Introduction
Adrenocortical carcinoma (ACC) is a rare endocrine
malignancy, with an incidence rate of 1.5 to 2 cases per million
people per year worldwide (1).
Albeit rare, ACC is often aggressive and lethal, with 10–20% of
patients surviving 5 years after diagnosis (2). To date, the main curative therapy for
ACC is surgical resection of the primary tumor. However, most
patients with an advanced stage of the disease have a median
survival time of <12 months, even after complete tumor resection
(3). Even with ostensibly complete
resections, the rates of local recurrence have typically ranged
from at least 19 to 34% in those patients with no residual disease
after surgery (4,5). Mitotane remains the only drug approved
for ACC treatment by the U.S. Food and Drug Administration and
European Medicine Executive Agency (6). However, mitotane is an adjuvant therapy
for patients with low to moderate risk for ACC recurrence, and its
efficacy is ultimately limited by its high lipophilicity, poor
pharmacokinetic properties, and dose-limiting toxicities (7). In addition, long-term radiotherapy may
be used in patients at an increased risk of local recurrence
(8). The high degree of malignancy,
strong invasiveness, poor traditional treatment efficacy and poor
prognosis of ACC have led to the exploration of targeted
therapy.
A recent study has demonstrated that endoplasmic
reticulum (ER) stress may play an important role in the development
of cancer (9). Alterations in the
homeostasis and appropriate functioning of the ER initiates a
cascade of signaling events known as the ER stress response or
unfolded protein response (UPR). When ER stress occurs, cancer
cells in colorectal, breast and cervical cancer, adapt the UPR to
alleviate the ER stress condition as a survival approach for
progression (10,11). Adaptation of cancer cells to adverse
conditions largely relies on the ability of a cell to perturb ER
stress-associated regulatory networks and prevent ER stress-induced
cell death (12,13). Autophagy is a highly conserved system
that delivers misfolded proteins and damaged organelles to the
lysosome for degradation, maintaining cell homeostasis (14). Li et al (15) showed that the ER stress-responsive
protein kinase R-like ER kinase (PERK)-eukaryotic initiation
factor-2α (eIF2α)-activating transcription factor (ATF) 4 pathway
contributes to ER stress-induced autophagy. Persistent ER stress
often results in the stimulation of autophagic activities (16).
Tauroursodeoxycholic acid (TUDCA) is a chemical
chaperone that stabilizes protein conformation and improves the
folding capacity of the ER (17).
Yang et al (18) showed that
TUDCA could downregulate ER stress in a dose-dependent manner using
human hepatocellular carcinoma cells. Guo et al (19) found that TUDCA reversed abnormal
autophagy and reduced ER stress in the liver of obese mice.
Therefore, TUDCA is a promising regulator for mediating ER stress,
which significantly relieves ER stress and inhibits cell apoptosis
in the aforementioned cells.
The present study aimed to identify whether ER
stress and autophagy are involved in the occurrence of ACC by TUDCA
interventions, providing a theoretical basis for the treatment of
ACC.
Materials and methods
Cell culture
The SW-13 cell line was obtained from Shanghai
Institutes for Biological Sciences, Chinese Academy of Sciences
(cat. no. TCHu221). The NCI-H295R cell line was obtained from the
American Type Culture Collection (cat. no. ATCC®
CRL-2128). Cells were grown in minimum essential medium
supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.) and 1% penicillin-streptomycin solution (Beijing
Solarbio Science and Technology Co., Ltd.). Cells were cultured at
37°C in a humidified atmosphere with 5% CO2 and 95%
humidity in an incubator. TUDCA was purchased from EMD Millipore.
SW-13 cells were treated with 0, 100, 200, 300 or 400 µM TUDCA, and
NCI-H295R cells were treated with 0, 100, 200, 400 or 600 µM
TUDCA.
Cell proliferation assay
SW-13 and NCI-H295R cells were seeded in 96-well
plates at a density of 1×103 cells/well and allowed to
attach for 24 h. Then, the cells were treated with different
concentrations of TUDCA as aforementioned. The cells were incubated
at 37°C for 12, 24, 48 and 72 h. Then, cell proliferation was
assessed using a Cell Counting Kit 8 (CCK8) assay (Dojindo
Molecular Technologies, Inc.) according to the manufacturer's
instructions. Finally, the optical density at 450 nm was detected
and cell proliferation calculated. Each set of experiments was
performed in triplicate.
Cell migration assay
After SW-13 and NCI-H295R cells were resuspended
with trypsin (Gibco; Thermo Fisher Scientific, Inc.),
5×105 cells/well were seeded in 6-well plates and
incubated in 10% serum-containing minimal essential medium (Gibco;
Thermo Fisher Scientific, Inc.) at 37°C for 24 h. When the cells
reached 100% confluence, scratches on the cells were made
perpendicular to the well plate with a small tip. The well plates
were washed once with PBS to remove the dislodged cells. Then,
SW-13 and NCI-H295R cells were treated with different
concentrations of TUDCA as aforementioned. The cells were cultured
in serum-free minimal essential medium at 37°C. Migration was
visualized at 0, 6, 12, and 24 h with an inverted light microscope
(TE2000; Nikon Corporation). Migration distances were measured
using ImageJ software version 1.8.0 (National Institutes of
Health).
Transwell invasion assays
SW-13 and NCI-H295R cells were treated with
aforementioned concentrations of TUDCA. The cells were cultured in
serum-free medium for 24 h at 37°C. Then, the upper chambers were
precoated with Matrigel matrix (Corning, Inc.), and cells were
added to the upper chambers at a concentration of 1×105
cells per well. Next, 500 µl serum-containing medium was added to
the lower chambers. After 24 h of incubation, the cells on the
upper side of the filters were removed with a cotton swab, and the
filters were washed with PBS. The cells were fixed in methanol for
30 min at room temperature, and nuclei were stained with 0.1%
crystal violet for 15 min at room temperature. Images were captured
under a light microscope, and cell counts were measured using
ImageJ software version 1.8.0 (National Institutes of Health).
Reverse transcription-quantitative PCR
(RT-qPCR) analysis
Total RNA was extracted from SW-13 cells cultured in
different concentrations of TUDCA (0, 100, 200, 300, or 400 µM for
48 h) using RNAiso Plus reagent (Takara Biotechnology Co., Ltd.).
Following RNA extraction, cDNA was synthesized using a Takara RT
kit (Takara Biotechnology Co., Ltd.) at 37°C for 15 min, 85°C for 5
sec and kept at 4°C until further experimentation. qPCR
amplification was conducted with SYBR-Green reagent (Takara
Biotechnology Co., Ltd.) using an ABI 7500 sequencer (Applied
Biosystems; Thermo Fisher Scientific, Inc.) and the following
conditions were used: Initial denaturation at 95°C for 30 sec, then
40 cycles of 95°C for 5 sec and 60°C for 34 sec. The experimental
steps were carried out according to the manufacturer's
instructions. The primers were designed and synthesized by Sangon
Biotech Co., Ltd. (Table I).
Cq values were generated using the default analysis
settings. ΔCq was calculated using the following
formula: (Cq gene of interest)-(CT GAPDH).
∆∆Cq was calculated using the following formula:
(ΔCq treated sample)-(Cq control sample).
Relative expression was calculated using the 2−ΔΔCq
method as previously described (20).
 | Table I.Primer sequence for reverse
transcription-quantitative PCR. |
Table I.
Primer sequence for reverse
transcription-quantitative PCR.
| Gene name | Primer sequence
(5′-3′) |
|---|
| GAPDH | F:
CAGGAGGCATTGCTGATGAT |
|
| R:
GAAGGCTGGGGCTCATTT |
| GRP78 | F:
CAGTTGTTACTGTACCAGCCTA |
|
| R:
CATTTAGGCCAGCAATAGTTCC |
| PERK | F:
CCAGTTTTGTACTCCAATTGCA |
|
| R:
CAGATACAGCTGGCCTCTATAC |
| CHOP | F:
GAGAATGAAAGGAAAGTGGCAC |
|
| R:
ATTCACCATTCGGTCAATCAGA |
| JNK | F:
ACACCACAGAAATCCCTAGAAG |
|
| R:
CACAGCATCTGATAGAGAAGGT |
| ATF6 | F:
CTGATGGCTGTTCAATACACAG |
|
| R:
GATCCCTTCGAAATGACACAAC |
| LC3-II/I | F:
AGCCCGTTTCTTTCATCATAACATC |
|
| R:
AAGATCTAAGCCTGTGCCATTTAC |
| Bax | F:
CGAACTGGACAGTAACATGGAG |
|
| R:
CAGTTTGCTGGCAAAGTAGAAA |
| Bcl-2 | F:
GACTTCGCCGAGATGTCCAG |
|
| R:
GAACTCAAAGAAGGCCACAATC |
Western blot analysis
SW-13 cells were treated with aforementioned
concentrations of TUDCA. After 48 h, SW-13 cells were lysed with
RIPA lysis buffer containing 1 mM PMSF (Beijing Solarbio Science
and Technology Co., Ltd.) for 30 min then the cells were
centrifuged at 14,000 × g for 10 min at 4°C, and the supernatant
was collected. Subsequently, the protein concentration was
determined using a BCA protein analysis kit (Beyotime Institute of
Biotechnology). A total of 50 µg of total protein was separated
using 10% SDS-PAGE and transferred to PVDF membranes
(Sigma-Aldrich; Merck KGaA). The membranes were blocked using 5%
skimmed milk at room temperature for 1 h and then incubated with
primary antibodies at 4°C overnight. Next, the membranes were
washed three times with TBS-Tween 20 and incubated with secondary
horseradish peroxidase-conjugated antibody (dilution 1:10,000; cat.
no. L3012-2; Signalway Antibody LLC) for 1 h at room temperature.
The protein bands were imaged using a densitometric scanner
(Bio-Rad Laboratories, Inc.) and analyzed with ImageJ software
version 1.8.0 (National Institutes of Health). Antibodies against
PERK (dilution 1:1,000; cat. no. 3192), JNK (dilution 1:1,000; cat.
no. 9252), ATF6 (dilution 1:1,000; cat. no. 65880), C/EBP
homologous protein (CHOP; dilution 1:1,000; cat. no. 2895),
microtubule-associated protein light chain 3 (LC3)-II/I (dilution
1:1,000; cat. no. 2775), Bax (dilution 1:1,000; cat. no. 2772) and
Bcl-2 (dilution 1:1,000; cat. no. 3498) were purchased from Cell
Signaling Technology, Inc. Antibodies against glucose-regulated
protein 78 (GRP78; dilution 1:1,000; cat. no. 33395) and GAPDH
(dilution 1:5,000; cat. no. 21612) were purchased from Signalway
Antibody LLC.
Statistical analysis
All experiments were repeated three times, and the
data were analyzed using SPSS version 17.0 software (SPSS, Inc.).
Graphical representation of data was prepared with GraphPad Prism
version 5.0 software (GraphPad Software, Inc.). The data are
expressed as the mean ± standard deviation. Differences in the data
among the groups were analyzed using one-way ANOVA combined with
Bonferroni's post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
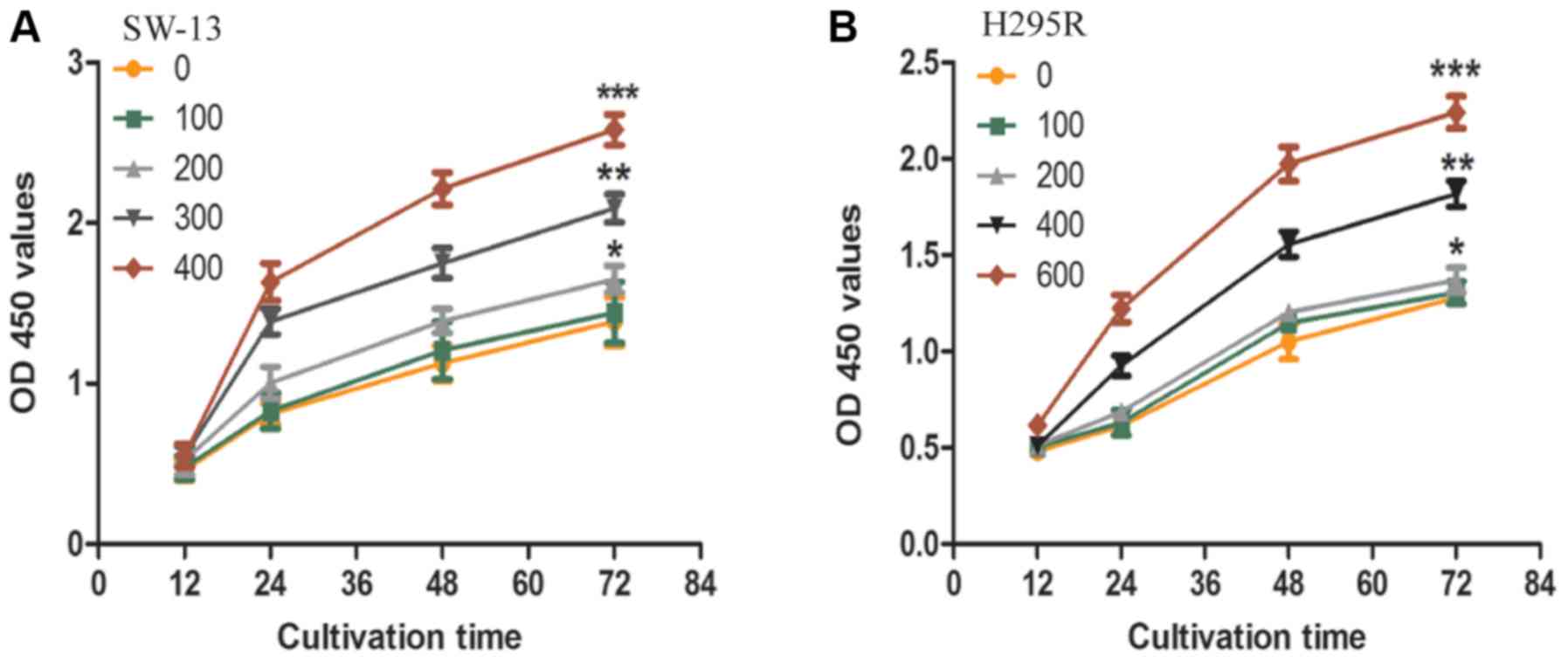
TUDCA promotes the proliferation,
migration, and invasiveness of ACC cells
The effects of TUDCA on SW-13 and NCI-H295R cell
proliferation, motility, and invasiveness were investigated. The
proliferation of SW-13 and NCI-H295R cells after treatment with
different concentrations of TUDCA was measured using a CCK8 assay.
The CCK8 assay revealed that TUDCA promoted SW-13 and NCI-H295R
cell proliferation. TUDCA at a concentration of 400 µM
significantly promoted the proliferation of SW-13 cells, while 600
µM TUDCA significantly promoted NCI-H295R cell proliferation
(Fig. 1A and B).
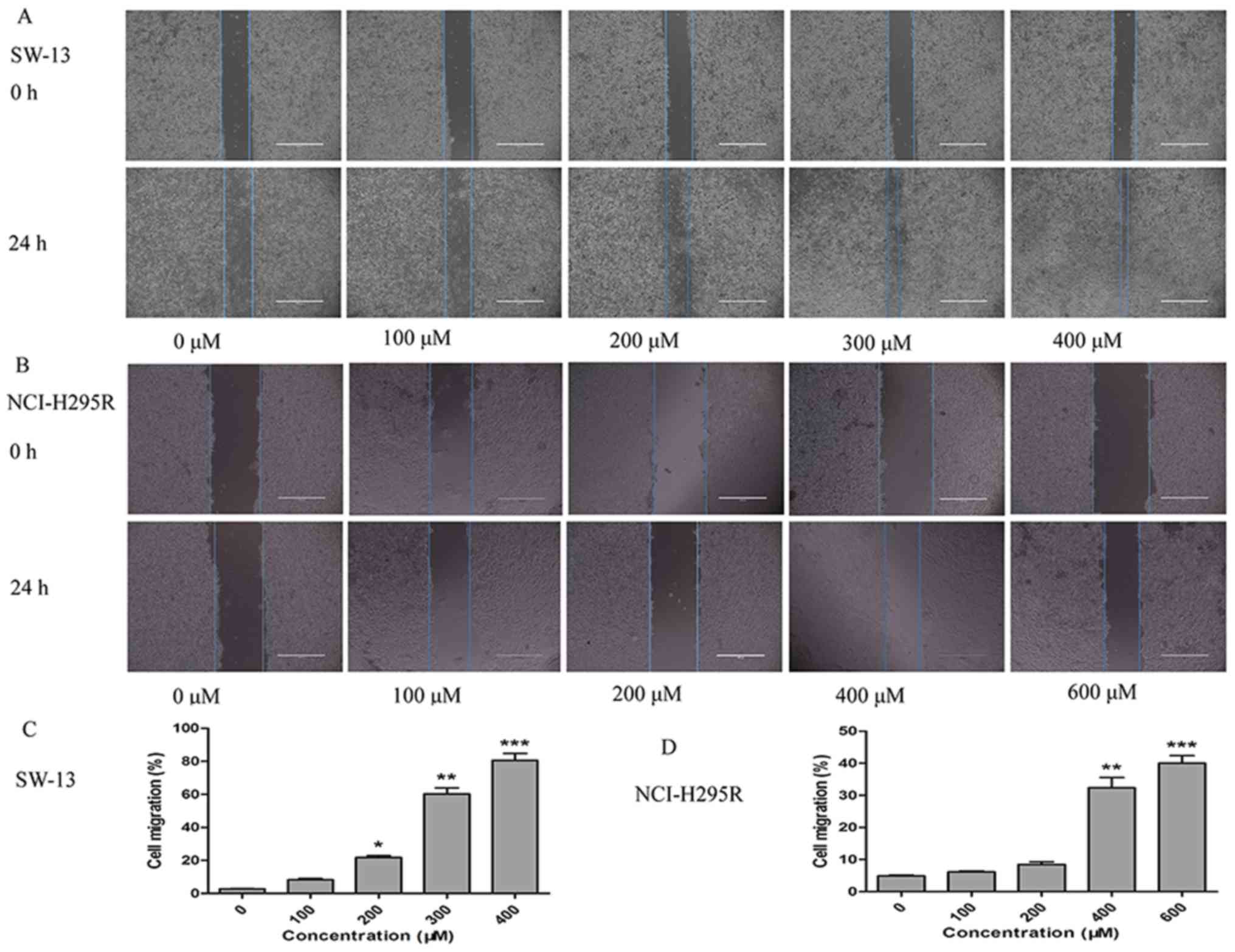
Since the migration and invasion of cancer cells are
crucial factors responsible for cancer progression (21), the effect of TUDCA on SW-13 and
NCI-H295R cell migration and invasion was examined. The wound
healing assays revealed that SW-13 and NCI-H295R cells had
different migration potentials after 24 h of intervention with
different concentrations of TUDCA. The group treated with 400 µM
TUDCA induced the highest percentage of SW-13 cell migration, while
600 µM TUDCA induced the highest percentage of NCI-H295R cell
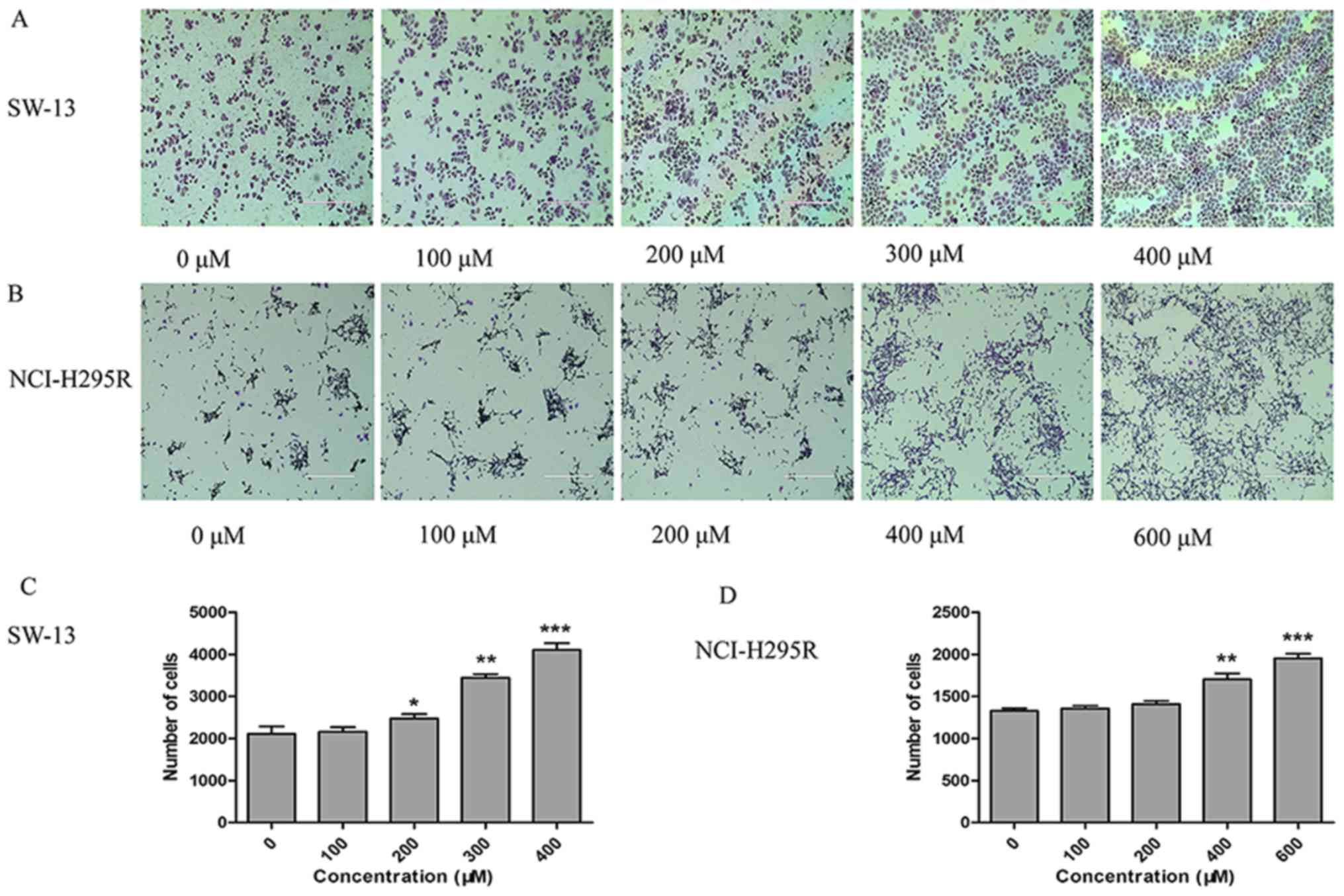
migration (Fig. 2A-D). The Transwell
assay confirmed that the invasiveness of SW-13 cells was
significantly increased in the 400 µM TUDCA group compared with
that in the 0 µM TUDCA group, and the invasiveness of NCI-H295R
cells was significantly increased in the 600 µM TUDCA group
compared with that in the 0 µM TUDCA group (Fig. 3A-D).
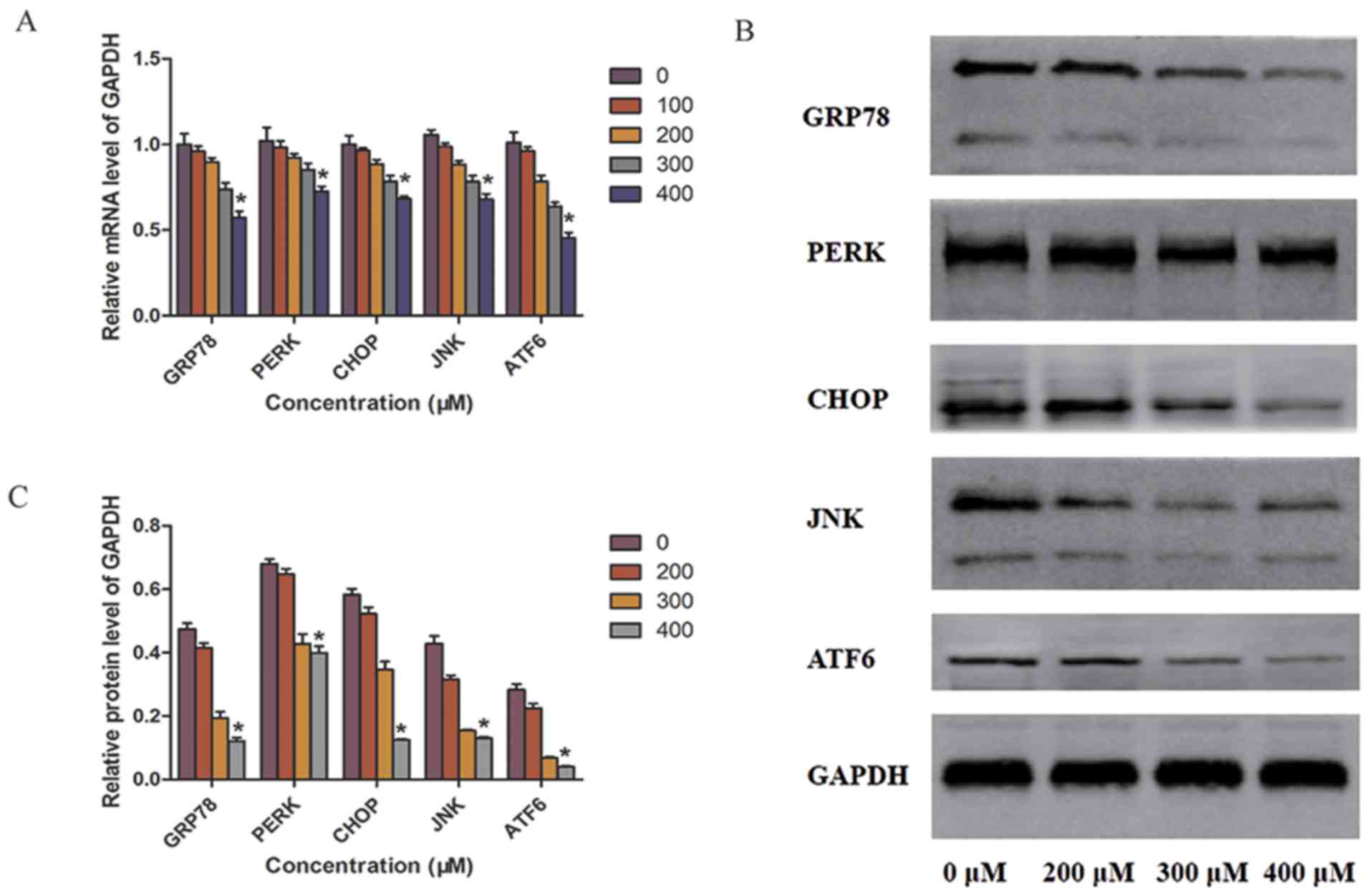
TUDCA alleviates the influence of ER
stress in SW-13 cells
In the present study, whether TUDCA regulated the ER
stress pathway in SW-13 cells was investigated using RT-qPCR and
western blot analyses. The mRNA expression of the ER stress
promoter GRP78 was significantly decreased in the 400 µM TUDCA
group. In accordance with the changes in mRNA expression, western
blot analysis showed that the expression of GRP78 protein was also
markedly decreased in the 400 µM TUDCA group compared with that in
the 0 µM TUDCA group after 48 h. Decreased expression of GRP78
alleviated ER stress in SW-13 cells. In addition, the expression of
ER-related factors was downregulated with TUDCA treatment; PERK and
ATF6 mRNA and protein expression levels were significantly reduced
in the 400 µM TUDCA group compared with those in the 0 µM TUDCA
group. The mRNA and protein expression of other downstream factors,
CHOP and JNK, also correspondingly decreased with TUDCA treatment.
There were no significant differences in the mRNA levels of ER
stress-related factors between the 100 µM TUDCA group and the 0 µM
TUDCA group. Therefore, the protein levels of 100 µM TUDCA group
was not compared using western blot analysis (Fig. 4A-C).
TUDCA induces autophagy and inhibits
apoptosis in SW-13 cells
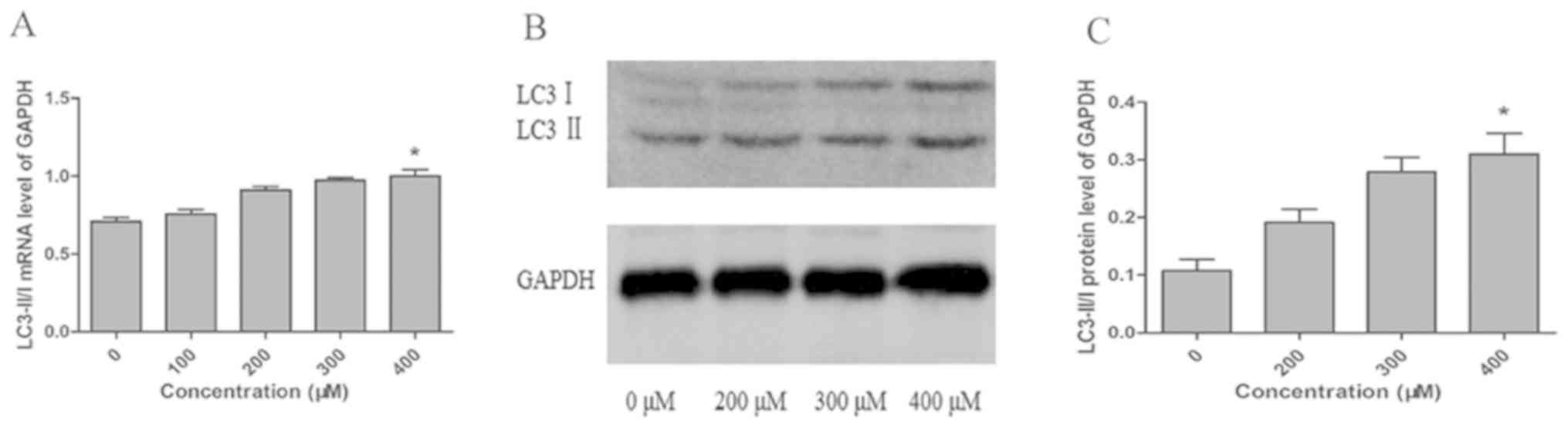
LC3 serves as one of the most important markers of
autophagy. When autophagy occurs, the LC3-I cytosolic form is
converted to the LC3-II lipid-conjugated membrane-bound form
(22). Our results showed that
LC3-II/I expression increased in a concentration-dependent manner
after TUDCA intervention, and LC3-II/I mRNA and protein expression
in the 400 µM TUDCA group was significantly higher compared with
that in the 0 µM TUDCA group (Fig.
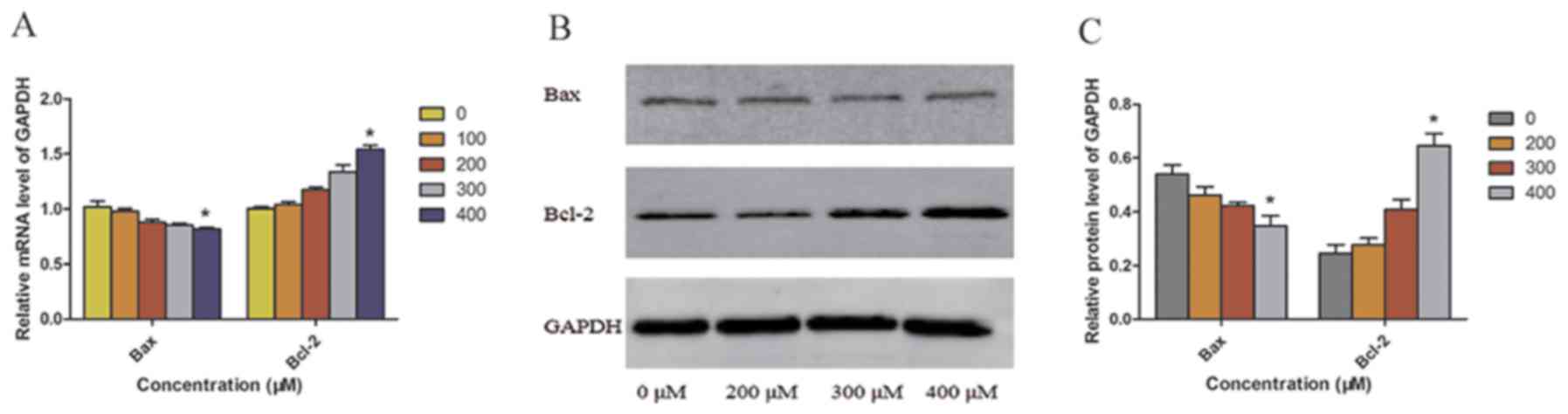
5A-C). In addition, TUDCA alleviated ER stress and induced
autophagy, thereby inhibiting apoptosis. In the 400 µM TUDCA group,
the expression of the anti-apoptotic factor Bcl-2 was upregulated,
and the mRNA and protein expression of Bcl-2 was significantly
higher in the 400 µM TUDCA group compared with that in the 0 µM
TUDCA group. However, the mRNA and protein expression of the
pro-apoptotic factor Bax exhibited the opposite expression pattern
from Bcl-2 (Fig. 6A-C).
Discussion
TUDCA is a well-known ER chaperone that alleviates
ER stress (23). Activation of the
UPR signaling pathway following ER stress plays an important role
in tumorigenesis (24). UPR
activation involves GRP78, PERK, inositol-requiring protein 1α
(IRE1α), and ATF6 (25). The
competitive binding of GRP78 with the hydrophobic surfaces of
misfolded proteins following ER stress releases these transducers
to induce UPR signaling. GRP78, the ER promoter, acts as a major
regulator of the UPR through directly interacting with the UPR
sensors, namely, PERK, IRE1α, and ATF6 (26). When TUDCA inhibited ER stress, three
ER sensors were also inhibited. The dissociation of GRP78 from
PERK, IRE1α, and ATF6 leads to their inhibition. Liu et al
(27) administered TUDCA to salivary
adenoid cystic carcinoma cell cultures 3 h before ceramide
treatment. Ceramide-induced X-box binding protein 1 mRNA splicing
was significantly inhibited by TUDCA pretreatment, and TUDCA
inhibited ceramide-induced eIF2α phosphorylation. TUDCA alleviated
ceramide-induced ER stress. Marquardt et al (28) reported that TUDCA ameliorated
maladaptive ER stress signaling and reduced the expression of ATF6
and CHOP. In the present study, it was observed that TUDCA
alleviates ER stress and promotes ACC cell proliferation. Thus, ER
stress-related signaling pathways play a role in ACC.
Autophagy is important for the secretion of diverse
proteins involved in intercellular signaling and cancer
progression. Autophagic flux can be upregulated in response to
stressful stimuli, such as nutritional, metabolic, oxidative,
pathogenic, genotoxic, or proteotoxic stress (29). This stimulus-induced autophagy serves
a cytoprotective function by helping cells adapt to stress and
promoting cell survival (30,31). As
one of the most important markers of autophagy, the transition of
LC3 from LC3-I to LC3-II mediates key features of autophagy
activation by promoting lysis, lipogenesis, and proteolysis
(22). Some studies showed that the
PERK-eIF2α-ATF4-CHOP pathway contributed to the ER stress-induced
activation of autophagy (32,33). A
study by B'Chir et al (32)
showed that the LC3-II/LC3-I ratio was increased after knockout of
CHOP. CHOP is a downstream indicator of the ER stress signaling
pathway and a key participant in apoptosis. CHOP can activate a
death program by inducing both the extrinsic and intrinsic
apoptotic pathways, decreasing the expression of the anti-apoptotic
protein Bcl-2 while increasing the expression of the pro-apoptotic
Bax (34). The present study found
that TUDCA alleviated ER stress via the PERK-ATF4-CHOP pathway and
induced autophagy in ACC SW-13 cells. Meanwhile, CHOP expression
was downregulated, which may inhibit ACC cell apoptosis. A previous
study has shown that TUDCA inhibited ER stress in pancreatic cancer
cells and blocked the pachymic acid-induced apoptosis of pancreatic
cancer (35). These results provide
evidence demonstrating that TUDCA alleviated ER stress and induced
autophagy in ACC cells, thereby inhibiting the apoptosis of ACC
cells.
In conclusion, TUDCA induced autophagy of ACC SW-13
cells and inhibited apoptosis of ACC SW-13 cells after alleviating
ER stress of ACC SW-13 cells. ER stress- and autophagy-related
signaling pathways are involved in the occurrence of ACC, which may
provide potential therapeutic targets for ACC treatment. There is a
complex interplay between ER stress and autophagy in ACC.
Therefore, an ER stress inducer such as thapsigargin will be used
in in vitro and in vivo experiments with ACC in
future studies.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China (grant no. 81060220), the Innovation
Project of Guangxi Graduate Education (grant no. YCBZ2018037), and
the Guangxi Medical and Health Self-financing Project (grant no.
Z20180636).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XH, DL and ZL designed the study. XH, LW, YK, XL,
XD, XHL, LL, HY, ZH, DL and ZL conducted the study and analyzed the
data. XH wrote the manuscript, and ZL revised the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ACC
|
adrenocortical carcinoma
|
|
ER
|
endoplasmic reticulum
|
|
TUDCA
|
tauroursodeoxycholic acid
|
|
UPR
|
unfolded protein response
|
|
PERK
|
protein kinase R-like ER kinase
|
|
eIF2α
|
eukaryotic initiation factor-2α
|
|
ATF4
|
activating transcription factor 4
|
|
GRP78
|
glucose-regulated protein 78
|
|
IRE1α
|
inositol-requiring protein 1α
|
References
|
1
|
Xie C, Tanakchi S, Raygada M, Davis JL and
Del Rivero J: Case report of an adrenocortical carcinoma associated
with germline CHEK2 mutation. J Endocr Soc. 3:284–290. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Xiao H, Xu D, Chen P, Zeng G, Wang X and
Zhang X: Identification of five genes as a potential biomarker for
predicting progress and prognosis in adrenocortical carcinoma. J
Cancer. 9:4484–4495. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li Y, Bian X, Ouyang J, Wei S, He M and
Luo Z: Nomograms to predict overall survival and cancer-specific
survival in patients with adrenocortical carcinoma. Cancer Manag
Res. 10:6949–6959. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Else T, Kim AC, Sabolch A, Raymond VM,
Kandathil A, Caoili EM, Jolly S, Miller BS, Giordano TJ and Hammer
GD: Adrenocortical carcinoma. Endocr Rev. 35:282–326. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gonzalez RJ, Tamm EP, Ng C, Phan AT,
Vassilopoulou-Sellin R, Perrier ND, Evans DB and Lee JE: Response
to mitotane predicts outcome in patients with recurrent adrenal
cortical carcinoma. Surgery. 142:867–875. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schteingart DE, Doherty GM, Gauger PG,
Giordano TJ, Hammer GD, Korobkin M and Worden FP: Management of
patients with adrenal cancer: Recommendations of an international
consensus conference. Endocrine-Related Cancer. 12:667–680. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mohan DR, Lerario AM and Hammer GD:
Therapeutic targets for adrenocortical carcinoma in the genomics
era. J Endocr Soc. 2:1259–1274. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Baudin E; Endocrine Tumor Board of Gustave
R, : Adrenocortical carcinoma. Endocrinol Metab Clin North Am.
44:411–434. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Teng Y, Zhao H, Gao L, Zhang W, Shull AY
and Shay C: FGF19 protects hepatocellular carcinoma cells against
endoplasmic reticulum stress via activation of FGFR4- GSK3β-Nrf2
Signaling. Cancer Res. 77:6215–6225. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kim C and Kim B: Anti-cancer natural
products and their bioactive compounds inducing ER stress-mediated
apoptosis: A review. Nutrients. 10(pii): E10212018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Limonta P, Moretti RM, Marzagalli M,
Fontana F, Raimondi M and Montagnani Marelli M: Role of endoplasmic
reticulum stress in the anticancer activity of natural compounds.
Int J Mol Sci. 20(pii): E9612019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang M, Law ME, Castellano RK and Law BK:
The unfolded protein response as a target for anticancer
therapeutics. Crit Rev Oncol Hematol. 127:66–79. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Moon HW, Han HG and Jeon YJ: Protein
quality control in the endoplasmic reticulum and cancer. Int J Mol
Sci. 19(pii): E30202018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhou F, Yang X, Zhao H, Liu Y, Feng Y, An
R, Lv X, Li J and Chen B: Down-regulation of OGT promotes cisplatin
resistance by inducing autophagy in ovarian cancer. Theranostics.
8:5200–5212. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li S, Guo L, Qian P, Zhao Y, Liu A, Ji F,
Chen L, Wu X and Qian G: Lipopolysaccharide induces autophagic cell
death through the PERK-dependent branch of the unfolded protein
response in human alveolar epithelial A549 cells. Cell Physiol
Biochem. 36:2403–2417. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Luchetti F, Crinelli R, Cesarini E,
Canonico B, Guidi L, Zerbinati C, Di Sario G, Zamai L, Magnani M,
Papa S and Iuliano L: Endothelial cells, endoplasmic reticulum
stress and oxysterols. Redox Biol. 13:581–587. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen Y, Liu CP, Xu KF, Mao XD, Lu YB, Fang
L, Yang JW and Liu C: Effect of taurine-conjugated ursodeoxycholic
acid on endoplasmic reticulum stress and apoptosis induced by
advanced glycation end products in cultured mouse podocytes. Am J
Nephrol. 28:1014–1022. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang Y, Tang X, Hao F, Ma Z, Wang Y, Wang
L and Gao Y: Bavachin induces apoptosis through mitochondrial
regulated ER stress pathway in HepG2 cells. Biol Pharm Bull.
41:198–207. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Guo Q, Shi Q, Li H, Liu J, Wu S, Sun H and
Zhou B: Glycolipid metabolism disorder in the liver of obese mice
is improved by TUDCA via the restoration of defective hepatic
autophagy. Int J Endocrinol. 2015:6879382015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li J, Wang Y, Ge J, Li W, Yin L, Zhao Z,
Liu S, Qin H, Yang J, Wang L, et al: Doublecortin-like kinase 1
(DCLK1) regulates B cell-specific moloney murine leukemia virus
insertion site 1 (Bmi-1) and is associated with metastasis and
prognosis in pancreatic cancer. Cell Physiol Biochem. 51:262–277.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rahman FU, Ali A, Duong HQ, Khan IU,
Bhatti MZ, Li ZT, Wang H and Zhang DW: ONS-donor ligand based
Pt(II) complexes display extremely high anticancer potency through
autophagic cell death pathway. Eur J Med Chem. 164:546–561. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vandewynckel YP, Laukens D, Devisscher L,
Paridaens A, Bogaerts E, Verhelst X, Van den Bussche A, Raevens S,
Van Steenkiste C, Van Troys M, et al: Tauroursodeoxycholic acid
dampens oncogenic apoptosis induced by endoplasmic reticulum stress
during hepatocarcinogen exposure. Oncotarget. 6:28011–28025. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wu CH, Silvers CR, Messing EM and Lee YF:
Bladder cancer extracellular vesicles drive tumorigenesis by
inducing the unfolded protein response in endoplasmic reticulum of
nonmalignant cells. J Biol Chem. 294:3207–3218. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Luo B and Lee AS: The critical roles of
endoplasmic reticulum chaperones and unfolded protein response in
tumorigenesis and anticancer therapies. Oncogene. 32:805–818. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Guzel E, Arlier S, Guzeloglu-Kayisli O,
Tabak MS, Ekiz T, Semerci N, Larsen K, Schatz F, Lockwood CJ and
Kayisli UA: Endoplasmic reticulum stress and homeostasis in
reproductive physiology and pathology. Int J Mol Sci. 18:E7922017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu Z, Xia Y, Li B, Xu H, Wang C, Liu Y,
Li Y, Li C, Gao N and Li L: Induction of ER stress-mediated
apoptosis by ceramide via disruption of ER Ca(2+) homeostasis in
human adenoid cystic carcinoma cells. Cell Biosci. 4:712014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Marquardt A, Al-Dabet MM, Ghosh S, Kohli
S, Manoharan J, ElWakiel A, Gadi I, Bock F, Nazir S and Wang H:
Farnesoid X receptor agonism protects against diabetic tubulopathy:
Potential add-on therapy for diabetic nephropathy. J Am Soc
Nephrol. 28:3182–3189. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Galluzzi L, Pietrocola F, Bravo-San Pedro
JM, Amaravadi RK, Baehrecke EH, Cecconi F, Codogno P, Debnath J,
Gewirtz DA, Karantza V, et al: Autophagy in malignant
transformation and cancer progression. EMBO J. 34:856–880. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Amaravadi R, Kimmelman AC and White E:
Recent insights into the function of autophagy in cancer. Genes
Dev. 30:1913–1930. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cotzomi-Ortega I, Aguilar-Alonso P,
Reyes-Leyva J and Maycotte P: Autophagy and its role in protein
secretion: Implications for cancer therapy. Mediators Inflamm.
2018:42315912018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
B'Chir W, Chaveroux C, Carraro V, Averous
J, Maurin AC, Jousse C, Muranishi Y, Parry L, Fafournoux P and
Bruhat A: Dual role for CHOP in the crosstalk between autophagy and
apoptosis to determine cell fate in response to amino acid
deprivation. Cell Signal. 26:1385–1391. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lei Y, Wang S, Ren B, Wang J, Chen J, Lu
J, Zhan S, Fu Y, Huang L and Tan J: CHOP favors endoplasmic
reticulum stress-induced apoptosis in hepatocellular carcinoma
cells via inhibition of autophagy. PLoS One. 12:e01836802017.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Puthalakath H, O'Reilly LA, Gunn P, Lee L,
Kelly PN, Huntington ND, Hughes PD, Michalak EM, McKimm-Breschkin
J, Motoyama N, et al: ER stress triggers apoptosis by activating
BH3-only protein Bim. Cell. 129:1337–1349. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cheng S, Swanson K, Eliaz I, McClintick
JN, Sandusky GE and Sliva D: Pachymic acid inhibits growth and
induces apoptosis of pancreatic cancer in vitro and in vivo by
targeting ER stress. PLoS One. 10:e01222702015. View Article : Google Scholar : PubMed/NCBI
|