Introduction
Breast cancer has become a major public health
problem. Its incidence accounts for 7–10% of all types of malignant
tumors, and its incidence is often related to heredity (1). The breast is not an important organ for
maintaining human life, but free cancer cells can spread throughout
the body with blood or lymphs because of the loose connection
between breast cancer cells (2).
Furthermore, it can cause metastasis and endanger life. Metastasis
and spread of cancer cells are the main cause of increased
mortality in breast cancer patients (3). There are many related factors affecting
the prognosis of breast cancer, among which the main factors are
tumor invasion and pathological biological characteristics
(4). Therefore, the best way to
reduce breast cancer patient mortality is early detection and
treatment.
As important regulators, microRNAs (miRNAs) regulate
various biological processes by interacting with some target genes.
The dysregulation of miRNAs have been reported to exert different
effects in progression of breast cancer. For example, miR-144
suppressed proliferation, invasion, and migration of breast cancer
cells through inhibiting CEP55 (5).
Inversely, miR-221/222 targets adiponectin receptor 1 to promote
epithelial-to-mesenchymal transition (EMT) in breast cancer
(6). The dysregulation of
microRNA-431 (miR-431) in human diseases and cancers has attracted
attention. For instance, miR-431 restrained trophoblast migration
and invasion via targeting ZEB1 in preeclampsia (7). Moreover, miR-431 can function as a
chemosensitizer and potentiator of drug activity in adrenocortical
carcinoma (8) and miR-431 was found
to promote differentiation and regeneration of old skeletal muscle
by targeting Smad4 (9). In human
cancers, miR-431 usually acts as a tumor inhibitor through
regulating target genes. Liu et al found that downregulation
of miR-431 expression was associated with lymph node metastasis and
promoted cell invasion in papillary thyroid carcinoma (10). Yang et al (11) demonstrated that miR-431 inhibited
cell proliferation and induced cell apoptosis via targeting CDK14
in pancreatic cancer. However, the specific role of miR-431 remains
blurry and needs to be illuminated in breast cancer.
The fibroblast growth factor (FGF) family containing
18 related proteins can be involved in skeletal development and
homeostasis (12). As a member of
FGF family, fibroblast growth factor 9 (FGF9) was associated with
poor prognosis in patients with resected non-small cell lung cancer
(13). Moreover, the promoting
effects of FGF9 on cell proliferation and migration were identified
in human hepatocellular carcinoma (14). FGF9, as a target gene, has been found
to be mediated by some miRNAs. Li et al (15) proposed that miR-665 inhibited
vascular smooth muscle cell proliferation via targeting FGF9.
miR-140-5p suppressed tumor growth and metastasis by suppressing
FGF9 expression in hepatocellular carcinoma (16). However, the interaction between
miR-431 and FGF9 has not been reported in previous studies. Thus,
we investigated their relationship as well as the functions of
miR-431 in breast cancer progression. This study explored a novel
biomarker for diagnosis of breast cancer patients.
Materials and methods
Clinical tissues
Ninety-eight breast cancer patients in Jining No. 1
People's Hospital (Jining, China) participated in the study.
Informed consents were obtained from all breast cancer patients.
Patients with breast cancer did not receive any treatment except
for surgery. Permission for this study was acquired from the
Institutional Ethics Committee of Jining No. 1 People's
Hospital.
Cell culture and transfection
Human breast epithelial cell line MCF10A and breast
cancer cells MDA-MB-231 were from the Cell Bank of Chinese Academy
of Sciences (Shanghai, China). The growth conditions were 5%
CO2, at 37°C and culture solution (90% DMEM medium + 10%
FBS). Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific,
Inc.) was applied to transfer miR-431 mimics, miR-431 inhibitors,
FGF9 siRNA or FGF9 plasmid (GenePharma Co., Ltd.) into MDA-MB-231
cells.
RNA isolation and RT-qPCR
Total RNA isolation was performed using TRIZOL
reagent (Invitrogen; Thermo Fisher Scientific, Inc.). In addition,
cDNA solution was obtained using PrimeScript reverse transcription
kit (Qiagen, Inc.). RT-qPCR assay was performing using miScript
SYBR Green PCR kit (Qiagen, Inc.) based on the manufacturer's
instruction. U6 or GAPDH was used as the control of miR-431 or
FGF9, which were quantified with the 2-∆∆cq method. The
primers used in our work were as follows: miR-431, forward primer:
5′-CAGGCCGTCATGCAAA-3′, reverse primer:
5′-CGCTTCAGAATTTGCGTGTCAT-3′; U6, forward primer:
5′-CTCGCTTCGGCAGCACA-3′, reverse primer:
5′-AACGCTTCACGAATTTGCGT-3′; FGF9 forward primer:
5′-GGACTAAACGGCACCAGAAA-3′, reverse primer:
5′-CCATCCAAGCCTCCATCATA-3′; GAPDH forward,
5′-ACATCGCTCAGACACCATG-3′, reverse,
5′-TGTAGTTGAGGTCAATGAAGGG-3′.
MTT assay
Transfected MDA-MB-231 cells (2×103 cells/well) were
prepared in a 96-well plate. MDA-MB-231 cells were incubated for
24, 48, 72 or 96 h in DMEM medium. Next, 10 µl of MTT solution was
added to incubate the cells for 4 h. MTT solution was aspirated and
Formazan solution was added to fully dissolve the crystals. The
absorbance at 490 nm was examined by a microscope (Olympus Corp.,
Tokyo, Japan).
Transwell assay
The upper chamber was added with 60 µl of diluted
Matrigel to observe cell invasion. After 30 min, MDA-MB-231 cell
suspension (2×103 cells/well) was added to the Transwell upper
chamber. Next, 500 µl of DMEM medium (10% FBS) was added to 24-well
plates in the lower chamber. After 24 h, 0.1% crystal violet was
applied to stain the invaded cells. Cell migration experiment is
the same as the cell invasion experimental step except that
Matrigel is not used. Observation and photographing were performed
by light microscopy.
Western blot analysis
Protein samples were acquired by using RIPA lysis
buffer (Beyotime). Protein was separated by 10% SDS-PAGE. Protein
samples were transferred to PVDF membranes. Blocked with 5% non-fat
milk, protein samples were incubated overnight at 4°C with
E-cadherin, N-cadherin, vimentin, Bcl-2, Bax and GAPDH primary
antibodies (Abcam). After washing, protein samples were incubated
with secondary antibodies (Abcam) for 1 h. ECL kit (Beyotime) was
used to assess protein bands. In addition, protein was quantified
with Image Lab Software (Bio-Rad).
Dual luciferase reporter assay
The pmirGLO luciferase reporter vector (Promega
Corporation) was inserted with 3′-UTR of wt-FGF9 or Mut-FGF9. Then,
MDA-MB-231 cells with the luciferase vector and miR-431 mimics were
incubated for 48 h. Finally, luciferase activities were assessed by
dual-luciferase reporter assay system (Promega Corporation).
Statistical analysis
Data were analyzed and illustrated using SPSS 17.0
and Graphpad Prism 6, respectively, and shown as mean ± SD.
Differences were analyzed using Student's t-test or one-way ANOVA.
Univariate Kaplan-Meier method with log-rank test and χ2
test was used to analyze the association between miR-431 and
patient survival rate or clinical features. Differences were
considered significant at P<0.05.
Results
Dysregulation of miR-431 was
identified in breast cancer
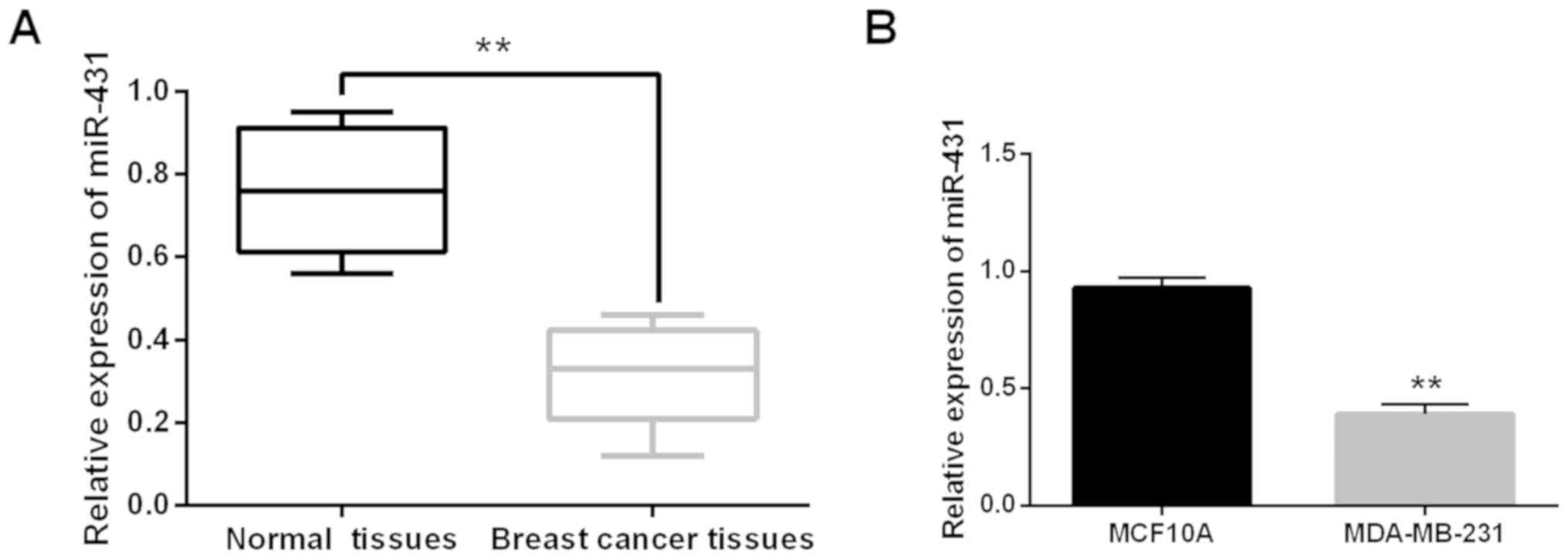
miR-431 expression was detected in breast cancer
tissues to explore dysregulation. It was found that miR-431 was
downregulated in breast cancer tissues compared to normal tissues
(P<0.01, Fig. 1A). Similarly,
miR-431 expression was lower in MDA-MB-231 cells than that in
MCF10A cells (P<0.01, Fig. 1B).
Correlation was identified between miR-431 expression and clinical
features in breast cancer patients. miR-431 expression was found to
be correlated with tumor size and lymph node metastasis (P<0.05,
Table I). The results indicated that
miR-431 may be dysregulated in breast cancer.
 | Table I.Relationship between miR-431
expression and the clinicopathological characteristics of breast
cancer patients. |
Table I.
Relationship between miR-431
expression and the clinicopathological characteristics of breast
cancer patients.
|
|
| miR-431 |
|
|---|
|
|
|
|
|
|---|
| Characteristics | Cases | High | Low | P-value |
|---|
| Age (years) |
|
|
| 0.07 |
| ≥50 | 40 | 16 | 24 |
|
|
<50 | 58 | 18 | 40 |
|
| Tumor size |
|
|
| 0.01a |
| <2
cm | 27 | 10 | 17 |
|
| ≥2
cm | 71 | 24 | 47 |
|
| Lymph node
metastasis |
|
|
| 0.005a |
| No | 28 | 9 | 19 |
|
| Yes | 70 | 25 | 45 |
|
| Her-2 status |
|
|
| 0.25 |
|
Positive | 52 | 18 | 34 |
|
|
Negative | 46 | 16 | 30 |
|
| ER status |
|
|
| 0.31 |
|
Positive | 45 | 15 | 30 |
|
|
Negative | 53 | 19 | 34 |
|
| PR status |
|
|
| 0.06 |
|
Positive | 36 | 16 | 20 |
|
|
Negative | 62 | 18 | 44 |
|
miR-431 restraines cell proliferation
and metastasis in breast cancer
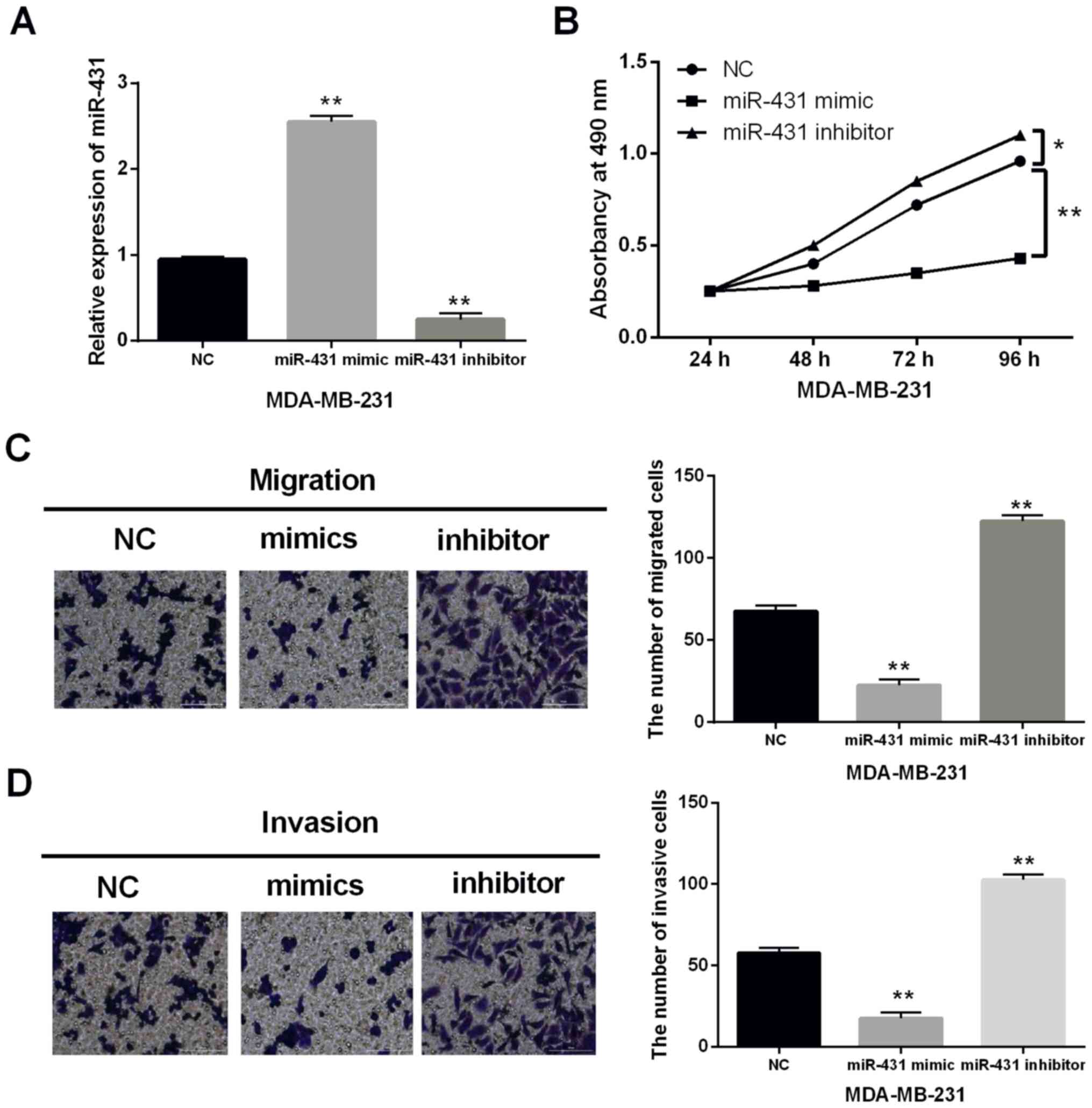
To further illuminate the role of miR-431 in breast
cancer, gain and loss functional experiment was performed in
MDA-MB-231 cells with miR-431 mimics or inhibitor. It was found
that miR-431 mimics enhanced the expression level in MDA-MB-231
cells, when miR-431 inhibitor decreased its expression (P<0.01,
Fig. 2A). MTT assay indicated that
MDA-MB-231 cell proliferation was repressed by miR-431 mimics and
accelerated by miR-431 inhibitor (P<0.01, Fig. 2B). Moreover, cell metastasis was
assessed by cell migration and invasion. We found that miR-431
mimics restrained cell migration and invasion, whereas miR-431
inhibitor facilitated cell metastasis in MDA-MB-231 cells
(P<0.01, Fig. 2C and D). Hence,
upregulation of miR-431 restrained cell proliferation and
metastasis in breast cancer.
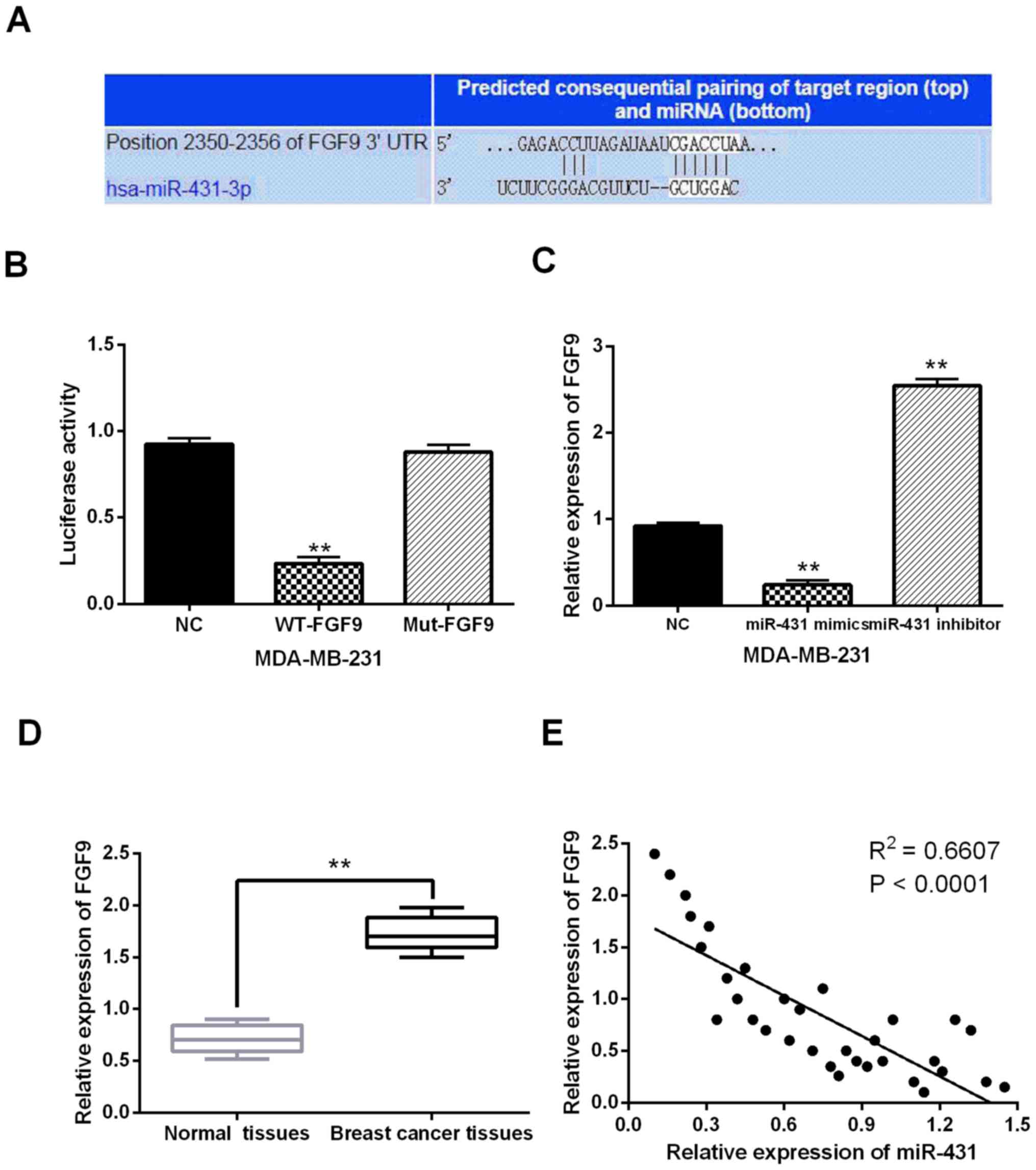
miR-431 directly targets FGF9
Targets of miR-431 were searched in TargetScan
database (http://www.targetscan.org/) to
explain its regulatory mechanism in breast cancer. It predicted
that miR-431 has a site binding to the 3′-UTR of FGF9 (Fig. 3A). Then, luciferase reporter assay
was conducted in MDA-MB-231 cells for verification. It showed that
miR-431 mimics reduced Wt-FGF9 luciferase activity, but did not
affect Mut-FGF9 luciferase activity (P<0.01, Fig. 3B). Moreover, we found that FGF9 was
downregulated by miR-431 mimics, and upregulated by miR-431
inhibitor in MDA-MB-231 cells (P<0.01, Fig. 3C). In addition, upregulation of FGF9
was identified in breast cancer tissues in contrast to normal
tissues (P<0.01, Fig. 3D).
Furthermore, a negative correlation between miR-431 and FGF9
expression was detected in breast cancer tissues (P<0.0001,
R2=0.6607; Fig. 3E). Collectively,
miR-431 directly targets FGF9 and the expression is inversely
regulated in breast cancer.
Upregulation of FGF9 impairs the
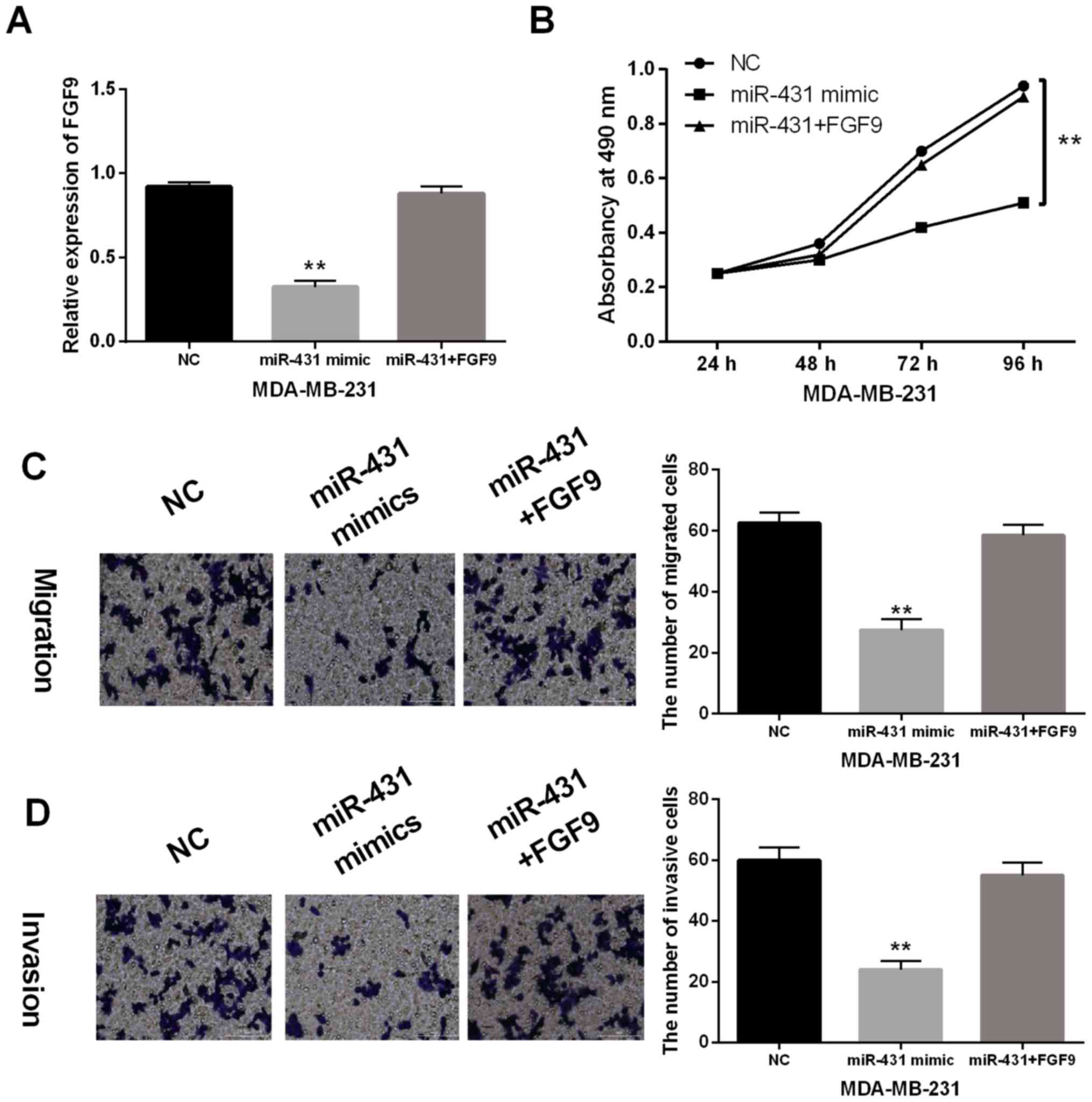
anti-tumor effect of miR-431 in breast cancer
FGF9 vector was transfected into MDA-MB-231 cells to
further explore their interaction in breast cancer. RT-qPCR
indicated that downregulation of FGF9 induced by miR-431 mimic was
recovered by FGF9 vector (Fig. 4A).
Similarly, the reverse effect of FGF9 on cell proliferation was
observed in MDA-MB-231 cells with miR-431 mimics (Fig. 4B). Moreover, the inhibitory effect of
miR-431 on MDA-MB-231 cell migration and invasion was impaired by
upregulation of FGF9 (Fig. 4C and
D). Based on these results, upregulation of FGF9 impaired the
anti-tumor effect of miR-431 in breast cancer.
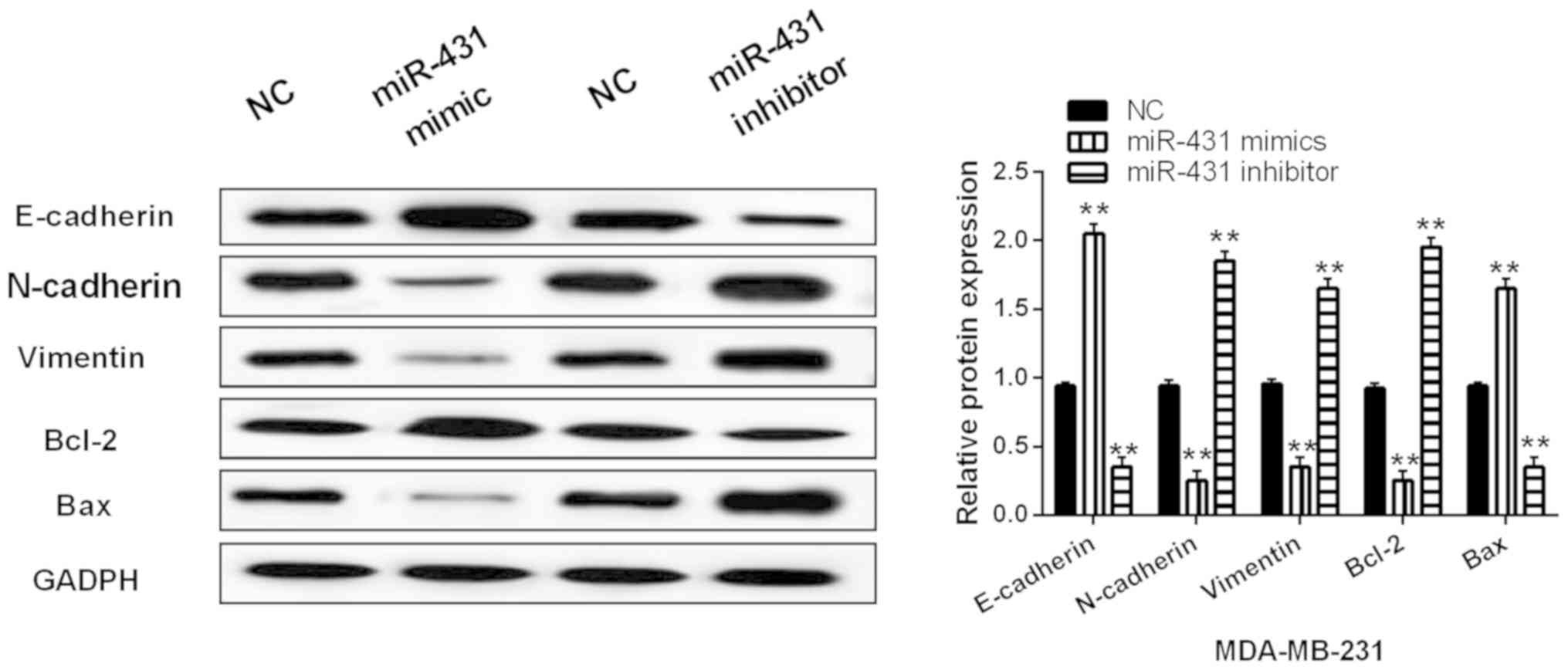
miR-431 hindered EMT and induced
apoptosis in breast cancers cells
Finally, expression of genes related to EMT and
apoptosis were assessed in MDA-MB-231 cells with miR-431 mimics or
inhibitor. miR-431 was found to regulate the expression of
E-cadherin, N-cadherin and vimentin. Upregulation of miR-431
promoted E-cadherin expression and suppressed expression of
N-cadherin and vimentin (P<0.01, Fig.
5). Downregulation of miR-431 showed the opposite effect on
expression of vimentin, N-cadherin and E-cadherin (P<0.01,
Fig. 5). Next, the expression of
apoptosis-associated proteins (Bcl-2/Bax) regulated by miR-431 was
detected in MDA-MB-231 cells. Bcl-2 expression was inhibited by
miR-431 overexpression and enhanced by knockdown of miR-431
(P<0.01, Fig. 5). Inversely,
miR-431 mimics promoted Bax expression. In addition, miR-431
inhibitor reduced Bax expression (P<0.01, Fig. 5). Thus, miR-431 hindered EMT and
induced apoptosis in breast cancers cells.
Discussion
Many miRNAs have been found to serve as tumor
promoter or suppressor in breast cancer, such as miR-29c and
miR-374a (17,18). In this study, miR-431 was found to
serve as a tumor inhibitor in breast cancer. In particular,
downregulation of miR-431 was identified in breast cancer, which
was associated with aggressive behavior. In addition, miR-431
restrained cell proliferation and induced apoptosis in breast
cancer. Furthermore, miR-431 blocked breast cancer metastasis and
EMT, and miR-431 directly targets FGF9. Moreover, upregulation of
FGF9 impaired the anti-tumor effect of miR-431 in breast cancer.
These results demonstrated that miR-431 was an important regulator
in breast cancer progression.
Consistent with our results, downregulation of
miR-431 was also found in lung cancer and hepatocellular carcinoma
(19,20). Furthermore, Pan et al
(21) showed that low miR-431
expression was related to lymph node metastasis and clinical TNM
stage in hepatocellular carcinoma. Similar results were also
identified in breast cancer. Functionally, it was reported that
miR-431 inhibited migration, invasion and EMT in hepatocellular
carcinoma cells by targeting ZEB1 (22). Moreover, miR-431 suppressed
proliferation and metastasis of lung cancer via down-regulating
DDX5 (23). The role of miR-431 in
breast cancer was the same as the above findings. In addition, we
also found that miR-431 induced breast cancer apoptosis through
enhancing Bax and repressing Bcl-2 expression. Similar results have
not been reported in previous studies. Moreover, previous studies
implied that miR-431 exerted effect in human diseases by mediating
certain target genes, including FOXA1 and UROC28 (24,25).
Here, FGF9 was confirmed to be a target of miR-431.
In the present study, FGF9 was upregulated in breast
cancer tissues. Furthermore, FGF9 had negative correlation with
miR-431 expression. Some other miRNAs were demonstrated to
negatively regulate FGF9 expression, such as miR-182 and miR-219a
(26,27). Moreover, it was reported that miR-26a
suppressed tumor growth and metastasis via targeting FGF9 in
gastric cancer (28). Liang et
al (29) reported that miR-187
repressed the proliferation of cervical cancer cells through
downregulation of FGF9. In addition, downregulation of miRNA-214
contributed to migration and invasion of gastric cancer cells
through targeting FGF9 and inducing EMT (30). Consistent with the above results,
miR-431 restrained cell viability and metastasis in breast cancer
through targeting FGF9. Furthermore, miR-431 blocked EMT and
induced apoptosis to play an inhibitory role in breast cancer.
In conclusion, downregulation of miR-431 was related
to aggressive behavior in breast cancer. Functionally, miR-431
restrained cell proliferation and metastasis in breast cancer.
Moreover, miR-431 hindered EMT and induced apoptosis in breast
cancer cells. In addition, miR-431 served as an inhibitor in breast
cancer through binding to FGF9. Although the role of miR-431 has
been illuminated in this study, the complex regulatory mechanisms
of miR-431 still need to be explored.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
WW wrote the manuscript, analyzed and interpreted
the patients' data. YD performed PCR and MTT assay. XL and YP were
responsible for Transwell assay, Western blot and Dual luciferase
reporter assay. JD and DL helped with statistical analysis. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
Jining No. 1 People's Hospital (Jining, China). Patients who
participated in this research had complete clinical data. Signed
informed consents were obtained from the patients or the
guardians.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jemal A, Center MM, DeSantis C and Ward
EM: Global patterns of cancer incidence and mortality rates and
trends. Cancer Epidemiol Biomarkers Prev. 19:1893–1907. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Strom C: Breast cancer: Diagnostic service
shares BRCA data. Nature. 522:342015. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhou M, Zhong L, Xu W, Sun Y, Zhang Z,
Zhao H, Yang L and Sun J: Discovery of potential prognostic long
non-coding RNA biomarkers for predicting the risk of tumor
recurrence of breast cancer patients. Sci Rep. 6:310382016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yin Y, Cai J, Meng F, Sui C and Jiang Y:
miR-144 suppresses proliferation, invasion, and migration of breast
cancer cells through inhibiting CEP55. Cancer Biol Ther.
19:306–315. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hwang MS, Yu N, Stinson SY, Yue P, Newman
RJ, Allan BB and Dornan D: miR-221/222 targets adiponectin receptor
1 to promote the epithelial-to-mesenchymal transition in breast
cancer. PLoS One. 8:e665022013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yang X and Meng T: MicroRNA-431 affects
trophoblast migration and invasion by targeting ZEB1 in
preeclampsia. Gene. 683:225–232. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kwok GTY, Zhao JT, Glover AR, Gill AJ,
Clifton-Bligh R, Robinson BG, Ip JCY and Sidhu SB: microRNA-431 as
a chemosensitizer and potentiator of drug activity in
adrenocortical carcinoma. Oncologist. 24:e241–e250. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lee KP, Shin YJ, Panda AC, Abdelmohsen K,
Kim JY, Lee SM, Bahn YJ, Choi JY, Kwon ES, Baek SJ, et al: miR-431
promotes differentiation and regeneration of old skeletal muscle by
targeting Smad4. Genes Dev. 29:1605–1617. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu Y, Li L, Liu Z, Yuan Q and Lu X:
Downregulation of miR-431 expression associated with lymph node
metastasis and promotes cell invasion in papillary thyroid
carcinoma. Cancer Biomark. 22:727–732. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang J, Zhu H, Jin Y and Song Y: miR-431
inhibits cell proliferation and induces cell apoptosis by targeting
CDK14 in pancreatic cancer. Eur Rev Med Pharmacol Sci.
22:4493–4499. 2018.PubMed/NCBI
|
|
12
|
Su N, Jin M and Chen L: Role of FGF/FGFR
signaling in skeletal development and homeostasis: Learning from
mouse models. Bone Res. 2:140032014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ohgino K, Soejima K, Yasuda H, Hayashi Y,
Hamamoto J, Naoki K, Arai D, Ishioka K, Sato T, Terai H, et al:
Expression of fibroblast growth factor 9 is associated with poor
prognosis in patients with resected non-small cell lung cancer.
Lung Cancer. 83:90–96. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang S, Lin H, Zhao T, Huang S, Fernig DG,
Xu N, Wu F, Zhou M, Jiang C and Tian H: Expression and purification
of an FGF9 fusion protein in E. coli, and the effects of the
FGF9 subfamily on human hepatocellular carcinoma cell proliferation
and migration. Appl Microbiol Biotechnol. 101:7823–7835. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li K, Pan J, Wang J, Liu F and Wang L:
miR-665 regulates VSMCs proliferation via targeting FGF9 and MEF2D
and modulating activities of Wnt/β-catenin signaling. Am J Transl
Res. 9:4402–4414. 2017.PubMed/NCBI
|
|
16
|
Yang H, Fang F, Chang R and Yang L:
MicroRNA-140-5p suppresses tumor growth and metastasis by targeting
transforming growth factor β receptor 1 and fibroblast growth
factor 9 in hepatocellular carcinoma. Hepatology. 58:205–217. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li W, Yi J, Zheng X, Liu S, Fu W, Ren L,
Li L, Hoon DSB, Wang J and Du G: miR-29c plays a suppressive role
in breast cancer by targeting the TIMP3/STAT1/FOXO1 pathway. Clin
Epigenetics. 10:642018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang J, He Y, Yu Y, Chen X, Cui G, Wang
W, Zhang X, Luo Y, Li J, Ren F, et al: Upregulation of miR-374a
promotes tumor metastasis and progression by downregulating LACTB
and predicts unfavorable prognosis in breast cancer. Cancer Med.
7:3351–3362. 2018. View Article : Google Scholar
|
|
19
|
Jiang Q, Cheng L, Ma D and Zhao Y:
FBXL19-AS1 exerts oncogenic function by sponging miR-431-5p to
regulate RAF1 expression in lung cancer. Biosci Rep. 39:392019.
View Article : Google Scholar
|
|
20
|
Li MF, Li YH, He YH, Wang Q, Zhang Y, Li
XF, Meng XM, Huang C and Li J: Emerging roles of hsa_circ_0005075
targeting miR-431 in the progress of HCC. Biomed Pharmacother.
99:848–858. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pan L, Ren F, Rong M, Dang Y, Luo Y, Luo D
and Chen G: Correlation between down-expression of miR-431 and
clinicopathological significance in HCC tissues. Clin Transl Oncol.
17:557–563. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sun K, Zeng T, Huang D, Liu Z, Huang S,
Liu J and Qu Z: MicroRNA-431 inhibits migration and invasion of
hepatocellular carcinoma cells by targeting the ZEB1-mediated
epithelial-mensenchymal transition. FEBS Open Bio. 5:900–907. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xu CM, Chen LX, Gao F, Zhu MF, Dai Y, Xu Y
and Qian WX: miR-431 suppresses proliferation and metastasis of
lung cancer via down-regulating DDX5. Eur Rev Med Pharmacol Sci.
23:699–707. 2019.PubMed/NCBI
|
|
24
|
Wu YZ, Chan KYY, Leung KT, Lam HS, Tam YH,
Lee KH, Li K and Ng PC: Dysregulation of miR-431 and target gene
FOXA1 in intestinal tissues of infants with necrotizing
enterocolitis. FASEB J. 33:5143–5152. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kong Q, Han J, Deng H, Wu F, Guo S and Ye
Z: miR-431-5p alters the epithelial-to-mesenchymal transition
markers by targeting UROC28 in hepatoma cells. OncoTargets Ther.
11:6489–6503. 2018. View Article : Google Scholar
|
|
26
|
Yu B, Qian T, Wang Y, Zhou S, Ding G, Ding
F and Gu X: miR-182 inhibits Schwann cell proliferation and
migration by targeting FGF9 and NTM, respectively at an early stage
following sciatic nerve injury. Nucleic Acids Res. 40:10356–10365.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rao C, Miao X, Zhao G, Zhang C, Shen H,
Dong C and Yang M: miR-219a-5p enhances cisplatin sensitivity of
human non-small cell lung cancer by targeting FGF9. Biomed
Pharmacother. 114:1086622019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Deng M, Tang HL, Lu XH, Liu MY, Lu XM, Gu
YX, Liu JF and He ZM: miR-26a suppresses tumor growth and
metastasis by targeting FGF9 in gastric cancer. PLoS One.
8:e726622013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liang H, Luo R, Chen X, Zhao Y and Tan A:
miR-187 inhibits the growth of cervical cancer cells by targeting
FGF9. Oncol Rep. 38:1977–1984. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang R, Sun Y, Yu W, Yan Y, Qiao M, Jiang
R, Guan W and Wang L: Downregulation of miRNA-214 in
cancer-associated fibroblasts contributes to migration and invasion
of gastric cancer cells through targeting FGF9 and inducing EMT. J
Exp Clin Cancer Res. 38:202019. View Article : Google Scholar : PubMed/NCBI
|