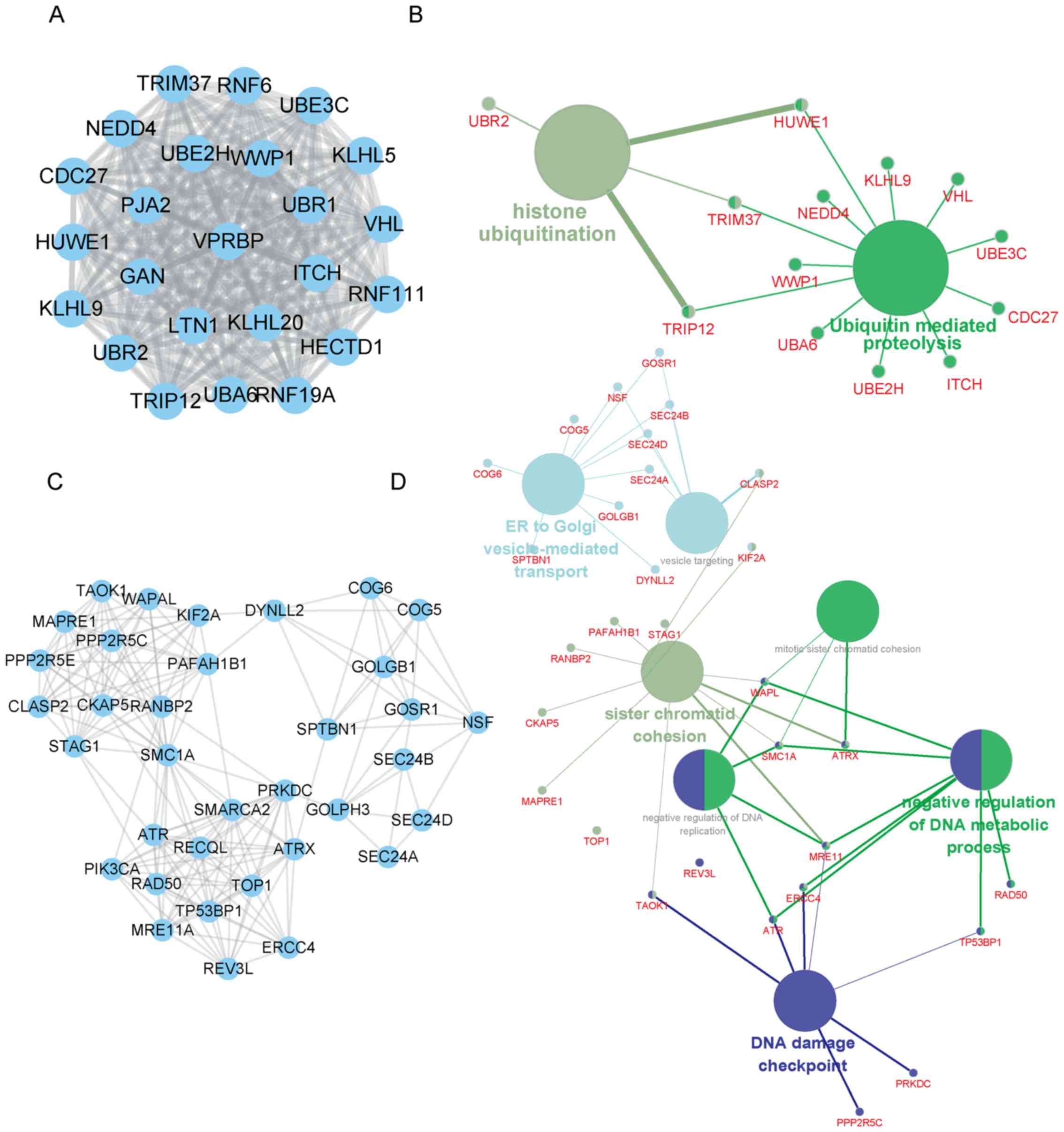
In addition, SNHG7 mediated PPI networks in thyroid
cancer were also constructed. The top two positively co-expressed
hub modules are presented in Fig. 3,
and the top two negatively co-expressed hub modules are presented
in Fig. 4. Positive-related module 1
contained 97 nodes and 4,438 edges, whereas positive-related module
2 contained 51 nodes and 697 edges. Furthermore, negative-related
module 1 contained 24 nodes and 276 edges, and negative-related
module 2 contained 35 nodes and 182 edges.
GO and KEGG pathway analysis of SNHG7
mediated hub PPI networks in THCA
GO and KEGG pathway analysis of SNHG7 mediated hub
PPI networks in thyroid cancer were subsequently performed using
Cytoscape's ClueGo plug-in. Significant biological processes and
pathways (P<0.05) are presented in the present study. The
results revealed that SNHG7 positive-related module 1 was involved
in regulating cytoplasmic translation, translation elongation,
signal-recognition particle-dependent co-translational protein
targeting to membrane and ribosome assembly, whereas SNHG7
positive-related module 2 was associated with positive regulation
of the viral life cycle, RNA processing and mRNA splicing.
Furthermore, SNHG7 negative-related module 1 was
revealed to be involved in regulating histone ubiquitination and
ubiquitin-mediated proteolysis, and SNHG7 negative-related module 1
was revealed to be involved in endoplasmic reticulum (ER)-to-Golgi
vesicle-mediated transport, sister chromatid cohesion and DNA
damage checkpoint regulation.
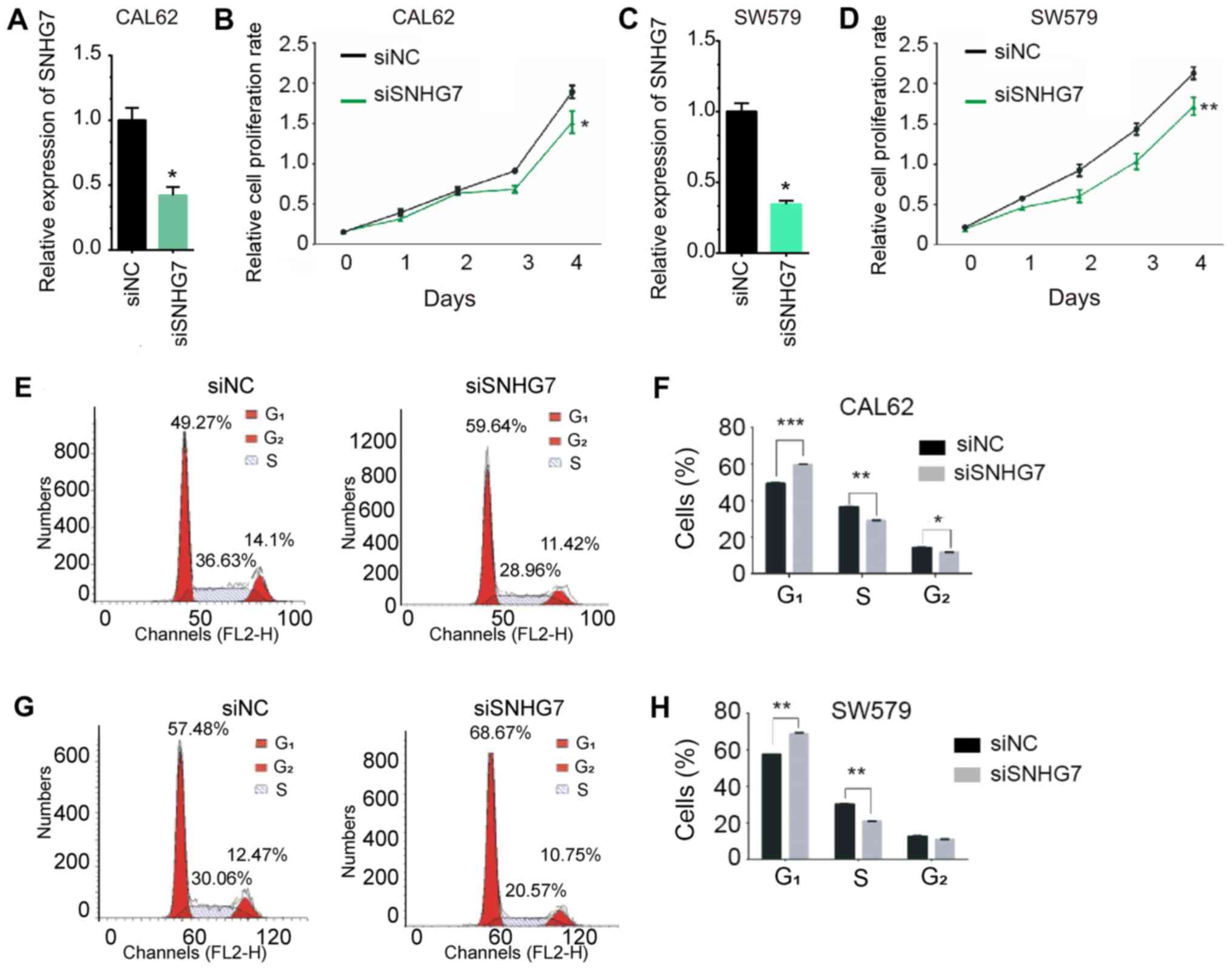
Knockdown of SNHG7 inhibits THCA cell
proliferation and cell cycle in vitro
In order to further validate the roles of SNHG7 in
thyroid cancer, a loss-of-function assay was performed in the CAL62
and SW579 cell lines. As shown Fig. 5A
and C, transfection with siSNHG7 significantly decreased the
mRNA expression of SNHG7 in CAL62 and SW579 cells compared with the
negative control (P<0.05). The CCK-8 assay was performed to
evaluate the effect of SNHG7 knockdown on cell proliferation. As
presented in Fig. 5, knockdown of
SNHG7 significantly inhibited proliferation in CAL62 (P<0.05;
Fig. 5B) and SW579 (P<0.01;
Fig. 5D) cells after 4 days.
As abnormal cell cycle progression is a hallmark of
cancer (29,30), the role of SNHG7 knockdown in
regulating the cell cycle was investigated by flow cytometry in
CAL62 and SW579 cells (Fig. 5E-H).
SNHG7 knockdown increased the percentage of cells in the
G1 phase and decreased the percentage of cells in the S
phase compared with control cells. Furthermore, SNHG7 knockdown
decreased the percentage of cells in G2 phase in the
CAL62 cells compared with control cells, but no significant
difference was observed in the SW579 cells (P<0.01).
Discussion
An increasing number of studies have demonstrated
the lncRNAs serve important roles in various types of human cancer,
including prostate (31,32), breast (33,34),
lung (35,36) and THCA (37). Specific lncRNAs have been reported to
be involved in THCA progression. For example, Pvt1 oncogene was the
first lncRNA revealed to be associated with the regulation of THCA
cell proliferation by increasing the expression of
thyroid-stimulating hormone receptor (38). Xu et al (39) observed that the lncRNAs
ENST00000537266 and ENST00000426615 served roles in papillary THCA
growth and metastasis. H19 imprinted maternally expressed
transcript was found to be a ceRNA by ‘sponging’ miR-17-5p to
regulate the expression of YES proto-oncogene 1 Src family tyrosine
kinase in THCA (40). The present
study revealed, to the best of the best of the authors' knowledge
for the first time, that lncRNA SNHG7 is upregulated in, and
associated with the prognosis of, THCA.
SNHG7 is a novel lncRNA that has been reported to
have an oncogenic role in different types of human cancer. SNHG7 is
upregulated in esophageal (41) and
prostate cancer (42), glioblastoma
(43), non-small cell lung (44) and gastric cancer (19). However, the roles of SNHG7 in THCA
have yet to be elucidated. In the present study, analysis of the
TCGA dataset revealed that SNHG7 was upregulated in THCA tissues
compared with normal samples. Notably, high expression of SNHG7 was
revealed to be associated with the progression of THCA. SNHG7 was
upregulated in advanced-stage (stages III and IV) compared with the
low-stage (stages I and II) THCA samples. Furthermore, the present
study revealed, to the best of best of the authors' knowledge for
the first time, that higher expression levels of SNHG7 were closely
associated with shorter overall survival and DFS times in THCA.
Taken together, these results demonstrated that SNHG7 may serve as
a biomarker for THCA.
Co-expression analysis has been widely used to
identify potential roles of lncRNAs. For example, Chen et al
(45) constructed an lncRNA-mRNA
co-expression network to reveal the roles of lncRNAs in zebra fish.
In the current study, an SNHG7-mediated co-expression network in
thyroid cancer was constructed. The four top hub modules mediated
by SNHG7 were identified in THCA. The present analysis revealed
that SNHG7 is associated with ‘translation’, ‘viral life cycle’,
‘RNA processing’, ‘mRNA splicing’, ‘histone ubiquitination’,
‘ER-to-Golgi vesicle-mediated transport’, ‘sister chromatid
cohesion’ and ‘DNA damage checkpoint regulation’. In order to
validate the roles of SNHG7 in THCA, an in vitro
loss-of-function assay was performed, and the results obtained
revealed that SNHG7 knockdown leads to a marked inhibition of cell
proliferation and cell cycle progression in THCA cell lines. Taken
together, the results obtained in the current study suggest that
SNHG7 is an oncogene in THCA.
In conclusion, the present study has demonstrated
that SNHG7 was markedly upregulated in THCA samples compared with
control samples through an analysis of TCGA datasets. SNHG7
expression levels were increased in advanced-stage compared with
early-stage THCA samples. Additionally, a higher expression level
of SNHG7 was associated with shorter survival times compared with a
lower expression level. The results obtained in the current study
suggest that SNHG7 may be used to predict the prognosis of patients
with THCA. The present study suggested that SNHG7 may serve as a
new therapeutic and prognostic target for THCA.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
LC and LJZ designed the study. LC, LJZ and JZ
developed the methodology. LC, LJZ and JZ analyzed and interpreted
the data, and wrote and revised the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ER
|
endoplasmic reticulum
|
|
lncRNAs
|
long non-coding RNAs
|
|
TNM
|
Tumor-Node-Metastasis
|
|
DFS
|
disease-free survival
|
|
SNHG7
|
small nucleolar RNA host gene 7
|
|
THCA
|
thyroid cancer
|
|
TCGA
|
The Cancer Genome Atlas
|
|
ceRNA
|
competing endogenous RNA
|
References
|
1
|
Chen AY, Jemal A and Ward EM: Increasing
incidence of differentiated thyroid cancer in the United States,
1988–2005. Cancer. 115:3801–3807. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pellegriti G, Frasca F, Regalbuto C,
Squatrito S and Vigneri R: Worldwide increasing incidence of
thyroid cancer: Update on epidemiology and risk factors. J Cancer
Epidemiol. 2013:9652122013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Huang P, Mao LF, Zhang ZP, Lv WW, Feng XP,
Liao HJ, Dong C, Kaluba B, Tang XF and Chang S: Down-Regulated
miR-125a-5p promotes the reprogramming of glucose metabolism and
cell malignancy by increasing levels of CD147 in thyroid cancer.
Thyroid. 28:613–623. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Choi D, Ramu S, Park E, Jung E, Yang S,
Jung W, Choi I, Lee S, Kim KE, Seong YJ, et al: Aberrant activation
of notch signaling inhibits PROX1 activity to enhance the malignant
behavior of thyroid cancer cells. Cancer Res. 76:582–593. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Prensner JR and Chinnaiyan AM: The
emergence of lncRNAs in cancer biology. Cancer Discov. 1:391–407.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang F, Yuan JH, Wang SB, Yang F, Yuan SX,
Ye C, Yang N, Zhou WP, Li WL, Li W and Sun SH: Oncofetal long
noncoding RNA PVT1 promotes proliferation and stem cell-like
property of hepatocellular carcinoma cells by stabilizing NOP2.
Hepatology. 60:1278–1290. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cao C, Sun J, Zhang D, Guo X, Xie L, Li X,
Wu D and Liu L: The long intergenic noncoding RNA UFC1, a target of
MicroRNA 34a, interacts with the mRNA stabilizing protein HuR to
increase levels of β-catenin in HCC cells. Gastroenterology.
148:415–426. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Engreitz JM, Pandya-Jones A, McDonel P,
Shishkin A, Sirokman K, Surka C, Kadri S, Xing J, Goren A, Lander
ES, et al: The Xist lncRNA exploits three-dimensional genome
architecture to spread across the X chromosome. Science.
341:12379732013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang F, Deng X, Ma W, Berletch JB, Rabaia
N, Wei G, Moore JM, Filippova GN, Xu J, Liu Y, et al: The lncRNA
Firre anchors the inactive X chromosome to the nucleolus by binding
CTCF and maintains H3K27me3 methylation. Genome Biol. 16:522015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yoon JH, Abdelmohsen K and Gorospe M:
Posttranscriptional gene regulation by long noncoding RNA. J Mol
Biol. 425:3723–3730. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sun W, Lan X, Zhang H, Wang Z, Dong W, He
L, Zhang T, Zhang P, Liu J and Qin Y: NEAT1_2 functions as a
competing endogenous RNA to regulate ATAD2 expression by sponging
microRNA-106b-5p in papillary thyroid cancer. Cell Death Dis.
9:3802018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen C, Zhou L, Wang H, Chen J, Li W, Liu
W, Shen M, Liu H and Fu X: Long noncoding RNA CNALPTC1 promotes
cell proliferation and migration of papillary thyroid cancer via
sponging miR-30 family. Am J Cancer Res. 8:192–206. 2018.PubMed/NCBI
|
|
13
|
Zhang R, Hardin H, Chen J, Guo Z and Lloyd
RV: Non-coding RNAs in thyroid cancer. Endocr Pathol. 27:12–20.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yang G, Lu X and Yuan L: LncRNA: A link
between RNA and cancer. Biochim Biophys Acta. 1839:1097–1109. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huang JK, Ma L, Song WH, Lu BY, Huang YB,
Dong HM, Ma XK, Zhu ZZ and Zhou R: LncRNA-MALAT1 Promotes
angiogenesis of thyroid cancer by modulating tumor-associated
macrophage FGF2 protein secretion. J Cell Biochem. 118:4821–4830.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li Q, Li H, Zhang L, Zhang C, Yan W and
Wang C: Identification of novel long non-coding RNA biomarkers for
prognosis prediction of papillary thyroid cancer. Oncotarget.
8:46136–46144. 2017.PubMed/NCBI
|
|
17
|
Guo LJ, Zhang S, Gao B, Jiang Y, Zhang XH,
Tian WG, Hao S, Zhao JJ, Zhang G, Hu CY, et al: Low expression of
long non-coding RNA GAS5 is associated with poor prognosis of
patients with thyroid cancer. Exp Mol Pathol. 102:500–504. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
She K, Huang J, Zhou H, Huang T, Chen G
and He J: lncRNA-SNHG7 promotes the proliferation, migration and
invasion and inhibits apoptosis of lung cancer cells by enhancing
the FAIM2 expression. Oncol Rep. 36:2673–2680. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang MW, Liu J, Liu Q, Xu QH, Li TF, Jin S
and Xia TS: LncRNA SNHG7 promotes the proliferation and inhibits
apoptosis of gastric cancer cells by repressing the P15 and P16
expression. Eur Rev Med Pharmacol Sci. 21:4613–4622.
2017.PubMed/NCBI
|
|
20
|
Gao J, Aksoy BA, Dogrusoz U, Dresdner G,
Gross B, Sumer SO, Sun Y, Jacobsen A, Sinha R, Larsson E, et al:
Integrative analysis of complex cancer genomics and clinical
profiles using the cBioPortal. Sci Signal. 6:pl12013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cerami E, Gao J, Dogrusoz U, Gross BE,
Sumer SO, Aksoy BA, Jacobsen A, Byrne CJ, Heuer ML, Larsson E, et
al: The cBio cancer genomics portal: An open platform for exploring
multidimensional cancer genomics data. Cancer Discov. 2:401–404.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Edge SB, Byrd DR, Compton CC, Fritz AG,
Greene FL and Trotti A: AJCC cancer staging manual7th. NY:
Springer; 2010
|
|
23
|
Barros-Filho MC, Marchi FA, Pinto CA,
Rogatto SR and Kowalski LP: High diagnostic accuracy based on
CLDN10, HMGA2, and LAMB3 transcripts in papillary thyroid
carcinoma. J Clin Endocrinol Metab. 100:E890–E899. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tomas G, Tarabichi M, Gacquer D, Hebrant
A, Dom G, Dumont JE, Keutgen X, Fahey TR, Maenhaut C and Detours V:
A general method to derive robust organ-specific gene
expression-based differentiation indices: Application to thyroid
cancer diagnostic. Oncogene. 31:4490–4498. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dom G, Tarabichi M, Unger K, Thomas G,
Oczko-Wojciechowska M, Bogdanova T, Jarzab B, Dumont JE, Detours V
and Maenhaut C: A gene expression signature distinguishes normal
tissues of sporadic and radiation-induced papillary thyroid
carcinomas. Br J Cancer. 107:994–1000. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Szklarczyk D, Gable AL, Lyon D, Junge A,
Wyder S, Huerta-Cepas J, Simonovic M, Doncheva NT, Morris JH, Bork
P, et al: STRING v11: Protein-protein association networks with
increased coverage, supporting functional discovery in genome-wide
experimental datasets. Nucleic Acids Res. 47:D607–D613. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bader GD and Hogue CW: An automated method
for finding molecular complexes in large protein interaction
networks. Bmc Bioinformatics. 4:22003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Williams GH and Stoeber K: The cell cycle
and cancer. J Pathol. 226:352–364. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kastan MB and Bartek J: Cell-cycle
checkpoints and cancer. Nature. 432:316–323. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Misawa A, Takayama KI and Inoue S: Long
non-coding RNAs and prostate cancer. Cancer Sci. 108:2107–2114.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wan X, Huang W, Yang S, Zhang Y, Pu H, Fu
F, Huang Y, Wu H, Li T and Li Y: Identification of
androgen-responsive lncRNAs as diagnostic and prognostic markers
for prostate cancer. Oncotarget. 7:60503–60518. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kumar M, DeVaux RS and Herschkowitz JI:
Molecular and cellular changes in breast cancer and new roles of
lncRNAs in breast cancer initiation and progression. Prog Mol Biol
Transl Sci. 144:563–586. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li T, Liu Y, Xiao H and Xu G: Long
non-coding RNA TUG1 promotes cell proliferation and metastasis in
human breast cancer. Breast Cancer. 24:535–543. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Park SM, Choi EY, Bae DH, Sohn HA, Kim SY
and Kim YJ: The LncRNA EPEL promotes lung cancer cell proliferation
through E2F target activation. Cell Physiol Biochem. 45:1270–1283.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tao H, Yang JJ, Zhou X, Deng ZY, Shi KH
and Li J: Emerging role of long noncoding RNAs in lung cancer:
Current status and future prospects. Respir Med. 110:12–19. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kim D, Lee WK, Jeong S, Seol MY, Kim H,
Kim KS, Lee EJ, Lee J and Jo YS: Upregulation of long noncoding RNA
LOC100507661 promotes tumor aggressiveness in thyroid cancer. Mol
Cell Endocrinol. 431:36–45. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhou Q, Chen J, Feng J and Wang J: Long
noncoding RNA PVT1 modulates thyroid cancer cell proliferation by
recruiting EZH2 and regulating thyroid-stimulating hormone receptor
(TSHR). Tumour Biol. 37:3105–3113. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Xu B, Shao Q, Xie K, Zhang Y, Dong T, Xia
Y and Tang W: The long non-coding RNA ENST00000537266 and
ENST00000426615 influence papillary thyroid cancer cell
proliferation and motility. Cell Physiol Biochem. 38:368–378. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu L, Yang J, Zhu X, Li D, Lv Z and Zhang
X: Long noncoding RNA H19 competitively binds miR-17-5p to regulate
YES1 expression in thyroid cancer. FEBS J. 283:2326–2339. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xu LJ, Yu XJ, Wei B, Hui HX, Sun Y, Dai J
and Chen XF: LncRNA SNHG7 promotes the proliferation of esophageal
cancer cells and inhibits its apoptosis. Eur Rev Med Pharmacol Sci.
22:2653–2661. 2018.PubMed/NCBI
|
|
42
|
Qi H, Wen B, Wu Q, Cheng W, Lou J, Wei J,
Huang J, Yao X and Weng G: Long noncoding RNA SNHG7 accelerates
prostate cancer proliferation and cycle progression through cyclin
D1 by sponging miR-503. Biomed Pharmacother. 102:326–332. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ren J, Yang Y, Xue J, Xi Z, Hu L, Pan SJ
and Sun Q: Long noncoding RNA SNHG7 promotes the progression and
growth of glioblastoma via inhibition of miR-5095. Biochem Biophys
Res Commun. 496:712–718. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
She K, Yan H, Huang J, Zhou H and He J:
miR-193b availability is antagonized by LncRNA-SNHG7 for
FAIM2-induced tumour progression in non-small cell lung cancer.
Cell Prolif. Nov 12–2017.(Epub ahead of print). doi:
10.1111/cpr.12406. PubMed/NCBI
|
|
45
|
Chen W, Zhang X, Li J, Huang S, Xiang S,
Hu X and Liu C: Comprehensive analysis of coding-lncRNA gene
co-expression network uncovers conserved functional lncRNAs in
zebrafish. Bmc Genomics. 19 (Suppl 2):S1122018. View Article : Google Scholar
|