Introduction
Lung cancer is the most common cancer worldwide in
terms of incidence and mortality rates, and non-small cell lung
cancer (NSCLC) is a histological subtype that represents ~85% of
cases (1). Despite advances in
diagnosis and treatment, at present, patients with NSCLC have a
poor prognosis; the overall 5-year survival rate of patients with
NSCLC was <20% between 2013 and 2018, with high mortality rates
due to high rates of invasion, metastasis and postoperative
recurrence (2,3). Chemotherapy is an efficient treatment
for patients with NSCLC (4).
However, frequent multidrug resistance during treatment markedly
restricts the curative effect (5).
Cisplatin (CDDP) is one of the first-line drugs for NSCLC
treatment; however, resistance to CDDP frequently limits its
efficacy in a clinical setting (6,7).
Therefore, a comprehensive understanding of the etiology and exact
molecular mechanism underlying CDDP resistance in NSCLC development
and progression is essential to improve the therapeutic efficacy
for patients with NSCLC.
Long non-coding RNAs (lncRNAs) are a type of
non-encoding RNA transcript of >200 nucleotides in length, which
do not encode proteins; however, they have the ability to regulate
numerous biological processes in tumorigenesis, such as
transcriptional and post-trantscriptional regulation, and protein
modification (8,9). lncRNAs have attracted large attention
due to their critical role in cancer progression (10). In addition, previous studies have
hypothesized that lncRNAs may interact with microRNAs (miRNAs) as
natural miRNA sponges or competitive endogenous RNAs (ceRNAs) to
regulate the expression of target genes, which serves a crucial
role in the network of ceRNAs in human diseases (11,12).
Myocardial infarction-associated transcript (MIAT)
is a type of lncRNA located on chromosome 22q12.1. It is 30,051 bp
in length and was first identified in the year 2000 (13). MIAT was originally reported to be
associated with myocardial infarction, and is abundantly expressed
in the nervous system and retinal tissue (14,15).
Recent studies have suggested that MIAT is upregulated and serves
as an oncogene in several types of cancer (16). MIAT is involved in a positive
feedback regulatory loop with its transcriptional regulator
octamer-binding protein 4 to promote cell proliferation in human
malignant mature B cell chronic lymphocytic leukemia and functions
as aceRNA of miRNA (miR)-155-5p to regulate dual specificity
protein phosphatase 7 in breast cancer (17,18).
MIAT was additionally demonstrated to be able to promote NSCLC
proliferation and metastasis through the activation of matrix
metalloproteinase 9 (19,20). Although a number of previous studies
demonstrated a partially oncogenic role of MIAT in NSCLC, the
biological mechanisms and miR-sponging role of MIAT in the
progression of NSCLC, particularly in CDDP resistance, have yet to
be comprehensively elucidated.
The present study hypothesized that MIAT over
expression was a characteristic molecular alteration that was
highly associated with poor prognosis in patients with NSCLC.
Materials and methods
Clinical specimens and cell lines
NSCLC tissues and corresponding adjacent normal
tissues (~4 mm away from the tumor) were collected during
therapeutic surgical resection from 60 patients (male, n=37;
female, n=23) with NSCLC between May 2017 and February 2019at the
First Affiliated Hospital of Gannan Medical University (Ganzhou,
China) according to the National Institutes of Health guidelines.
The patients underwent surgery without radiotherapy, chemotherapy
or any other treatment, and the patient age ranged between 37 and
79 years (≥60 years, n=32; <60 years, n=28; mean age,
58.88±12.65 years). The clinical stage was determined according to
the 8th edition of the TNM classification for lung cancer from
American Joint Committee on Cancer (21). Tissues were frozen in liquid nitrogen
following RNA extraction. The procedures in this study involving
animal and human tissue were approved by the Human Ethics Committee
of the First Affiliated Hospital of Gannan Medical University.
Written informed consent was obtained from each patient prior to
the use of their tissues for scientific research. A 2-year
follow-up survival survey was conducted; overall survival (OS) was
defined as the interval between resection and mortality or the date
of last follow-up. The human NSCLC H1299 and CDDP resistant H1299
(H1299-CDDP) cells used in the present study were obtained from the
cell bank of Type Culture Collection of the Chinese Academy of
Sciences and were cultured in 90% RPMI-1640 medium (Gibco; Thermo
Fisher Scientific, Inc.) supplemented with 10% fetal bovine serum
(Gibco; Thermo Fisher Scientific, Inc.), 1% penicillin and
streptomycin at 37°C in a humidified atmosphere containing 5%
CO2.
RNA interference and cell
transfection
The small interfering (si)RNA oligonucleotides
si-MIAT
(5′-GATCCCCGGACAGAGAATGCAAATAATTCAAGAGATTATTTGCATTCTCTGTCCTTTTTA-3′)
and anti-miR-184 (5′-GCUAGAACUUGUCCUAGCGCUUGGCACAUAUTT-3′), in
addition topcDNA3.1-MIAT (MIAT) and miR-184-mimics (miR-184,
5′-TCTACAGTGCCAAGGACTGCACCTCACGTGTCTCCAG-3′) were purchased from
Shanghai GenePharma Co., Ltd. H1299 and H1299-CDDP cells were
transfected with siRNA or negative controls using RNAiMAX
Lipofectamine® (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. H1299 Cells were
seeded in 6-well plates at a density of 2×105
cells/well. In total, 2 µg plasmid DNA or 100 pmol siRNA, which was
suspended in 50 µl Opti-minimum essential medium (MEM) (Invitrogen;
Thermo Fisher Scientific, Inc.), was mixed with 10 µl
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) and left for 20 min at 25°C to form a mixture.
Prior to replacing the cell culture medium with 400 µl Opti-MEM,
H1299 cells were incubated with the mixture at 37°C in a humidified
atmosphere containing 5% CO2. After 6 h, the mixture was
discarded and the cells were cultured in complete medium at 37°C
for 24 h. Subsequently, the cells were harvested for reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) and
western blot analyses.
Dual-luciferase reporter assay
TargetScan (http://www.targetscan.org/) and miRcode (http://www.mircode.org/) were used with bioinformatics
databases Starbase v2.0 (http://starbase.sysu.edu.cn/starbase2/), miRcode
(http://www.mircode.org/) and TargetScanHuman
Release 7.2 (http://www.targetscan.org/vert_72/) to identify the
potential target sequences of miR-184. MIAT sequences or splicing
factor 1 (SF1) 3′UTR fragments containing corresponding wild-type
or mutant-type miR-184 binding site were amplified by PCR and
subcloned into pcDNA3.1 luciferase reporter vectors. The
pcDNA3.1-MIAT wild-type (WT) or mutant (MUT) and pcDNA3.1-SF1 WT or
mutant vectors, with and without designed miR-184 binding sites,
were obtained from Shanghai GenePharma Co., Ltd., and
co-transfected with miR-184-mimics or negative controls in
H1299-CDDP cells using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.). At 24–48 h following transfection,
firefly and Renilla luciferase activities were measured
using a Dual-Luciferase Reporter Assay System (Promega
Corporation). All values were obtained from at least three
independent experiments.
Cell proliferation assay
The cell viability of H1299 and H1299-CDDP cells was
assessed by MTT assay. Following incubation of the cells in 96-well
plates for 0, 12, 24, 48, 72 h, 20 µl 5 mg/ml thiazolyl blue in PBS
was added to each well, and the plates were incubated at 37°C for
an additional 2 h. The medium was subsequently removed and the
purple colored formazan precipitates were dissolved in 150 µl
dimethyl sulfoxide. The absorbance was recorded at 490 nm using a
Thermo microplate reader (Rayto Life and Analytical Sciences Co.,
Ltd.). A colony formation assay was performed, according to a
standard protocol (22). H1299 and
H1299-CDDP cells were seeded in 6-well plates (2,000 cells/well)
and incubated under the aforementioned culture conditions for ~10
days. Subsequently, cells were fixed with methanol for 15 min and
stained with 0.5% crystal violet solution for 25 min at room
temperature. Colonies (>50 cells) were counted under a light
microscope (×4 magnification). The data are presented as the mean
of three independent experiments.
Cell apoptosis assay
A cell apoptosis assay was performed with an Annexin
V-fluorescein isothiocyanate (FITC) apoptosis detection kit (BD
Biosciences) according to the manufacturer's protocol. H1299 and
H1299-CDDP cells (1–2×105 cells/well) were seeded in a
6-well plate for 48 h. Subsequently, cells were harvested and
washed twice with ice-cold PBS, and incubated with 5 µl Annexin
V-FITC and 2 µl propidium iodide (BD Biosciences) in 100 µl binding
buffer for 15 min at room temperature. The levels of apoptosis were
examined using the aforementioned apoptosis detection kit,
according to the manufacturer's protocol, by flow cytometry
(FACScan; BD Biosciences). The data were analyzed using Cell Quest
software (version 5.1; BD Biosciences).
RT-qPCR
Total RNA from transfected H1299-CDDP cells, tumor
and adjacent normal tissues was isolated with TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. RT was conducted with random primers
using an RT-PCR kit (cat. no. DWA005; Takara Biotechnology Co.,
Ltd.) at 37°C for 15 min and 85°C 15 sec. qPCR was performed with
iQ SYBR Green Supermix (Bio-Rad Laboratories, Inc.) according to
the manufacturer's protocol. The thermocycling conditions were as
follows: 95°C for 30 sec, followed by 40 cycles of 95°C for 5 sec
and 60°C for 34 sec. The primer sequences used were as follows:
MIAT, forward: 5′-TCTTCATGTCAGAACACGCTTTA-3′ and reverse:
3′-AAGGTCACCCGAGGTCCAA-5′; miR-184 stem loop,
5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACTTCCCA-3′, forward,
5′-TGGACGGAGAACTGATAAGGGT-3′ and reverse,
3′-CCTTATCAGTTCTCCGTCCATT-5′; and GAPDH forward,
5′-TCTCTGCTCCTCCTGTTC-3′ and reverse,
3′-GGTTGAGCACAGGGTACTTTATTGA-5′. GAPDH mRNA was used as an
endogenous control for mRNA. The relative expression level was
calculated using the 2−∆∆Cq method (23) and each experiment was repeated at
least three times.
Western blot analysis
Total protein was extracted with
Radio-Immunoprecipitation Assay Buffer, quantified by BCA protein
detection method (Beyotime Institute of Biotechnology) and
separated by 10% SDS-PAGE using 20 µg of protein per lane. The
proteins were subsequently transferred to polyvinylidene difluoride
(PVDF) membranes (EMD Millipore). The PVDF membranes were blocked
with 5% skim milk at room temperature for 2–4 h. Subsequently, the
PVDF membranes were incubated with primary antibodies (all 1:1,000;
Abcam) against β-actin (ab179467), B-cell lymphoma-2
(Bcl-2)-associated X protein (Bax) (ab232479), Bcl-2(ab32124),
cleaved caspase-3 (ab2302) and SF1 (ab65815) at 4°C overnight.
Following incubation with the relevant secondary antibodies
(horseradish peroxidase-conjugated goat anti-mouse IgG; 1:5,000;
abs20001ss; ABSIN) for 2 h at 37°C, the proteins were visualized by
autoradiography using enhanced chemiluminescence (Bio-Rad
Laboratories, Inc.). The relative expression of the protein of
interest was presented as a grayscale ratio of the protein to
β-actin and the results were analyzed using Graph Pad Prism
software (GraphPad Software 6.0, Inc.).
In vivo tumor formation assay
A total of 20 male BALB/c nude mice (~4 weeks old)
were obtained from Beijing Huafukang Bioscience Co. Ltd. The in
vivo xenograft assay was performed according to guidelines of
the Institutional Animal Care and Use Committee (IACUC) and
approved by the institutional guidelines of Gannan Medical
University. H1299-CDDP cells (1×107 cells/mouse)
transfected with si-MIAT or si-MIAT negative controls were injected
into the right flanks of 30 mice. After 7 days, mice bearing tumors
~100 mm3 were divided into four groups (n=5 mice/group):
si-NC + PBS, si-NC + CDDP, si-MIAT + PBS and si-MIAT + CDDP. Mice
were treated with CDDP (4 mg/kg) or an equal volume of PBS once
every 3 days through tail vein injections, and the tumor size was
measured simultaneously using a simplified equation (length ×
width2 ×0.5). The weights of the mice were measured
every three days following treatment. The mice were sacrificed by
cervical dislocation at the humane endpoint (excessive weight loss)
and tumors were excised at the end of the treatment period.
Statistical analysis
All experiments were performed at least three times
and the data are presented as the mean ± standard deviation.
Significant differences between two groups were analyzed using a
Student's t-test (parametric) or Mann-Whitney U test
(non-parametric). Tukey's post-hoc test was performed following
one-way ANOVA; least significant difference test was performed
following repeated measures ANOVA to analyze tumor size over time.
The Kaplan-Meier test was used to estimate OS. The log-rank test
was used to analyze the effect of clinical variables and MIAT
expression on patients' OS. P<0.05 was considered to indicate a
statistically significant difference. The statistical analysis was
conducted using Graph Pad software (GraphPad Software 6.0,
Inc.).
Results
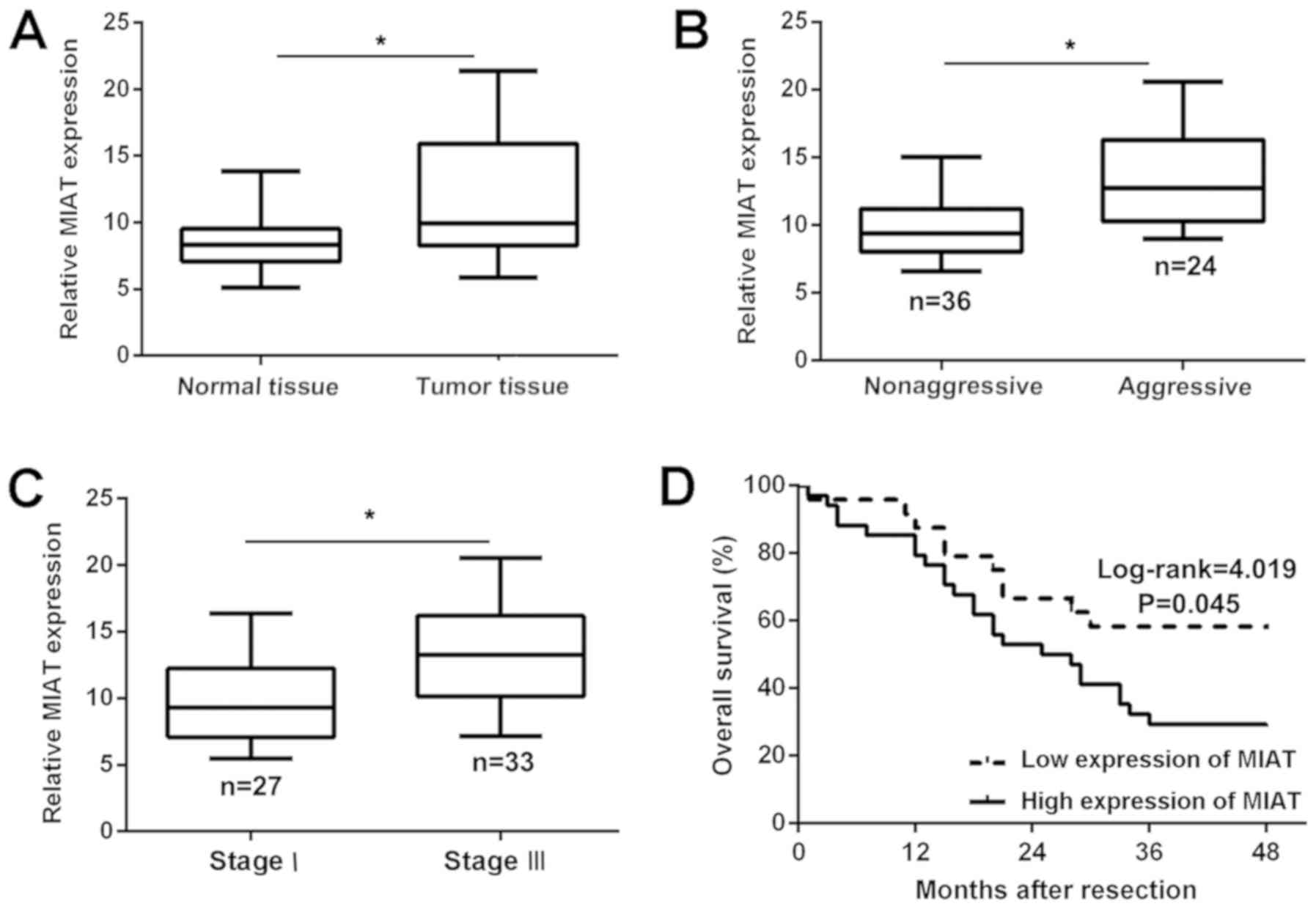
MIAT is overexpressed in NSCLC tissues
and associated with a poor prognosis
The MIAT expression profile in NSCLC and adjacent
normal lung tissues was assessed. In total, 60 pairs of specimens
were analyzed by RT-qPCR, and the results demonstrated that the
expression of MIAT was significantly (P<0.05) upregulated in
NSCLC tissue compared with that in adjacent normal tissue (Fig. 1A). In addition, the expression of
MIAT was revealed to be associated with the metastasis of NSCLC
(Fig. 1B). Association analysis
additionally demonstrated that the overexpression of MIAT was
highly associated with clinical stage (Fig. 1C). Further Kaplan-Meier survival
analysis demonstrated that upregulated expression of MIAT was
positively associated with a poor 2-year survival of NSCLC
(Fig. 1D). Taken together, these
results demonstrated that the overexpression of MIAT may be
associated with metastasis and a poor prognosis in NSCLC.
Knockdown of MIAT inhibits the
proliferation of NSCLC H1299 and H1299-CDDP cells
To examine the role of MIAT in the resistance of
NSCLC to CDDP, the expression profile of MIAT in NSCLC H1299 cells
and CDDP-resistantH1299 cells was determined. H1299 is a common
NSCLC cell line, and the first-line drug for NSCLC in a clinical
setting is a platinum compound, which is prone to drug resistance
(24). Therefore, the present study
investigated and compared H1299 vs. H1299-CDDP cells. Notably, the
expression of MIAT in H1299-CDDP was markedly elevated when
compared with that in H1299, which suggested that MIAT may be
associated with CDDP resistance (Fig.
2A). To investigate the biological role of MIAT in NSCLC, an
overexpression plasmid of MIAT (plasmid-MIAT) was transfected into
H1299 cells, and siRNA targeting the coding region of MIAT
(si-MIAT) was transfected into H1299-CDDP cells. The transfection
efficiencies of plasmid-MIAT and si-MIAT were determined by
RT-qPCR; the results demonstrated that the expression level of MIAT
was decreased in MIAT-siRNA-transfected H1299-CDDP cells and was
significantly (P<0.001) increased in plasmid-MIAT-transfected
H1299 cells (Fig. 2B). MTT and
colony formation assays demonstrated that transfection with MIAT
significantly (P<0.001) attenuated the proliferation of H1299
and H1299-CDDP cells (Fig. 2C-E). In
addition, a flow cytometry was performed to further determine the
effect of MIAT on the apoptosis of H1299-CDDP cells, and a
significantly (P<0.001) increased percentage of apoptotic cells
in si-MIAT-transfected H1299-CDDP cells was observed (Fig. 2F). Furthermore, apoptotic-associated
proteins were analyzed by western blot analysis, which revealed
that the silencing of MIAT led to a decrease in the expression
level of apoptosis by inhibiting protein Bcl-2 expression, whereas
an increased expression level of the pro-apoptotic protein cleaved
caspase-3 was observed (Fig. 2G).
These data demonstrated that MIAT was able to promote NSCLC cell
proliferation.
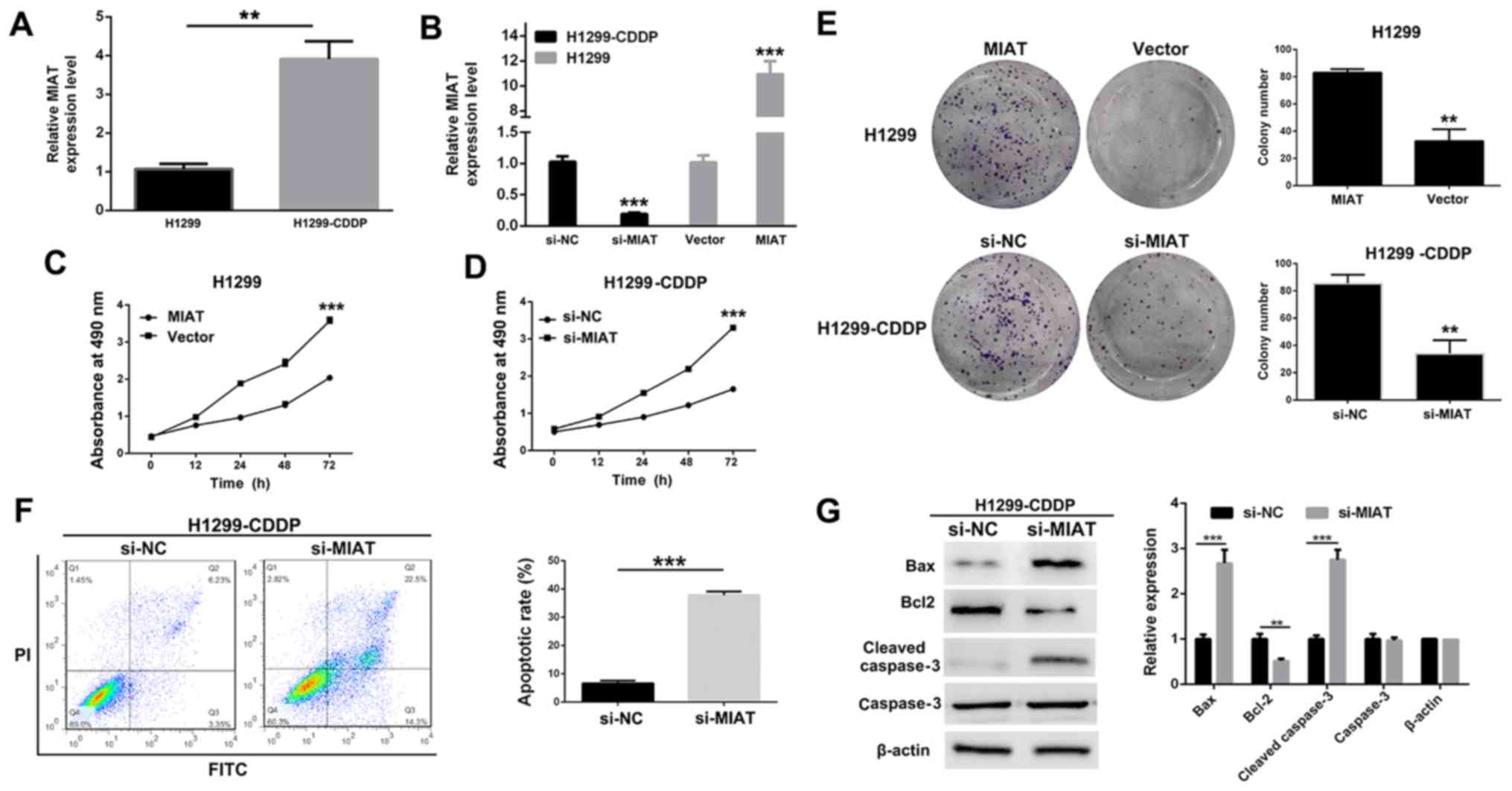 | Figure 2.Functional analysis of MIAT in NSCLC
cells. (A) The MIAT expression level in H1299 and H1299-CDDP cells
was determined by RT-qPCR. (B) The MIAT expression level in
H1299-CDDP cells transfected with si-MIAT or NC-siRNA, as well as
in H1299 cells transfected with pcDNA3.1-MIAT plasmid (MIAT) or
empty vector was determined by RT-qPCR. (C and D) Growth curves of
(C) H1299 cells transfected with MIAT or vector and (D) H1299-CDDP
cells transfected with si-MIAT or si-NC were determined via MTT
assays. (E) Colony formation assays of H1299 cells transfected with
MIAT or vector and H1299-CDDP cells transfected with si-MIAT or
si-NC were performed. (F) Flow cytometry assays were performed to
analyze the apoptosis of H1299-CDDP cells following treatment with
si-MIAT or si-NC. (G) The levels of Bax, Bcl-2 and cleaved
caspase-3 following MIAT silencing in H1299-CDDP cells were
determined via western blotting. *P<0.05, **P<0.01 and
***P<0.001. MIAT, myocardial infarction-associated transcript;
NSCLC, non-small cell lung cancer; NC, negative control; si, small
interfering; CDDP, cisplatin; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; Bcl-2, B-cell
lymphoma-2; Bax, Bcl-2-associated X protein. |
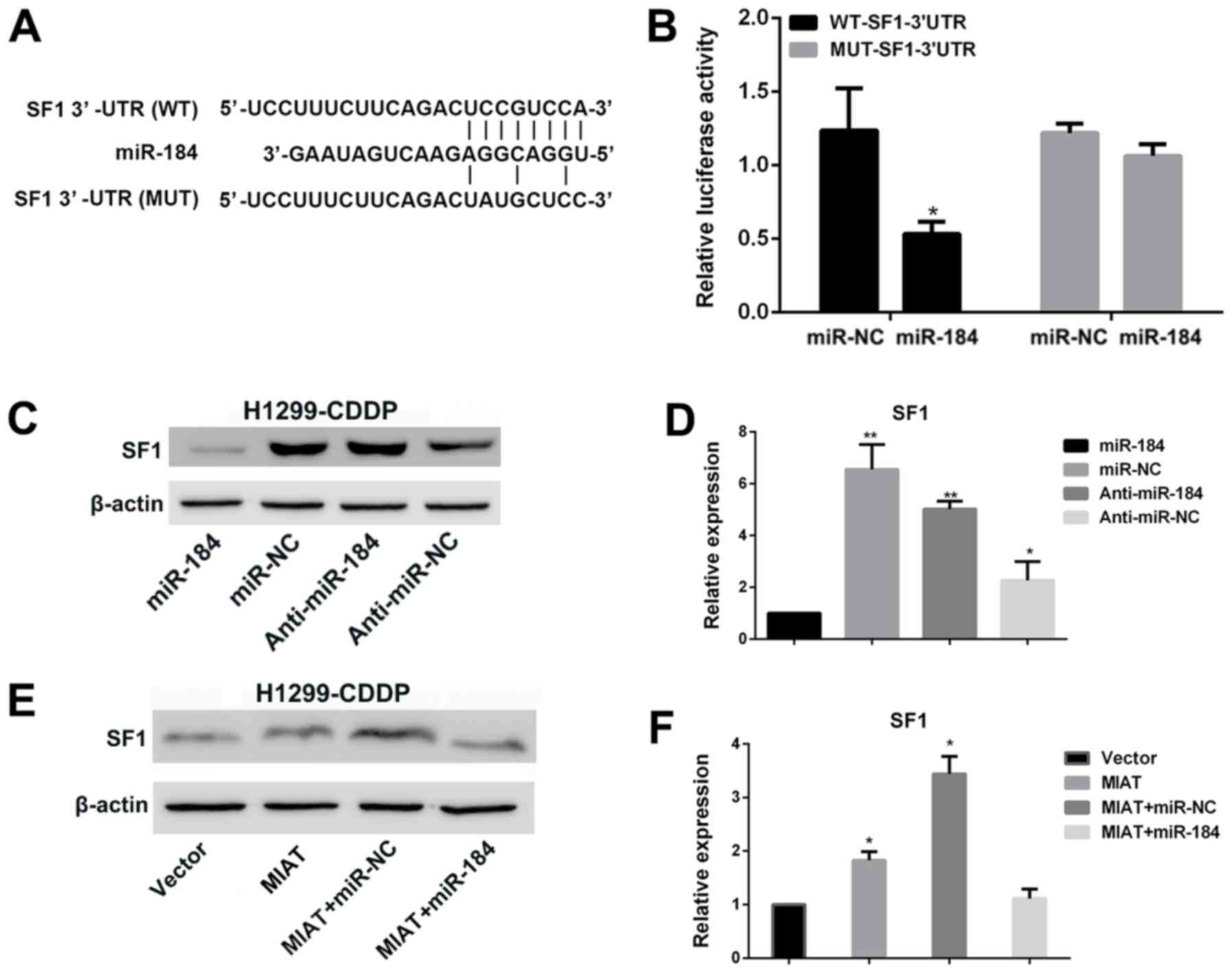
MIAT sponges miR-184 as a ceRNA
To determine whether MIAT was able to interact with
miRs as previously demonstrated, TargetScan (http://www.targetscan.org/) and miRcode (http://www.mircode.org/) were used with bioinformatics
databases (Starbase v2.0, miRcode and TargetScan Human Release
7.2). Putative binding sites between MIAT and miR-184 were
identified (Fig. 3A). In order to
verify the interaction between miR-184 and MIAT at the endogenous
level, a dual-luciferase reporter assay was performed in H1299-CDDP
cells. The data demonstrated that the luciferase activity was
notably decreased when the WT-MIAT-3′untranslated region (3′UTR)
and miR-184 were co-transfected into H1299-CDDP cells compared with
the activity in H1299-CDDP cells co-transfected with MUT-MIAT-3′UTR
and the miR-184 negative control. By contrast, the luciferase
activity of MUT-MIAT-3′UTR exhibited no statistically significant
differences (Fig. 3B). In addition,
the miR-184 expression level was increased when MIAT was silenced
in H1299-CDDP cells (Fig. 3C).
Colony formation and MTT assays demonstrated knocking down MIAT
expression significantly (P<0.001) attenuated the proliferation
of H1299-CDDP cells. However, co-transfection of si-MIAT and
anti-miR-184 into H1299-CDDP cells led to increased proliferation
and colony formation compared with that of cells co-transfected
with si-MIAT and anti-miR-negative control (Fig. 3D-F). The results suggested that MIAT
functions as a ceRNA by sponging miR-184.
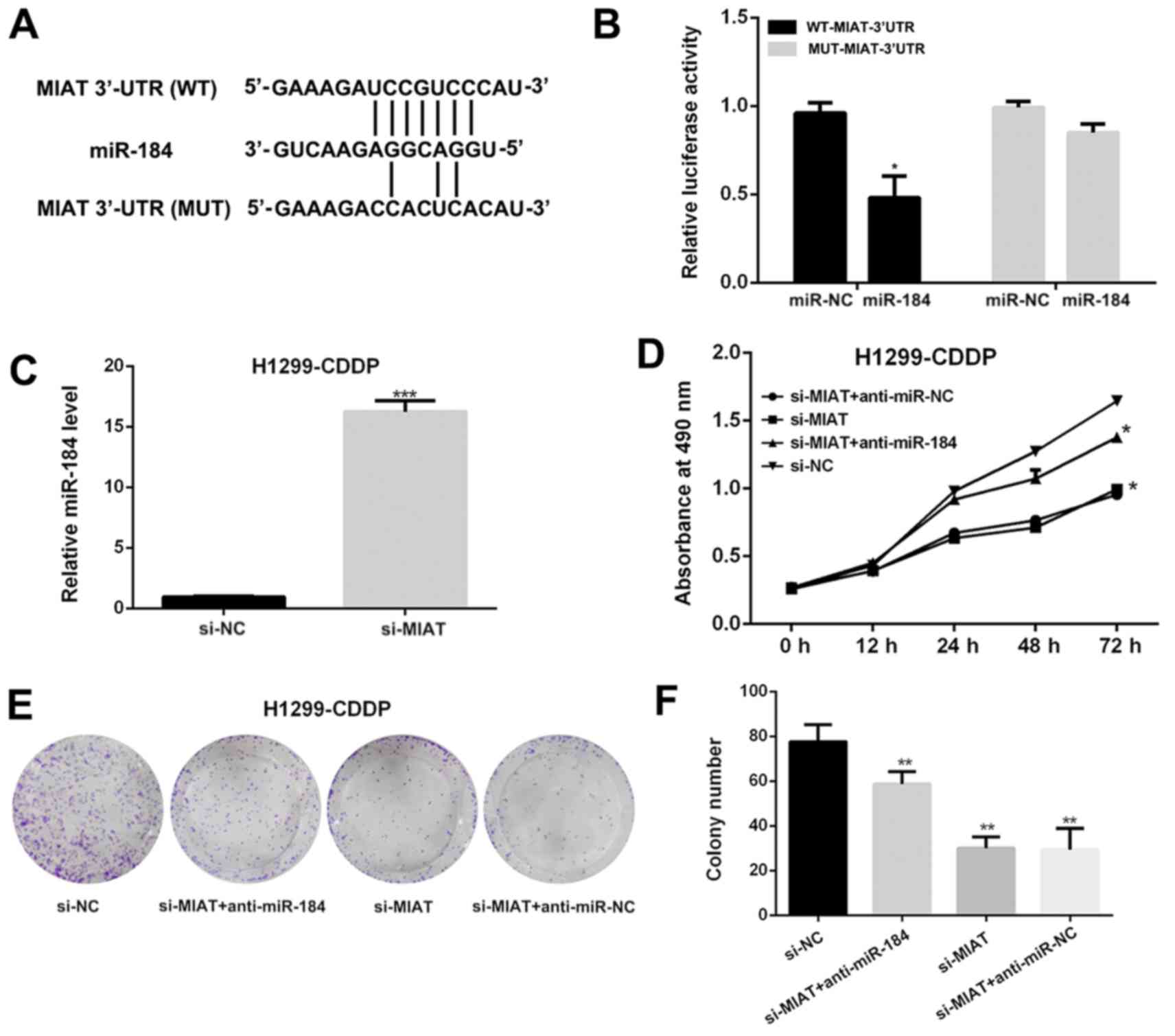 | Figure 3.MIAT acts as a competitive endogenous
RNA by sponging miR-184. (A) The predicted binding sites of MIAT to
miR-184 sequence were illustrated. (B) The luciferase activity of
H1299-CDDP cells co-transfected with miR-184 mimics and luciferase
reporters containing WT-MIAT-3′-UTR or MUT-MIAT-3′-UTR transcripts
were analyzed. (C) The relative miR-184 level in H1299-CDDP cells
transfected with si-MIAT or si-NC was determined by reverse
transcription-quantitative polymerase chain reaction. (D) Growth
curves of H1299-CDDP cells transfected with si-MIAT + anti-miR-NC,
si-MIAT, si-MIAT + anti-miR-184 or si-NC were determined via MTT
assays. (E) Images of colony formation assays of H1299-CDDP cells
transfected with si-MIAT + anti-miR-NC, si-MIAT, si-MIAT +
anti-miR-184 or si-NC were performed. (F) The statistical analysis
of colony formation assays of H1299-CDDP cells transfected with
si-MIAT + anti-miR-NC, si-MIAT, si-MIAT + anti-miR-184 or si-NC.
*P<0.05, **P<0.01 and ***P<0.001 vs. negative control.
MIAT, myocardial infarction-associated transcript; NSCLC, non-small
cell lung cancer; NC, negative control; si, small interfering; WT,
wild type; MUT, mutant; URT, untranslated region; CDDP, cisplatin;
miR, microRNA. |
MIAT promotes the expression of SF1 by
sponging miR-184 in H1299-CDDP cells
Accumulating evidence has suggested that lncRNAs may
serve as ceRNAs to regulate the expression of miRs and mRNAs
(25,26). Therefore, the present study aimed to
elucidate the mechanism underlying the MIAT/miR-184 axis in NSCLC
cells. The bioinformatics tool TargetScan revealed that SF1 is a
candidate target of miR-184 (Fig.
4A). To validate the interaction between miR-184 and SF1, a
dual-luciferase reporter assay was performed in H1299-CDDP cells.
The results demonstrated that the luciferase activity was markedly
decreased whenWT-SF1-3′UTR and miR-184 were co-transfected into
H1299-CDDP cells compared with the activity in H1299-CDDP cells
co-transfected with MUT-SF1-3′UTR and miR-184negative control. By
contrast, the luciferase activity of MUT-SF1-3′UTR exhibited no
statistically significant differences (Fig. 4B). Furthermore, western blot analysis
demonstrated that the overexpression of miR-184led to a markedly
decreased expression level of SF1 compared with that of miR-184
negative control; by contrast, SF1expression was markedly increased
when miR-184 was silenced by anti-miR-184 compared with that caused
byanti-miR-184 negative control in H1299-CDDP cells (Fig. 4C and D). Overexpression of MIAT
additionally induced a notably increased expression level of SF1
(Fig. 4E and F). Although MIAT +
miR-NC upregulated the expression of SF1, the expression level of
SF1 in cells treated with MIAT + miR-184 was significantly
downregulated compared with that in cells treated with MIAT. These
results suggested that upregulated MIAT acts as a ceRNA of miR-184
to regulate SF1 expression.
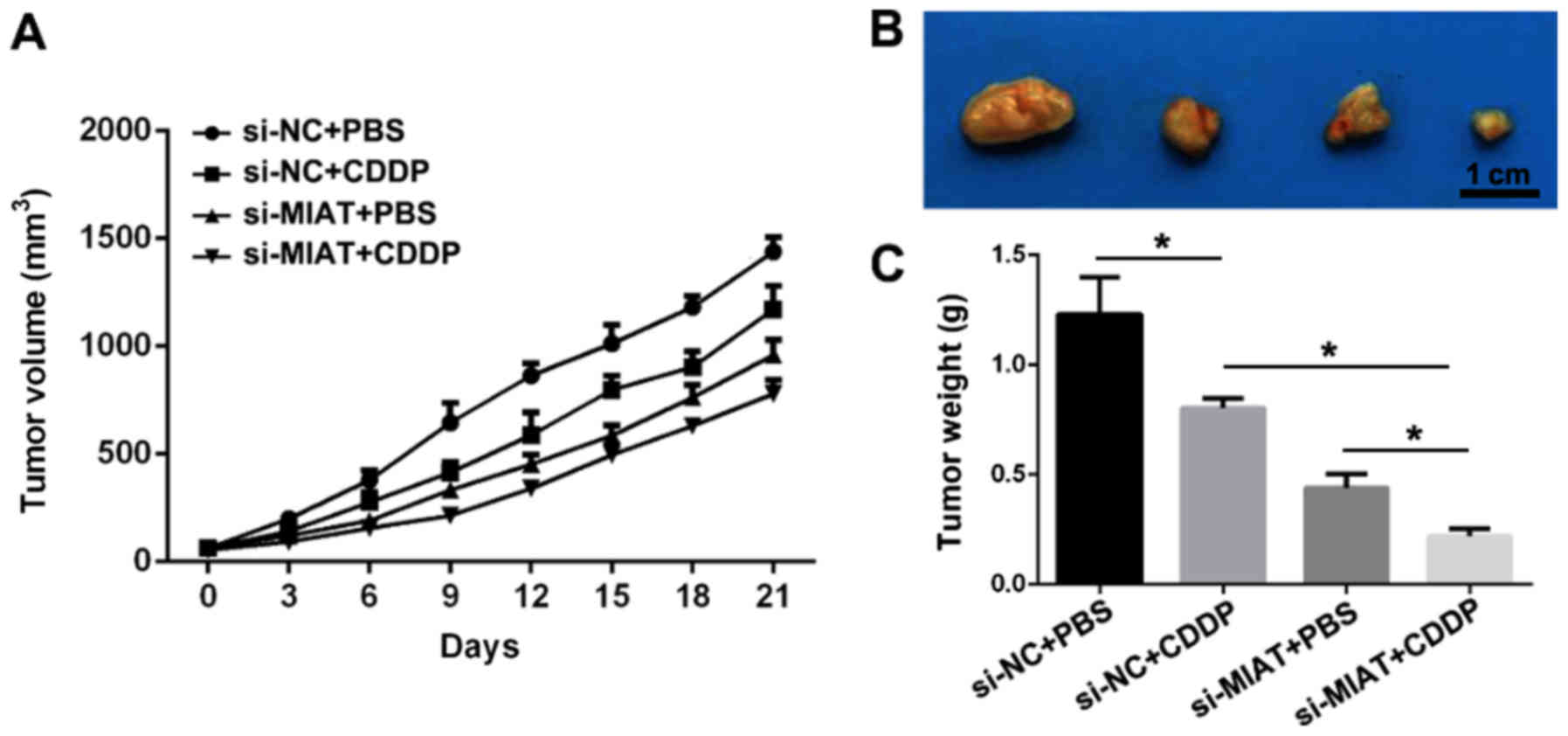
Knockdown of MIAT suppresses tumor
growth and enhances the sensitivity of NSCLC to CDDP in nude
mice
In order to examine the association between MIAT and
CDDP resistance in vivo, a subcutaneous xenograft tumor
model in nude mice was established. H1299-CDDP cells transfected
with si-MIAT or a si-MIAT negative control were subcutaneously
injected into the right flanks of the nude mice. The results
demonstrated that treatment with CDDP or MIAT knockdown markedly
attenuated the tumor growth and tumor volume (Fig. 5A and B), and the tumor weight was
also decreased (Fig. 5C). In
addition, silencing MIAT further intensified the repression
efficacy of CDDP on tumor growth. Taken together, these data
suggested that knockdown of MIAT arrested tumor growth and
increased CDDP sensitivity in NSCLC.
Discussion
Accumulating evidence has demonstrated that lncRNAs
exert complex effects in cancer progression (27,28).
Abnormally expressed lncRNAs have been recognized to exert
oncogenic properties in NSCLC, for instance, upregulated lncRNA
maternally expressed 3 was observed to inhibit NSCLC cell
proliferation and induce apoptosis by affecting p53 expression
(29). In addition, a previous study
demonstrated that overexpressed lncRNA sprout RTK signaling
antagonist 4-intronic transcript 1 promoted NSCLC cell
proliferation and metastasis by affecting the
epithelial-mesenchymal transition via histone-lysine
N-methyltransferase enhancer of zeste 2 (30). Aberrant expression of microRNAs is
associated with tumor progression and metastasis (31,32).
Drug resistance remains a principal challenge in chemotherapy,
since it impedes the curative effect when administered to patients
(7). MIAT has been recognized as a
potent oncogene in breast cancer (17), chronic lymphocytic leukemia (18) and colorectal cancer (33). In the present study, a significant
overexpression of MIAT was observed in NSCLC tissues compared with
that in adjacent normal lung tissues. In addition, the expression
level of MIAT in patients with NSCLC was significantly associated
with increased aggressiveness, a lower OS rate and advanced
clinical stage. Furthermore, the expression of MIAT in H1299-CDDP
cells was increased compared with that in H1299 cells. Knockdown
experiments identified the tumorigenic role of MIAT in promoting
cellular proliferation in NSCLC cells. Furthermore, the in
vivo assay demonstrated that the silencing of MIAT was able to
inhibit tumor growth and enhance the sensitivity of NSCLC to
CDDP.
A ceRNA hypothesis suggested that lncRNAs may
interact with miRs directly in order to regulate the expression of
target genes, and a novel regulatory network has been identified in
the crosstalk between lncRNAs and miRs (11). lncRNA MIAT has been recognized as a
sponge of miR-155-5p to promote breast cancer progression by
regulating dual specificity protein phosphatase expression
(17). Knockdown of MIAT was able to
inhibit breast cancer cell proliferation, migration and invasion by
sponging miR-155-5p (17).
Therefore, the present study used TargetScan to identify the
putative target miR of MIAT. Putative binding sites were identified
between MIAT and miR-184. The endogenous interaction between MIAT
and miR-184 was revealed by a dual-luciferase assay, and the
miR-184 expression levels were restored as MIAT was silenced. This
further verified that MIAT served as a ceRNA by sponging miR-184.
In addition, it was identified that SF1 is a target gene of the
MIAT/miR-184 axis in NSCLC cells using bioinformatics analysis,
which was subsequently demonstrated by dual-luciferase reporter
assays. SF1 is an RNA-binding protein, which is involved in the
spliceosome-mediated assembly of specific pre-mRNAs (13). Previous studies identified a
correlation between SF1 and cancer. For example, SF1 was able to
promote the tumorigenesis of testicular germ cell tumors (34,35). In
the present study, it was observed that the expression levels of
SF1 were regulated by MIAT by sponging miR-184 in NSCLC cells.
However, additional studies are required to examine the function
and molecular mechanism of SF1 in various types of cancer. The
in vivo analysis of the present study further validated that
depletion of MIAT was able to suppress tumor growth and enhance the
sensitivity of NSCLC to CDDP. These results suggested that
inhibiting the expression of MIAT may be a potential therapeutic
strategy to promote therapeutic activity and enhance CDDP
susceptibility in NSCLC.
In summary, overexpressed MIAT in CDDP-resistant
H1299 cells and NSCLC tissue specimens promoted proliferation and
enhanced resistance to CDDP in NSCLC cells in vitro and
in vivo, which suggests that MIAT exerts oncogenic
properties in NSCLC. The biological functional analysis
demonstrated that MIAT exerted its oncogenic effect partially by
upregulating the expression of SF1 as a miR-184 sponge. The results
of the present study provided novel insights into the molecular
signaling network involved in NSCLC carcinogenesis and CDDP
resistance, and demonstrated the potential therapeutic application
of MIAT in NSCLC progression.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Science and
Technology research project of the Department of Education of
Jiangxi Province (grant no. GJJ170099).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding authors on reasonable
request.
Authors' contributions
LW conceived and designed the study. CL acquired and
analyzed the data. LW and ZZ interpreted the data and wrote the
manuscript. ZZ critically revised the manuscript. All authors read
and approved the final version of the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Human Ethics
Committee of the First Affiliated Hospital of Gannan Medical
University. Written informed consent was obtained from all patients
prior to using their tissues for scientific research.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Youlden DR, Cramb SM and Baade PD: The
international epidemiology of lung cancer-Geographical distribution
and secular trends. J Thorac Oncol. 3:819–831. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Reck M, Heigener DF, Mok T, Soria JC and
Rabe KF: Management of non-small-cell lung cancer: Recent
developments. Lancet. 382:709–719. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Molina JR, Yang P, Cassivi SD, Schild SE
and Adjei AA: Non-small cell lung cancer: Epidemiology, risk
factors, treatment, and survivorship. Mayo Clin Proc. 83:584–594.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Duma N, Santana-Davila R and Molina JR:
Non-small cell lung cancer: Epidemiology, screening, diagnosis, and
treatment. Mayo Clin Proc. 94:1623–1640. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gottesman MM, Lavi O, Hall MD and Gillet
JP: Toward a better understanding of the complexity of cancer drug
resistance. Annu Rev Pharmacol Toxicol. 56:85–102. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fennell DA, Summers Y, Cadranel J, Benepal
T, Christoph DC, Lal R, Das M, Maxwell F, Visseren-Grul C and Ferry
D: Cisplatin in the modern era: The backbone of first-line
chemotherapy for non-small cell lung cancer. Cancer Treat Rev.
44:42–50. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Galluzzi L, Vitale I, Michels J, Brenner
C, Szabadkai G, Harel-Bellan A, Castedo M and Kroemer G: Systems
biology of cisplatin resistance: Past, present and future. Cell
Death Dis. 5:e12572014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Esteller M: Non-coding RNAs in human
disease. Nat Rev Genet. 12:861–874. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shi X, Sun M, Liu H, Yao Y and Song Y:
Long non-coding RNAs: A new frontier in the study of human
diseases. Cancer Lett. 339:159–166. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Prensner JR and Chinnaiyan AM: The
Emergence of lncRNAs in cancer biology. Cancer Discov. 1:391–407.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Anastasiadou E, Jacob LS and Slack FJ:
Non-coding RNA networks in cancer. Nat Rev Cancer. 18:5–18. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liz J and Esteller M: lncRNAs and
microRNAs with a role in cancer development. Biochim Biophys Acta.
1859:169–176. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ishii N, Ozaki K, Sato H, Mizuno H, Saito
S, Takahashi A, Miyamoto Y, Ikegawa S, Kamatani N, Hori M, et al:
Identification of a novel non-coding RNA, MIAT, that confers risk
of myocardial infarction. J Hum Genet. 51:1087–1099. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liao J, He Q, Li M, Chen Y, Liu Y and Wang
J: LncRNA MIAT: Myocardial infarction associated and more. Gene.
578:158–161. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sun C, Huang L, Li Z, Leng K, Xu Y, Jiang
X and Cui Y: Long non-coding RNA MIAT in development and disease: A
new player in an old game. J Biomed Sci. 25:232018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang R, Zhao L, Ji L, Bai L and Wen Q:
Myocardial infarction associated transcript (MIAT) promotes
papillary thyroid cancer progression via sponging miR-212. Biomed
Pharmacother. 118:1092982019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Luan T, Zhang X, Wang S, Song Y, Zhou S,
Lin J, An W, Yuan W, Yang Y, Cai H, et al: Long non-coding RNA MIAT
promotes breast cancer progression and functions as ceRNA to
regulate DUSP7 expression by sponging miR-155-5p. Oncotarget.
8:76153–76164. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sattari A, Siddiqui H, Moshiri F, Ngankeu
A, Nakamura T, Kipps TJ and Croce CM: Upregulation of long
noncoding RNA MIAT in aggressive form of chronic lymphocytic
leukemias. Oncotarget. 7:54174–54182. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lai IL, Yang CA, Lin PC, Chan WL, Lee YT,
Yen JC, Chang YS and Chang JG: Long noncoding RNA MIAT promotes
non-small cell lung cancer proliferation and metastasis through
MMP9 activation. Oncotarget. 8:98148–98162. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang HY, Zheng FS, Yang W and Lu JB: The
long non-coding RNA MIAT regulates zinc finger E-box binding
homeobox 1 expression by sponging miR-150 and promoteing cell
invasion in non-small-cell lung cancer. Gene. 633:61–65. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shirasawa M, Fukui T, Kusuhara S, Hiyoshi
Y, Ishihara M, Kasajima M, Nakahara Y, Otani S, Igawa S, et al:
Prognostic significance of the 8th edition of the TNM
classification for patients with extensive disease small cell lung
cancer. Cancer Manag Res. 10:6039–6047. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zeng A, Hua H, Liu L and Zhao J: Betulinic
acid induces apoptosis and inhibits metastasis of human colorectal
cancer cells in vitro and in vivo. Bioorg Med Chem. 27:2546–2552.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Faehling M, Achenbach J, Staib P, Steffen
U, Tessen HW, Gaillard VE and Brugger W: Erlotinib in routine
clinical practice for first-line maintenance therapy in patients
with advanced non-small cell lung cancer (NSCLC). J Cancer Res Clin
Oncol. 144:1375–1383. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tu L, Huang Q, Fu S and Liu D: Aberrantly
expressed long noncoding RNAs in hypertrophic scar fibroblasts in
vitro: A microarray study. Int J Mol Med. 41:1917–1930.
2018.PubMed/NCBI
|
|
26
|
Zhou RS, Zhang EX, Sun QF, Ye ZJ, Liu JW,
Zhou DH and Tang Y: Integrated analysis of lncRNA-miRNA-mRNA ceRNA
network in squamous cell carcinoma of tongue. BMC Cancer.
19:7792019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang CG, Yin DD, Sun SY and Han L: The
use of lncRNA analysis for stratification management of prognostic
risk in patients with NSCLC. Eur Rev Med Pharmacol Sci. 21:115–119.
2017.PubMed/NCBI
|
|
28
|
Zhu H, Yu J, Zhu H, Guo Y and Feng S:
Identification of key lncRNAs in colorectal cancer progression
based on associated protein-protein interaction analysis. World J
Surg Oncol. 15:1532017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lu KH, Li W, Liu XH, Sun M, Zhang Ml, Wu
WQ, Xie WP and Hou YY: Long non-coding RNA MEG3 inhibits NSCLC
cells proliferation and induces apoptosis by affecting p53
expression. BMC Cancer. 13:4612013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sun M, Liu XH, Lu KH, Nie FQ, Xia R, Kong
R, Yang JS, Xu TP, Liu YW, Zou YF, et al: EZH2-mediated epigenetic
suppression of long noncoding RNA SPRY4-IT1 promotes NSCLC cell
proliferation and metastasis by affecting the
epithelial-mesenchymal transition. Cell Death Dis. 5:e12982014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ma YJ, Ha CF, Bai ZM, Li HN, Xiong Y and
Jiang J: Overexpression of microRNA-205 predicts lymph node
metastasis and indicates an unfavorable prognosis in endometrial
cancer. Oncol Lett. 12:4403–4410. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pradhan AK, Emdad L, Das SK, Sarkar D and
Fisher PB: The enigma of miRNA regulation in cancer. Adv Cancer
Res. 135:25–52. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu Z, Wang H, Cai H, Hong Y, Li Y, Su D
and Fan Z: Long non-coding RNA MIAT promotes growth and metastasis
of colorectal cancer cells through regulation of miR-132/Derlin-1
pathway. Cancer Cell Int. 18:592018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhu R, Heaney J, Nadeau JH, Ali S and
Matin A: Deficiency of splicing Factor 1 suppresses the occurrence
of testicular germ cell tumors. Cancer Res. 70:7264–7272. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shitashige M, Naishiro Y, Idogawa M, Honda
K, Ono M, Hirohashi S and Yamada T: Involvement of Splicing
Factor-1 in beta-catenin/T-Cell Factor-4-mediated gene
transactivation and Pre-mRNA splicing. Gastroenterology.
132:1039–1054. 2007. View Article : Google Scholar : PubMed/NCBI
|