Colorectal cancer (CRC) ranks third (13.5%) and
second (9.5%) among the incidence of malignancies worldwide in male
and female patients, respectively, and is a serious hazard to human
health (1). Previous studies have
demonstrated that the molecular pathogenesis of CRC is mostly
caused by genetic mutations (2,3).
Numerous studies over the past two decades have reported that
genetic mutations are associated with the prognosis and treatment
of CRC, and targeted therapies have been developed (4–7). The
progression of CRC is usually accompanied by the activation of the
KRAS and BRAF genes and the inhibition of the p53
tumour suppressor gene expression; mutations in these genes are
associated with changes in the number and structure of chromosomes
(8–10). However, >15% of sporadic CRCs
occur through completely different molecular pathogenesis. For
example, serrated precancerous lesions usually manifest as a result
of the methylation of the CpG locus and mutation of the gene
(11).
The prognosis of CRC is poor due to a lack of
effective diagnostic methods at an early stage. Therefore, an
effective solution can only be provided for subsequent diagnosis
and treatment by better understanding the gene expression of CRC
during its occurrence and development and identifying the genes
that may be involved in the occurrence and progression of
cancer.
With the rapid development of science and
technology, microarray technology has become increasingly accurate
and has been widely used to explore changes in animal and plant
gene expression (12–14). Microarray technology aids in the
discovery of changes in gene expression during cancer development
and progression (15–17). However, it can be difficult to obtain
reliable results with single microarray analysis. The present study
aimed to identify genetic changes in CRC in three mRNA microarray
datasets from the Gene Expression Omnibus (GEO) database by
obtaining differentially expressed genes (DEGs) between CRC and
normal tissues. Gene Ontology (GO) and Kyoto Encyclopedia of Genes
and Genomes (KEGG) were used for functional enrichment analysis,
and a protein-protein interaction (PPI) network was used to analyse
the associations between the DEGs. A total of 142 DEGs and 10 hub
genes were identified, which may be candidate biomarkers of
CRC.
The Database for Annotation, Visualization and
Integrated Discovery (DAVID; version 6.7; http://david.ncifcrf.gov) (19) is an online bioinformatics database
that integrates biological data and analysis tools and provides
gene annotation information and protein data. KEGG is a database
resource for understanding advanced functions and biological
systems from large-scale molecular data generated by
high-throughput experimental techniques (20). GO is a major bioinformatics tool for
annotating genes and analysing their biological processes (BPs),
molecular functions (MFs) and cellular components (CCs) (21). To analyse the function of the DEGs,
analysis was performed using DAVID. P<0.05 was considered to
indicate a statistically significant difference. The results based
on the top ten BPs, MFs, CCs and KEGG were visualized.
A PPI network was predicted using the Search Tool
for the Retrieval of Interacting Genes/Proteins (STRING; version
10.0; http://string-db.org) database (22). The PPI network of the DEGs was
constructed, and interactions with a combined score >0.4 were
considered statistically significant. Cytoscape (version 3.4.0) is
an open source bioinformatics platform for visualizing molecular
interaction networks (23).
Cytoscape plug-in Molecular Complex Detection (MCODE; version
1.4.2) provides topology-based clustering for a network (24). Cytoscape was used to draw a PPI
network, and MCODE was used to identify the most important modules
in the network. The selection criteria were as follows: MCODE score
>5, degree cut-off =2, node score cut-off =0.2, maximum depth
=100, and k score =2. Subsequently, KEGG and GO analysis of the
genes in this module was performed using DAVID.
The selection criterion for the hub genes was degree
of connectivity ≥10. The functions of the genes were identified
using the GeneCards online analysis tool (https://www.genecards.org). The BP analysis and
visualization of the hub genes were performed using the cBioPortal
(http://www.cbioportal.org) online
platform (25). The functions of the
hub genes were identified using GO and KEGG analysis. Hierarchical
clustering of the hub genes was constructed using the University of
California Santa Cruz Cancer Genomics Browser (http://genome-cancer.ucsc.edu) (26). The overall survival analysis was
performed using Kaplan-Meier curves in cBioPortal. The log-rank
test was used to for statistical analysis. The expression profiles
of DNA topoisomerase IIα (TOP2A) and phosphoribosylaminoimidazole
carboxylase (PAICS) were analysed using the Oncomine database
(http://www.oncomine.com). The expression levels
of genes in normal tissues and CRC tissues were also analysed using
the Oncomine database (27).
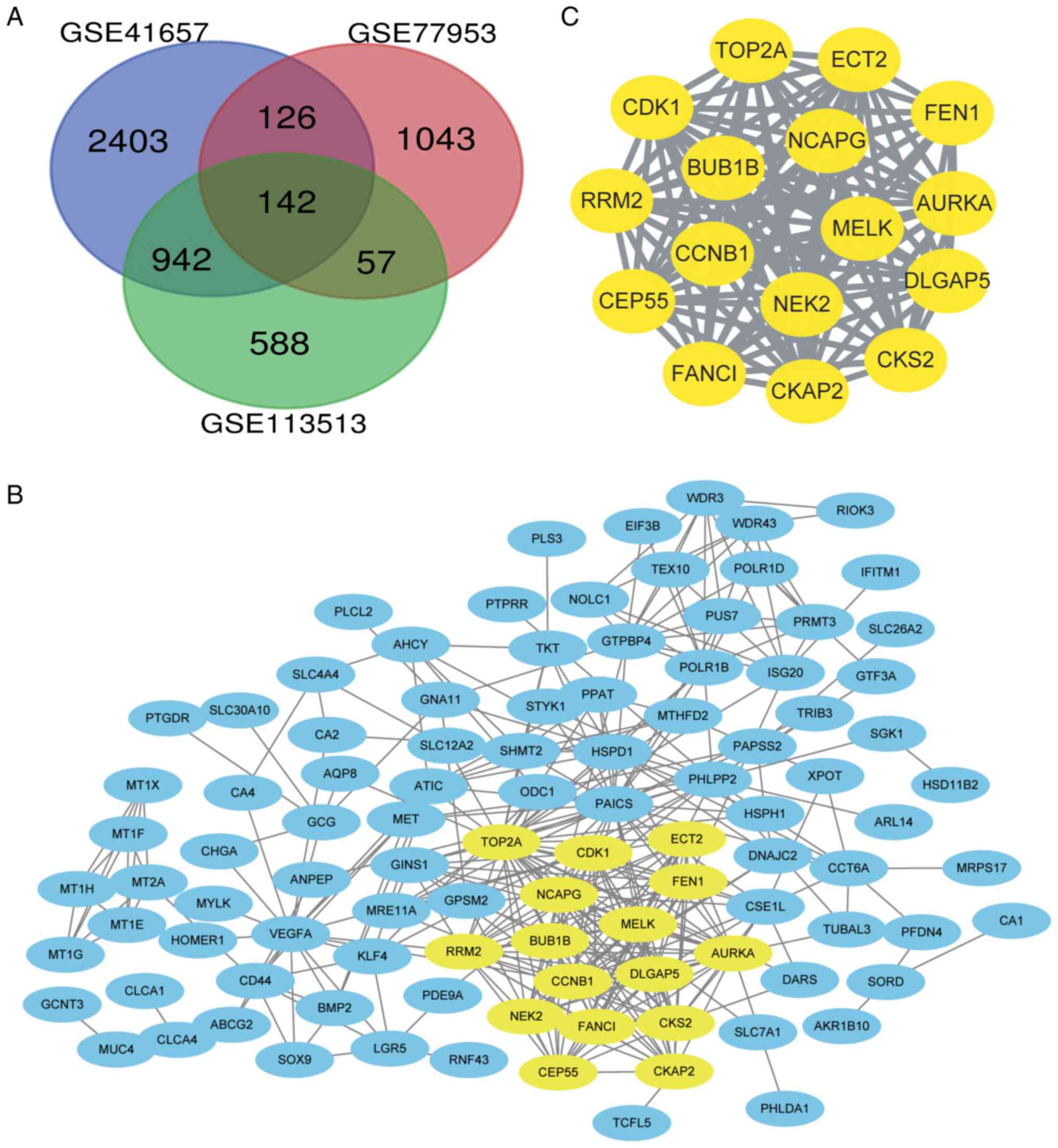
DEGs were identified by standardized microarray
results (GSE41657, GSE77953 and GSE113513). A total of 142 genes
overlapped in the three datasets, as demonstrated in the Venn
diagram (Fig. 1A).
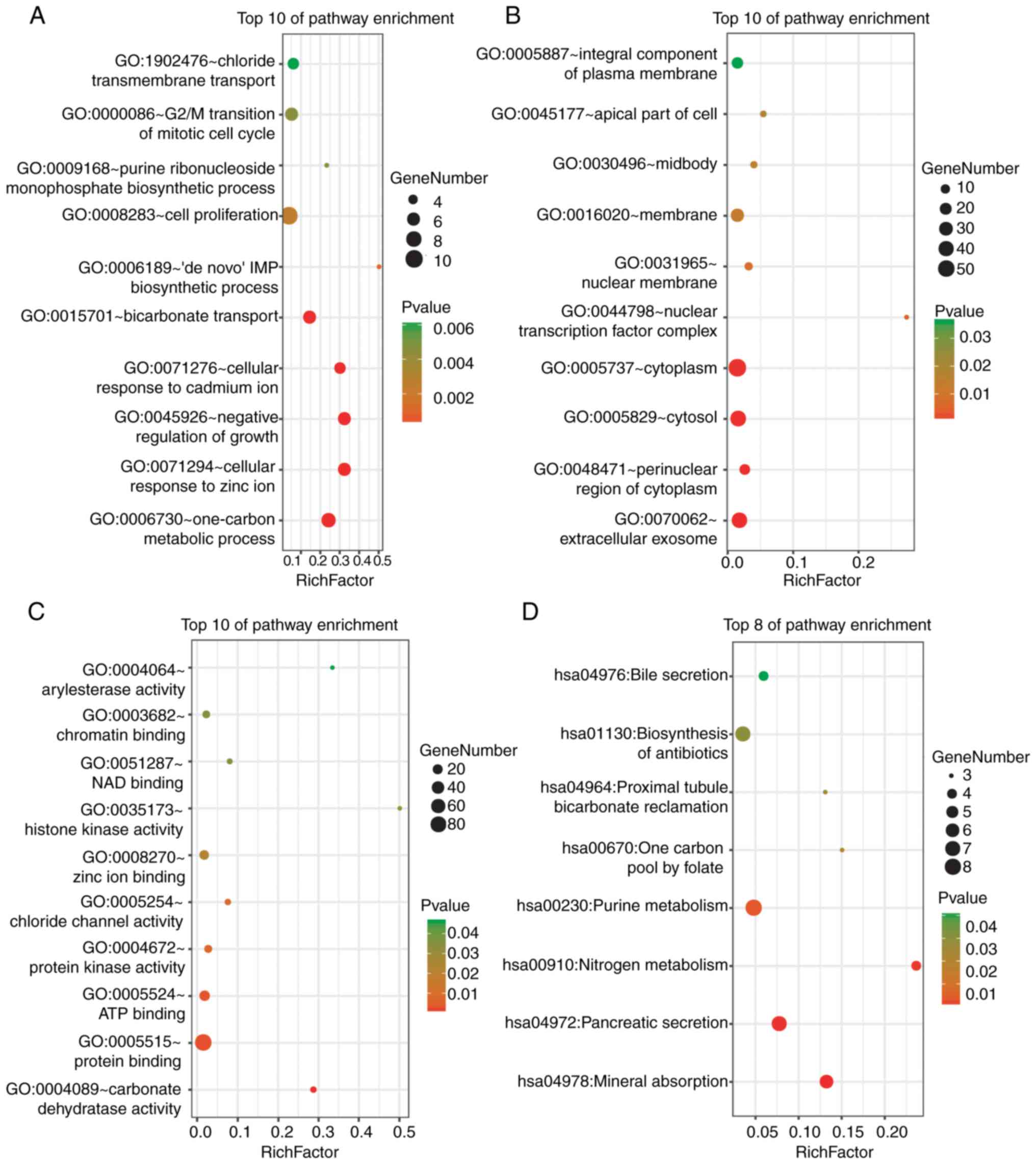
DAVID was used for the functional enrichment
analysis of DEGs. The results indicated that the DEGs were mainly
enriched in ‘cell proliferation’, ‘G2/M transition of mitotic cell
cycle’ and ‘one-carbon metabolism’ BPs (Fig. 2A); ‘cytoplasm’, ‘cytosol’ and
‘extracellular exosome’ cellular components (CCs) (Fig. 2B); and ‘protein binding’ and ‘ATP
binding’ molecular functions (MFs) (Fig.
2C). KEGG pathway analysis revealed strong enrichment in
‘biosynthesis of antibiotics’, ‘purine metabolism’, ‘pancreatic
secretion’ and ‘mineral absorption’ (Fig. 2D).
The functional results of the genes in the most
significant module indicated that these genes were mainly enriched
in processes associated with cell cycle progression, ATP binding,
nucleotide binding, the p53 signalling pathway and oocyte meiosis
(Table I).
Genes with a degree of connectivity ≥10 were
identified as the hub genes. The names, abbreviations and functions
of these genes are presented in Table
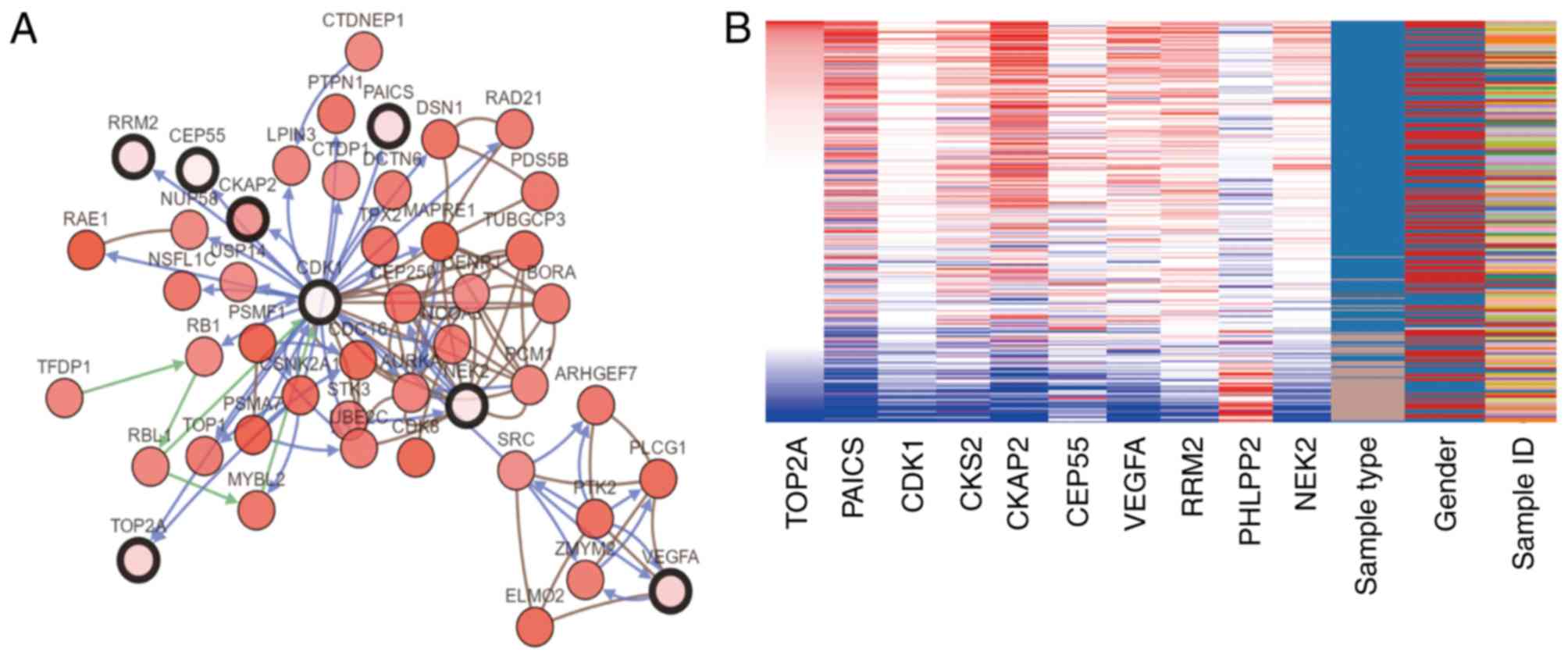
II. The cBioPortal online platform was used to construct a hub
gene network (Fig. 3A), and
functional enrichment analysis of DEGs was performed using DAVID
(Table III). The heat map
constructed using the UCSC Cancer Genomics Browser indicated that
the hub genes may be used to distinguish CRC tissue samples from
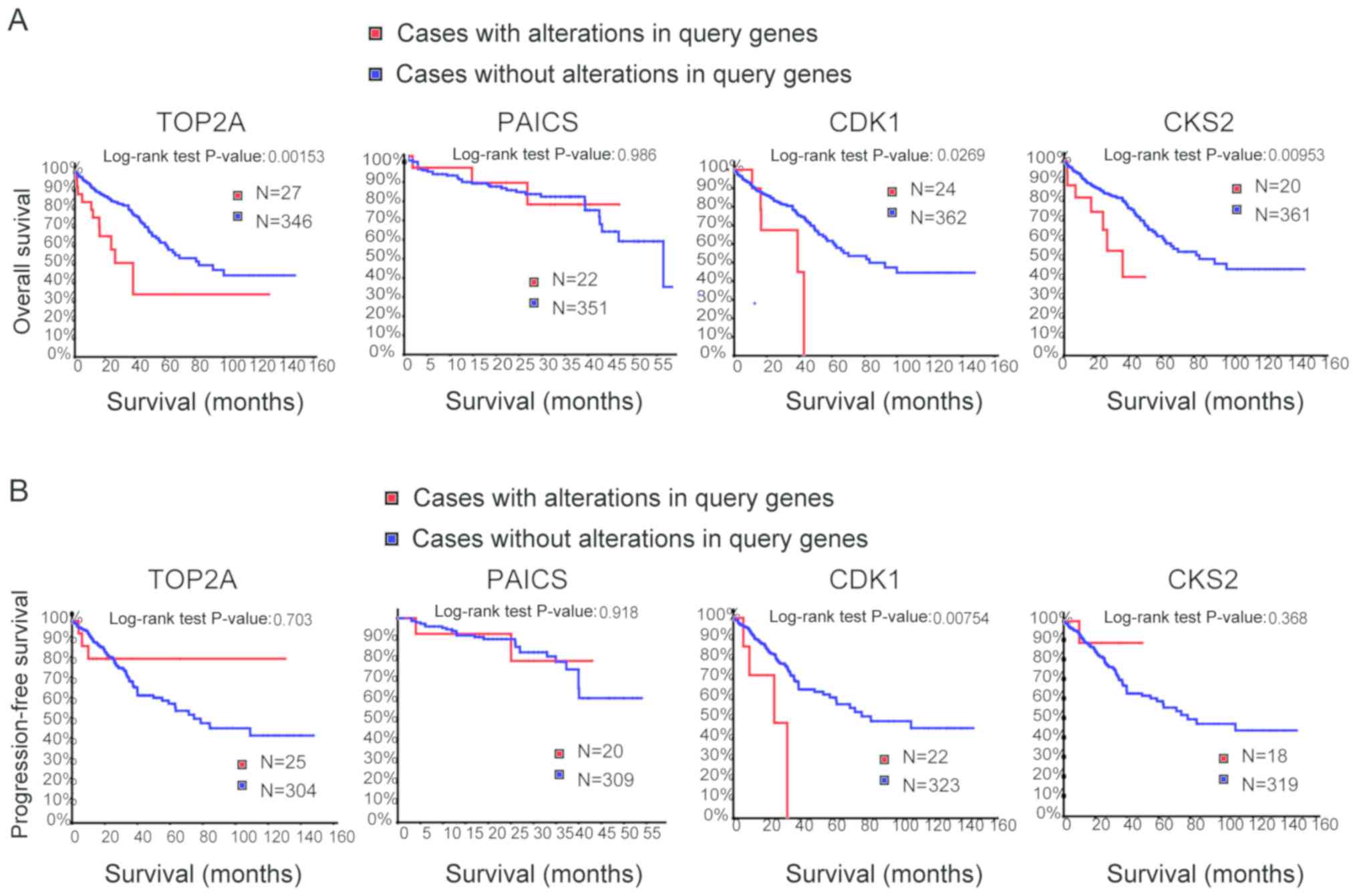
normal intestinal cancer tissue samples (Fig. 3B). Overall and progression-free
survival based on alterations in the hub genes was subsequently
analysed using Kaplan-Meier curves. Alterations in TOP2A,
cyclin-dependent kinase 1 (CDK1) and CDC28 protein kinase
regulatory subunit 2 (CKS2) in patients with CRC were associated
with a poor overall survival rate (Fig.
4A). However, patients with alterations in TOP2A and CKS2 did
not exhibit significant differences in progression-free survival
(Fig. 4B).
Among these genes, TOP2A exhibited a node degree of
34, and PAICS exhibited the highest degree of connectivity among
the hub genes (34 and 26, respectively). These two genes may serve
important roles in the occurrence or development of CRC. Based on
the survival analysis, alterations in PAICS exhibited no
statistically significant differences in overall and
progression-free survival in patients with CRC (overall survival
P=0.986; progression-free survival P=0.918; Fig. 4). Overall survival rates were
significantly different for patients with TOP2A alterations, but
progression-free survival was not significantly different (overall
survival P=0.002; progression-free survival P=0.703; Fig. 4). The expression profiles of TOP2A
and PAICS in human tissues were analysed using the Oncomine
database; TOP2A and PAICS mRNA was upregulated in various cancer
tissues compared with normal tissues (Fig. 5A and B). Further analysis revealed
significant increases in the expression of TOP2A and PAICS in CRC
(Fig. 5C and D).
CRC is a common malignant tumour of the digestive
tract, and its mortality rate (9.2%) ranks second among all types
of cancer (1). The main causes of
CRC include dietary and environmental factors, as well as genetic
mutations (3). The simultaneous
methylation of the CpG site and mutation of the BRAF gene are also
important factors in the development and progression of CRC
(11). Failure to detect CRC early
may be one of the reasons for poor prognosis in patients.
Therefore, there is an urgent need for efficient diagnostic and
therapeutic methods.
In the present study, DEGs between CRC and
non-cancerous tissues were obtained from three mRNA microarray
datasets. The functions of the DEGs were identified by GO and KEGG
enrichment analysis. The results demonstrated that the DEGs were
mainly enriched in the cell cycle, proliferation, mitotic cell
cycle and carbon metabolism. Previous studies have reported that
dysregulation of the cell cycle and the mitotic cell cycle serves
an important role in the development and progression of CRC
(28–31). In a study by Chamberlain et al
(32), functional B vitamin was used
to assess the association of carbon metabolism with CRC, and a
similar association between total B vitamin status and CRC risk was
identified. Ducker (33) et
al also revealed that the single-carbon metabolism of the
cytosol can support tumourigenesis. These findings are consistent
with the results of the present study. GO cluster and KEGG analysis
in the present study also revealed that the changes in the most
important module were mainly enriched in processes associated with
cell cycle progression, ATP and nucleotide binding and in KEGG
pathways associated with the cell cycle, progesterone-mediated
oocyte maturation and oocyte cell meiosis.
A total of 10 DEGs were selected as the hub genes.
Among them, TOP2A and PAICS were the top two nodes with the highest
degree of connectivity. TOP2A is involved in DNA replication,
transcription and chromosome segregation, and is essential for
tumorigenesis and cancer development (34). TOP2A has been demonstrated to be
associated with chemoresistance and tumour recurrence and has been
recognized as a target for anticancer drugs (35,36). In
addition, TOP2A is highly expressed in lung, colon and breast
cancer, involved in the inhibition of apoptosis, proliferation and
chemoresistance of CRC and may be considered an important biomarker
for the diagnosis, prognosis and treatment of tumours (37–39). In
the present study, the PPI network indicated that TOP2A interacted
directly with CDK1, ribonucleoside-diphosphate reductase subunit M2
(RRM2) and CKS2, indicating a key role for TOP2A in CRC. PAICS is
involved in purine nucleotide synthesis; a previous study has
demonstrated that high expression of PAICS in lung and prostate
cancer is associated with an altered metabolic state of cells,
apoptosis inhibition in cancer cells and enhanced cancer cell
invasion (40). In addition, PAICS
has been reported to promote tumourigenesis and progression in
breast and bladder cancer (41,42). The
results of the analysis in the Oncomine database in the present
study demonstrated that TOP2A and PAICS expression was
significantly upregulated in CRC compared with that in normal
tissues; however, to the best of our knowledge, there are currently
no studies focusing on the association between PAICS expression and
CRC. The association between TOP2A and PAICS and overall and
disease-free survival was also analysed; changes in TOP2A were
significantly associated with overall survival but were independent
of disease-free survival. Changes in PAICS exhibited a reduction in
overall and disease-free survival; however, these observations were
not statistically significant. These results may require further
research for verification.
CDK1 is a non-redundant cyclin-dependent kinase that
serves an important role in mitosis (43,44).
Perturbations in chromosomal stability and aspects of S phase and
G2/M control mediated by CDK2 and CDK1 are pivotal tumorigenic
events (45). A previous study has
demonstrated that CDK1 is required for the survival of cells
overexpressing MYC, and CDK1 has a therapeutic effect in the
treatment of human malignancies that overexpress MYC (46). The combination of CKS2 and CDK is
essential for promoting cancer cell metastasis in diseases such as
colon cancer (47,48). Vascular endothelial growth factor A
(VEGFA) is a class of cytokines called antigen growth factors that
stimulate the formation of new blood vessels; tumour angiogenesis
is primarily dependent on VEGFA-driven responses (49). Anti-VEGF-A treatment can reduce Treg
proliferation in CRC patients, and VEGFA inhibitors have been used
to treat CRC (50,51). In the present study revealed, the
expression levels of CDK11, CSK2 and VEGFA in CRC were analysed;
the results revealed high expression of these genes in CRC, which
was consistent with previous studies. In addition, high expression
levels of CDK1 and CKS2 were associated with poor survival.
In summary, the aim of the present study was to
identify DEGs that may be involved in the development or
progression of CRC. The results demonstrated that TOP2A and CDK1
may be involved in the survival prognosis of patients with CRC. The
present study also revealed that these genes may be biomarkers for
the diagnosis of CRC. However, the present study had limitations,
and other databases such as The Cancer Genome Atlas, as well as
in vivo and in vitro experiments may be needed to
clarify the biological functions of these genes in CRC.
Not applicable.
The present study was supported by the Fujian
Provincial Key Medical Specialist Construction Project (grant no.
2016-SLCZD), the Training Project of Young Talents in the Health
System of Fujian Province (grant no. 2016-ZQN-45) and the Fujian
Medical University Start up Fund for Scientific Research (grant no.
2017-XQ1061).
The datasets used and analysed during the present
study are available from the corresponding author on reasonable
request.
SQC and ZHC designed the study and obtained funding
YLL drafted the manuscript and supervised the study. YLL
participates in the design of the study, obtaining data and writing
manuscripts. YZ, YSL, JG and SYL collected and analysed the data.
All authors read and approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Van Cutsem E, Lenz HJ, Kohne CH, Heinemann
V, Tejpar S, Melezinek I, Beier F, Stroh C, Rougier P, van Krieken
JH and Ciardiello F: Fluorouracil, leucovorin, and irinotecan plus
cetuximab treatment and RAS mutations in colorectal cancer. J Clin
Oncol. 33:692–700. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tong K, Pellón-Cárdenas O, Sirihorachai
VR, Warder BN, Kothari OA, Perekatt AO, Fokas EE, Fullem RL, Zhou
A, Thackray JK, et al: Degree of tissue differentiation dictates
susceptibility to BRAF-driven colorectal cancer. Cell Rep.
21:3833–3845. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sanz-Garcia E, Argiles G, Elez E and
Tabernero J: BRAF mutant colorectal cancer: Prognosis, treatment,
and new perspectives. Ann Oncol. 28:2648–2657. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Corcoran RB, André T, Atreya CE, Schellens
JHM, Yoshino T, Bendell JC, Hollebecque A, McRee AJ, Siena S,
Middleton G, et al: Combined BRAF, EGFR, and MEK inhibition in
patients with BRAF(V600E)-mutant colorectal cancer. Cancer Discov.
8:428–443. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hamzehzadeh L, Khadangi F, Ghayoor
Karimiani E, Pasdar A and Kerachian MA: Common KRAS and NRAS gene
mutations in sporadic colorectal cancer in Northeastern Iranian
patients. Curr Probl Cancer. 42:572–581. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Reggiani Bonetti L, Barresi V, Bettelli S,
Caprera C, Manfredini S and Maiorana A: Analysis of KRAS, NRAS,
PIK3CA, and BRAF mutational profile in poorly differentiated
clusters of KRAS-mutated colon cancer. Hum Pathol. 62:91–98. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Misale S, Yaeger R, Hobor S, Scala E,
Janakiraman M, Liska D, Valtorta E, Schiavo R, Buscarino M,
Siravegna G, et al: Emergence of KRAS mutations and acquired
resistance to anti-EGFR therapy in colorectal cancer. Nature.
486:532–536. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cooks T, Pateras IS, Jenkins LM, Patel KM,
Robles AI, Morris J, Forshew T, Appella E, Gorgoulis VG and Harris
CC: Mutant p53 cancers reprogram macrophages to tumor supporting
macrophages via exosomal miR-1246. Nat Commun. 9:7712018.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fu X, Huang Y, Fan X, Deng Y, Liu H, Zou
H, Wu P, Chen Z, Huang J, Wang J, et al: Demographic trends and
KRAS/BRAFV600E mutations in colorectal cancer patients
of South China: A single-site report. Int J Cancer. 144:2109–2117.
2019.PubMed/NCBI
|
|
11
|
O'Brien MJ, Yang S, Mack C, Xu H, Huang
CS, Mulcahy E, Amorosino M and Farraye FA: Comparison of
microsatellite instability, CpG island methylation phenotype, BRAF
and KRAS status in serrated polyps and traditional adenomas
indicates separate pathways to distinct colorectal carcinoma end
points. Am J Surg Pathol. 30:1491–1501. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Saito M, Momma T and Kono K: Targeted
therapy according to next generation sequencing-based panel
sequencing. Fukushima J Med Sci. 64:9–14. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Deshiere A, Berthet N, Lecouturier F,
Gaudaire D and Hans A: Molecular characterization of Equine
Infectious Anemia Viruses using targeted sequence enrichment and
next generation sequencing. Virology. 537:121–129. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Meng H, Wang L, You H, Huang C and Li J:
Circular RNA expression profile of liver tissues in an EtOH-induced
mouse model of alcoholic hepatitis. Eur J Pharmacol.
862:1726422019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Harada K, Okamoto W, Mimaki S, Kawamoto Y,
Bando H, Yamashita R, Yuki S, Yoshino T, Komatsu Y, Ohtsu A, et al:
Comparative sequence analysis of patient-matched primary colorectal
cancer, metastatic, and recurrent metastatic tumors after adjuvant
FOLFOX chemotherapy. BMC Cancer. 19:2552019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hu Y, He C, Liu JP, Li NS, Peng C, Yang-Ou
YB, Yang XY, Lu NH and Zhu Y: Analysis of key genes and signaling
pathways involved in Helicobacter pylori-associated gastric cancer
based on the cancer genome atlas database and RNA sequencing data.
Helicobacter. 23:e125302018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nakagawa H and Fujita M: Whole genome
sequencing analysis for cancer genomics and precision medicine.
Cancer Sci. 109:513–522. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Barrett T, Wilhite SE, Ledoux P,
Evangelista C, Kim IF, Tomashevsky M, Marshall KA, Phillippy KH,
Sherman PM, Holko M, et al: NCBI GEO: Archive for functional
genomics data sets-update. Nucleic Acids Res. 41:D991–D995. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huang DW, Sherman BT, Tan Q, Collins JR,
Alvord WG, Roayaei J, Stephens R, Baseler MW, Lane HC and Lempicki
RA: The DAVID gene functional classification tool: A novel
biological module-centric algorithm to functionally analyze large
gene lists. Genome Biol. 8:R1832007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kanehisa M, Furumichi M, Tanabe M, Sato Y
and Morishima K: KEGG: New perspectives on genomes, pathways,
diseases and drugs. Nucleic Acids Res. 45:D353–D361. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The gene
ontology consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Franceschini A, Szklarczyk D, Frankild S,
Kuhn M, Simonovic M, Roth A, Lin J, Minguez P, Bork P, von Mering C
and Jensen LJ: STRING v9.1: Protein-protein interaction networks,
with increased coverage and integration. Nucleic Acids Res.
41:D808–D815. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Smoot ME, Ono K, Ruscheinski J, Wang PL
and Ideker T: Cytoscape 2.8: New features for data integration and
network visualization. Bioinformatics. 27:431–432. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bandettini WP, Kellman P, Mancini C,
Booker OJ, Vasu S, Leung SW, Wilson JR, Shanbhag SM, Chen MY and
Arai AE: MultiContrast Delayed Enhancement (MCODE) improves
detection of subendocardial myocardial infarction by late
gadolinium enhancement cardiovascular magnetic resonance: A
clinical validation study. J Cardiovasc Magn Reson. 14:832012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cerami E, Gao J, Dogrusoz U, Gross BE,
Sumer SO, Aksoy BA, Jacobsen A, Byrne CJ, Heuer ML, Larsson E, et
al: The cBio cancer genomics portal: An open platform for exploring
multidimensional cancer genomics data. Cancer Discov. 2:401–404.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Haeussler M, Zweig AS, Tyner C, Speir ML,
Rosenbloom KR, Raney BJ, Lee CM, Lee BT, Hinrichs AS, Gonzalez JN,
et al: The UCSC genome browser database: 2019 update. Nucleic Acids
Res. 47:D853–D858. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rhodes DR, Yu J, Shanker K, Deshpande N,
Varambally R, Ghosh D, Barrette T, Pandey A and Chinnaiyan AM:
ONCOMINE: A cancer microarray database and integrated data-mining
platform. Neoplasia. 6:1–6. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang ZZ, Yang J, Jiang BH, Di JB, Gao P,
Peng L and Su XQ: KIF14 promotes cell proliferation via activation
of Akt and is directly targeted by miR-200c in colorectal cancer.
Int J Oncol. 53:1939–1952. 2018.PubMed/NCBI
|
|
29
|
Wu J, Yi J, Wu Y, Chen X, Zeng J, Wu J and
Peng W: 3, 3′-dimethylquercetin inhibits the proliferation of human
colon cancer RKO cells through Inducing G2/M cell cycle arrest and
apoptosis. Anticancer Agents Med Chem. 19:402–409. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cheng J, Dwyer M, Okolotowicz KJ, Mercola
M and Cashman JR: A novel inhibitor targets both wnt signaling and
ATM/p53 in colorectal cancer. Cancer Res. 78:5072–5083. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li J, Liu YY, Yang XF, Shen DF, Sun HZ,
Huang KQ and Zheng HC: Effects and mechanism of STAT3 silencing on
the growth and apoptosis of colorectal cancer cells. Oncol Lett.
16:5575–5582. 2018.PubMed/NCBI
|
|
32
|
Chamberlain JA, Dugué PA, Bassett JK,
Hodge AM, Brinkman MT, Joo JE, Jung CH, Makalic E, Schmidt DF,
Hopper JL, et al: Dietary intake of one-carbon metabolism nutrients
and DNA methylation in peripheral blood. Am J Clin Nutr.
108:611–621. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ducker GS, Chen L, Morscher RJ,
Ghergurovich JM, Esposito M, Teng X, Kang Y and Rabinowitz JD:
Reversal of cytosolic one-carbon flux compensates for loss of the
mitochondrial folate pathway. Cell Metab. 23:1140–1153. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nitiss JL: DNA topoisomerase II and its
growing repertoire of biological functions. Nat Rev Cancer.
9:327–337. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pommier Y, Leo E, Zhang H and Marchand C:
DNA topoisomerases and their poisoning by anticancer and
antibacterial drugs. Chem Biol. 17:421–433. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Deweese JE and Osheroff N: The DNA
cleavage reaction of topoisomerase II: Wolf in sheep's clothing.
Nucleic Acids Res. 37:738–748. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
McLeod HL, Douglas F, Oates M, Symonds RP,
Prakash D, van der Zee AG, Kaye SB, Brown R and Keith WN:
Topoisomerase I and II activity in human breast, cervix, lung and
colon cancer. Int J Cancer. 59:607–611. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shibao K, Takano H, Nakayama Y, Okazaki K,
Nagata N, Izumi H, Uchiumi T, Kuwano M, Kohno K and Itoh H:
Enhanced coexpression of YB-1 and DNA topoisomerase II alpha genes
in human colorectal carcinomas. Int J Cancer. 83:732–737. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Coss A, Tosetto M, Fox EJ, Sapetto-Rebow
B, Gorman S, Kennedy BN, Lloyd AT, Hyland JM, O'Donoghue DP,
Sheahan K, et al: Increased topoisomerase IIalpha expression in
colorectal cancer is associated with advanced disease and
chemotherapeutic resistance via inhibition of apoptosis. Cancer
Lett. 276:228–238. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Goswami MT, Chen G, Chakravarthi BV, Pathi
SS, Anand SK, Carskadon SL, Giordano TJ, Chinnaiyan AM, Thomas DG,
Palanisamy N, et al: Role and regulation of coordinately expressed
de novo purine biosynthetic enzymes PPAT and PAICS in lung cancer.
Oncotarget. 6:23445–23461. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Meng M, Chen Y, Jia J, Li L and Yang S:
Knockdown of PAICS inhibits malignant proliferation of human breast
cancer cell lines. Biol Res. 51:242018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chakravarthi BVSK, Rodriguez Pena MDC,
Agarwal S, Chandrashekar DS, Hodigere Balasubramanya SA, Jabboure
FJ, Matoso A, Bivalacqua TJ, Rezaei K, Chaux A, et al: A role for
de novo purine metabolic enzyme PAICS in bladder cancer
progression. Neoplasia. 20:894–904. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Santamaría D, Barrière C, Cerqueira A,
Hunt S, Tardy C, Newton K, Cáceres JF, Dubus P, Malumbres M and
Barbacid M: Cdk1 is sufficient to drive the mammalian cell cycle.
Nature. 448:811–815. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Brown NR, Korolchuk S, Martin MP, Stanley
WA, Moukhametzianov R, Noble MEM and Endicott JA: CDK1 structures
reveal conserved and unique features of the essential cell cycle
CDK. Nat Commun. 6:67692015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Asghar U, Witkiewicz AK, Turner NC and
Knudsen ES: The history and future of targeting cyclin-dependent
kinases in cancer therapy. Nat Rev Drug Discov. 14:130–146. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Goga A, Yang D, Tward AD, Morgan DO and
Bishop JM: Inhibition of CDK1 as a potential therapy for tumors
over-expressing MYC. Nat Med. 13:820–827. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Martinsson-Ahlzén HS, Liberal V,
Grünenfelder B, Chaves SR, Spruck CH and Reed SI: Cyclin-dependent
kinase-associated proteins Cks1 and Cks2 are essential during early
embryogenesis and for cell cycle progression in somatic cells. Mol
Cell Biol. 28:5698–5709. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Li M, Lin YM, Hasegawa S, Shimokawa T,
Murata K, Kameyama M, Ishikawa O, Katagiri T, Tsunoda T, Nakamura Y
and Furukawa Y: Genes associated with liver metastasis of colon
cancer, identified by genome-wide cDNA microarray. Int J Oncol.
24:305–312. 2004.PubMed/NCBI
|
|
49
|
Claesson-Welsh L and Welsh M: VEGFA and
tumour angiogenesis. J Intern Med. 273:114–127. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ferrara N and Adamis AP: Ten years of
anti-vascular endothelial growth factor therapy. Nat Rev Drug
Discov. 15:385–403. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Terme M, Pernot S, Marcheteau E, Sandoval
F, Benhamouda N, Colussi O, Dubreuil O, Carpentier AF, Tartour E
and Taieb J: VEGFA-VEGFR pathway blockade inhibits tumor-induced
regulatory T-cell proliferation in colorectal cancer. Cancer Res.
73:539–549. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Seki A and Fang G: CKAP2 is a
spindle-associated protein degraded by APC/C-Cdh1 during mitotic
exit. J Biol Chem. 282:15103–15113. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Tsuchihara K, Lapin V, Bakal C, Okada H,
Brown L, Hirota-Tsuchihara M, Zaugg K, Ho A, Itie-Youten A,
Harris-Brandts M, et al: Ckap2 regulates aneuploidy, cell cycling,
and cell death in a p53-dependent manner. Cancer Res. 65:6685–6691.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Weinberger P, Ponny SR, Xu H, Bai S,
Smallridge R, Copland J and Sharma A: Cell cycle M-phase genes are
highly upregulated in anaplastic thyroid carcinoma. Thyroid.
27:236–252. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wang Y, Jin T, Dai X and Xu J:
Lentivirus-mediated knockdown of CEP55 suppresses cell
proliferation of breast cancer cells. Biosci Trends. 10:67–73.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Jiang W, Wang Z, Chen G and Jia Y:
Prognostic significance of centrosomal protein 55 in stage I
pulmonary adenocarcinoma after radical resection. Thorac Cancer.
7:316–322. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
D'Angiolella V, Donato V, Forrester FM,
Jeong YT, Pellacani C, Kudo Y, Saraf A, Florens L, Washburn MP and
Pagano M: Cyclin F-mediated degradation of ribonucleotide reductase
M2 controls genome integrity and DNA repair. Cell. 149:1023–1034.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Brognard J and Newton AC: PHLiPPing the
switch on Akt and protein kinase C signaling. Trends Endocrinol
Metab. 19:223–230. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Brognard J, Sierecki E, Gao T and Newton
AC: PHLPP and a second isoform, PHLPP2, differentially attenuate
the amplitude of Akt signaling by regulating distinct Akt isoforms.
Mol Cell. 25:917–931. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Liao WT, Li TT, Wang ZG, Wang SY, He MR,
Ye YP, Qi L, Cui YM, Wu P, Jiao HL, et al: microRNA-224 promotes
cell proliferation and tumor growth in human colorectal cancer by
repressing PHLPP1 and PHLPP2. Clin Cancer Res. 19:4662–4672. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Cai J, Fang L, Huang Y, Li R, Yuan J, Yang
Y, Zhu X, Chen B, Wu J and Li M: miR-205 targets PTEN and PHLPP2 to
augment AKT signaling and drive malignant phenotypes in non-small
cell lung cancer. Cancer Res. 73:5402–5415. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Santarpia L, Qi Y, Stemke-Hale K, Wang B,
Young EJ, Booser DJ, Holmes FA, O'Shaughnessy J, Hellerstedt B,
Pippen J, et al: Mutation profiling identifies numerous rare drug
targets and distinct mutation patterns in different clinical
subtypes of breast cancers. Breast Cancer Res Treat. 134:333–343.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Hu CM, Zhu J, Guo XE, Chen W, Qiu XL, Ngo
B, Chien R, Wang YV, Tsai CY, Wu G, et al: Novel small molecules
disrupting Hec1/Nek2 interaction ablate tumor progression by
triggering Nek2 degradation through a death-trap mechanism.
Oncogene. 34:1220–1230. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Hayward DG, Clarke RB, Faragher AJ, Pillai
MR, Hagan IM and Fry AM: The centrosomal kinase Nek2 displays
elevated levels of protein expression in human breast cancer.
Cancer Res. 64:7370–7376. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Hawkins SM, Loomans HA, Wan YW,
Ghosh-Choudhury T, Coffey D, Xiao W, Liu Z, Sangi-Haghpeykar H and
Anderson ML: Expression and functional pathway analysis of nuclear
receptor NR2F2 in ovarian cancer. J Clin Endocrinol Metab.
98:E1152–E1162. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Neal CP, Fry AM, Moreman C, McGregor A,
Garcea G, Berry DP and Manson MM: Overexpression of the Nek2 kinase
in colorectal cancer correlates with beta-catenin relocalization
and shortened cancer-specific survival. J Surg Oncol. 110:828–838.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Lu L, Zhai X and Yuan R: Clinical
significance and prognostic value of Nek2 protein expression in
colon cancer. Int J Clin Exp Pathol. 8:15467–15473. 2015.PubMed/NCBI
|
|
68
|
Xu H, Zeng L, Guan Y, Feng X, Zhu Y, Lu Y,
Shi C, Chen S, Xia J, Guo J, et al: High NEK2 confers to poor
prognosis and contributes to cisplatin-based chemotherapy
resistance in nasopharyngeal carcinoma. J Cell Biochem.
120:3547–3558. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Zhang Y, Wang W, Wang Y, Huang X, Zhang Z,
Chen B, Xie W, Li S, Shen S and Peng B: NEK2 promotes
hepatocellular carcinoma migration and invasion through modulation
of the epithelial-mesenchymal transition. Oncol Rep. 39:1023–1033.
2018.PubMed/NCBI
|