Introduction
Lung cancer is the leading cause of mortality among
patients with cancer worldwide and non-small cell lung cancer
(NSCLC) is the major type of lung cancer, accounting for nearly 85%
of lung cancer cases (1). Most
patients with NSCLC are diagnosed in the advanced phase and the
5-year survival rate is <5% despite advances in treatment
(1). Chemotherapy plays a critical
role in lung cancer treatment (2).
Cisplatin (DDP) is one of the first-line chemotherapeutic agents
used to treat NSCLC. However, the majority of patients suffer from
DDP resistance, leading to treatment failure, limiting its clinical
application (3). In addition,
DDP-based regimens are frequently associated with dose-associated
toxicity, which results in decreases in drug tolerability and
therapeutic efficacy (4). Thus,
investigating the underlying molecular mechanism of DDP-resistance
and developing new therapeutic agents that are more effective and
less toxic in sensitizing NSCLC to chemotherapy is urgently
required.
The epithelial-to-mesenchymal transition (EMT) has
been reported to promote drug resistance in a number of previous
studies (5–7) and thus, ongoing research has focused on
investigating ways to reverse or delay the EMT to prevent drug
resistance (6). Cancerous inhibitor
of protein phosphatase 2A (CIP2A) is a novel human oncoprotein that
is overexpressed in numerous types of human malignancy and is a
predictor for poor prognosis, including NSCLC (8–10). CIP2A
inhibits protein phosphatase 2A (PP2A) tumor suppressor activity
and facilitates tumor growth, progression, metastasis, apoptosis
and drug resistance in numerous different types of cancer (9,10). In
addition, CIP2A facilitates the EMT process via the
mitogen-activated protein kinase (MEK)/extracellular
signal-regulated kinase-1 (ERK) pathway (11). CIP2A also mediates the anticancer
effects of numerous compounds and its potential role in mediating
the resistance of cancer cells to numerous chemotherapeutic agents
has been confirmed (12–15). Notably, CIP2A mediated protein kinase
B (AKT) phosphorylation (14,16) and
the AKT downstream mammalian target of rapamycin (mTOR)/p70S6K1
pathway plays roles in mediating DDP resistance in lung cancer
(17). Therefore, the CIP2A
signaling axis may be an effective target in reversing DDP
resistance and improving the response to DDP-based therapy in
NSCLC.
Rhizoma of Paris polyphylla (RPS), a
traditional Chinese medicinal herb, has been widely prescribed by
herbal practitioners to treat infection, hemorrhage, immune
diseases and tumors (18).
Polyphyllin I (PPI) and polyphyllin VII (PPVII) are two of the
primary active components isolated from RPS, which exhibit strong
anticancer effects in a number of different types of cancer by
regulating the signaling pathways associated with proliferation,
apoptosis and autophagy (19–27).
Notably, PPI and PPVII have also been demonstrated to possess
chemosensitizing effects in a number of different types of cancer
(28–32). However, the effects of PPI and PPVII
on DDP-resistant lung cancer cells remain unclear. In the present
study, the chemosensitizing effects of PPI and PPVII on A549/DDP
cells were investigated in vitro, alongside further
investigation into the potential underlying molecular
mechanisms.
Materials and methods
Cell culture and reagents
Human NSCLC cell lines A549 and DDP-resistant
A549/DDP cells (OriGene Technologies, Inc.) were cultured in
RPMI-1640 (Gibco; Thermo Fisher Scientific, Inc.) supplemented with
10% fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.) and
penicillin (100 U/ml)-streptomycin (100 mg/ml) at 37°C, with 5%
CO2. A549/DDP cells were cultured in RPMI-1640 with
1,000 ng/ml DDP (Sigma Aldrich; Merck KGaA). PPI and PPVII were
purchased from Beijing Solarbio Science & Technology Co., Ltd.
Antibodies used in the present study were: anti-E-cadherin (cat.
no. 3195T), anti-vimentin (cat. no. 12826S) anti-α-smooth muscle
actin (SMA; cat. no. 19245T), anti-CIP2A (cat. no. 14805S),
anti-AKT (cat. no. 4691T), anti-phosphorylated AKT (p-AKT; cat. no.
4060S), anti-p-mTOR (cat. no. 5536T), anti-mTOR (cat. no. 2983T),
anti-poly (ADP-ribose) polymerase 1 (PARP; cat. no. 9532T),
anti-cleaved PARP (cat. no. 9542T), anti-p53 (cat. no. 2527T),
anti-Bcl-2 (cat. no. 15071T), anti-Bax (cat. no. 5023T),
anti-caspase-3 (cat. no. 14220T), anti-cleaved-caspase-3 (cat. no.
9664T) and anti-GAPDH (cat. no. 5174T), which were all purchased
from Cell Signaling Technologies, Inc.
Cell proliferation and viability
assays
Cells that had reached the logarithmic growth phase
were adjusted to a cell density of 5.0×104/ml and 100 µl
was seeded in 96-well plates at a density of 5.0×103
cells/well. Cells were allowed to adhere for 24 h, after which the
cells were treated with the indicated drugs and cultured for a
further 24 h. A total of 100 µl sterile Cell Counting Kit-8 (CCK-8)
solution (Dojindo Molecular Technologies, Inc.) was added to the
culture medium of each well and incubated at 37°C for 2 h, and the
absorbance was measured at 450 nm using a microplate reader. The
effect of each drug on the cell viability was calculated according
to the following formula: Cell survival rate (cell viability)
%=[optical density (OD) of dosing group A/OD of control group A]
×100%.
Annexin V-fluorescein isothiocyanate
(FITC)/propidium iodide (PI) apoptosis analysis
Apoptotic cells were measured using an Annexin
V-FITC Apoptosis Detection kit (BD Biosciences) according to the
manufacturer's protocol. A549/DDP cells were treated with the
indicated drugs for 24 h and then washed with PBS. The cells were
resuspended in 500 µl binding buffer, 5 µl Annexin V-FITC and 5 µl
PI were added and the mixture was incubated in the dark at room
temperature for 15 min. The cells were then analyzed using a BD
FACS Canto™ III flow cytometry device and Kaluza Analysis software
(version 1.3; Beckman Coulter, Inc.). Early apoptotic cells were
Annexin V-FITC (+)/PI (−), while late apoptotic or necrotic cells
were Annexin V-FITC (+)/PI (+).
Western blot analysis
Cells were washed twice with PBS and lysed with RIPA
buffer (Beyotime Insititute of Biotechnology) according to the
manufacturer's protocol. Using BCA protein concentration assay,
lysates were normalized and protein samples were prepared with
SDS-PAGE protein sample buffer (Beyotime Institute of
Biotechnology) at a volume ratio of 4:1. Samples containing equal
quantity of protein (25 µg) were separated via SDS-PAGE (10% gel)
and then electro-transferred to polyvinylidene fluoride membranes
(EMD Millipore) using a constant current of 200 mA for at least 2 h
at 0°C. The membranes were blocked with 5% non-fat milk made with
Tris-buffered saline (containing 0.1% Tween 20) at room temperature
for 1 h, followed by incubation with the primary antibodies
overnight at 4°C: anti-E-cadherin (1:1,000), anti-vimentin
(1:1,000), anti-α-SMA (1:1,000), anti-CIP2A (1:1,000), anti-AKT
(1:1,000), anti-p-AKT (1:2,000), anti-p-mTOR (1:1,000), anti-mTOR
(1:1,000), anti-PARP (1:1,000), anti-cleaved PARP (1:1,000),
anti-p53 (1:1,000), anti-Bcl-2 (1:1,000), anti-Bax (1:1,000),
anti-caspase-3 (1:1,000), anti-cleaved-caspase-3 (1:1,000) and
anti-GAPDH (1:1,000). Then the membranes were washed and incubated
with the corresponding secondary horseradish peroxidase-conjugated
antibodies (1:2,500; cat. nos. ZB-2305 and ZB-2301; OriGene
Technologies, Inc.) at room temperature for 1 h. The bound antibody
complexes were detected using the immobilon western
chemiluminescent HRP substrate(cat. no. WBKLS0100; EMD Millipore)
and analyzed using Quantity One software (version 4.6; Bio-Rad
Laboratories).
Statistical analysis
All experiments were repeated at least three times
and the data were processed using SPSS software (version 19.0; IBM
Corp.). The relative expression levels were presented as the mean ±
standard deviation. Differences between data groups were evaluated
for significance using one-way analysis of variance followed by the
Tukey test. P<0.05 was considered to indicate a statistically
significant result.
Results
Characterization of A549 and A549/DDP
cells
A549 and A549/DDP cells were exposed to different
concentrations of DDP (1–32 µg/ml) for 24 h (Fig. 1A). The 50% inhibitory concentration
(IC50) of DDP against A549 cells was 5.49 µg/ml, while
the IC50 of DDP against A549/DDP cells was 33.36 µg/ml.
The DDP cytotoxicity in A549 cells was higher compared with
A549/DDP cells.
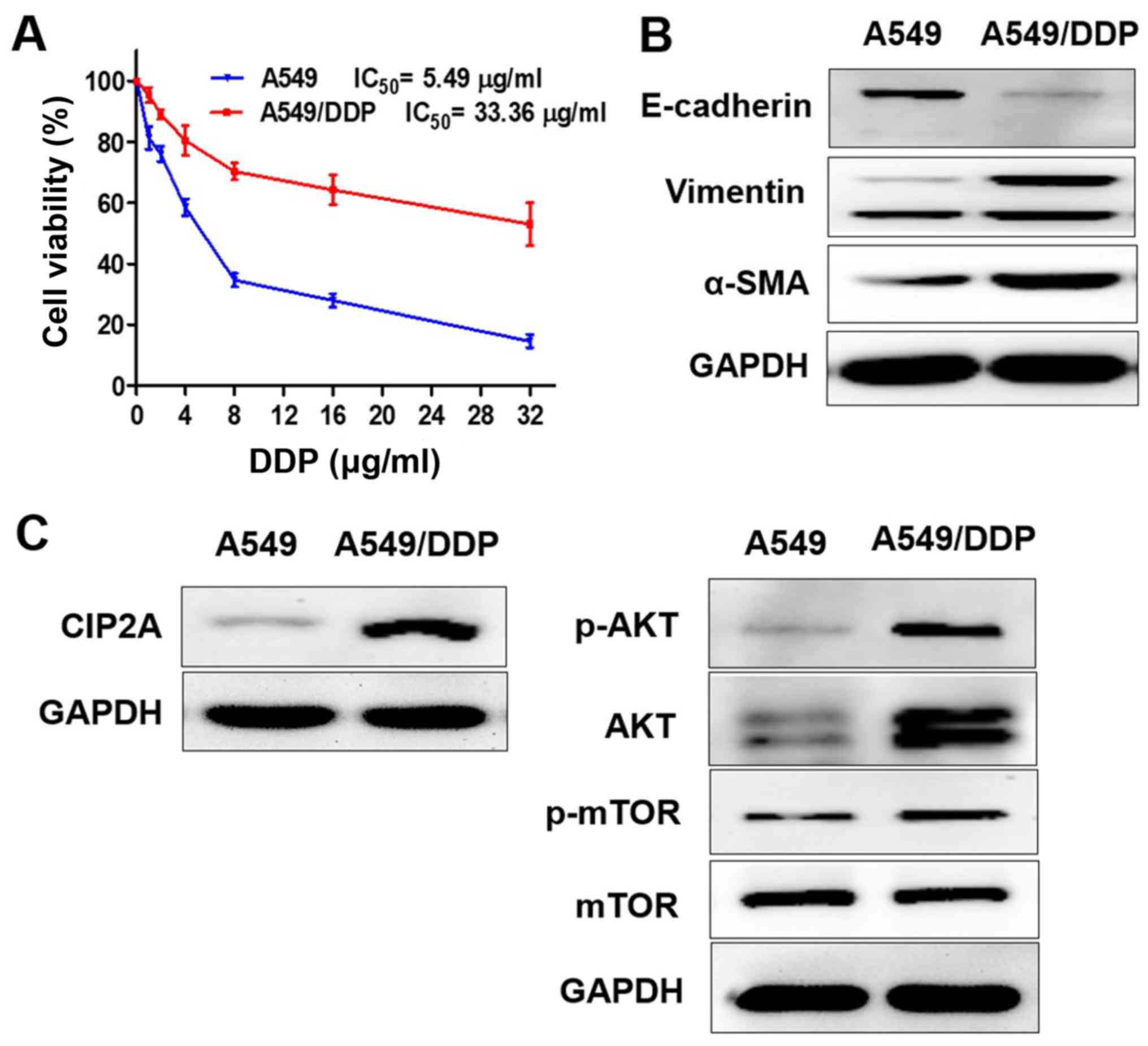 | Figure 1.Characterization of A549 and A549/DDP
cells. (A) A549 and A549/DDP cells were exposed to different
concentrations of DDP (1–32 µg/ml) for 24 h, and cell viability was
determined by Cell Counting Kit-8 assay. The protein expression
levels of (B) E-cadherin, vimentin, α-SMA and (C) CIP2A, p-AKT,
AKT, p-mTOR were measured using western blotting; E, epithelial;
SMA, smooth muscle actin; p-mTOR, phosphorylated mammalian target
of rapamycin; AKT, protein kinase B; DDP, cisplatin; CIP2A,
cancerous inhibitor of protein phosphatase 2A; IC50,
half maximal inhibitory concentration. |
In order to determine whether the EMT and CIP2A
pathways are affected when A549 cells acquire resistance to DDP,
the present study compared the levels of EMT markers and CIP2A,
AKT, p-AKT, p-mTOR in A549 and A549/DDP cells. The results
confirmed that the epithelial cell marker E-cadherin exhibited low
expression, while the mesenchymal transition markers vimentin and
α-SMA were overexpressed in A549/DDP cells. The A549/DDP cell line
exhibited the EMT phenotype (Fig.
1B). CIP2A and AKT, and p-AKT and p-mTOR were overexpressed in
A549/DDP cells, while expression levels were decreased in A549
cells (Fig. 1C).
Inhibitory effects of PPI and PPVII on
A549 and A549/DDP cells
Chemical structure of PPI and PPVII are shown in
Fig. 2A. A549 and A549/DDP cells
were seeded in 96-well plates for 24 h and then treated with
different concentrations of PPI (0.5–8 µg/ml) and PPVII (0.5–8
µg/ml) for 24 h. Cell viability was determined using a CCK-8 assay
and presented as a percentage of the control (Fig. 2B). The results revealed that PPI and
PPVII both caused moderate cytotoxicity in A549 and A549/DDP cells
in a dose-dependent manner.
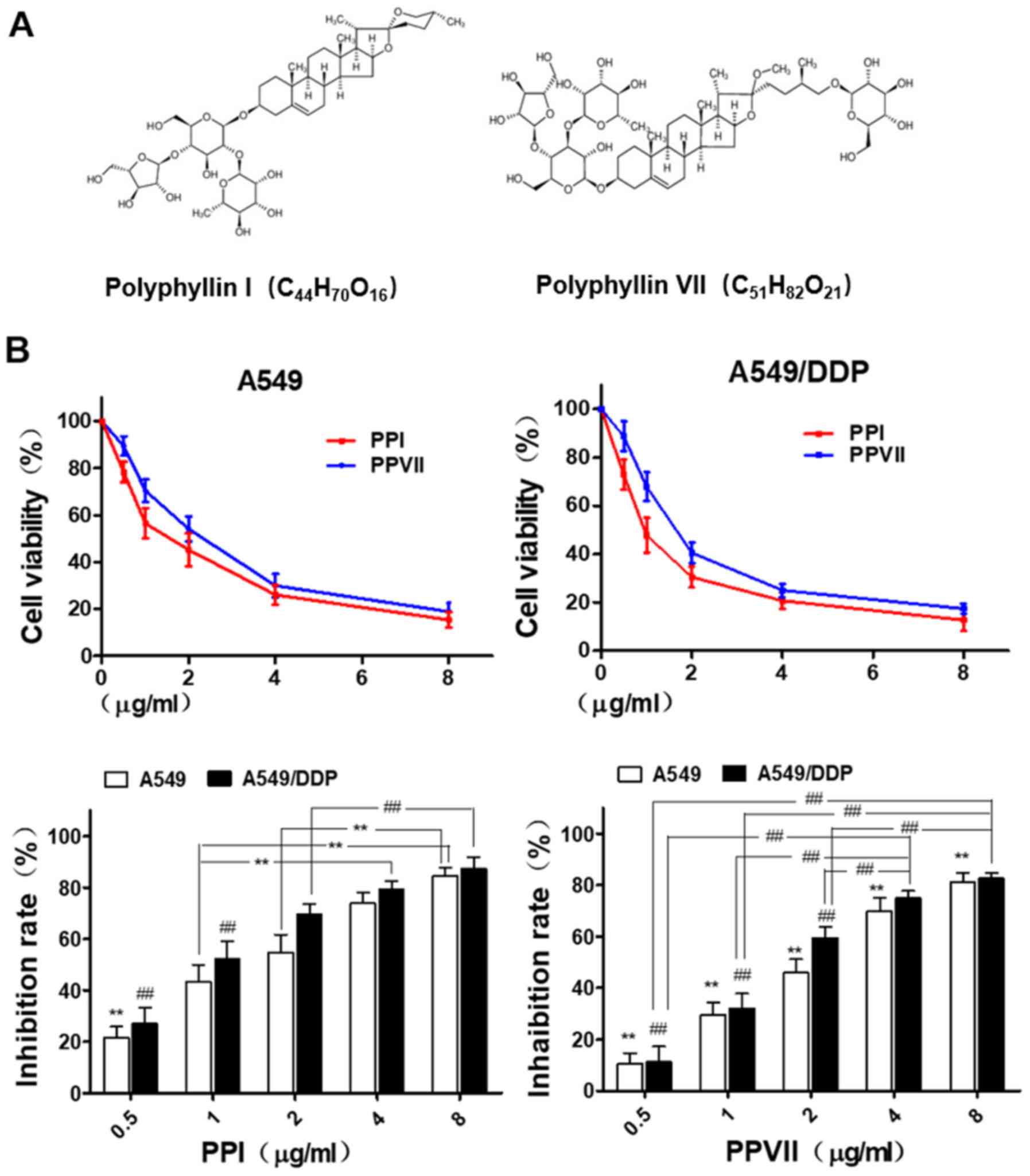 | Figure 2.PPI and PPVII induce cytotoxicity in
A549 and A549/DDP cells. (A) Chemical structure of PPI and PPVII.
(B) A549 and A549/DDP cells were treated with different dose of PPI
(0.5–8 µg/ml) and PPVII (0.5–8 µg/ml) for 24 h, respectively. In
A549 cells, the cell inhibition rate of 0.5 µg/ml PPI was
significantly different from other concentration groups
(**P<0.01); 1.0 µg/ml was significantly different from 4.0 and
8.0 µg/ml (**P<0.01), and 2.0 µg/ml was significantly different
from 8.0 µg/ml (**P<0.01). As for A549/DDP cells, the cell
inhibition rates of 0.5 µg/ml PPI and 1.0 µg/ml were both
significantly different from other concentration groups
(##P<0.01), and 2.0 µg/ml was significantly different
from 8.0 µg/ml (##P<0.01). In A549 cells, there were
significant differences in cell inhibition rates of PPVII among
different concentration groups (**P<0.01); as for A549/DDP
cells, there were significant differences among different
concentration groups except for 4.0 and 8.0 µg/ml
(##P<0.01). PPI, Polyphyllin I; PPVII, polyphyllin
VII; DDP, cisplatin. |
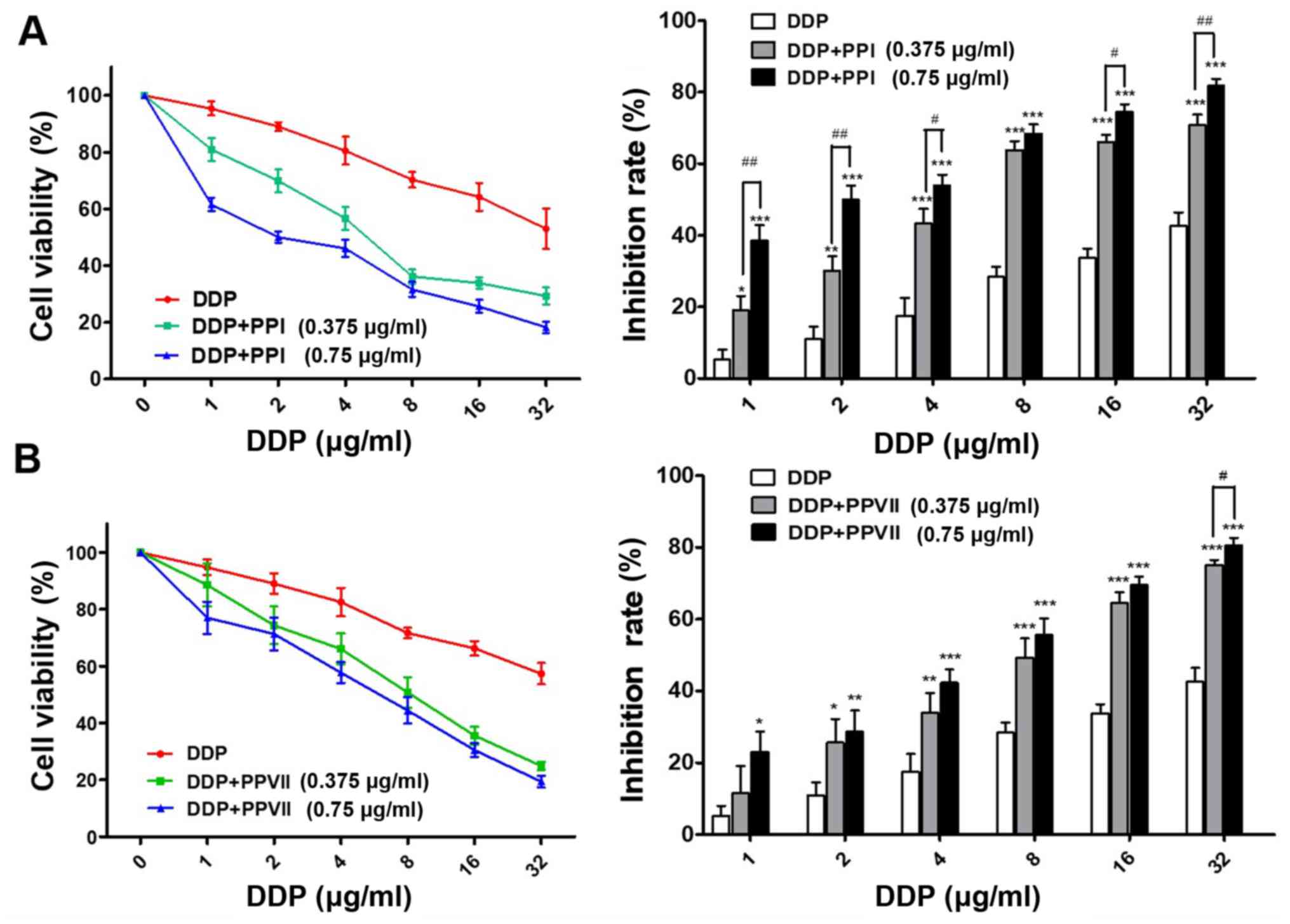
Chemosensitizing effects of PPI and
PPVII on A549/DDP cells
In order to determine whether PPI and PPVII can
reverse the drug resistance of A549/DDP cells to DDP, the present
study incubated cells with a range of doses of DDP in the absence
or presence of 0.375 µg/ml PPI, 0.75 µg/ml PPI, 0.375 µg/ml PPVII
and 0.75 µg/ml PPVII for 24 h. Using the CCK-8 assay, it was
revealed that PPI or PPVII in combination with DDP treatment
drastically inhibited the cell viability of A549/DDP cells compared
with DDP treatment alone; either PPI or PPVII with DDP treated
cells demonstrated increased sensitivity to DDP at all
concentrations (Fig. 3A and B).
Furthermore, the IC50 value of the DDP plus 0.375 µg/ml
PPI group (6.17±0.88 µg/ml) and DDP plus 0.75 µg/ml PPI group
(2.30±0.33 µg/ml) was significantly decreased compared with the
IC50 value of the DDP group (33.42±4.05 µg/ml;
P<0.01; P<0.01; Table I). The
IC50 value of the DDP plus 0.375 µg/ml PPVII group
(8.37±1.17 µg/ml) and DDP plus 0.75 µg/ml PPVII group (5.72±1.03
µg/ml) were also significantly decreased compared with the
IC50 value of the DDP group (33.42±4.05 µg/ml;
P<0.05; P<0.01, respectively; Table I). These results suggested that PPI
and PPVII both have chemosensitizing effects on A549/DDP cells.
 | Table I.PPI and PPVII reduce the
IC50 value of cisplatin in A549/DDP cells |
Table I.
PPI and PPVII reduce the
IC50 value of cisplatin in A549/DDP cells
| µg/ml | DDP | DDP+PPI (0.375
µg/ml) | DDP+PPI (0.75
µg/ml) |
|---|
| IC50 of
DDP | 33.42±4.05 |
6.17±0.88b |
2.30±0.33b |
|
|
| DDP+PPVII (0.375
µg/ml) | DDP+PPVII (0.75
µg/ml) |
|
|
|
8.37±1.17a |
5.72±1.03b |
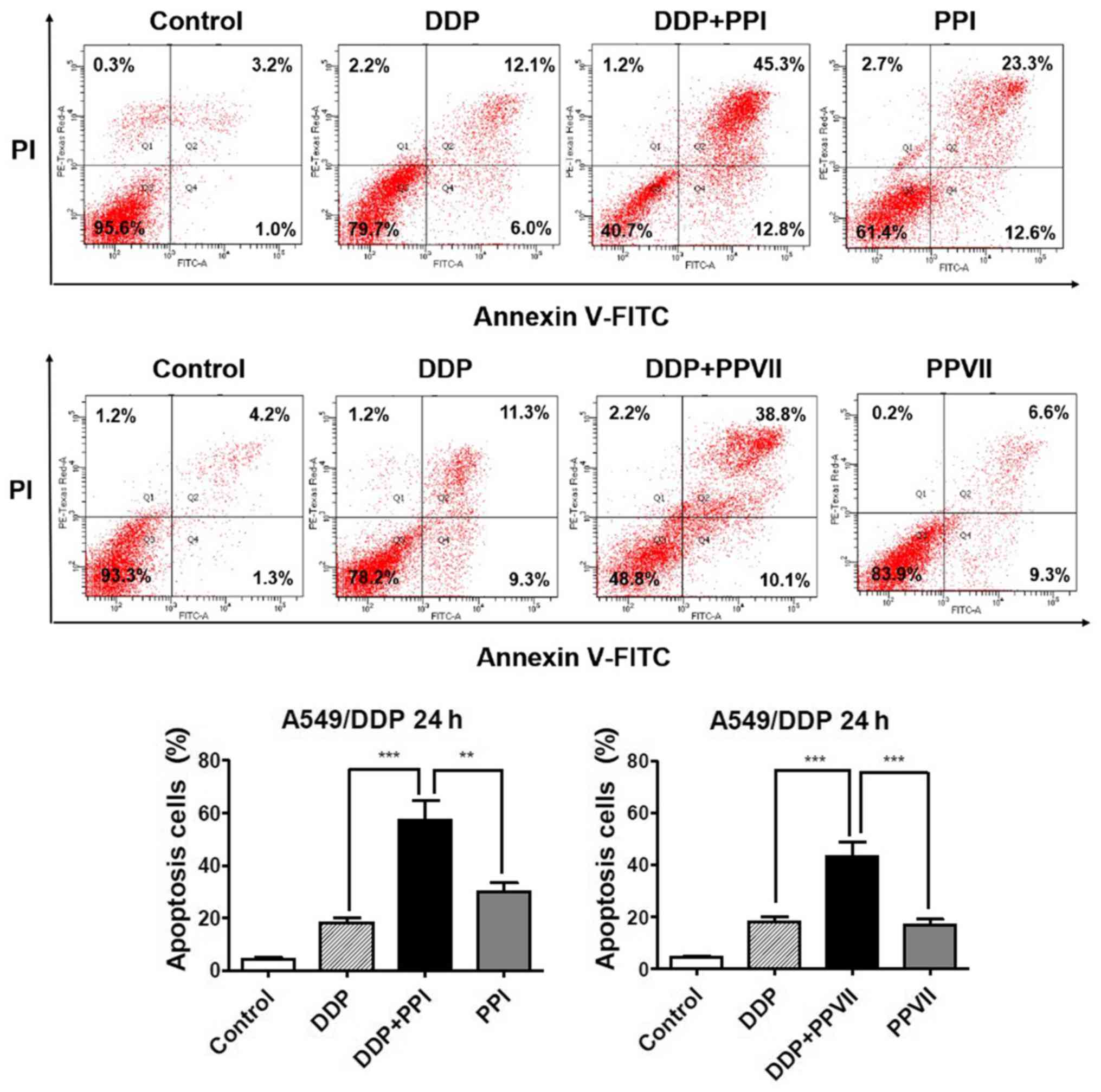
PPI and PPVII increases DDP-induced
apoptosis in A549/DDP cells
Apoptosis is one of the major mechanisms responsible
for DDP inhibiting cell proliferation (4) and failure of the apoptotic pathway may
be the leading factor in DDP resistance (32). The present study subsequently
investigated the roles of PPI and PPVII in DDP-induced apoptosis in
A549/DDP cells using the Annexin V- FITC/PI apoptosis analysis
(Fig. 4). The results demonstrated
that the combination of DDP (6 µg/ml) with PPI (0.75 µg/ml)
significantly increased apoptosis rate (57.2±5.2%) compared with
DDP (18.6±1.4%), or PPI alone (29.9±4.3%; P<0.01; P<0.01,
respectively) and combination of DDP (6 µg/ml) with PPVII (0.75
µg/ml) also significantly increased apoptosis rate (43.3±3.7%)
compared with DDP (18.6±1.4%), or PPVII alone (16.7±1.8%;
P<0.01; P<0.01, respectively). These results revealed that
PPI and PPVII could promote DDP-induced apoptosis in A549/DDP
cells.
PPI and PPVII enhance DDP-induced
apoptosis via the p53 pathway and caspase-dependent pathway in
A549/DDP cells
In order to determine the molecular mechanism
underlying the way in which PPI and PPVII enhance the DDP-induced
apoptosis in A549/DDP cells, the present study investigated the
expression of key regulators, including p53, Bax, Bcl-2, PARP,
cleaved-PARP, pro-caspase-3 and cleaved-caspase-3 protein, which
are the most important apoptosis regulators. The western blotting
results demonstrated that DDP in combination with either PPI or
PPVII treatment exhibited synergistic apoptosis-promoting effects
in A549/DDP cells. Compared with the control group, DDP treatment
upregulated the expression levels of p53, while the combination of
DDP with either PPI or PPVII markedly enhanced this effect, which
consequently upregulated the pro-apoptotic protein Bax and
downregulated anti-apoptotic protein Bcl-2 expression compared with
DDP treatment alone (Fig. 5A). The
ratio of Bax to Bcl-2 was enhanced, which led to the enhanced
activation of PARP, as well as enhanced activation of caspase-3 and
ultimately initiated enhanced cell apoptosis. A decrease in PARP
and pro-caspase-3 was observed, as well as an increase in
cleaved-PARP and cleaved-caspase-3 in DDP with either the PPI or
PPVII treated group when compared with the DDP treated group
(Fig. 5B). These results indicated
that PPI and PPVII enhanced DDP-induced apoptosis via the p53
pathway and caspase-dependent pathway in A549/DDP cells.
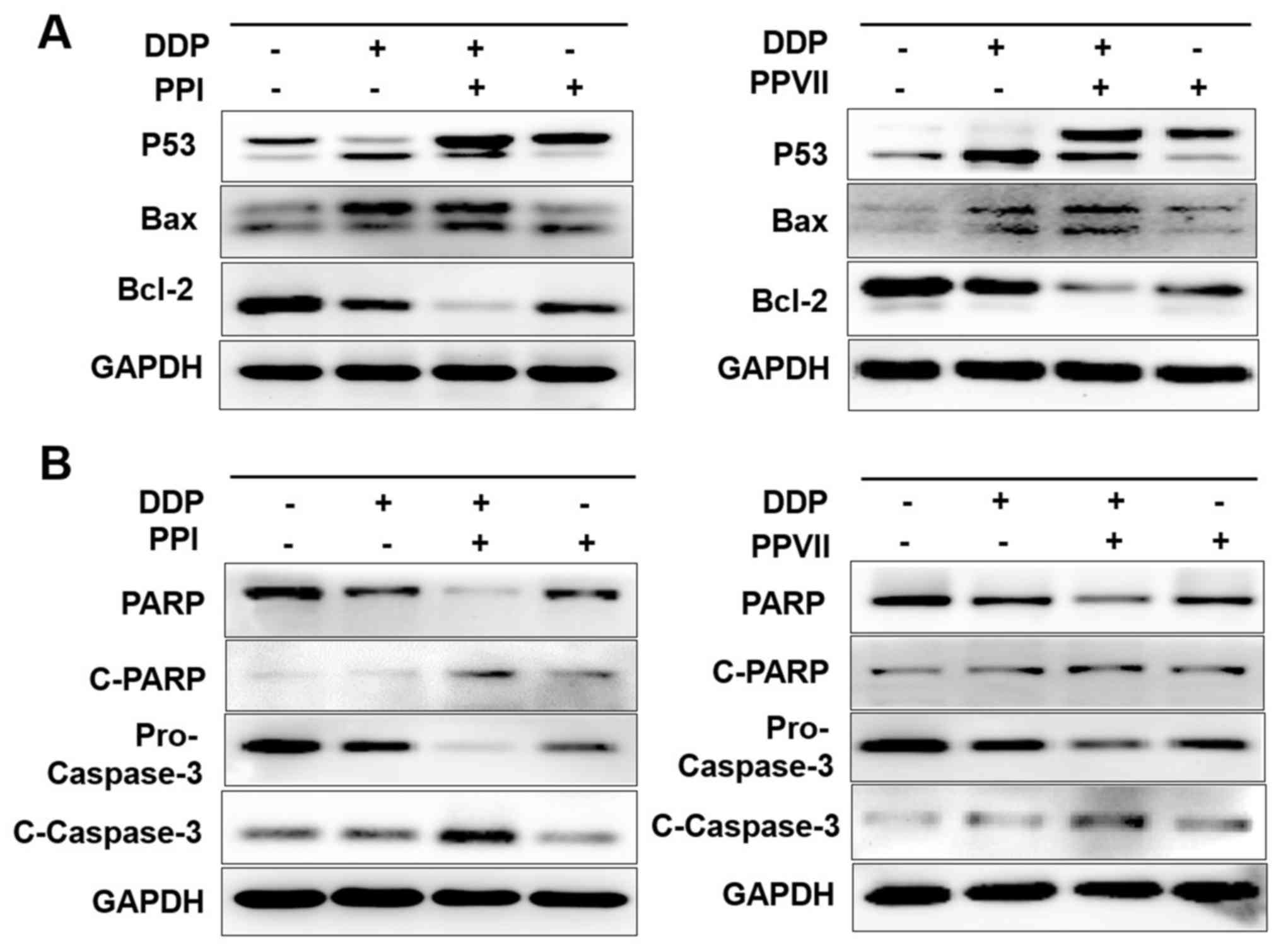 | Figure 5.PPI and PPVII enhance DDP-induced
apoptosis through the P53 pathway and caspases-dependent pathway.
A549/DDP cells were treated with DDP (6 µg/ml), PPI (0.75 µg/ml),
PPVII (0.75 µg/ml), or DDP (6 µg/ml) in combination with either PPI
(0.75 µg/ml) or PPVII (0.75 µg/ml) for 24 h, respectively. The
protein expression levels of P53, Bax, Bcl-2 (A), PARP, C-PARP,
pro-Caspase-3, C-Caspase-3 (B) were measured using western
blotting. PPI, Polyphyllin I; PPVII, polyphyllin VII; DDP,
cisplatin; C-PARP, cleaved-poly (ADP-ribose) polymerase 1. |
PPI and PPVII reverse the EMT and suppress the
CIP2A/AKT/mTOR pathway. It has previously been demonstrated that
the A549/DDP cell line exhibited the EMT phenotype and that CIP2A,
p-AKT, AKT and p-mTOR were overexpressed in A549/DDP cells
(Fig. 1B and C). Therefore, the
present study investigated the effect of PPI and PPVII on the EMT
and CIP2A/AKT/mTOR pathway in A549/DDP cells. A549/DDP cells were
incubated with DDP (6 µg/ml), PPI (0.75 µg/ml), PPVII (0.75 µg/ml)
or DDP (6 µg/ml) in combination with either PPI (0.75 µg/ml) or
PPVII (0.75 µg/ml) for 24 h. Western blot analysis revealed that
treatment with PPI and PPVII reversed the EMT in A549/DDP cells, as
demonstrated by the upregulation of epithelial marker E-cadherin
and downregulation of the mesenchymal markers vimentin and α-SMA
(Fig. 6A). DDP or PPI, PPVII
treatment alone could all downregulate the expression of CIP2A and
inhibit its downstream AKT phosphorylation and mTOR
phosphorylation, while the combination of DDP with either PPI or
PPVII enhanced these effects (Fig.
6B). PPI and PPVII could downregulate the AKT expression level,
which may further suppress the AKT pathway. These results suggested
that the combination of DDP with either PPI or PPVII had
synergistic inhibitory effects on CIP2A/AKT/mTOR pathway in
A549/DDP cells.
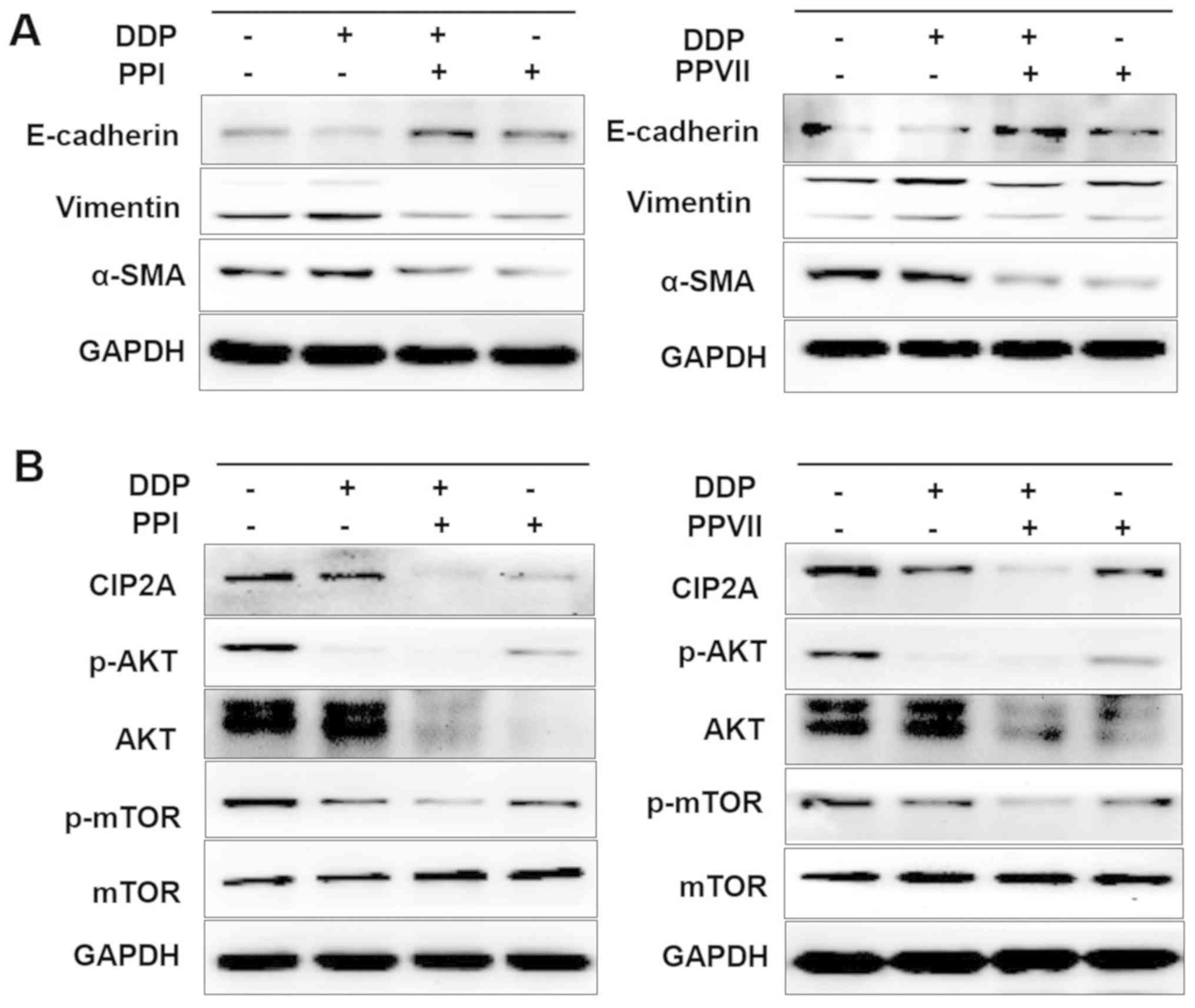 | Figure 6.PPI and PPVII reverse EMT and
suppress the CIP2A/AKT/mTOR pathway. A549/DDP cells were incubated
with DDP (6 µg/ml), PPI (0.75 µg/ml), PPVII (0.75 µg/ml), or DDP (6
µg/ml) in combination with either PPI (0.75 µg/ml) or PPVII (0.75
µg/ml) for 24 h, then cells were harvested for further western
blotting analysis. (A) The protein expression levels of E-cadherin,
vimentin, α-SMA in A549/DDP cells were measured. (B) The protein
expression levels of CIP2A, p-AKT, AKT, p-mTOR, m-TOR were
measured. PPI, Polyphyllin I; PPVII, polyphyllin VII; DDP,
cisplatin; p-AKT, phosphorylated-protein kinase B; mTOR, mammalian
target of rapamycin; CIP2A, cancerous inhibitor of protein
phosphatase 2A; SMA, smooth muscle actin; E, epithelial. |
Discussion
DDP is the backbone of numerous mainstream
chemotherapeutic regimens for advanced NSCLC. However, resistance
to DDP in NSCLC is increasing and is a major limitation in its
clinical application. Therefore, novel therapeutic strategies
utilizing natural compounds may diminish the side-effects of
chemotherapy agents. The present study identified the
chemosensitizing effects of natural compounds PPI and PPVII in
DDP-resistant NSCLC with the aim of gaining insight into the
underlying molecular mechanism. The data revealed the
chemosensitizing activities of PPI and PPVII in combination with
DDP against A549/DDP cells.
As is already known, DDP acts by binding to DNA and
inhibiting DNA synthesis, suppressing cell division and inducing
apoptosis (33). Apoptosis of tumor
cells is a key indicator for measuring the effectiveness of
chemotherapy (34). One of the
hallmarks of chemoresistance is evading apoptosis (33). In the present study, PPI and PPVII at
low doses markedly enhanced the DDP-induced apoptosis effect in
A549/DDP cells. The effects of PPI or PPVII on the expression of
key apoptosis regulators were further investigated and the results
revealed that PPI and PPVII treatment upregulated the expression of
P53 and Bax, and downregulated Bcl-2 expression. The ratio of Bax
to Bcl-2 was enhanced, which led to the activation of PARP, as well
as activation of caspase-3. This was reflected by a decrease in
PARP and an increase of cleaved-PARP and cleaved-caspase3, which
ultimately initiated cell apoptosis. These results indicated that
PPI and PPVII enhanced DDP-induced apoptosis via p53 and the
caspase-dependent pathway in A549/DDP cells. In addition, the
results of the present study showed an interesting phenomenon that
P53 was double- or multiple-banded and the adjacent bands are very
close to each other, even if multiple western blot experiments were
repeated or switched to other P53 monoclonal antibodies. To solve
this problem, the literature was consulted and possible
explanations was investigated. P53 has many modifications, such as
phosphorylation, ubiquitination, acetylation and methylation
(35–37). According to the current experimental
results, it is unclear what type of modification P53 underwent, but
the present study hypothesizes that this is a valuable research
direction and further experimental studies may be needed to
determine which modification of P53 is mainly affected by these
drugs.
A number of studies have demonstrated the
association between EMT to the emergence of drug resistance
(5–7), therefore ongoing research has been
focused on determining ways to reverse the EMT with the aim of
delaying or preventing drug resistance. CIP2A is a novel human
oncoprotein that is overexpressed in NSCLC and associated with poor
prognosis (8). CIP2A acts as an
‘oncogenic nexus’ to participate in multiple pathways, including
the PI3K/AKT/mTOR, RAS/MEK/ERK and the Wnt/β-catenin pathway
(9). In addition, CIP2A cooperates
with the oncogene H-Ras via the MEK/ERK pathway to facilitate the
EMT process (11). Furthermore,
CIP2A expression is associated with DDP resistance (15,16) and
its downstream AKT/mTOR pathway plays an important role in
mediating DDP resistance in lung cancer (15–17). The
results from the present study revealed that the A549/DDP cell line
exhibited the EMT phenotype and CIP2A, p-AKT, AKT and p-mTOR were
overexpressed in A549/DDP cells. PPI and PPVII could reverse the
EMT of A549/DDP cells, as evidenced by upregulation of the
epithelial marker E-cadherin and downregulation of the mesenchymal
markers vimentin and α-SMA. DDP treatment alone could downregulate
the expression of CIP2A and inhibit the downstream phosphorylation
of AKT and mTOR, while the combination of DDP with either PPI or
PPVII enhanced these effects. In addition, PPI and PPVII could
downregulate the AKT expression level. These results suggest that
the combination of DDP with either PPI or PPVII had synergistic
inhibitory effects on the CIP2A/AKT/mTOR pathway in A549/DDP
cells.
The function of CIP2A in apoptosis is not yet fully
understood. A number of studies confirmed that CIP2A is involved in
apoptosis and its depletion contributes to cell apoptosis in a
number of different types of cancer (14,38,39).
Functional studies also confirmed the potential role of CIP2A
depletion or downregulation in sensitizing cancer cells to several
chemotherapeutic agents, including DDP (17,40). In
the present study, PPI and PPVII enhanced DDP-induced apoptosis via
p53 and the caspase-dependent pathway; at the same time, PPI and
PPVII had synergistic effects with DDP on CIP2A and its downstream
AKT/mTOR signaling cascade in A549/DDP cells. As a tumor
suppressor, p53 is activated in response to DNA-damaging stress,
which can induce apoptosis in either transient or permanent cell
cycle arrests. p53 has complicated interactions with CIP2A via
PPP2R1A (9) and there is also
abundant crosstalk between p53 and the AKT/mTOR signaling pathway,
which could determine the choice of cell response to p53 (41) and requires further investigation.
The results of the present study determined that PPI
and PPVII enhanced sensitivity to DDP, potentially through inducing
increased apoptosis via the p53 pathway, reversing the EMT and
suppressing the CIP2A/AKT/mTOR pathway in A549/DDP cells. These
data strongly suggested the potential beneficial effects of PPI and
PPVII on patients suffering with DDP-resistant NSCLC.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81473485) and the
Natural Science Foundation of Shandong Province (grant no.
2014ZRE27321).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
FF and PC analyzed the data and wrote the
manuscript. FF and PC performed the experiments. CW and YW assisted
in preparing the experiments. WW designed the study. All authors
have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Szejniuk WM, Robles AI, McCulloch T,
Falkmer UGI and Røe OD: Epigenetic predictive biomarkers for
response or outcome to platinum-based chemotherapy in non-small
cell lung cancer, current state-of-art. Pharmacogenomics J.
19:5–14. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cao L, Chen J, Ou B, Liu C, Zou Y and Chen
Q: GAS5 knockdown reduces the chemo-sensitivity of non-small cell
lung cancer (NSCLC) cell to cisplatin (DDP) through regulating
miR-21/PTEN axis. Biomed Pharmacother. 93:570–579. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dasari S and Tchounwou PB: Cisplatin in
cancer therapy: Molecular mechanisms of action. Eur J Pharmacol.
740:364–378. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Du B and Shim JS: Targeting
epithelial-mesenchymal transition (EMT) to overcome drug resistance
in cancer. Molecules. 21:E9652016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shibue T and Weinberg RA: EMT, CSCs, and
drug resistance: The mechanistic link and clinical implications.
Nat Rev Clin Oncol. 14:611–629. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Song Y, Ye M, Zhou J, Wang Z and Zhu X:
Targeting E-cadherin expression with small molecules for digestive
cancer treatment. Am J Transl Res. 11:3932–3944. 2019.PubMed/NCBI
|
|
8
|
Liu Z, Ma L, Wen ZS, Hu Z, Wu FQ, Li W,
Liu J and Zhou GB: Cancerous inhibitor of PP2A is targeted by
natural compound celastrol for degradation in non-small-cell lung
cancer. Carcinogenesis. 35:905–914. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
De P, Carlson J, Leyland-Jones B and Dey
N: Oncogenic nexus of cancerous inhibitor of protein phosphatase 2A
(CIP2A): An oncoprotein with many hands. Oncotarget. 5:4581–4602.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tang M, Shen JF, Li P, Zhou LN, Zeng P,
Cui XX, Chen MB and Tian Y: Prognostic significance of CIP2A
expression in solid tumors: A meta-analysis. PLoS One.
13:e01996752018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu Y, Gu TT and Zheng PS: CIP2A cooperates
with H-Ras to promote epithelial mesenchymal transition in
cervical-cancer progression. Cancer Lett. 356:646–655. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lin YC, Chen KC, Chen CC, Cheng AL and
Chen KF: CIP2A-mediated Akt activation plays a role in
bortezomib-induced apoptosis in head and neck squamous cell
carcinoma cells. Oral Oncol. 48:585–593. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yu HC, Chen HJ, Chang YL, Liu CY, Shiau
CW, Cheng AL and Chen KF: Inhibition of CIP2A determines
erlotinib-induced apoptosis in hepatocellular carcinoma. Biochem
Pharmacol. 85:356–366. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu P, Yao J, He J, Zhao L, Wang X, Li Z
and Qian J: CIP2A down regulation enhances the sensitivity of
pancreatic cancer cells to gemcitabine. Oncotarget. 7:14831–14840.
2016.PubMed/NCBI
|
|
15
|
Zhang X, Xu B, Sun C, Wang L and Miao X:
Knockdown of CIP2A sensitizes ovarian cancer cells to cisplatin: An
in vitro study. Int J Clin Exp Med. 8:16941–16947. 2015.PubMed/NCBI
|
|
16
|
Gao F, Wang X, Chen S, Xu T, Wang X, Shen
Y, Dong F, Zhong S and Shen Z: CIP2A depletion potentiates the
chemosensitivity of cisplatin by inducing increased apoptosis in
bladder cancer cells. Oncol Rep. 40:2445–2454. 2018.PubMed/NCBI
|
|
17
|
Liu LZ, Zhou XD, Qian G, Shi X, Fang J and
Jiang BH: AKT1 amplification regulates cisplatin resistance in
human lung cancer cells through the mammalian target of
rapamycin/p70S6K1 pathway. Cancer Res. 67:6325–6332. 2017.
View Article : Google Scholar
|
|
18
|
Ho JW, Leung YK and Chan CP: Herbal
medicine in the treatment of cancer. Curr Med Chem Anticancer
Agents. 2:209–214. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shi YM, Yang L, Geng YD, Zhang C and Kong
LY: Polyphyllin I induced-apoptosis is enhanced by inhibition of
autophagy in human hepatocellular carcinoma cells. Phytomedicine.
22:1139–49. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gu L, Feng J, Zheng Z, Xu H and Yu W:
Polyphyllin I inhibits the growth of ovarian cancer cells in nude
mice. Oncol Lett. 12:4969–4974. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chang J, Li Y, Wang X, Hu S, Wang H, Shi
Q, Wang Y and Yang Y: Polyphyllin I suppresses human osteosarcoma
growth by inactivation of Wnt/β-catenin pathway in vitro and in
vivo. Sci Rep. 7:76052017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dong R, Guo J, Zhang Z, Zhou Y and Hua Y:
Polyphyllin I inhibits gastric cancer cell proliferation by
downregulating the expression of fibroblast activation protein
alpha (FAP) and hepatocyte growth factor (HGF) in cancer-associated
fibroblasts. Biochem Biophys Res Commun. 497:1129–1134. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang Q, Chen W, Xu Y, Lv X, Zhang M and
Jiang H: Polyphyllin I modulates MALAT1/STAT3 signaling to induce
apoptosis in gefitinib-resistant non-small cell lung cancer.
Toxicol Appl Pharmacol. 356:1–7. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang C, Jia X, Bao J, Chen S, Wang K,
Zhang Y, Li P, Wan JB, Su H, Wang Y, et al: Polyphyllin VII induces
apoptosis in HepG2 cells through ROS-mediated mitochondrial
dysfunction and MAPK pathways. BMC Complement Altern Med.
16:582016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lin Z, Liu Y, Li F, Wu J, Zhang G, Wang Y,
Lu L and Liu Z: Anti-lung cancer effects of polyphyllin vi and vii
potentially correlate with apoptosis in vitro and in vivo.
Phytother Res. 29:1568–1576. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hsieh MJ, Chien SY, Lin JT, Yang SF and
Chen MK: Polyphyllin G induces apoptosis and autophagy cell death
in human oral cancer cells. Phytomedicine. 23:1545–1554. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cai X, Guo L, Pei F, Chang X and Zhang R:
Polyphyllin G exhibits antimicrobial activity and exerts anticancer
effects on human oral cancer OECM-1 cells by triggering G2/M cell
cycle arrest by inactivating cdc25C-cdc2. Arch Biochem Biophys.
644:93–99. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Han W, Hou G and Liu L: Polyphyllin I
(PPI) increased the sensitivity of hepatocellular carcinoma HepG2
cells to chemotherapy. Int J Clin Exp Med. 8:20664–20669.
2015.PubMed/NCBI
|
|
29
|
Al Sawah E, Marchion DC, Xiong Y, Ramirez
IJ, Abbasi F, Boac BM, Bush SH, Bou Zgheib N, McClung EC,
Khulpateea BR, et al: The Chinese herb polyphyllin D sensitizes
ovarian cancer cells to cisplatin-induced growth arrest. J Cancer
Res Clin Oncol. 141:237–42. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lou W, Chen Y, Zhu KY, Deng H, Wu T and
Wang J: Polyphyllin I overcomes EMT-associated resistance to
erlotinib in lung cancer cells via IL-6/STAT3 Pathway Inhibition.
Biol Pharm Bull. 40:1306–1313. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li Y, Fan L, Sun Y, Miao X, Zhang F, Meng
J, Han J, Zhang D, Zhang R, Yue Z and Mei Q: Paris saponin VII from
trillium tschonoskii reverses multidrug resistance of Adriamycin
resistant MCF-7/ADR cells via P-glycoprotein inhibition and
apoptosis augmentation. J Ethnopharmacol. 154:728–734. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang H, Fei Z and Jiang H: Polyphyllin VII
increases sensitivity to gefitinib by modulating the elevation of
P21 in acquired gefitinib resistant non-small cell lung cancer. J
Pharmacol Sci. 134:190–196. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Galluzzi L, Senovilla L, Vitale I, Michels
J, Martins I, Kepp O, Castedo M and Kroemer G: Molecular mechanisms
of DDP resistance. Oncogene. 31:1869–1883. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu H, Li P, Li B, Sun P, Zhang J, Wang B
and Jia B: RKIP promotes Cisplatin-induced gastric cancer cell
death through NF-κB/Snail pathway. Tumour Biol. 36:1445–1153. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xu Y: Regulation of p53 responses by
post-translational modifications. Cell Death Differ. 10:400–403.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bode AM and Dong Z: Post-translational
modification of p53 in tumorigenesis. Nat Rev Cancer. 4:793–805.
2004. View
Article : Google Scholar : PubMed/NCBI
|
|
37
|
Park JH, Yang SW, Park JM, Ka SH, Kim JH,
Kong YY, Jeon YJ, Seol JH and Chung CH: Positive feedback
regulation of p53 transactivity by DNA damage-induced ISG15
modification. Nat Commun. 7:125132016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liu X, Duan C, Ji J, Zhang T, Yuan X,
Zhang Y, Ma W, Yang J, Yang L, Jiang Z, et al: Cucurbitacin B
induces autophagy and apoptosis by suppressing CIP2A/PP2A/mTORC1
signaling axis in human cisplatin resistant gastric cancer cells.
Oncol Rep. 38:271–278. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chao TT, Wang CY, Chen YL, Lai CC, Chang
FY, Tsai YT, Chao CH, Shiau CW, Huang YC, Yu CJ and Chen KF:
Afatinib induces apoptosis in NSCLC without EGFR mutation through
Elk-1-mediated suppression of CIP2A. Oncotarget. 6:2164–2179. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Xu P, Yao J, He J, Zhao L, Wang X, Li Z
and Qian J: CIP2A down regulation enhances the sensitivity of
pancreatic cancer cells to gemcitabine. Oncotarget. 7:14831–14840.
2016.PubMed/NCBI
|
|
41
|
Duan L and Maki CG: The IGF-1R/AKT pathway
determines cell fate in response to p53. Transl Cancer Res.
5:664–675. 2016. View Article : Google Scholar : PubMed/NCBI
|