Introduction
Colorectal cancer (CRC) is the third most common
malignancy worldwide in 2012 (1).
The incidence, in developing countries, has increased by 2–4-fold
over the last two decades (2,3). Similar
to other malignancies, metastases are the primary cause of
CRC-associated mortality (4). The
discovery of new therapeutic strategies or potential co-therapeutic
agents against CRC will be of benefit to numerous patients.
Patients with this malignancy often experience depression due to
various stresses, which can aggravate clinical manifestations and,
as a result, affect disease progression, prognosis and outcome
(5,6). The inhibition of monoamine transmitter
reuptake by targeting monoamine transporters is currently the most
successful mechanism for treating depression (7). Epidemiological and animal studies
suggest that the use of antidepressants may be associated with
decreased risk of CRC (8–10). However, the mechanism underlying this
decreased risk remains elusive.
The norepinephrine transporter (NET) belongs to the
monoamine transporter class; monoamine transporters are located in
the neurons and glial cells of the central nervous system and in
tissues of the peripheral organs innervated by the sympathetic
ganglia, including the heart, adrenal medulla, gastrointestinal
tract and placenta (11). Upon
binding to its substrate, norepinephrine (NE), NET co-transports NE
along with a Na+ ion into the cytoplasm to maintain
noradrenergic transmission homeostasis, which is targeted in the
treatment of depression (12,13). NET
not only regulates the longevity of NE in the synapse but also
plays a role in presynaptic and postsynaptic homeostasis. Several
intracellular and extracellular signaling molecules can regulate
its function (12–14). In addition, loss or disruption of NET
function, caused by factors such as genetic polymorphisms, has been
epidemiologically demonstrated to be closely associated with
various neuropsychiatric diseases, cardiovascular diseases and
cancer (15–17).
The loss of cell adhesion is a key step in the
cascade leading to malignancy and metastasis. In numerous
instances, epithelial tumors lose epithelial (E)-cadherin-mediated
adhesions as they progress toward malignancy (18). Preoperatively elevated soluble
E-cadherin levels are a prediction marker of metastasis and a
pre-therapeutic prognostic marker for patients with CRC and hepatic
metastases (19,20). Notch signaling, a critical pathway in
tissue development, was also reported to contribute to
tumorigenesis and tumor metastasis (21). Furthermore, Notch signaling was
demonstrated to regulate E-cadherin expression in breast cancer,
hepatocarcinoma and CRC (22–24). The
loss of NET function was found to be associated with genetic
alterations in both neural crest cells and the adult superior
cervical ganglion and locus ceruleus; the inhibitor of Notch
signaling numb-like (Numbl) exhibited increased expression and the
Notch signaling pathway was subsequently inhibited in NET-deficient
mice (25). However, little is known
about the function of NET and its downstream mediators in
cancer.
In the present study, the significance of NET in CRC
was determined using publicly available CRC gene expression RNA
sequencing (RNAseq) datasets from The Cancer Genome Atlas (TCGA)
database and by immunohistochemistry assays. NET was highly
expressed and associated with CRC metastasis. Subsequently, the
effect of NET depletion was determined by transfection of human
colon cancer HCT116 and SW480 cells with NET-targeting small
interfering (si)RNA. Knockdown of NET resulted in decreased
invasion of CRC cells, inhibition of Notch1 activation and
increased E-cadherin expression. These findings revealed that NET
was highly expressed and closely associated with the invasion of
human CRC cells by influencing cell-cell adhesion through the
Notch1-E-cadherin pathway.
Materials and methods
Materials
Antibodies against human NET (cat nos. GTX47102 and
Ab41559) were purchased from GeneTex, Inc. and Abcam, respectively.
Anti-E-cadherin (cat. no. 20874-1-AP), anti-N-cadherin (cat. no.
22018-1-AP), anti-Notch1 (cat. no. 20687-1-AP) and anti-Snail1
(cat. no. 13099-1-AP) were purchased from Wuhan Sanying
Biotechnology, while anti-GAPDH (cat. co. Sc-32233) was obtained
from Santa Cruz Biotechnology, Inc.
Bioinformatics analysis
Data of NET expression in tumor tissues and
Tumor-Node-Metastasis (TNM) classification of 183 CRC cases were
extracted from the RNAseq Illumina HiSeq dataset in TCGA of the
University of California Santa Cruz Genomics Institute. The
datasets were downloaded from TCGA tools cancer browser (https://xenabrowser.net/hub/). As the previous reports
(26,27) demonstrated, the procedure of select
datasets (TCGA.COADREAD.sampleMap/HiSeqV2).
Clinical tissue specimens
Formalin-fixed paraffin-embedded tissue blocks of
CRC tumors and adjacent normal mucosal tissues were obtained from
35 patients between December 2017 and June 2018 at the Second
Affiliated Hospital of Xi'an Jiaotong University (Xi'an, China).
None of the patients received prior chemotherapy, radiotherapy or
systemic therapies, or had additional malignant tumors. The
clinicopathological parameters of the patients are presented in
Table SI. There were 15 cases with
metastasis, including lymph node metastasis and distant metastasis,
and 20 cases without metastasis. All samples were obtained with the
informed consent of each patient before collection. The present
study was approved by the Medical Ethics Committee of Xi'an
Jiaotong University (approval no. 2017-488).
Tissue immunohistochemistry
Immunohistochemical detection of NET was performed
as previously described (28).
Anti-NET antibody was used at 1:100 dilution and incubated
overnight at 4°C; the human Biotin-Streptavidin HRP Detection kit
(cat. no. SP-9001; ZSGB-BIO) including the anti-rabbit secondary
antibody (incubation for 30 min at room temperature) and DAB
chromogenic agents (incubation for 15 min at room temperature) were
used according to the manufacturer's protocol. All samples were
counterstained with hematoxylin for 30 sec at room temperature.
NET positivity appeared as brown or yellow staining
in the plasma membrane or cytoplasm. Scores by two independent
investigators were averaged for evaluation of NET expression. The
proportion of positive cells was scored as follows: 0, no positive
cells; 1, <10% positive cells; 2, 10–50% positive cells; 3,
50–80% positive cells; and 4, ≥80% positive cells. Staining
intensity was rated as follows: 0, no staining; 1, light yellow and
weak staining; 2, yellow and moderate staining; and 3, brown and
strong staining. The product of staining intensity and proportion
of positive cells comprised the staining index (SI). The ratio of
NET SI of CRC tumor tissues to NET SI of adjacent normal mucosal
tissues was used to evaluate differences in NET expression between
patients with metastatic and non-metastatic CRC.
Cell lines and cell culture
Human colorectal cancer cells (HCT116, RKO, HT29,
SW480, SW620 and CaCo-2), purchased from the American Type Culture
Collection (ATCC), were kindly provided by Professor Hiroyuki
Kuwano (Graduate School of Medicine, Gunma University, Maebashi,
Japan). The normal colonic epithelial cell line NCM460 was
purchased from the ATCC. The cells were cultured in RPMI-1640
medium (Hyclone; GE Healthcare Sciences) supplemented with 10%
fetal bovine serum (FBS; Biological Industries) at 37°C in a
humidified atmosphere of 95% air and 5% CO2, with medium
change every 2 days. Cells in the mid-log phase were used for the
experiments in the present study.
siRNA synthesis and transfection
Human NET siRNAs (siNET1 sense,
5′-GAUUUCGUGACUGUAGUUU-3′; siNET1 antisense,
5′-AAACUACAGUCACGAAAUC-3′; siNET2 sense, 5′-GGAGAAGGAGAGCUACCAA-3′;
and siNET2 antisense, 5′-UUGGUAGCUCUCCUUCUCC-3′) and negative
control siRNA (siNC sense, 5′-UUCUCCGAACGUGUCACGU-3′; and siNC
antisense, 5′-ACGUGACACGUUCGGAGAA-3′) were chemically synthesized
by Shanghai Jima Industrial, Co., Ltd. Lipofectamine®
2000 (Invitrogen; Thermo Fisher Scientific, Inc.) was used to
transfect the cells. The siRNAs were diluted to 30 nM in the wells
plated with HCT116 or SW480 cells. Following incubation for 12 h,
cells were incubated with normal medium to observe changes in cell
viability or harvested to be seeded into a 24-well cell culture
Transwell insert system (8 µm pore size; Becton Dickinson and
Company) for observing changes in cell invasion. The transfection
efficiency was determined using reverse transcription-quantitative
(RT-q)PCR and western blot analysis.
RNA isolation and RT-qPCR
Total cellular RNA from the human colorectal cancer
cells was extracted using TRIzol® reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions. mRNA was reverse transcribed into complementary DNA
(cDNA) (at 37°C for 30 min and 85°C for 5 min) using the
PrimeScript RT Reagent kit (Takara Bio, Inc.). qPCR was performed
using the FastStart Essential DNA Green Master kit (Roche
Diagnostics) on a LightCycler®480 II instrument,
according to the manufacturer's instructions: 95°C for 10 min; 95°C
for 15 sec, 60°C for 1 min, 72°C for 30 sec (40 cycles); and 94°C
for 90 sec, 60°C for 3 min and 94°C for 10 sec. The relative
expression of NET to β-actin (internal control) was calculated
using the 2−ΔΔCq method (29). The primers were synthesized by
Tsingke Biotech, and the sequences were as follows: NET forward,
5′-GCGCTCATCCCAGTGTCTAA-3′; NET reverse, 5′-GGATCAAGAAGGCACCGCC-3′;
β-actin forward, 5′-CCAGAGGCGTACAGGGATAG-3′; and β-actin reverse,
5′-CCAACCGCGAGAAGATGA-3′. All reactions were performed in
triplicate.
Cell invasion assay
Cell invasion was examined in a 24-well cell culture
Transwell insert system. The chambers were coated with Matrigel for
determination of cell invasive ability. The lower chamber was
filled with 600 µl RPMI-1640 medium containing 10% FBS, and the
upper chamber was filled with 200 µl RPMI-1640 medium without FBS.
HCT116 (6×104) and SW480 (10×104) cells were
seeded onto the upper chamber. Following incubation for 24 h, the
remaining cells in the upper chamber were completely removed using
a cotton swab, and the invading cells on the bottom surface were
fixed with 4% polyformaldehyde for 15 min at room temperature and
stained with 0.1% crystal violet for 30 min at room temperature.
Digital images were obtained using a photomicroscope at ×100
magnification (Nikon Corporation). In addition, stained cells on
the lower surface of the chamber were dissolved with 150 µl DMSO
and quantified by measuring the absorbance at 590 nm using a
microplate reader (BMG Labtech GmbH).
Cell viability assay
Cells (HCT116, 3×103 cells/well; and
SW480, 3.5×103 cells/well) were seeded onto a 96-well
plate. Following incubation for 24 h, MTT (0.5 mg/ml) was added and
incubated at 37°C for 4 h, and the absorbance of each well was
determined spectrophotometrically at 570 nm using the FLUOstar
OPTIMA (BMG Labtech GmbH). All samples were evaluated in
quintuplicate.
Western blot analysis
Cells were harvested from culture dishes and lysed
in RIPA buffer supplemented with protease inhibitors and
phosphatase inhibitors (Invitrogen; Thermo Fisher Scientific,
Inc.). The protein concentration was determined using the Pierce
BCA Protein Assay Kit (Thermo Fisher Scientific, Inc.). The cell
lysates (20 µg/lane) were separated by SDS-PAGE on a 10%
polyacrylamide gel and transferred to PVDF membranes for blotting.
The membranes were blocked in 5% skim milk for 30 min at room
temperature and incubated with primary antibodies (all 1:1,000) at
4°C overnight. An antibody against GAPDH was used as the loading
control. This was followed by incubation with horseradish
peroxidase-conjugated IgG anti-rabbit (cat. no. 111-035-144) and
anti-mouse (cat. no. 115-035-146) secondary antibodies (Jackson
ImmunoResearch Laboratories, Inc.) at 1:5,000 dilution for 1 h. The
immunoreactive bands were visualized by enhanced chemiluminescence
(EMD Millipore). The band intensity was measured by densitometry
and quantified using the Gel Plotting Macros of NIH Image 1.62
software (National Institutes of Health). The relative expression
ratio of NET to GAPDH was calculated and normalized to the siNC
samples on the same membrane.
Statistical analysis
All experiments were performed with three
independent replicates. The data were expressed as the mean ±
standard deviation and analyzed with SPSS 17.0 software (SPSS,
Inc.). The Kruskal-Wallis and least significance difference (LSD)
post hoc test were used to analyze the expression of NET among
patients with CRC exhibiting different T, N and M. One-way ANOVA
followed by Tamhane's T2 post hoc test was used to determine the
associations between NET expression level and clinical
characteristics of patients with CRC. P<0.05 was considered to
indicate a statistically significant difference.
Results
NET is highly expressed in CRC tissues
with metastasis and in human colon cancer cells
In order to determine the significance of NET in
CRC, publicly available CRC gene expression RNAseq datasets from
TCGA database were analyzed. NET expression was not associated with
tumor topography (Fig. 1A); however,
it was significantly associated with lymphatic metastasis
(P<0.05; Fig. 1B), distant
metastasis (P<0.05; Fig. 1C) and
clinical stages (P<0.05; Fig. 1D)
in patients with CRC (30). The
expression of NET was further examined in 35 CRC tissues and
adjacent normal tissues (Fig. 2A).
In tumor tissues of patients with metastatic CRC, the expression of
NET was significantly higher than in adjacent normal tissues, and
its fold increase was higher than that of patients with
non-metastatic CRC. The protein levels of NET in CRC HCT116, RKO,
HT29, SW480, SW620 and CaCo-2 cells were significantly higher than
those in normal colonic epithelial NCM460 cells (P<0.001;
Fig. 2B). These findings indicate
that NET is highly expressed in CRC and is associated with CRC
metastasis.
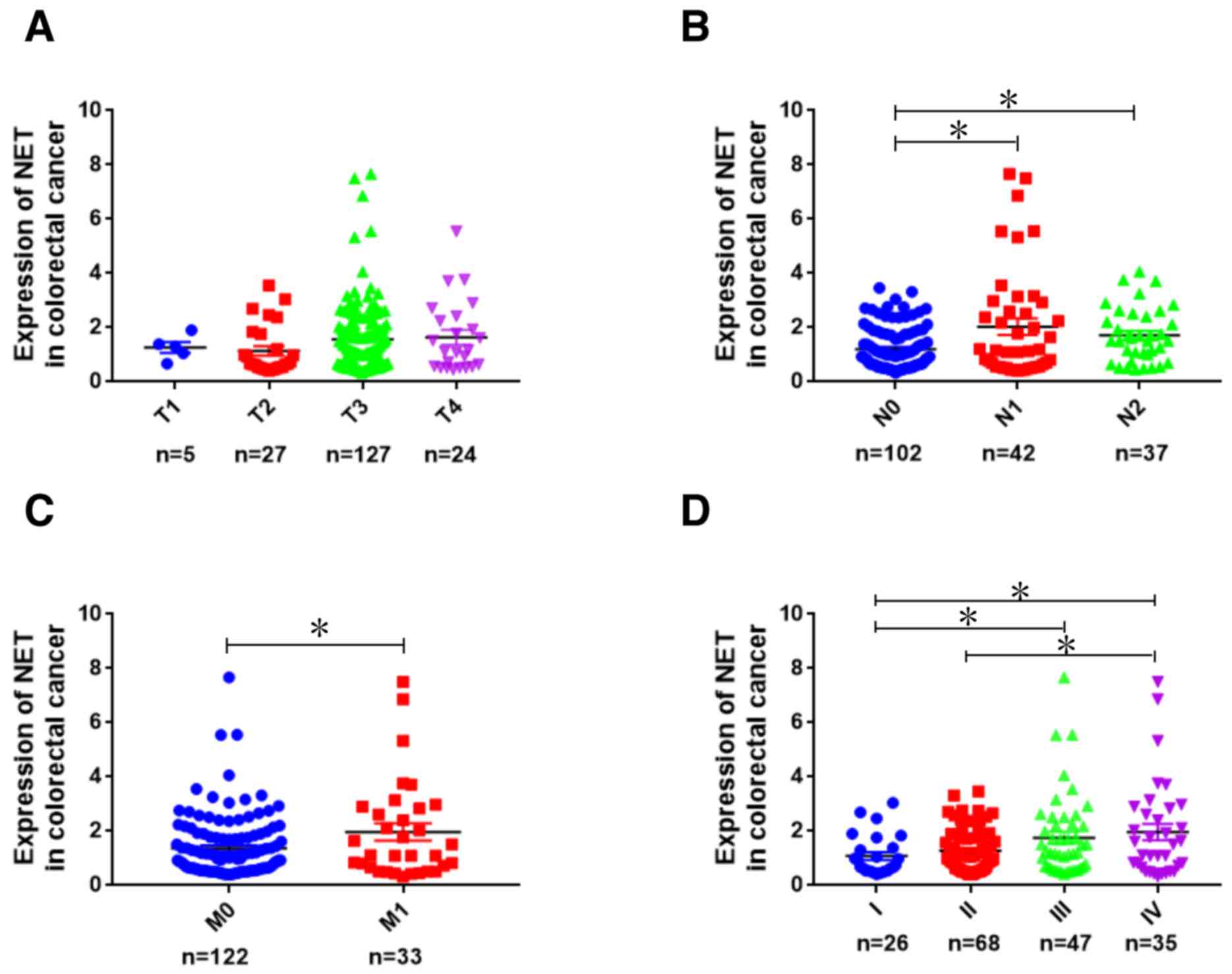 | Figure 1.Analysis of NET expression in human
CRC using TCGA database. Using publicly available CRC gene
expression RNAseq datasets in TCGA database, the expression
patterns of NET were analyzed bioinformatically and shown according
to the grade of (A) topography (T1, T2, T3 and T4), (B) lymph node
metastasis (N0, N1 and N2), (C) distant metastasis (M0 and M1) and
(D) clinical stages (I, II, III and IV) of patients with CRC.
Kruskal-Wallis test followed by least significance difference post
hoc test was used to analyze NET expression among different T, N, M
and clinical stage groups. *P<0.05. TCGA, The Cancer Genome
Atlas; NET, norepinephrine transporter; CRC, colorectal cancer. |
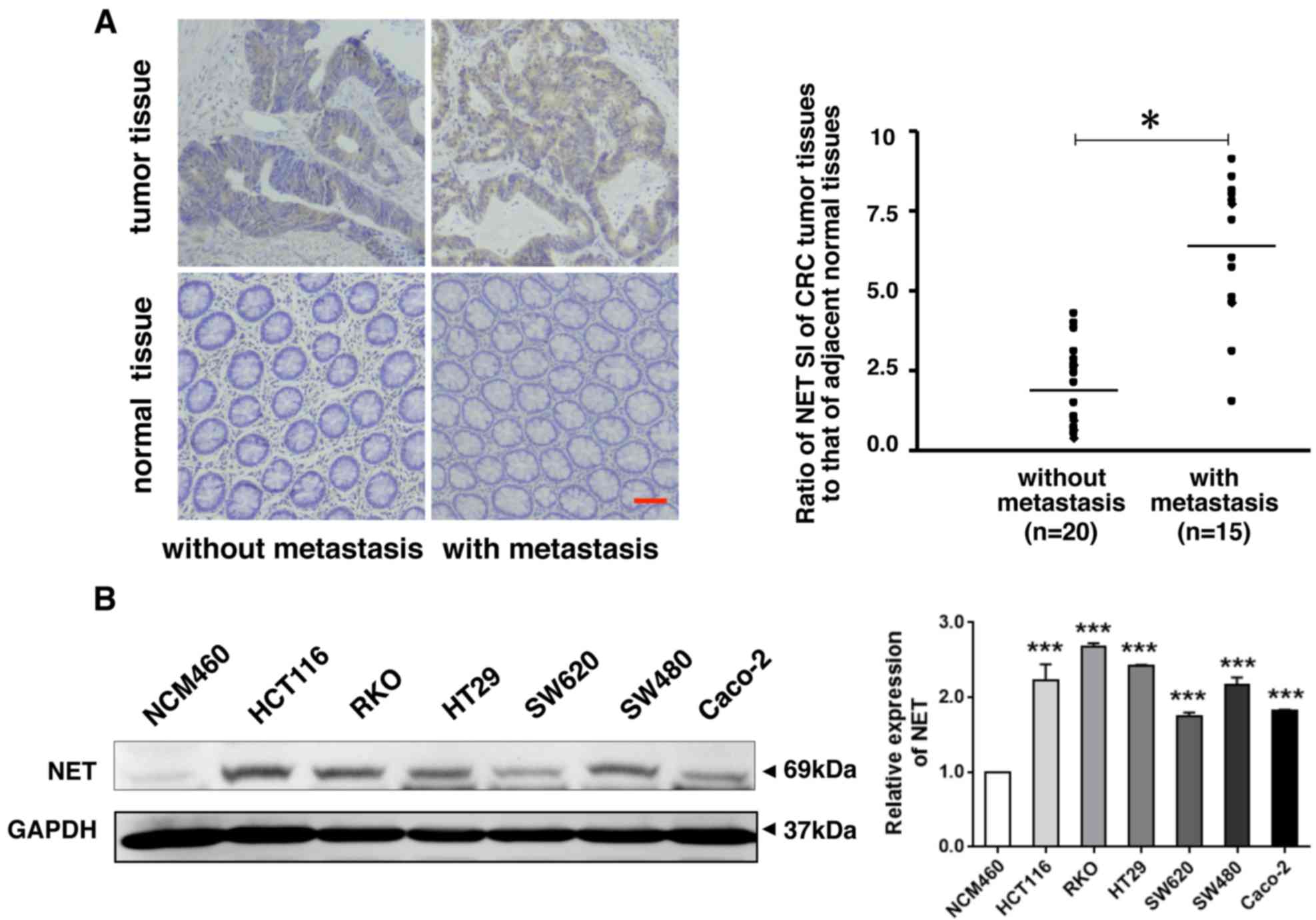 | Figure 2.NET is highly expressed in human CRC
with metastasis and in human CRC cells. (A) Clinical CRC tissues
and adjacent normal tissues were collected and used for paraffin
sections. NET immunohistochemistry was performed, which revealed
increased expression of NET in cancer tissues of patients with
metastatic CRC (scale bar, 50 µm). The proportion and intensity of
staining were evaluated and used to calculate the staining index
(SI). The ratio of NET SI of CRC tumor tissues to NET SI of
adjacent normal mucosal tissues was used to evaluate differences in
NET expression between patients with metastatic and non-metastatic
CRC. One-way ANOVA followed by Tamhane's T2 post hoc test was used
to compare the expression of NET between tumor tissues of patients
with CRC with metastasis and that of those without metastasis. (B)
Lysates of human CRC cell lines (HCT116, RKO, HT29, SW620, SW480
and CaCo-2) and a normal colonic epithelial cell line, NCM460, were
harvested. Western blotting revealed high expression of NET in
human CRC cells (left). The band intensities were quantified, and
the relative expression of NET to GAPDH was calculated and
normalized to the NCM460 cell line. One-way ANOVA and least
significance differenceposthoc test were used to conduct multiple
comparisons between the NET expression levels in human CRC cell
lines and in NCM460 cells. *P<0.05, ***P<0.001. CRC,
colorectal cancer; NET, norepinephrine transporter. |
Knockdown of NET inhibits the invasive
capability of human colon cancer cells
In order to determine the role of high NET
expression in CRC metastasis, two siRNAs specifically targeting
human NET (siNET1 and siNET2) and a negative control (siNC) were
synthesized (Table I). Due to the
high expression level of NET in human colon cancer HCT116 and SW480
cells (Fig. 2B), these cells were
selected for use in the subsequent experiments. HCT116 and SW480
cells were transfected with siNET1, siNET2 or siNC, and total RNA
and protein lysates were extracted at 48 h post-transfection.
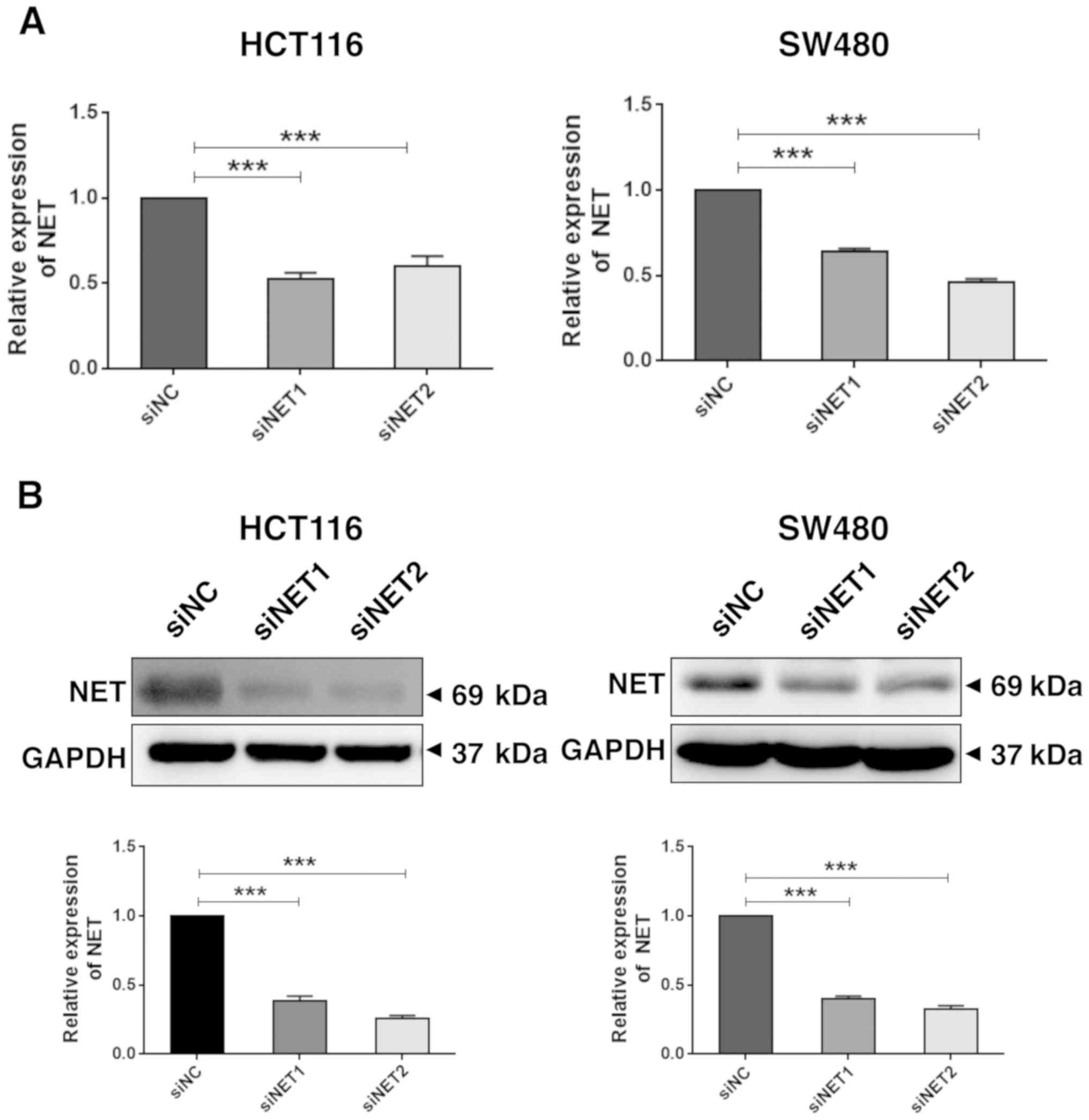
RT-qPCR and western blot analysis revealed significantly decreased
mRNA (P<0.001; Fig. 3A) and
protein (P<0.001; Fig. 3B)
expression levels of NET in HCT116 and SW480 cells transfected with
both siNET1 and siNET2, compared with those observed in cells
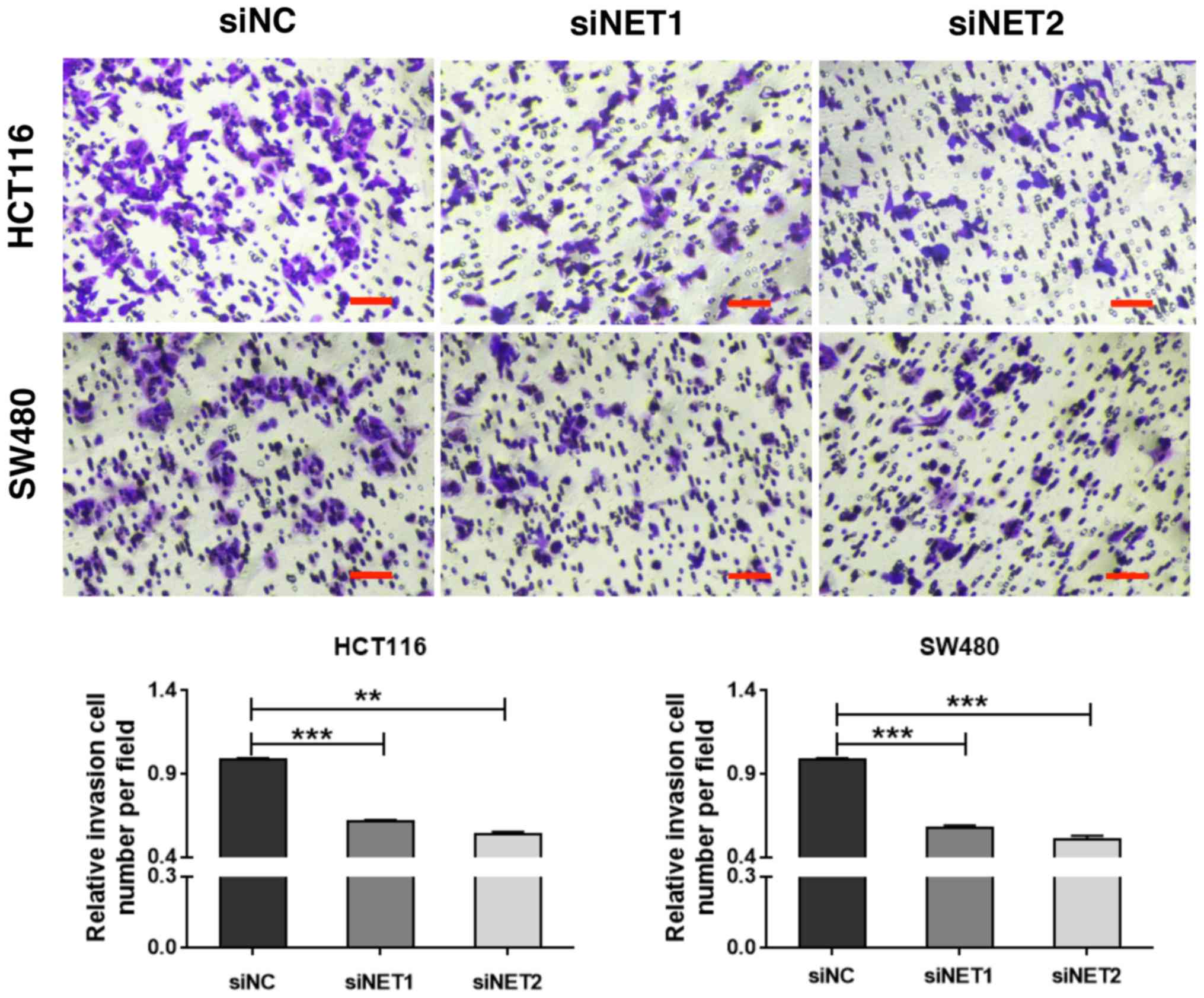
transfected with siNC. Subsequently, the effect of NET knockdown on
cell invasion was examined. As shown in Fig. 4, the Transwell assay demonstrated
significantly suppressed invasive capability of siNET1- and
siNET2-transfected HCT116 and SW480 cells compared with that of
cells transfected with siNC; with decreases of 38.1 and 46.0%
observed for HCT116 cells, and 42.3 and 49.0% observed for SW480
cells, respectively. In addition, there were no significant
differences in cell viability between the siNET and siNC groups
(Fig. S1). These findings indicate
that the depletion of NET inhibits the invasive capabilities of
human colon cancer cells.
 | Table I.Sequences of primers and
oligonucleotides used in the present study. |
Table I.
Sequences of primers and
oligonucleotides used in the present study.
| Name | Sequence (5′ to
3′) |
|---|
| siNET1 sense |
GAUUUCGUGACUGUAGUUUTT |
| siNET1
antisense |
AAACUACAGUCACGAAAUCTT |
| siNET2 sense |
GGAGAAGGAGAGCUACCAATT |
| siNET2
antisense |
UUGGUAGCUCUCCUUCUCCTT |
| siNC sense |
UUCUCCGAACGUGUCACGUTT |
| siNC antisense |
ACGUGACACGUUCGGAGAATT |
| NET forward |
GCGCTCATCCCAGTGTCTAA |
| NET reverse |
GGATCAAGAAGGCACCGCC |
| β-actin
forward |
CCAGAGGCGTACAGGGATAG |
| β-actin
reverse |
CCAACCGCGAGAAGATGA |
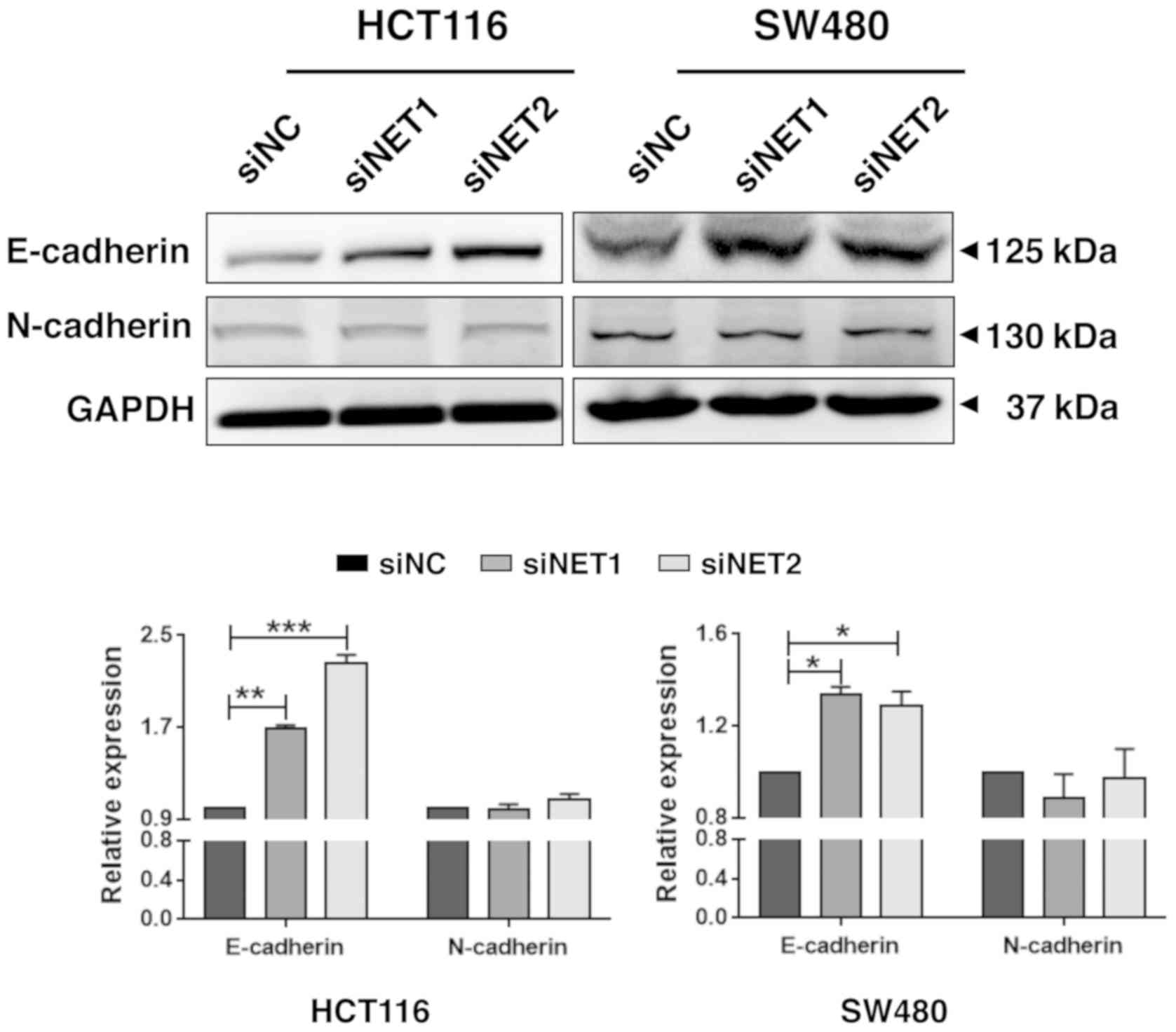
Knockdown of NET increases E-cadherin
and inhibits Notch1 signaling activity
E-cadherin is an important cell-cell adhesion
molecule. Several studies indicated that expression of E-cadherin
was negatively correlated with the degree of differentiation,
invasiveness and metastasis of malignant tumors, and positively
correlated with prognosis (18–20).
Thus, the changes in the expression of E-cadherin were determined
following the knockdown of NET in HCT116 and SW480 cells. As shown
in Fig. 5, although there were no
significant changes in N-cadherin protein level, the protein level
of E-cadherin was significantly increased in HCT116 and SW480 cells
transfected with siNET1 (P<0.01 and P<0.05, respectively) and
siNET2 (P<0.001 and P<0.05, respectively) compared with that
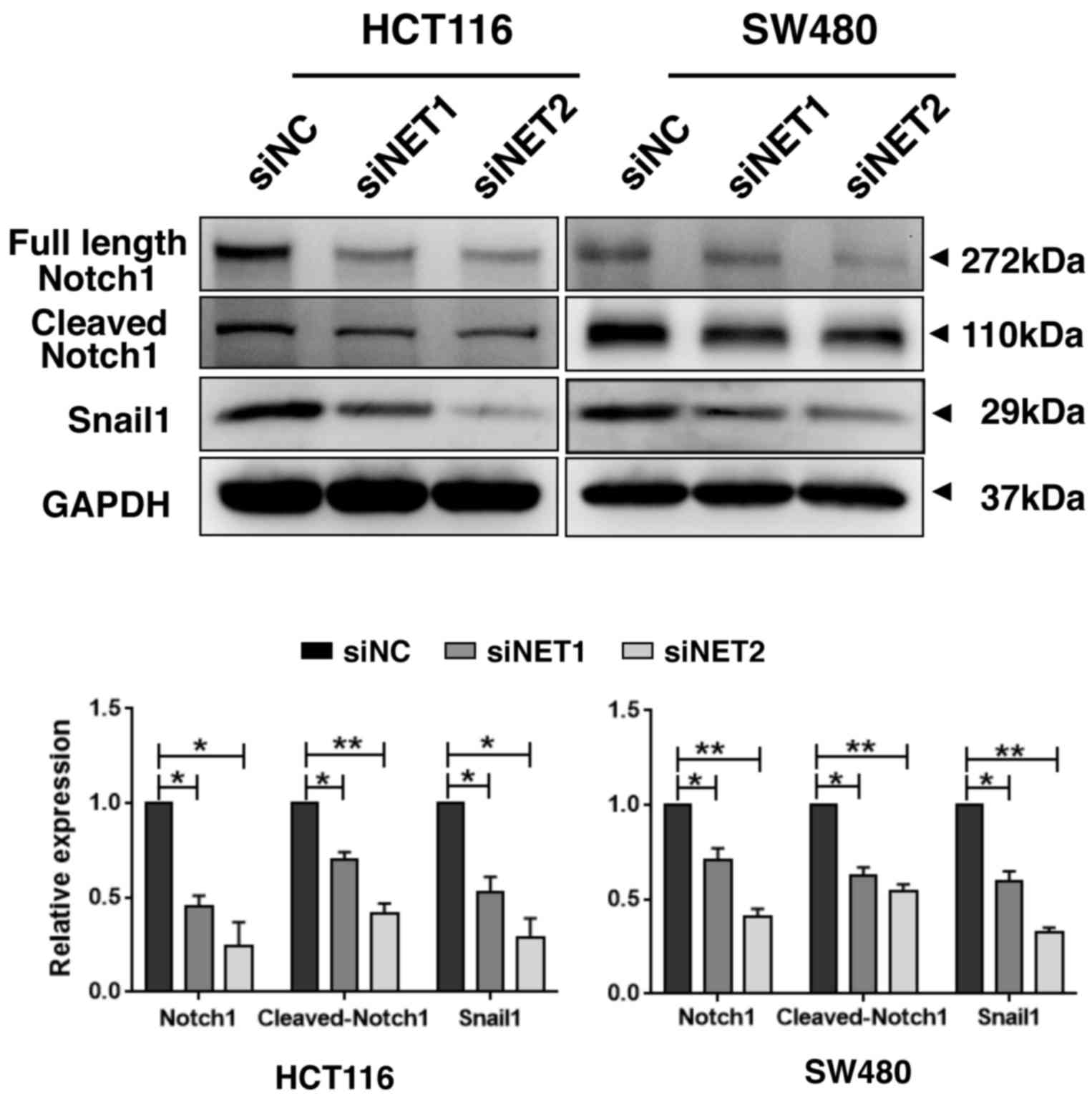
observed in cells transfected with siNC. Moreover, compared with
those of the siNC group, the expression levels of full length
Notch1, cleaved Notch1 and the downstream transcription factor
Snail1 (31) were all significantly
decreased in both HCT116 and SW480 cells transfected with siNET1 or
siNET2, indicating Notch1 signaling inhibition in NET-depleted
human CRC cells (Fig. 6). In
addition to the key roles during the normal development of multiple
tissues and organs, Notch signaling was shown to be associated with
the occurrence and development of invasion and metastasis in
various types of cancer. Chen et al (22) revealed that hypoxia-mediated Notch
signaling may have an important role in the initiation of
epithelial-mesenchymal transition and possess subsequent potential
for breast cancer metastasis. Wang et al (23) demonstrated that abnormal Notch1
expression is strongly associated with metastatic hepatocellular
carcinoma, which may be mediated through the
Notch1-Snail1-E-cadherin signaling pathway. Vinson et al
summarized that Notch1 signaling regulates the formation and
maintenance of colorectal cancer stem cells, which lead to
metastasis and tumorigenesis (21–23,31).
Furthermore, Notch signaling was demonstrated to regulate
E-cadherin expression in several types of cancer, including in CRC
cells, and Notch1-Hairy enhancer of Split-1 (HES1)-E-cadherin was
shown to promote invasiveness and metastasis, and was associated
with poor survival (24). Combined
with the findings of the present study, it is speculated that the
depletion of NET results in the inhibition of Notch1 signaling,
increases E-cadherin expression and decreases the invasive
capability of human colon cancer cells.
Discussion
Epidemiological and in vivo studies suggested
that the use of antidepressants was correlated with decreased risk
of CRC (8–10). However, the mechanism underlying this
decreased risk remains elusive. NET, a target of antidepressants,
is distributed within neurons, glial cells and peripheral
sympathetic nerve fibers that innervate tissue organs, including
the gastrointestinal tract. The loss or disruption of NET function
was shown to be associated with several neuropsychiatric diseases
and tumors, for which the underlying mechanisms are unknown.
Studies focusing on the SNP 1287 G/A (rs5569), located in exon 10
of hNET, have demonstrated an association with depression,
attention-deficit/hyperactivity disorder, personality traits,
alcohol dependence, panic disorder, schizophrenia, and bipolar
disorder. Höpfner et al (15)
revealed that changes of hNET level can influence the effect of
meta-iodobenzylguanidine on neuroendocrine gastrointestinal tumors
(15–17,32). The
present study revealed that NET was highly expressed in CRC tissues
with metastasis, compared with that found in adjacent normal
tissues, and its fold increase was higher than that of patients
with non-metastatic CRC. The knockdown of NET resulted in the
inhibition of the invasive capability of human colon cancer cells.
In addition, E-cadherin expression increased and Notch1 signaling
was inhibited upon knockdown of NET in colon cancer cells. These
results suggest that high expression of NET in CRC is associated
with the metastasis of human colon cancer cells by influencing
cell-cell adhesion via the Notch1-E-cadherin pathway.
The activation of the β-adrenergic system was
demonstrated to contribute primarily to stress-associated
acceleration of cancer progression (33). NE is the principal chemical messenger
employed in central noradrenergic and peripheral sympathetic
synapses. Inhibitors of monoamine transmitter reuptake are widely
used as antidepressants in clinical practice and are reportedly
associated with a decreased risk of CRC (8–10). In
addition to their actions against depression, these antidepressants
also inhibit tumor cell viability (9,10,34,35).
The antidepressant fluoxetine was demonstrated to have a direct
anti proliferative effect on human colon cancer cells in
vitro. Moreover, changes in cell viability, cell cycle,
apoptosis and NF-κB signaling induced by fluoxetine were also
demonstrated. However, the molecular targets associated with this
antidepressant were not explored; a series of future experiments
will be based on the present findings with the aim of clarifying
them and provide evidence for their practice. NET is an important
target of antidepressants. Genetic changes or drug interventions
are frequently reported to cause several diseases, including
cancer. The present study demonstrated high expression of NET to be
associated with human colon cancer cell invasion and metastasis. To
the best of our knowledge, this is the first study to investigate
the target of monoamine transmitter reuptake inhibitors, NET, with
respect to its function in human colon cancer cells. However, in
vivo studies are required to validate the findings of the
present study. NET-targeting short hairpin RNA are currently being
constructed, which will be used to knockdown NET in mice for future
in vivo studies.
The loss of cell adhesion is a key step in the
development of malignancy and metastasis. E-cadherin is a
prognostic indicator in metastasis, and its decreased expression
was correlated with enhanced metastasis in several malignancies
(18–20). In the present study, increased
expression of E-cadherin was detected in human colon cancer cells
upon the knockdown of NET, which may explain the suppressed
invasive capabilities of NET-depleted colon cancer cells. Notch
signaling, a critical pathway for tissue development, was also
shown to be associated with tumorigenesis and tumor metastasis
(21). After extracellular
activation by Jagged 1, the transmembrane protein Notch is cleaved
by γ-secretase, and the cleaved intracellular domain of Notch
activates associated transcription factors to enhance tumor cell
metastasis (36). Moreover, Notch
signaling regulates E-cadherin expression, and in CRC cells,
Notch1-HES1-E-cadherin was demonstrated to promote invasiveness and
metastasis (24). The present study
also demonstrated the inhibition of Notch1 signaling activity upon
knockdown of NET in human colon cancer cells. This result is
remarkable in light of a previous report that demonstrated NET
function to be involved in noradrenergic cell differentiation; the
expression of the Notch signaling inhibitor Numbl was increased,
and Notch signaling activity was subsequently inhibited, in both
neural crest cells and adult superior cervical ganglion and locus
ceruleus in NET-deficient mice (25). NET was highly expressed and
associated with metastasis in human CRC tissues, and the depletion
of NET inhibited Notch signaling, increased E-cadherin levels and
inhibited the invasive capability of human CRC cells. Thus,
antidepressants that target and inhibit NET may decrease the risk
of human CRC (8–10,34,35). To
the best of our knowledge, this is also the first study to describe
the molecular mechanisms downstream of NET in human CRC, which
requires further investigation.
Overall, the present study revealed a novel function
for NET in colon cancer cells and identified its downstream
effector molecules, providing valuable information for application
in future studies.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Professor Hiroyuki
Kuwano and Professor Seiji Torii (Graduate School of Medicine,
Gunma University, Maebashi, Japan) for their kind assistance; and
Mr. Xiaofei Wang, Ms. Lin Yu and Ms. Lei Ni (Key Laboratory of
Environment and Genes Related to Diseases, Xi'an Jiaotong
University Health Science Center, Xi'an, P.R. China) for their
technical support.
Funding
This work was supported by a grant from the National
Natural Science Foundation of China (grant no. 81760510) and the
Fundamental Research Funds of Xi'an Jiaotong University (grant no.
xjj2016076).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
HZ, JM, KT, XZ, XJ and NH performed the majority of
the experiments and wrote the manuscript. CH, NH and JH supported
the design and interpretation of this study. FL, WX and JZ
performed the statistical analysis. All authors read and approved
the final version of the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Medical Ethics
Committee of Xi'an Jiaotong University (Shaanxi, China). All
participants provided written informed consent. Personal
information for the samples involved in the study was
anonymized.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sung JJ, Lau JY, Goh KL and Leung WK; Asia
Pacific Working Group on Colorectal Cancer, : Increasing incidence
of colorectal cancer in asia: Implications for screening. Lancet
Oncol. 6:871–876. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Vatandoust S, Price TJ and Karapetis CS:
Colorectal cancer: Metastases to a single organ. World J
Gastroenterol. 21:11767–11776. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lutgendorf SK, Sood AK and Antoni MH: Host
factors and cancer progression: Biobehavioral signaling pathways
and interventions. J Clin Oncol. 28:4094–4099. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Worster B and Holmes S: The preoperative
experience of patients undergoing surgery for colorectal cancer: A
phenomenological study. Eur J Oncol Nurs. 12:418–424. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Alberts D, Lluria-Prevatt M, Kha S and
Weihs K: Supportive Cancer Care. 1st. Springer International
Publishing; Switzerland: pp. 45–76. 2016, PubMed/NCBI
|
|
8
|
Chubak J, Boudreau DM, Rulyak SJ and
Mandelson MT: Colorectal cancer risk in relation to antidepressant
medication use. Int J Cancer. 128:227–232. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Koh SJ, Kim JM, Kim LK, Kim N, Jung HC,
Song IS and Kim JS: Fluoxetine inhibits NF-κB signaling in
intestinal epithelial cells and ameliorates experimental colitis
and colitis-associated colon cancer in mice. Am J Physiol
Gastrointest Liver Physiol. 301:G9–G19. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kannen V, Garcia SB, Silva WA Jr, Dasser
M, Mönch R, Alho EJ, Heinsen H, Scholz CJ, Friedrich M, Heinze KG,
et al: Oncostatic effects of fluoxetine in experimental colon
cancer models. Cell Signal. 27:1781–1788. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kristensen AS, Andersen J, Jørgensen TN,
Sørensen L, Eriksen J, Loland CJ, Strømgaard K and Gether U: SLC6
neurotransmitter transporters: Structure, function and regulation.
Pharmacol Rev. 63:585–640. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mandela P and Ordway GA: The
norepinephrine transporter and its regulation. J Neurochem.
97:310–313. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pramod AB, Foster J, Carvelli L and Henry
LK: SLC6 transporters: Structure, function, regulation, disease
association and therapeutics. Mol Aspects Med. 34:197–219. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zahniser NR and Doolen S: Chronic and
acute regulation of Na+/Cl− dependent
neurotransmitter transporters: Drugs, substrates, presynaptic
receptors, and signaling systems. Pharmacol Ther. 92:21–55. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Höpfner M, Sutter AP, Huether A,
Ahnert-Hilger G and Scherübl H: A novel approach in the treatment
of neuroendocrine gastrointestinal tumors: Additive
antiproliferative effects of interferon-gamma and
meta-iodobenzylguanidine. BMC Cancer. 4:232004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hahn MK and Blakely RD: The functional
impact of SLC6 transporter genetic variation. Annu Rev Pharmacol
Toxicol. 47:401–441. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Matthay KK, George RE and Yu AL: Promising
therapeutic targets in neuroblastoma. Clin Cancer Res.
18:2740–2753. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cavallaro U and Christofori G: Cell
adhesion and signaling by cadherins and Ig-CAMs in cancer. Nat Rev
Cancer. 4:118–132. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Weiss JV, Klein-Scory S, Kübler S,
Reinacher-Schick A, Stricker I, Schmiegel W and Schwarte-Waldhoff
I: Soluble E-cadherin as a serum biomarker candidate: Elevated
levels in patients with late-stage colorectal carcinoma and FAP.
Int J Cancer. 128:1384–1392. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Okugawa Y, Toiyama Y, Inoue Y, Iwata T,
Fujikawa H, Saigusa S, Konishi N, Tanaka K, Uchida K and Kusunoki
M: Clinical significance of serum soluble E-cadherin in colorectal
carcinoma. J Surg Res. 175:e67–e73. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hu YY, Zheng MH, Zhang R, Liang YM and Han
H: Notch signaling pathway and cancer metastasis. Adv Exp Med Biol.
727:186–198. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen J, Imanaka N, Chen J and Griffin JD:
Hypoxia potentiates notch signaling in breast cancer leading to
decreased E-cadherin expression and increased cell migration and
invasion. Br J Cancer. 102:351–360. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang XQ, Zhang W, Lui EL, Zhu Y, Lu P, Yu
X, Sun J, Yang S, Poon RT and Fan ST: Notch1-Snail1-E-cadherin
pathway in metastastic hepatocellular carcinoma. Int J Cancer.
131:E163–E172. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yuan R, Ke J, Sun L, He Z, Zou Y, He X,
Chen Y, Wu X, Cai Z, Wang L, et al: HES1 promotes metastasis and
predicts poor survival in patients with colorectal cancer. Clin Exp
Metastasis. 32:169–179. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hu YF, Caron MG and Sieber-Blum M:
Norepinephrine transport-mediated gene expression in noradrenergic
neurogenesis. BMC Genomics. 10:1512009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cancer Genome Atlas Network: Comprehensive
molecular characterization of human colon and rectal cancer.
Nature. 487:330–337. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang MY, Liu XX, Li H, Li R, Liu X and Qu
YQ: Elevates mRNA levels of AURKA, CDC20 and TPX2 are associated
with poor prognosis of smoking related lung adenocarcinoma using
bioinformatics analysis. Int J Med Sci. 15:1676–1685. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hou N, Zhang X, Zhao LY, Zhao XG, Li ZF,
Song TS and Huang C: A novel chronic stress-induced shift in the
Th1 to Th2 response promotes colon cancer growth. Biochem Biophys
Res Commun. 439:471–476. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhou X, Zhao F, Wang ZN, Song YX, Chang H,
Chiang Y and Xu HM: Altered expression of miR-152 and miR-148a in
ovarian cancer is related to cell proliferation. Oncol Rep.
27:447–454. 2012.PubMed/NCBI
|
|
30
|
Ouyang W, Ren L, Liu G, Chi X and Wei H:
LncRNA MIR4435-2HG predicts poor prognosis in patients with
colorectal cancer. Peer J. 7:e66832019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tania M, Khan MA and Fu J: Epithelial to
mesenchymal transition inducing transcription factors and
metastatic cancer. Tumor Biol. 35:7335–7342. 2014. View Article : Google Scholar
|
|
32
|
Pandit-Taskar N and Modak S:
Norepinephrine transporter as a target for imaging and therapy. J
Nucl Med. 58 (Suppl 2):39S–53S. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tang J, Li Z, Lu L and Cho CH:
β-Adrenergic system, a backstage manipulator regulating tumour
progression and drug target in cancer therapy. Semin Cancer Biol.
23:533–542. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kannen V, Hintzche H, Zanette DL, Silva WA
Jr, Garcia SB, Waaga-Gasser AM and Stopper H: Antiproliferative
effects of fluoxetine on colon cancer cells and in a colonic
carcinogen mouse model. PLoS One. 7:e500432012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Stopper H, Garcia SB, Waaga-Gasser AM and
Kannen V: Antidepressant fluoxetine and its potential against colon
tumors. World J Gastrointest Oncol. 6:11–21. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Leong KG, Niessen K, Kulic I, Raouf A,
Eaves C, Pollet I and Karsan A: Jagged1-mediated notch activation
induces epithelial- to-mesenchymal transition through Slug-induced
repression of E-cadherin. J Exp Med. 204:2935–2948. 2007.
View Article : Google Scholar : PubMed/NCBI
|