Introduction
Colon cancer is one of the most common malignancies
in the world and one of the leading causes of cancer-associated
deaths (1–3). Colon cancer ranks second among the most
common types of cancer in China (4,5). The
5-year average survival rate of patients with colon cancer is 50%,
and the 5-year survival rate of advanced high-grade colon cancer
patients is less than 10% (1–4). At
present, the primary therapeutic method for treating patients with
colon cancer is surgery combined with adjuvant chemotherapy
(5,6). Oxaliplatin, 5-fluorouracil and
capecitabine are the primary chemotherapeutic drugs used to treat
colon cancer (4,6). Nevertheless, recurrence and multidrug
resistance reduce the survival rate of patients with colon cancer
(3,7,8).
Researchers have demonstrated that multiple mechanisms are involved
in the initiation, development, proliferation and metastasis of
colon cancer, including the upregulation of oncogenes and
downregulation of tumor suppressor genes, and dysregulated
expression of miRNAs or other noncoding RNAs (9–11).
However, the potential mechanism underlying human colon cancer
development remains unclear. Additional studies will facilitate an
improved understanding of colon cancer and identify potential
targets for new treatments.
USP22 is a ubiquitin-specific protease that belongs
to the deubiquitinase family of proteins (12,13). In
human tissues, USP22 is primarily expressed in the brain, skeletal
muscle and heart tissues, and it is expressed at lower levels in
the liver and lung (12,14). USP22 is one subunit of SAGA
(Spt-Ada-Gcn5-acetyltransferase), and is involved in the
deubiquitination of histones H2A and H2B and the acetylation of
histone H4 in order to regulate the transcription and expression of
numerous genes (15,16). USP22 participates in a number of
physiological processes, including regulating the cell cycle, cell
growth, cell differentiation and cell signal transduction (17–19).
USP22 has also been demonstrated to be involved in the c-myc and
p53 signaling pathways (17).
Recently, USP22 has been reported to serve an important role in a
number of different types of cancer, including gastric cancer
(19), hepatocellular carcinoma
(20), non-small-cell lung cancer
(21) and breast cancer (22). However, the function of USP22 in
human colon cancer has not been demonstrated.
In the present study, the protein expression levels
of USP22 in 80 colon cancer tissues and 64 normal colon tissues
were determined, and the association between USP22 expression and
clinicopathological features was analyzed. Furthermore, the effects
of USP22 on colon cancer cell proliferation and metastasis were
evaluated through cell functional assays. Finally, the molecular
mechanism by which USP22 promotes biological behavior in colon
cancer cells was examined.
Materials and methods
Clinical tissue samples
A total of 80 paraffin-embedded colon cancer tissues
and 64 normal colon tissues were obtained from 86 male patients and
58 female patients aged 61.2 years (range, 45–78 years) at the
First Affiliated Hospital of Anhui Medical University (Hefei,
China) between January 2013 and December 2014. These patients with
colon cancer were followed up for at least 5 years, and their
clinicopathological parameters were collected. Patients with colon
cancer and another disease or who had undergone special therapies
prior to surgery were excluded. The clinicopathological parameters
of these colon cancer patients were determined and confirmed based
on WHO systems (23). Every patient
provided written informed consent, and the study was approved by
the Biomedical Ethics Committee of Anhui Medical University.
Immunohistochemistry (IHC)
USP22 expression in paraffin embedded sections (4 µm
thick) of human colon cancer tissues and in normal colon tissues
were obtained from the hospital pathology department having been
fixed in 10% neutral formalin for 24 h at room temperature and
paraffin embedded. The samples were examined by IHC. A two-step
histostaining method (Fuzhou Maixin Biotech Co., Ltd.) was used for
IHC as described previously (24).
Samples were incubated with a USP22 antibody (cat. no. LS-C99567;
1:200; LifeSpan BioSciences, Inc.) for 4 h at room temperature.
Subsequently, the samples were incubated with a horseradish
peroxidase-conjugated universal detection reagent (Fuzhou Maixin
Biotech Co., Ltd.) for 30 min at room temperature.
3,3′-Diaminobenzidine tetrahydrochloride (Fuzhou Maixin Biotech
Co., Ltd.) was used for detection. The stained sections were
observed and scored under a light microscope (magnification, ×400;
Olympus Corporation). Positive signals of USP22 protein in the
cells were primarily located in the nucleus and stained dark brown.
Sections with ≥10% USP22-positive cells were designated
USP22-positive, and sections with <10% USP22-positive cells were
designated USP22-negative.
Cell culture
The human colon cancer cell lines SW480, SW620,
HCT116, HT29 (colorectal adenocarcinoma cell line), and colon
epithelium cell line NCM460 were obtained from American Type
Culture Collection and cultured in DMEM with 10% fetal bovine serum
(Biological Industries, Inc.), and cells were maintained at 37°C
with 5% CO2 in a humidified incubator.
Small interfering (si)RNA
transfection
USP22-siRNA1 (USP22-homo-695), USP22-siRNA2
(USP22-homo-1088) and negative control siRNA were obtained from
Shanghai GenePharma Co., Ltd. siRNA (2.5 µg) transfection was
performed using Lipofectamine® 2000 (Qiagen China Co.,
Ltd.) according to the manufacturer's protocol. Western blotting,
reverse transcription-quantitative (RT-q)PCR and cell functional
assays were performed 48 h after transfection. The sequences of the
siRNAs were: siRNA1, 5′-GGAGAAAGAUCACCUCGAATT-3′; siRNA2,
5′-GCAUCAUAGACCAGAUCUUTT-3′; and negative control siRNA,
5′-UUCUCCGAAGGUGUCACGUTT-3′.
Western blotting
Western blot analysis was performed in order to
examine the protein levels of USP22 essentially as described in
previous studies (24). Protein was
extracted from cells using lysis buffer (Beyotime, Inc.) and the
protein concentration was determined using the BCA Protein assay
kit (Thermo Fisher Scientific, Inc.). Proteins (30 µg) were
separated via SDS-PAGE (10% gel) and transferred to polyvinylidene
fluoride membranes (EMD Millipore). Membranes were blocked at room
temperature with 5% skimmed milk for 45 min and incubated at room
temperature for 2 h with the following antibodies: USP22 rabbit
polyclonal antibody (LifeSpan BioSciences, Inc.; cat. no.
LS-C99567; 1:800), Bmi-1 rabbit polyclonal antibody (Abcam; cat.
no. ab126783; 1:500) and Cyclin D2 rabbit polyclonal antibody
(OriGene Technologies, Inc.; cat. no. TA323121; 1:500), and
subsequently with horseradish peroxidase-conjugated goat
anti-rabbit/mouse secondary antibodies (Abcam; cat. no. ab150077;
1:1,000) at room temperature for 1 h. The blots were incubated by
WesternBright chemiluminescence (ECL) (Western blotting detection
kit; Advansta, Inc.) for 1 min, and developed using ImageQuant™
LAS4000 (GE Healthcare Life Sciences). Relative expression level
was normalized to endogenous control β-actin mouse monoclonal
antibody (Sigma-Aldrich; Merck KGaA; cat. no. A5316; 1:5,000) using
Image J software (version 1.51; National Institutes of Health).
RT-qPCR
RT-quantitative PCR (RT-qPCR) assay was performed in
order to investigate the mRNA levels of USP22 in HCT116 and HT29
cell lines following siRNA transfection as previously described
(24). Total RNA was isolated from
cells and cells using TRIzol® reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) and cDNA was synthesized at 42°C
for 30 min using the Transcript All-in-One First-Strand cDNA
Synthesis Super Mix for qPCR kit (TransGen Biotech, Inc.). The gene
expression levels of USP22 were determined using a SYBR Green qPCR
kit (TransGen Biotech, Inc.), and calculated using the
2−ΔΔCq method (25). The
PCR thermocycling conditions were as follows: 95°C for 40 sec
followed by 40 cycles of 95°C for 5 sec, 60°C for 30 sec, and 95°C
for 15 sec, 60°C for 57 sec, 95°C for 15 sec. GAPDH was used as the
housekeeping gene. The sequences of the primers used were: USP22
forward, 5′-CACTTCTGCGGGACT-3′ and USP22-reverse,
5′-TACGGGATGTGAGGG-3′; and GAPDH-forward
5′-AGCAAGAGCACAAGAGGAAG-3′, and GAPDH-reverse
5′-GGTTGAGCACAGGGTACTTT-3′.
Cell proliferation assays
Cell proliferation was measured using an MTT assay
and colony formation assay as previously described (24). For the MTT assay, cells (1,500 per
well) were seeded into 96-well plates, and MTT values were detected
every day for 5 days. A total of 10 µl of MTT reagent was added
into each well. After 2 h, the medium was removed and 100 µl of
DMSO detection reagent was added. Light absorption values were
detected at 570 nm. For the cell colony formation assay, cells
(1,000 per well) were seeded into 6-well plates, when the number of
cells in the majority of single colonies were >50, cells were
washed three times with PBS and fixed with 90% ethanol for 1 h at
room temperature. Subsequently, cells were stained with 0.1%
crystal violet for 10 min at room temperature, and the number of
visible colonies was counted using the naked eye.
Migration and invasion assays
For the cell migration assay, cells
(1×106 per well) were mixed with medium without serum,
and plated in upper chambers (Corning Inc.) without Matrigel (BD
Biosciences); below the chambers, medium with 10% serum as the
chemoattractant was added. After 40 h, chambers were washed three
times with PBS and fixed with 4% paraformaldehyde for 1 h at room
temperature, and subsequently stained with 0.1% crystal violet for
10 min at room temperature. Finally, the stained cells from four
random fields were counted, and the images were captured using a
light microscope (magnification, ×100; Olympus Corporation). For
the cell invasion assay, cells were plated into 24-well transwell
chambers (Corning Inc.) coated with Matrigel (BD Biosciences), the
number of invaded cells were counted after 70 h. All other
conditions were the same as the migration assay.
Cell cycle assay
For the cell cycle assay, a Cell Cycle and Apoptosis
Analysis kit (Beyotime Institute of Biotechnology) was used. Cells
were collected, washed with PBS and fixed with 70% ice-cold ethanol
>2 h at 4°C. The cells were then washed with 1 ml ice-cold PBS.
A total of 0.5 ml PI solution was added into the cells and
incubated further for 1 h at 4°C in the dark. Flow cytometry (BD
Biosciences FACSVerse) was used to detect the cell cycle stage.
Statistical analysis
Each experiment was repeated three times. An
unpaired two-tailed t-tests was used to analyze the mean values
between two groups, and a one-way ANOVA was used to compare the
mean values among multiple groups with a Bonferroni's multiple
comparison post-hoc test. A Pearson's χ2 test was used
for analysis of data from tissue immunohistochemistry and
clinicopathological parameter association analysis. Patient
relapse-free survival (RFS) and overall survival (OS) analysis was
performed using Kaplan-Meier curves, and the log-rank test was used
for statistical analysis. P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression of USP22 is upregulated in
human colon cancer tissues
A total of 80 colon cancer tissues and 64 normal
colon tissues were collected, and the expression of USP22 in these
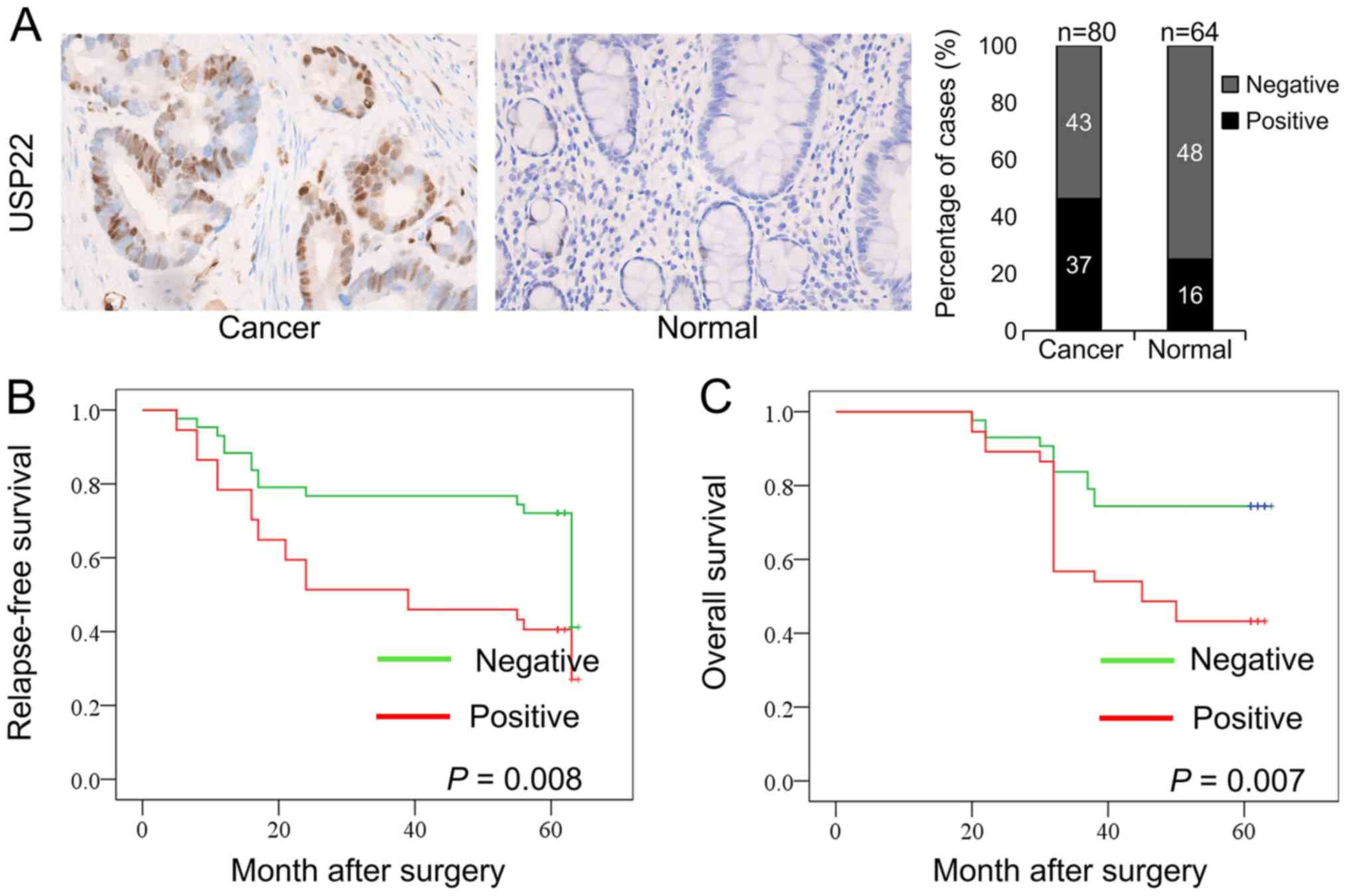
tissues was determined using IHC. Positive signals for USP22
protein in the cells were primarily located in the nucleus
(Fig. 1A). The expression of USP22
was significantly higher in colon cancer tissues compared with
normal tissues. The percentages of USP22-positive and
USP22-negative cells in these tissues were calculated. Among the 80
colon cancer tissues, 37 (46.25%) were USP22-positive, and 43
(53.75%) were USP22-negative. Among the 64 normal colon tissues, 16
(25%) were USP22-positive, and 48 (75%) were USP22-negative. There
was a significant difference in USP22 expression between colon
cancer tissues and normal colon tissues (P=0.009; Table I). Therefore, the expression of USP22
was significantly higher in human colon cancer tissues compared
with the normal colon tissues.
 | Table I.Expression of USP22 in colon cancer
and normal tissues. |
Table I.
Expression of USP22 in colon cancer
and normal tissues.
|
|
| USP22
expression |
|---|
|
|
|
|
|---|
| Group | n | Negative, n
(%) | Positive, n
(%) |
|---|
| Colon cancer | 80 | 43 (53.75) | 37
(46.25)a |
| Normal | 64 | 48 (75.0) | 16 (25.0) |
Association between USP22 expression
and clinicopathological parameters and survival rates in patients
with colon cancer
The clinicopathological parameters of the 80 colon
cancer patients were collected to analyze the association between
USP22 expression and clinicopathological parameters. Patient age,
tumor size, lymph node metastasis, tumor grade and the
Tumor-Node-Metastasis staging system were included in this analysis
(23). Table II shows that the percentage of
tissues with high USP22 expression in colon cancer patients with
lymph node metastasis (20 out of 31; 64.5%) was significantly
higher compared with patients with colon cancer without lymph node
metastasis (17 out of 49; 34.7%; P=0.009). The percentage of
tissues with high USP22 expression in late-stage colon cancer
(stage III–IV; 19 out of 30, 63.3%) was significantly higher
compared with patients with early-stage colon cancer (stage I–II;
18 out of 50, 36.0%; P=0.018). However, there were no significant
associations between USP22 expression and patient age, tumor size
or tumor grade (all P>0.05).
| Table II.Association between USP22 expression
and clinicopathological parameters in patients with colon
cancer. |
Table II.
Association between USP22 expression
and clinicopathological parameters in patients with colon
cancer.
| Parameter | n | USP22 positive
expression, n (%) | P-value |
|---|
| Age, years |
|
| 0.698 |
|
≤55 | 20 | 10 (50.0) |
|
|
>55 | 60 | 27 (45.5) |
|
| Tumor size, cm |
|
| 0.779 |
| ≤5 | 66 | 31 (47.0) |
|
|
>5 | 14 | 6 (42.9) |
|
| Lymph node
metastasis |
|
| 0.009b |
|
Absent | 49 | 17 (34.7) |
|
|
Present | 31 | 20 (64.5) |
|
| Grade |
|
| 0.210 |
|
Well | 10 | 3 (30.0) |
|
|
Moderate | 58 | 26 (44.8) |
|
|
Poor | 12 | 8 (66.7) |
|
| Stage |
|
| 0.018a |
| I +
II | 50 | 18 (36.0) |
|
| III+
IV | 30 | 19 (63.3) |
|
Kaplan-Meier curves were generated to analyze the
association between USP22 expression and RFS and OS in the 80 colon
cancer samples. All patients were followed-up for ≥5 years.
Compared with patients in the USP22-negative group, patients in the
USP22-positive group exhibited significantly lower RFS (P=0.008)
and OS rates (P=0.007; Fig. 1B and
C). Therefore, high expression of USP22 was associated with a
poor prognosis.
Downregulation of USP22
expression
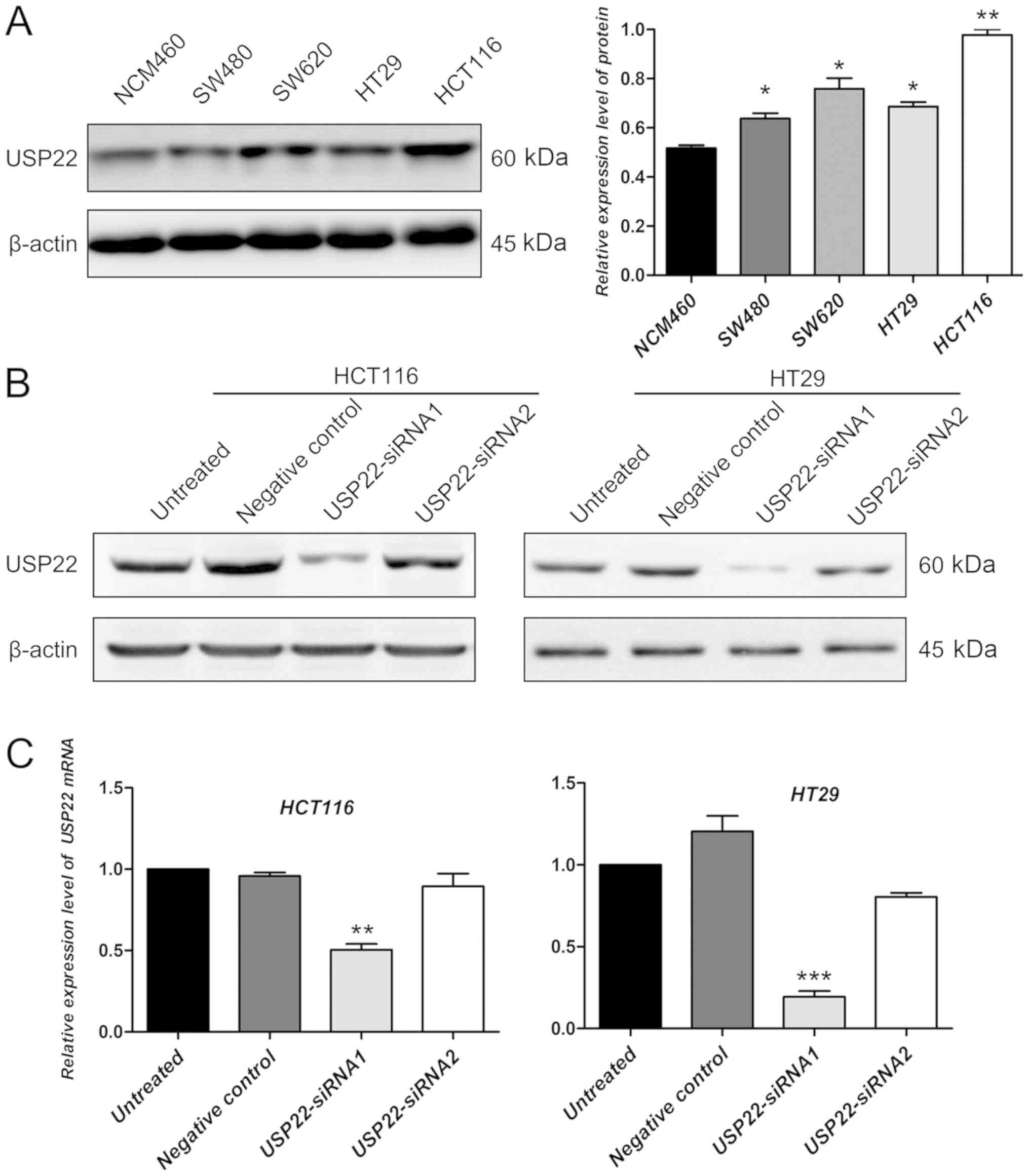
The protein expression levels of USP22 in colon
cancer cell lines (included HT29, colorectal adenocarcinoma cell
line) and a colon epithelium cell line were determined by western
blotting. Compared with the colon epithelium cell line, the
expression of USP22 was significantly higher in colon cancer cell
lines (P<0.05; Fig. 2A). The
human colon cancer cell line HCT116 and colorectal adenocarcinoma
cell line HT29 were selected to perform the cell functional
experiments using siRNA transfection for USP22 knockdown. Compared
with untreated cells and the negative control, in both cell lines,
USP22-siRNA1 decreased the protein expression level of USP22
(Fig. 2B). However, the decrease in
USP22 protein levels in cells transfected with siRNA2 was not
significant compared with that in control cells (Fig. 2B). To confirm this result, the mRNA
expression levels of USP22 were also examined in untreated,
negative control, and USP22-siRNA1 and USP22-siRNA2 transfected
cancer cells. Compared with the control, the mRNA expression levels
of USP22 decreased significantly in cells transfected with
USP22-siRNA1 in HCT116 and HT29 cells (P<0.01; Fig. 2C). Therefore, cells transfected with
USP22- siRNA1 were used for all subsequent experiments.
Knockdown of USP22 reduces
proliferation in human colon cancer cells
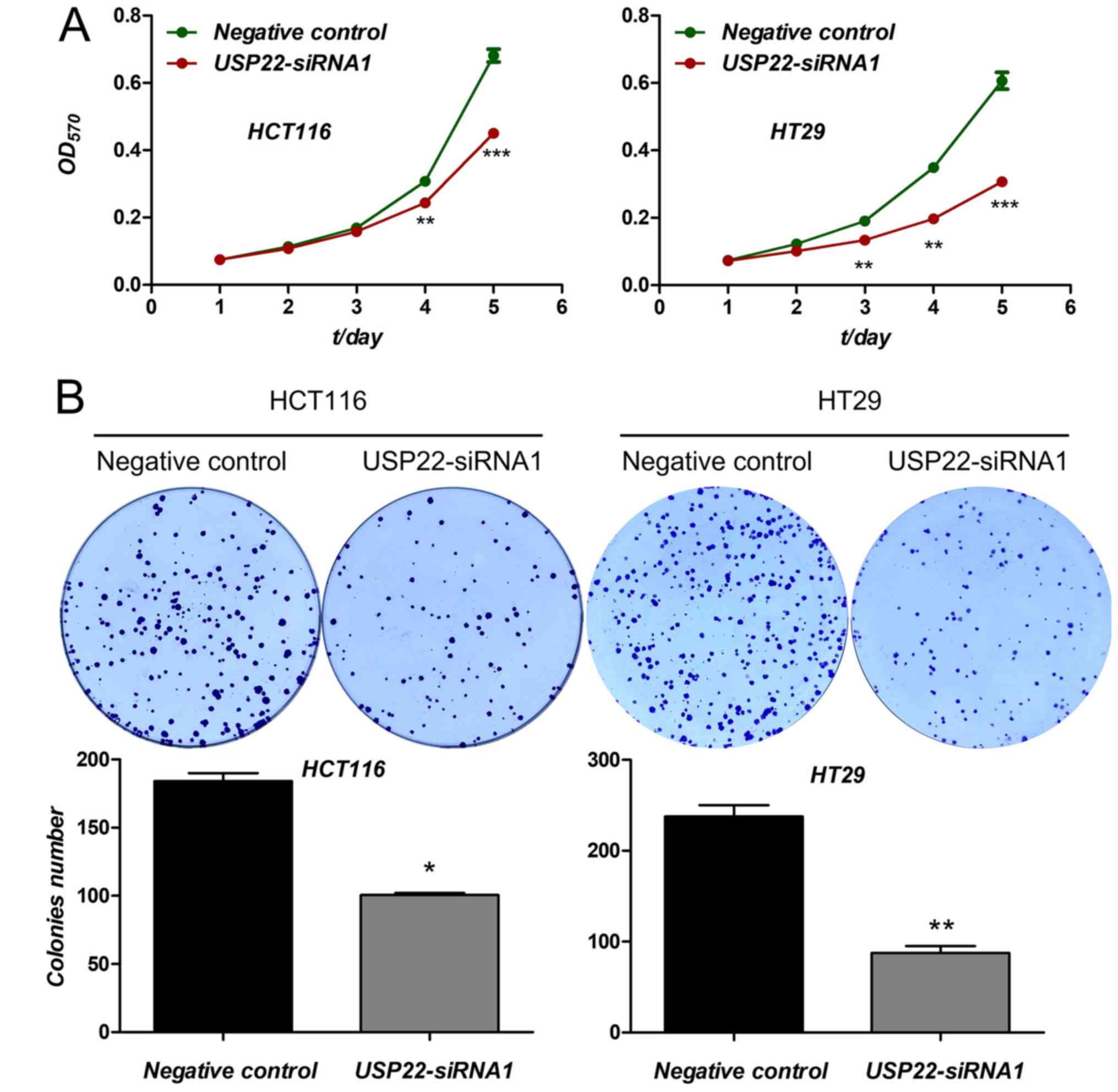
The effect of USP22 knockdown on proliferation as
determined. Compared with the negative control, cell viability,
evaluated using an MTT assay, was significantly decreased in cells
transfected with USP22-siRNA1 in both cancer cell lines after 3
(HCT116 cells) and 4 days (HT29 cells) (P<0.01; Fig. 3A). Furthermore, compared with the
negative control, cell colony formation was also reduced in both
cancer cell lines transfected with USP22-siRNA1 (P<0.05;
Fig. 3B). Therefore, knockdown of
USP22 significantly reduced cell proliferation in human colon
cancer cells. These results suggest that USP22 acts as a promoter
of tumor cell proliferation in human colon cancer cells.
Knockdown of USP22 reduces metastasis
in human colon cancer cells
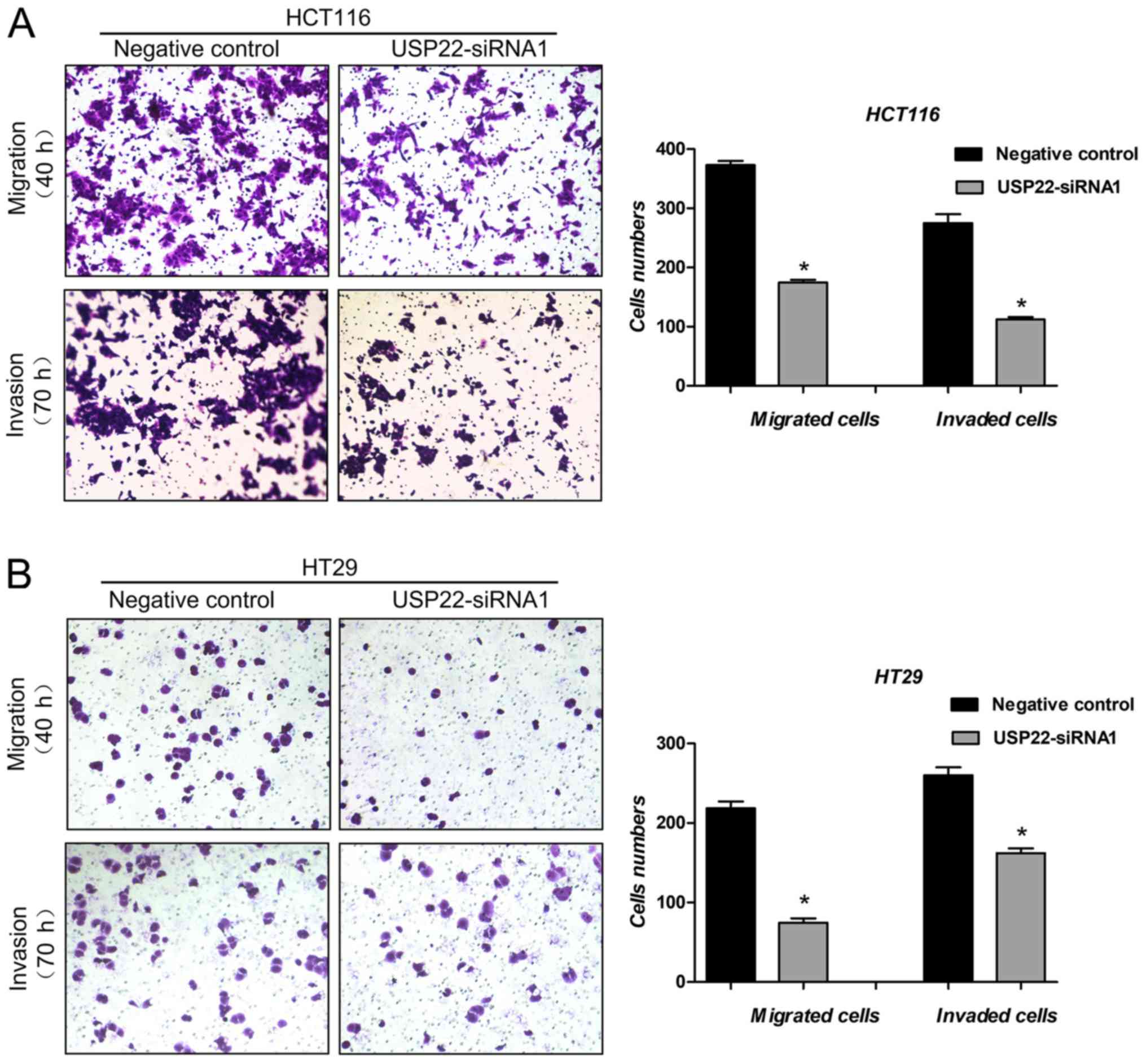
Cell migration and invasion assays were used to
examine the effect of USP22 on metastasis of human colon cancer
cells. Compared with the negative control, in cells transfected
with USP22-siRNA1, both cell migration (P<0.05) and cell
invasion (P<0.05) were significantly decreased in both colon
cancer cell lines (Fig. 4A and B).
These results suggest that USP22 promotes metastasis in human colon
cancer cells.
USP22 regulates the expression of
Bmi-1 and Cyclin D2
Several candidate genes were selected to identify
the downstream mechanism by which USP22 exerted its effects on
human colon cancer cells (data not shown). Similar to previous
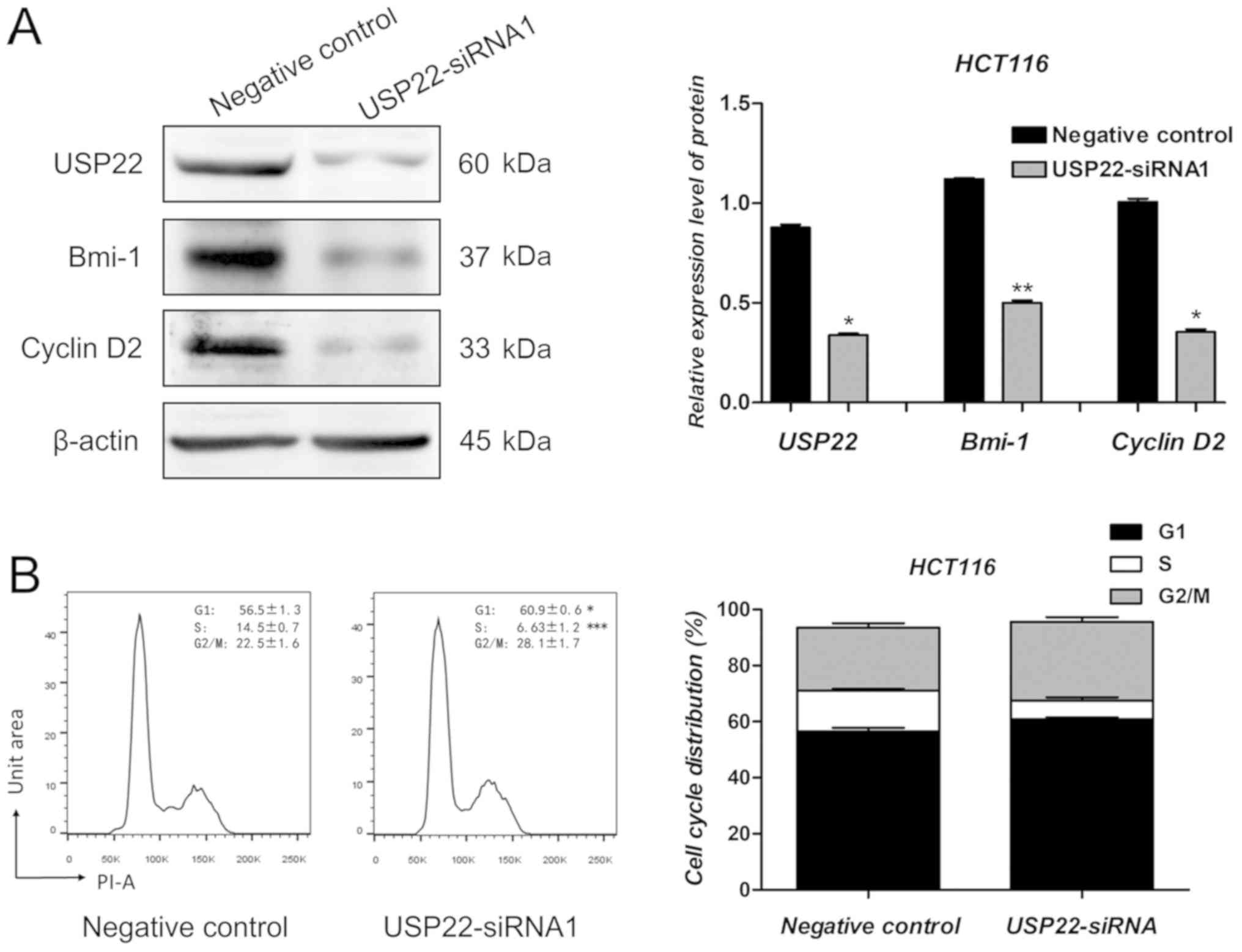
results, the protein expression levels of USP22 decreased
significantly following transfection with USP22-siRNA1 in HCT116
cells. Compared with the negative control cells, Bmi-1 and Cyclin
D2 protein expression levels were decreased significantly in cells
transfected with USP22-siRNA1 (P<0.05; Fig. 5A). As reported previously, Bmi-1 and
Cyclin D2 are oncogenes in human colon cancer cells (26–28). To
determine whether USP22 knockdown affected cell cycle regulation,
flow cytometry analysis was performed following transfection with
USP22-siRNA1 and negative control siRNA. As shown in Fig. 5B, the number of USP22-siRNA1 cells in
the G1 phase increased significantly compared with the negative
control (P<0.05; Fig. 5B).
Therefore, Bmi-1 and Cyclin D2 may be involved in the promotion of
proliferation and metastasis of colon cancer cells by USP22.
Discussion
In the present study, the role of USP22 in human
colon cancer was determined through clinical analyses and cell
functional assays. IHC analysis and clinicopathological parameter
association analysis showed that the expression of USP22 was higher
in colon cancer tissues compared with normal colon tissues, and
that overexpression of USP22 was positively correlated with tumor
lymph node metastasis and tumor stage. Kaplan-Meier curves showed
that USP22 was negatively associated with patient RFS and OS rates
in the 80 patients with colon cancer. Cell functional assays were
performed. Knockdown of USP22 by siRNA transfection significantly
decreased cell viability, cell colony formation, cell migration and
cell invasion. Therefore, USP22 promoted both cell proliferation
and cell metastasis in colon cancer cells and upregulation of USP22
predicted poor prognosis for patients with colon cancer.
USP22 belongs to the deubiquitinase family of
proteins and strictly regulates gene expression through histone
deubiquitination or acetylation procession (12,13,15,16). It
has been reported that USP22 serves an important role in driving
transcription and cell cycle progression (17–19).
USP22 has been demonstrated to be a vital biomarker of cancer stem
cells (16,29). In the present study, the oncogenic
role of USP22 in human colon cancer was determined. As reported
previously, Wang et al (30)
showed that USP22 was upregulated in malignant colon carcinoma, and
expression was associated with the degree of differentiation,
invasion, lymph node metastasis and tumor stage in patients with
colon carcinoma (30). The results
of the present study are consistent with the results observed by
Wang et al (30). However, Ao
et al (31) reported that
USP22 promoted cell proliferation but inhibited cell invasion in
SW480 colon cancer through the STAT3/MMP9 pathway (31). In these experiments, USP22 increased
the proliferation in HCT116 and SW480 colon cancer cell lines,
whereas the metastasis promoting effects on colon cancer cells were
cell specific. Furthermore, it has been reported that USP22
positively regulates c-Myc and promotes tumorigenic activity in
human breast cancer (22). Tang
et al (32) reported that
elevated expression of USP22 was associated with poor prognosis in
breast cancer patients. Ma et al (19) demonstrated that USP22 maintained
gastric cancer cell stemness and promoted gastric cancer
progression by stabilizing the Bmi-1 protein (19). USP22 was also reported to serve as an
oncogene in human hepatocellular carcinoma; overexpression of USP22
indicated poor prognosis for patients with hepatocellular carcinoma
and USP22 mediated multidrug resistance in hepatocellular carcinoma
(20,32). USP22 also served as an oncogene in a
number of different types of cancer, including non-small-cell lung
cancer (21), papillary thyroid
carcinoma (29) and glioma (33). Therefore, USP22 has been shown to be
oncogenic in a large number of different types of cancer including
colon cancer, and specific inhibitors of USP22 may serve as
potential therapeutic options for treating patients with cancer
where upregulation of USP22 is observed.
It was determined that Bmi-1 and Cyclin D2 were
positively regulated by USP22. As reported previously, Bmi-1 is
oncogenic in human colon cancer cells; the expression level of
Bmi-1 is associated with tumor progression and prognosis of colon
cancer, and Bmi-1 promotes migration and invasion of colon cancer
stem cells by regulating E-cadherin (26,27).
Furthermore, Bmi-1 has also been reported to serve as an oncogene
in human hepatocellular carcinoma (34), oral cancer (35), breast cancer (36), gastric cancer (37) and lung cancer (38) amongst others. Cyclin D2 is an
important protein involved in cell cycle regulation. Cyclin D2
promotes both proliferation and metastasis of human colon cancer
cells (28,39). Furthermore, Cyclin D2 also serves as
an oncogene in a number of different types of cancer, including
breast cancer (40), prostate cancer
(41), oral squamous carcinoma
(42) and non-small-cell lung cancer
(43). Therefore, Bmi-1 and Cyclin
D2 are important oncogenes in cancer and may mediate the promoting
role of USP22 in human colon cancer.
In conclusion, the present study systematically
examined the oncogenic role of USP22 in human colon cancer.
Overexpression of USP22 is associated with enhanced malignant
properties in colon cancer cells and a poorer prognosis in patients
with colon cancer. The present study highlights the role of USP22
in cancer progression, and USP22 may serve as a potential
therapeutic target for treating patients with colon cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Scientific
Research Foundation of Anhui Medical University, National Natural
Science Foundation of China (grant nos. 81572350 and 81500373),
Natural Science Foundation of Anhui Province (grant no.
1608085MH193) and the Excellent Talents Supporting Program of the
University of Anhui Education Department (grant no.
gxyq2018011).
Availability of data and material
The datasets used and/or analyzed during the present
study are available from the author upon reasonable request.
Authors' contributions
WW designed the study. XY collected the data and HW
performed the experiments. AX, XZ and YZ performed the statistical
analyses. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Each patient signed informed consent for the present
study. The present study was approved by Biomedical Ethics
Committee of Anhui Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Meng X and Fu R: miR-206 regulates 5-FU
resistance by targeting Bcl-2 in colon cancer cells. Onco Targets
Ther. 11:1757–1765. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Evert J, Pathak S, Sun XF and Zhang H: A
study on effect of oxaliplatin in MicroRNA expression in human
colon cancer. J Cancer. 9:2046–2053. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li Y, Gong P, Hou JX, Huang W, Ma XP, Wang
YL, Li J, Cui XB and Li N: miR-34a regulates multidrug resistance
via positively modulating OAZ2 signaling in colon cancer cells. J
Immunol Res. 2018:74985142018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dou H, Shen R, Tao J, Huang L, Shi H, Chen
H, Wang Y and Wang T: Curcumin Suppresses the Colon Cancer
Proliferation by Inhibiting Wnt/β-catenin pathways via miR-130a.
Front Pharmacol. 8:8772017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li L and Ma BB: Colorectal cancer in
Chinese patients: Current and emerging treatment options. Onco
Targets Ther. 7:1817–1828. 2014.PubMed/NCBI
|
|
6
|
Mishra J, Drummond J, Quazi SH, Karanki
SS, Shaw JJ, Chen B and Kumar N: Prospective of colon cancer
treatments and scope for combinatorial approach to enhanced cancer
cell apoptosis. Crit Rev Oncol Hematol. 86:232–250. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu YR, Liang L, Zhao JM, Zhang Y, Zhang
M, Zhong WL, Zhang Q, Wei JJ, Li M, Yuan J, et al: Twist1 confers
multidrug resistance in colon cancer through upregulation of
ATP-binding cassette transporters. Oncotarget. 8:52901–52912.
2017.PubMed/NCBI
|
|
8
|
He K, Chen D, Ruan H, Li X, Tong J, Xu X,
Zhang L and Yu J: BRAFV600E-dependent Mcl-1 stabilization leads to
everolimus resistance in colon cancer cells. Oncotarget.
7:47699–47710. 2016.PubMed/NCBI
|
|
9
|
Segditsas S and Tomlinson I: Colorectal
cancer and genetic alterations in the Wnt pathway. Oncogene.
25:7531–7537. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hollis M, Nair K, Vyas A, Chaturvedi LS,
Gambhir S and Vyas D: MicroRNAs potential utility in colon cancer:
Early detection, prognosis, and chemosensitivity. World J
Gastroenterol. 21:8284–8292. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yu YJ, Nangia-Makker P, Farhana L and
Majumdar APN: A novel mechanism of lncRNA and miRNA interaction:
CCAT2 regulates miR-145 expression by suppressing its maturation
process in colon cancer cells. Mol Cancer. 16:1552017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gong Z, Liu J, Xie X, Xu X, Wu P, Li H,
Wang Y, Li W and Xiong J: Identification of potential target genes
of USP22 via ChIP-seq and RNA-seq analysis in HeLa cells. Genet Mol
Biol. 41:488–495. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang A, Ning Z, Lu C, Gao W, Liang J, Yan
Q, Tan G and Liu J: USP22 induces cisplatin resistance in lung
adenocarcinoma by regulating γH2AX-Mediated DNA damage repair and
Ku70/Bax-mediated apoptosis. Front Pharmacol. 8:2742017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee HJ, Kim MS, Shin JM, Park TJ, Chung HM
and Baek KH: The expression patterns of deubiquitinating enzymes,
USP22 and Usp22. Gene Expr Patterns. 6:277–284. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Armour SM, Bennett EJ, Braun CR, Zhang XY,
McMahon SB, Gygi SP, Harper JW and Sinclair DA: A high-confidence
interaction map identifies SIRT1 as a mediator of acetylation of
USP22 and the SAGA coactivator complex. Mol Cell Biol.
33:1487–1502. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang XY, Varthi M, Sykes SM, Phillips C,
Warzecha C, Zhu W, Wyce A, Thorne AW, Berger SL and McMahon SB: The
putative cancer stem cell marker USP22 is a subunit of the human
SAGA complex required for activated transcription and cell-cycle
progression. Mol Cell. 29:102–111. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang X, Zang H, Luo Y, Wu J, Fang Z, Zhu W
and Li Y: High expression of USP22 predicts poor prognosis and
advanced clinicopathological features in solid tumors: A
meta-analysis. Onco Targets Ther. 11:3035–3046. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hoeller D and Dikic I: Targeting the
ubiquitin system in cancer therapy. Nature. 458:438–444. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ma Y, Fu HL, Wang Z, Huang H, Ni J, Song
J, Xia Y, Jin WL and Cui DX: USP22 maintains gastric cancer stem
cell stemness and promotes gastric cancer progression by
stabilizing BMI1 protein. Oncotarget. 8:33329–33342.
2017.PubMed/NCBI
|
|
20
|
Ling S, Li J, Shan Q, Dai H, Lu D, Wen X,
Song P, Xie H, Zhou L, Liu J, et al: USP22 mediates the multidrug
resistance of hepatocellular carcinoma via the SIRT1/AKT/MRP1
signaling pathway. Mol Oncol. 11:682–695. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu G, Cai J, Wang L, Jiang LY, Huang JB,
Hu R and Ding F: MicroRNA-30e-5p suppresses non-small cell lung
cancer tumorigenesis by regulating USP22-mediated Sirt1/JAK/STAT3
signaling. Exp Cell Res. 362:268–278. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kim D, Hong A, Park HI, Shin WH, Yoo L,
Jeon SJ and Chung KC: Deubiquitinating enzyme USP22 positively
regulates c-Myc stability and tumorigenic activity in mammalian and
breast cancer cells. J Cell Physiol. 232:3664–3676. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bosman FT, Carneiro F, Hruban RH and
Theise ND: WHO classification of tumours of the digestive
system4th. 3. International Agency for Research on Cancer; Lyon:
2010
|
|
24
|
Wang B, Wu ZS and Wu Q: CMIP promotes
proliferation and metastasis in human glioma. Biomed Res Int.
2017:53401602017.PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li DW, Tang HM, Fan JW, Yan DW, Zhou CZ,
Li SX, Wang XL and Peng ZH: Expression level of Bmi-1 oncoprotein
is associated with progression and prognosis in colon cancer. J
Cancer Res Clin Oncol. 136:997–1006. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang Z, Bu X, Chen H, Wang Q and Sha W:
Bmi-1 promotes the invasion and migration of colon cancer stem
cells through the downregulation of E-cadherin. Int J Mol Med.
38:1199–1207. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Schlormann W, Naumann S, Renner C and Glei
M: Influence of miRNA-106b and miRNA-135a on butyrate-regulated
expression of p21 and Cyclin D2 in human colon adenoma cells. Genes
Nutr. 10:502015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li ZH, Yu Y, Du C, Fu H, Wang J and Tian
Y: RNA interference-mediated USP22 gene silencing promotes human
brain glioma apoptosis and induces cell cycle arrest. Oncol Lett.
5:1290–1294. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang Z, Zhu L, Guo T, Wang Y and Yang J:
Decreased H2B monoubiquitination and overexpression of
ubiquitin-specific protease enzyme 22 in malignant colon carcinoma.
Hum Pathol. 46:1006–1014. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ao N and Liu Y, Bian X, Feng H and Liu Y:
Ubiquitin-specific peptidase 22 inhibits colon cancer cell invasion
by suppressing the signal transducer and activator of transcription
3/matrix metalloproteinase 9 pathway. Mol Med Rep. 12:2107–2113.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tang B, Tang F, Li B, Yuan SG, Xu Q,
Tomlinson S, Jin JF, Hu W and He SQ: High USP22 expression
indicates poor prognosis in hepatocellular carcinoma. Oncotarget.
6:12654–12667. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhao H, Tang H, Huang Q, Qiu B, Liu X, Fan
D, Gong L, Guo H, Chen C, Lei S, et al: MiR-101 targets USP22 to
inhibit the tumorigenesis of papillary thyroid carcinoma. Am J
Cancer Res. 6:2575–2586. 2016.PubMed/NCBI
|
|
34
|
Shao Y, Zhang D, Li X, Yang J, Chen L,
Ning Z, Xu Y, Deng G, Tao M, Zhu Y and Jiang J: MicroRNA-203
increases cell radiosensitivity via directly targeting Bmi-1 in
hepatocellular carcinoma. Mol Pharm. 15:3205–3215. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kim JS, Choi DW, Kim CS, Yu SK, Kim HJ, Go
DS, Lee SA, Moon SM, Kim SG, Chun HS, et al: MicroRNA-203 induces
apoptosis by targeting Bmi-1 in YD-38 oral cancer cells. Anticancer
Res. 38:3477–3485. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yan Y, Wang Y, Zhao P, Ma W, Hu Z and
Zhang K: BMI-1 promotes self-renewal of radio- and temozolomide
(TMZ)-resistant breast cancer cells. Reprod Sci. 24:1620–1629.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li N, Jiang K, Fang LP, Yao LL and Yu Z:
Knockdown of long noncoding RNA CCAT1 inhibits cell growth,
invasion and peritoneal metastasis via downregulation of Bmi-1 in
gastric cancer. Neoplasma. 65:736–744. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Koh H, Park H, Chandimali N, Huynh DL,
Zhang JJ, Ghosh M, Gera M, Kim N, Bak Y, Yoon DY, et al:
MicroRNA-128 suppresses paclitaxel-resistant lung cancer by
inhibiting MUC1-C and BMI-1 in cancer stem cells. Oncotarget.
8:110540–110551. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang Y, Xue J, Kuang H, Zhou X, Liao L and
Yin F: microRNA-1297 inhibits the growth and metastasis of
colorectal cancer by suppressing Cyclin D2 expression. DNA Cell
Biol. 36:991–999. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhong H, Yang J, Zhang B, Wang X, Pei L,
Zhang L, Lin Z, Wang Y and Wang C: LncRNA GACAT3 predicts poor
prognosis and promotes cell proliferation in breast cancer through
regulation of miR-497/CCND2. Cancer Biomark. 22:787–797. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Huang F, Zhao H, Du Z and Jiang H: MiR-615
inhibits prostate cancer cell proliferation and invasion by
directly targeting Cyclin D2. Oncol Res. 27:293–299. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zeng Q, Tao X, Huang F, Wu T, Wang J,
Jiang X, Kuang Z and Cheng B: Overexpression of miR-155 promotes
the proliferation and invasion of oral squamous carcinoma cells by
regulating BCL6/Cyclin D2. Int J Mol Med. 37:1274–1280. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Li YL, Wang J, Zhang CY, Shen YQ, Wang HM,
Ding L, Gu YC, Lou JT, Zhao XT, Ma ZL and Jin YX: MiR-146a-5p
inhibits cell proliferation and cell cycle progression in NSCLC
cell lines by targeting CCND1 and CCND2. Oncotarget. 7:59287–59298.
2016.PubMed/NCBI
|