Autophagy is upregulated to maintain cytosolic
homeostasis when the innate immune defence recognizes invasive
bacterial pathogens (16). However,
autophagy can be upregulated or downregulated in gastric epithelial
cells during H. pylori infection (17,18). The
present review focused on the molecular mechanisms currently
considered to be associated with H. pylori-mediated
autophagy. The hypothesis that the induction of autophagy can be a
novel therapeutic target for persistent H. pylori infection
was presented.
The 2016 Nobel Prize in Physiology or Medicine was
awarded to Yoshinori Ohsumi, who first illustrated that nutrient
deficiency induced extensive autophagy in yeast cells in 1992
(19). Autophagy is defined as the
segregation of organelles and cellular components within double
membrane vacuoles called autophagosomes (20). The fusion of autophagosomes and
lysosomes generates autophagolysosomes, which degrade cytoplasmic
contents (20). Therefore, autophagy
can be stimulated as an intracellular defence mechanism to
eliminate pathogens following their recognition by the innate
immune system (21). Autophagosomes
can deliver pathogens to lysosomes. Furthermore, autophagolysosomes
can degrade pathogens for cellular homeostasis (20,21).
Non-canonical autophagy is another type of ATG7- and
ATG3-independent autophagy, which has been described during the
development of the Drosophila midgut (28). Non-canonical autophagy has also
reported as an ATG5-independent signaling pathway of autophagy
(29). The non-canonical process of
autophagy does not occur from a double-membrane autophagosome and
is called LC3-associated phagocytosis (LAP) (22,30). LAP
promotes phagosome maturation and lysosomal fusion (31).
Autophagy not only eradicates pathogens, but also
serves a dual role in carcinogenesis. In 1980, a study demonstrated
that the process of autophagy could be induced in leukemic cells
following treatment with an antiproliferative drug (32). Our previous studies indicated that
matrine had potent antitumour activity against gastric cancer cells
(33,34). Autophagy is upregulated in gastric
cancer cells during this antitumour process, and autophagy acts as
a cytoprotective mechanism to overcome lethal stress (33). Additionally, combination treatment
with matrine and autophagy inhibitors can enhance the antitumour
effect of matrine in gastric cancer (34). Our previous study further
demonstrated that matrine exhibited antitumour activity and induced
autophagy in hepatocellular carcinoma cells (35). The extensive activation of autophagy
induces autophagic cell death (35).
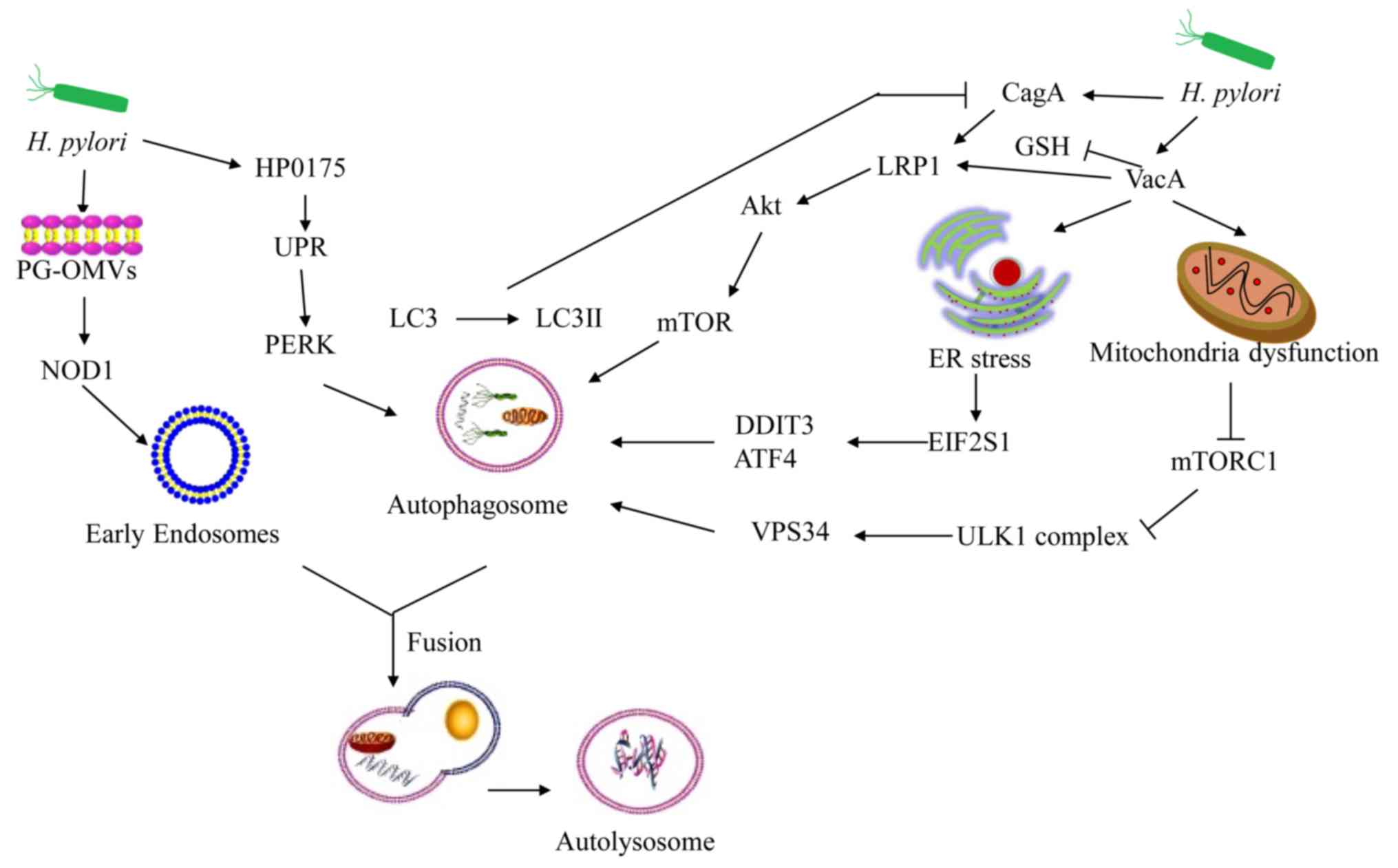
A physiological mechanism of outer membrane vesicles
(OMVs) from bacteria can deliver peptidoglycans into the host cell
cytosol and induce an immune response in vivo (51). OMVs from H. pylori can induce
autophagy, which is essential for proinflammatory chemokine
production (52). OMVs rely on the
nucleotide-binding oligomerization domain-1-receptor interacting
serine/threonine kinase 2 signaling pathway, which is essential for
the induction of autophagy and the production of interleukin 8
(52,53). In addition, H. pylori OMVs
induce autophagosome formation, which is not dependent on VacA
(52). H. pylori secretes
HP0175, which has been identified as an inducer of apoptosis in
gastric epithelial cells (54).
HP0175 can also upregulate the expression of autophagy-associated
genes independent of functional VacA during acute infection
(17).
VacA is a critical virulence factor involved in the
pathogenesis of peptic ulceration and gastric cancer (55). The toxins of VacA can induce a series
of intracellular alterations, including cell vacuolation, membrane
channel formation, disruption of endosomal/lysosomal function,
apoptosis and immunomodulation (56). VacA localizes in the mitochondria and
induces their dysfunction (57).
VacA relies on the inhibition of rapamycin complex 1 (mTORC1),
which coordinates nutrients and energy stress signals in order to
promote metabolic homeostasis (58).
In VacA-intoxicated cells, the VacA-dependent inhibition of mTORC1
signaling results in the activation of cellular autophagy via the
ULK1 complex (59). Low-density
lipoprotein receptor-related protein-1 (LRP1) is the receptor for
VacA-induced autophagy (60). VacA
forms LRP1 conjugates in order to regulate the formation of
autophagosomes and autolysosomes (60). Additionally, VacA can induce
autophagy via endoplasmic reticulum (ER) stress (61). Inhibition of autophagy can decrease
VacA-induced cell death in AGS cells (61). Tribble pseudokinase 3 (TRIB3) serves
an important role in ER stress-induced autophagy (61,62).
VacA can trigger ER stress and increase the expression of TRIB3 in
AGS cells (61). Knockdown of the ER
stress effector protein can significantly decrease the formation of
autolysosomes and cell death (61).
Therefore, VacA causes autophagic cell death via ER stress in
gastric epithelial cells. Additionally, VacA-induced autophagy can
degrade the toxins and limit host cell damage, leading to the
maintenance of cellular homeostasis (63). VacA-induced autophagy does not affect
the formation of VacA-large vacuoles (49).
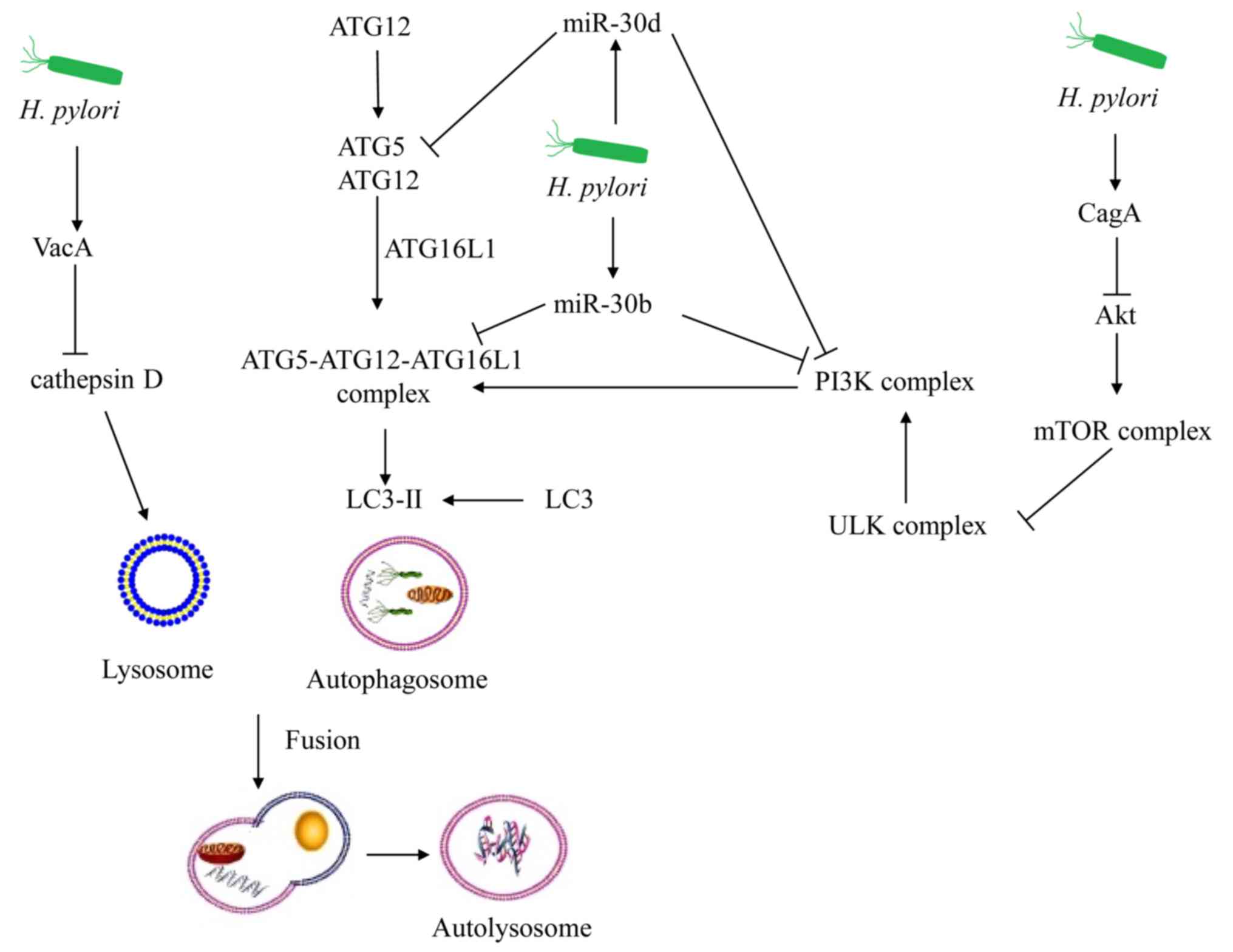
Exposure to VacA for prolonged periods may mimic the
chronic infection model of VacA+ H. pylori strains (76). Autophagy is disrupted by the
prolonged co-culture of VacA, since cathepsin D expression is
inhibited in autophagosomes (18).
Therefore, VacA can further inhibit autophagy in gastric epithelial
cells during chronic infection of H. pylori.
Our previous studies demonstrated that autophagy
exhibited a cytoprotective function in cancer cells and that it
could induce autophagic cell death at different stages of cancer
formation (33–35). H. pylori may disturb
homeostasis in host cells during acute infection, notably via the
secretion of virulence factors. Autophagy is an important defence
mechanism that can restrict bacterial survival and growth. Gastric
epithelial cells can induce canonical autophagy in order to
maintain homeostasis during acute infection with H. pylori.
Chronic infections with H. pylori can cause the dysfunction
of autophagy-associated proteins. The inhibition of autophagy can
lead to persistent infection. H. pylori can resist
antibiotic treatment, and, as a consequence, the chronic infection
of this bacterial strain has become a global health issue. During
infection, the induction of autophagy, which maintains cellular
homeostasis, is inhibited. By upregulating autophagy-associated
proteins in gastric epithelial cells, H. pylori can be
eliminated. This strategy can be applied with the use of autophagy
inducers as novel therapeutic agents. Although the mechanism of
multi-drug-resistance acquired by H. pylori relies on
associated virulence factors to cause downregulation of autophagy
and maintenance of persistent infection, the precise identification
of the proteins involved in this signaling pathway remains
unclear.
Not applicable.
The present study was supported by the National
Natural Science Foundation of China (grant no. 31770537).
Data sharing is not applicable to this article, as
no datasets were generated or analyzed during the current
study.
YL contributed to the planning and design of the
study. FZ, CC, JH, RS, JZ, ZH, HC and YL were responsible for the
literature search and the writing of the manuscript. FZ and YL
performed revisions of the manuscript. All authors have read and
approved the final manuscript for publication.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Blaser MJ: Gastric Campylobacter-like
organisms, gastritis, and peptic ulcer disease. Gastroenterology.
93:371–383. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Strugatsky D, McNulty R, Munson K, Chen
CK, Soltis SM, Sachs G and Luecke H: Structure of the proton-gated
urea channel from the gastric pathogen Helicobacter pylori.
Nature. 493:255–258. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Schistosomes, liver flukes and
Helicobacter pylori, . IARC Working Group on the Evaluation of
Carcinogenic Risks to Humans. Lyon, 7–14 June 1994. IARC monographs
on the evaluation of carcinogenic risks to humans. 61:1–241.
1994.PubMed/NCBI
|
|
4
|
Fukase K, Kato M, Kikuchi S, Inoue K,
Uemura N, Okamoto S, Terao S, Amagai K, Hayashi S and Asaka M;
Japan Gast Study Group, : Effect of eradication of Helicobacter
pylori on incidence of metachronous gastric carcinoma after
endoscopic resection of early gastric cancer: An open-label,
randomised controlled trial. Lancet. 372:392–397. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yoon SB, Park JM, Lim CH, Cho YK and Choi
MG: Effect of Helicobacter pylori eradication on
metachronous gastric cancer after endoscopic resection of gastric
tumors: A meta-analysis. Helicobacter. 19:243–248. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lin TF and Hsu PI: Second-line rescue
treatment of Helicobacter pylori infection: Where are we
now? World J Gastroenterol. 24:4548–4553. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Song Z, Zhou L, Zhang J, He L, Bai P and
Xue Y: Hybrid therapy as first-line regimen for Helicobacter
pylori eradication in populations with high antibiotic
resistance rates. Helicobacter. 21:382–388. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tan B, Yang JC, Young CL, Bishu S, Owyang
SY, El-Zaatari M, Zhang M, Grasberger H, Qian JM and Kao JY:
Helicobacter pylori Antimicrobial susceptibility
testing-guided salvage therapy in the USA: A real life experience.
Dig Dis Sci. 63:437–445. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lee JW, Kim N, Nam RH, Park JH, Kim JM,
Jung HC and Song IS: Mutations of Helicobacter pylori
associated with fluoroquinolone resistance in Korea. Helicobacter.
16:301–310. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kalach N, Bontems P and Raymond J:
Helicobacter pylori infection in children. Helicobacter. 22
(Suppl 1):2017.doi: 10.1111/hel.12414. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ranjbar R, Khamesipour F, Jonaidi-Jafari N
and Rahimi E: Helicobacter pylori in bottled mineral water:
Genotyping and antimicrobial resistance properties. BMC Microbiol.
16:402016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Taneike I, Nami A, O'Connor A, Fitzgerald
N, Murphy P, Qasim A, O'Connor H and O'Morain C: Analysis of drug
resistance and virulence-factor genotype of Irish Helicobacter
pylori strains: Is there any relationship between resistance to
metronidazole and cagA status? Aliment Pharmacol Ther. 30:784–790.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kawahara T, Kohjima M, Kuwano Y, Mino H,
Teshima-Kondo S, Takeya R, Tsunawaki S, Wada A, Sumimoto H and
Rokutan K: Helicobacter pylori lipopolysaccharide activates
Rac1 and transcription of NADPH oxidase Nox1 and its organizer
NOXO1 in guinea pig gastric mucosal cells. Am J Physiol Cell
Physiol. 288:C450–C457. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shimizu T, Marusawa H, Watanabe N and
Chiba T: Molecular pathogenesis of Helicobacter
pylori-related gastric cancer. Gastroenterol Clin North Am.
44:625–638. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang JC and Chien CT: A new approach for
the prevention and treatment of Helicobacter pylori
infection via upregulation of autophagy and downregulation of
apoptosis. Autophagy. 5:413–414. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Levine B and Deretic V: Unveiling the
roles of autophagy in innate and adaptive immunity. Nat Rev
Immunol. 7:767–777. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Halder P, Datta C, Kumar R, Sharma AK,
Basu J and Kundu M: The secreted antigen, HP0175, of
Helicobacter pylori links the unfolded protein response
(UPR) to autophagy in gastric epithelial cells. Cell Microbiol.
17:714–729. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Raju D, Hussey S, Ang M, Terebiznik MR,
Sibony M, Galindo-Mata E, Gupta V, Blanke SR, Delgado A,
Romero-Gallo J, et al: Vacuolating cytotoxin and variants in
Atg16L1 that disrupt autophagy promote Helicobacter pylori
infection in humans. Gastroenterology. 142:1160–1171. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Takeshige K, Baba M, Tsuboi S, Noda T and
Ohsumi Y: Autophagy in yeast demonstrated with proteinase-deficient
mutants and conditions for its induction. J Cell Biol. 119:301–311.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Randow F and Youle RJ: Self and nonself:
How autophagy targets mitochondria and bacteria. Cell Host Microbe.
15:403–411. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huang J and Brumell JH: Bacteria-autophagy
interplay: A battle for survival. Nat Rev Microbiol. 12:101–114.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Scambler T, Feeley C and McDermott MF:
Protection against lupus-like inflammatory disease is in the LAP of
non-canonical autophagy. Ann Transl Med. 4:5132016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Parzych KR and Klionsky DJ: An overview of
autophagy: Morphology, mechanism, and regulation. Antioxid Redox
Signal. 20:460–473. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nishimura T and Mizushima N: The ULK
complex initiates autophagosome formation at phosphatidylinositol
synthase-enriched ER subdomains. Autophagy. 13:1795–1796. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Russell RC, Tian Y, Yuan H, Park HW, Chang
YY, Kim J, Kim H, Neufeld TP, Dillin A and Guan KL: ULK1 induces
autophagy by phosphorylating Beclin-1 and activating VPS34 lipid
kinase. Nat Cell Biol. 15:741–750. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Leidal AM, Levine B and Debnath J:
Autophagy and the cell biology of age-related disease. Nat Cell
Biol. 20:1338–1348. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Krokowski S and Mostowy S: Interactions
between Shigella flexneri and the Autophagy Machinery. Front Cell
Infect Microbiol. 6:172016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chang TK, Shravage BV, Hayes SD, Powers
CM, Simin RT, Wade Harper J and Baehrecke EH: Uba1 functions in
Atg7- and Atg3-independent autophagy. Nat Cell Biol. 15:1067–1078.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lindqvist LM, Simon AK and Baehrecke EH:
Current questions and possible controversies in autophagy. Cell
Death Discov. 1(pii): 150362015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lai SC and Devenish RJ: LC3-associated
phagocytosis (LAP): Connections with host autophagy. Cells.
1:396–408. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Martinez J, Malireddi RK, Lu Q, Cunha LD,
Pelletier S, Gingras S, Orchard R, Guan JL, Tan H, Peng J, et al:
Molecular characterization of LC3-associated phagocytosis reveals
distinct roles for Rubicon, NOX2 and autophagy proteins. Nature
Cell Biol. 17:893–906. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mikles-Robertson F, Dave C and Porter CW:
Apparent autophagocytosis of mitochondria in L1210 leukemia cells
treated in vitro with
4,4′-diacetyl-diphenylurea-bis(guanylhydrazone). Cancer Res.
40:1054–1061. 1980.PubMed/NCBI
|
|
33
|
Li Y, Zhang J, Ma H, Chen X, Liu T, Jiao
Z, He W, Wang F, Liu X and Zeng X: Protective role of autophagy in
matrineinduced gastric cancer cell death. Int J Oncol.
42:1417–1426. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang J, Li Y, Chen X, Liu T, Chen Y, He
W, Zhang Q and Liu S: Autophagy is involved in anticancer effects
of matrine on SGC-7901 human gastric cancer cells. Oncol Rep.
26:115–124. 2011.PubMed/NCBI
|
|
35
|
Zhang JQ, Li YM, Liu T, He WT, Chen YT,
Chen XH, Li X, Zhou WC, Yi JF and Ren ZJ: Antitumor effect of
matrine in human hepatoma G2 cells by inducing apoptosis and
autophagy. World J Gastroenterol. 16:4281–4290. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gutierrez MG, Master SS, Singh SB, Taylor
GA, Colombo MI and Deretic V: Autophagy is a defense mechanism
inhibiting BCG and Mycobacterium tuberculosis survival in
infected macrophages. Cell. 119:753–766. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dortet L, Mostowy S, Samba-Louaka A, Gouin
E, Nahori MA, Wiemer EA, Dussurget O and Cossart P: Recruitment of
the major vault protein by InlK: A Listeria monocytogenes
strategy to avoid autophagy. PLoS Pathog. 7:e10021682011.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Steele S, Brunton J, Ziehr B, Taft-Benz S,
Moorman N and Kawula T: Francisella tularensis harvests nutrients
derived via ATG5-independent autophagy to support intracellular
growth. PLoS Pathog. 9:e10035622013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Choy A, Dancourt J, Mugo B, O'Connor TJ,
Isberg RR, Melia TJ and Roy CR: The Legionella effector RavZ
inhibits host autophagy through irreversible Atg8 deconjugation.
Science. 338:1072–1076. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Newton HJ, Kohler LJ, McDonough JA,
Temoche-Diaz M, Crabill E, Hartland EL and Roy CR: A screen of
Coxiella burnetii mutants reveals important roles for
Dot/Icm effectors and host autophagy in vacuole biogenesis. PLoS
Pathog. 10:e10042862014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ligeon LA, Moreau K, Barois N, Bongiovanni
A, Lacorre DA, Werkmeister E, Proux-Gillardeaux V, Galli T and
Lafont F: Role of VAMP3 and VAMP7 in the commitment of Yersinia
pseudotuberculosis to LC3-associated pathways involving single-
or double-membrane vacuoles. Autophagy. 10:1588–1602. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Starr T, Child R, Wehrly TD, Hansen B,
Hwang S, López-Otin C, Virgin HW and Celli J: Selective subversion
of autophagy complexes facilitates completion of the Brucella
intracellular cycle. Cell Host Microbe. 11:33–45. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yu HB, Croxen MA, Marchiando AM, Ferreira
RB, Cadwell K, Foster LJ and Finlay BB: Autophagy facilitates
Salmonella replication in HeLa cells. MBio. 5:e00865–14. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
MacMicking JD, Taylor GA and McKinney JD:
Immune control of tuberculosis by IFN-gamma-inducible LRG-47.
Science. 302:654–659. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kimmey JM and Stallings CL: Bacterial
pathogens versus autophagy: Implications for therapeutic
interventions. Trends Mol Med. 22:1060–1076. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Starr T, Ng TW, Wehrly TD, Knodler LA and
Celli J: Brucella intracellular replication requires trafficking
through the late endosomal/lysosomal compartment. Traffic.
9:678–694. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Terebiznik MR, Vazquez CL, Torbicki K,
Banks D, Wang T, Hong W, Blanke SR, Colombo MI and Jones NL:
Helicobacter pylori VacA toxin promotes bacterial
intracellular survival in gastric epithelial cells. Infect Immun.
74:6599–6614. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Catrenich CE and Chestnut MH: Character
and origin of vacuoles induced in mammalian cells by the cytotoxin
of Helicobacter pylori. J Med Microbiol. 37:389–395. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Raju D and Jones NL: Methods to monitor
autophagy in H. pylori vacuolating cytotoxin A
(VacA)-treated cells. Autophagy. 6:138–143. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang YH, Wu JJ and Lei HY: When
Helicobacter pylori invades and replicates in the cells.
Autophagy. 5:540–542. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Parker H and Keenan JI: Composition and
function of Helicobacter pylori outer membrane vesicles.
Microbes Infect. 14:9–16. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Irving AT, Mimuro H, Kufer TA, Lo C,
Wheeler R, Turner LJ, Thomas BJ, Malosse C, Gantier MP, Casillas
LN, et al: The immune receptor NOD1 and kinase RIP2 interact with
bacterial peptidoglycan on early endosomes to promote autophagy and
inflammatory signaling. Cell Host Microbe. 15:623–635. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Kaparakis M, Turnbull L, Carneiro L, Firth
S, Coleman HA, Parkington HC, Le Bourhis L, Karrar A, Viala J, Mak
J, et al: Bacterial membrane vesicles deliver peptidoglycan to NOD1
in epithelial cells. Cell Microbiol. 12:372–385. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Basak C, Pathak SK, Bhattacharyya A,
Pathak S, Basu J and Kundu M: The secreted peptidyl prolyl cis,
trans-isomerase HP0175 of Helicobacter pylori induces
apoptosis of gastric epithelial cells in a TLR4- and apoptosis
signal-regulating kinase 1-dependent manner. J Immunol.
174:5672–5680. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Yamaoka Y: Mechanisms of disease:
Helicobacter pylori virulence factors. Nat Rev Gastroenterol
Hepatol. 7:629–641. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Isomoto H, Moss J and Hirayama T:
Pleiotropic actions of Helicobacter pylori vacuolating
cytotoxin, VacA. Tohoku J Exp Med. 220:3–14. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Willhite DC, Cover TL and Blanke SR:
Cellular vacuolation and mitochondrial cytochrome c release are
independent outcomes of Helicobacter pylori vacuolating
cytotoxin activity that are each dependent on membrane channel
formation. J Biol Chem. 278:48204–48209. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Kroemer G, Marino G and Levine B:
Autophagy and the integrated stress response. Mol Cell. 40:280–293.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Kim IJ, Lee J, Oh SJ, Yoon MS, Jang SS,
Holland RL, Reno ML, Hamad MN, Maeda T, Chung HJ, et al:
Helicobacter pylori Infection modulates host cell metabolism
through VacA-dependent inhibition of mTORC1. Cell Host Microbe.
23:583–593.e8. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yahiro K, Satoh M, Nakano M, Hisatsune J,
Isomoto H, Sap J, Suzuki H, Nomura F, Noda M, Moss J and Hirayama
T: Low-density lipoprotein receptor-related protein-1 (LRP1)
mediates autophagy and apoptosis caused by Helicobacter
pylori VacA. J Biol Chem. 287:31104–31115. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Zhu P, Xue J, Zhang ZJ, Jia YP, Tong YN,
Han D, Li Q, Xiang Y, Mao XH and Tang B: Helicobacter pylori
VacA induces autophagic cell death in gastric epithelial cells via
the endoplasmic reticulum stress pathway. Cell Death Dis.
8:32072017. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Zhang K and Kaufman RJ: Identification and
characterization of endoplasmic reticulum stress-induced apoptosis
in vivo. Methods Enzymol. 442:395–419. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Terebiznik MR, Raju D, Vázquez CL,
Torbricki K, Kulkarni R, Blanke SR, Yoshimori T, Colombo MI and
Jones NL: Effect of Helicobacter pylori's vacuolating
cytotoxin on the autophagy pathway in gastric epithelial cells.
Autophagy. 5:370–379. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Jungblut PR, Bumann D, Haas G, Zimny-Arndt
U, Holland P, Lamer S, Siejak F, Aebischer A and Meyer TF:
Comparative proteome analysis of Helicobacter pylori. Mol
Microbiol. 36:710–725. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Odenbreit S, Puls J, Sedlmaier B, Gerland
E, Fischer W and Haas R: Translocation of Helicobacter
pylori CagA into gastric epithelial cells by type IV secretion.
Science. 287:1497–1500. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Kumar S and Dhiman M: Inflammasome
activation and regulation during Helicobacter pylori
pathogenesis. Microb Pathog. 125:468–474. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Ishikawa S, Ohta T and Hatakeyama M:
Stability of Helicobacter pylori CagA oncoprotein in human
gastric epithelial cells. FEBS Lett. 583:2414–2418. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Tsugawa H, Suzuki H, Saya H, Hatakeyama M,
Hirayama T, Hirata K, Nagano O, Matsuzaki J and Hibi T: Reactive
oxygen species-induced autophagic degradation of Helicobacter
pylori CagA is specifically suppressed in cancer stem-like
cells. Cell Host Microbe. 12:764–777. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Fan X, Long A, Goggins M, Fan X, Keeling
PW and Kelleher D: Expression of CD44 and its variants on gastric
epithelial cells of patients with Helicobacter pylori
colonisation. Gut. 38:507–512. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Xiao B, Liu Z, Li BS, Tang B, Li W, Guo G,
Shi Y, Wang F, Wu Y, Tong WD, et al: Induction of microRNA-155
during Helicobacter pylori infection and its negative
regulatory role in the inflammatory response. J Infect Dis.
200:916–925. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
71
|
Tang B, Li N, Gu J, Zhuang Y, Li Q, Wang
HG, Fang Y, Yu B, Zhang JY, Xie QH, et al: Compromised autophagy by
MIR30B benefits the intracellular survival of Helicobacter
pylori. Autophagy. 8:1045–1057. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Yang XJ, Si RH, Liang YH, Ma BQ, Jiang ZB,
Wang B and Gao P: Mir-30d increases intracellular survival of
Helicobacter pylori through inhibition of autophagy pathway.
World J Gastroenterol. 22:3978–3991. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Muhammad JS, Nanjo S, Ando T, Yamashita S,
Maekita T, Ushijima T, Tabuchi Y and Sugiyama T: Autophagy
impairment by Helicobacter pylori-induced methylation
silencing of MAP1LC3Av1 promotes gastric carcinogenesis. Int J
Cancer. 140:2272–2283. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Castano-Rodriguez N, Kaakoush NO, Goh KL,
Fock KM and Mitchell HM: Autophagy in Helicobacter pylori
infection and related gastric cancer. Helicobacter. 20:353–369.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Tanaka S, Nagashima H, Uotani T, Graham DY
and Yamaoka Y: Autophagy-related genes in Helicobacter
pylori infection. Helicobacter. 222017.doi:
10.1111/hel.12376.
|
|
76
|
Raju D, Hussey S and Jones NL: Crohn
disease ATG16L1 polymorphism increases susceptibility to infection
with Helicobacter pylori in humans. Autophagy. 8:1387–1388.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Suzuki M, Mimuro H, Kiga K, Fukumatsu M,
Ishijima N, Morikawa H, Nagai S, Koyasu S, Gilman RH, Kersulyte D,
et al: Helicobacter pylori CagA phosphorylation-independent
function in epithelial proliferation and inflammation. Cell Host
Microbe. 5:23–34. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Li N, Tang B, Jia YP, Zhu P, Zhuang Y,
Fang Y, Li Q, Wang K, Zhang WJ, Guo G, et al: Helicobacter
pylori CagA protein negatively regulates autophagy and promotes
inflammatory response via c-Met-PI3K/Akt-mTOR signaling pathway.
Front Cell Infect Microbiol. 7:4172017. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Luo MX, Wong SH, Chan MT, Yu L, Yu SS, Wu
F, Xiao Z, Wang X, Zhang L, Cheng AS, et al: Autophagy mediates
HBx-induced nuclear Factor-kappaB activation and release of IL-6,
IL-8, and CXCL2 in hepatocytes. J Cell Physiol. 230:2382–2389.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Li N, Wu X, Holzer RG, Lee JH, Todoric J,
Park EJ, Ogata H, Gukovskaya AS, Gukovsky I, Pizzo DP, et al: Loss
of acinar cell IKKa triggers spontaneous pancreatitis in mice. J
Clin Invest. 123:2231–2243. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Gukovskaya AS, Gukovsky I, Algul H and
Habtezion A: Autophagy, inflammation, and immune dysfunction in the
pathogenesis of pancreatitis. Gastroenterology. 153:1212–1226.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Yang JC, Shun CT, Chien CT and Wang TH:
Effective prevention and treatment of Helicobacter pylori
infection using a combination of catechins and sialic acid in AGS
cells and BALB/c mice. J Nutr. 138:2084–2090. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Chu YT, Wang YH, Wu JJ and Lei HY:
Invasion and multiplication of Helicobacter pylori in
gastric epithelial cells and implications for antibiotic
resistance. Infect Immun. 78:4157–4165. 2010. View Article : Google Scholar : PubMed/NCBI
|