Introduction
Lung carcinoma is the first leading malignancy in
males, and its incidence in females is on the rise. Globally, lung
carcinoma is a fetal malignant cancer that seriously endangers
human lives (1). In China, the
incidence of lung carcinoma ranks first in urban population and
second in rural ones (2). Non-small
cell lung carcinoma (NSCLC) is the majority subtype of lung
carcinoma, accounting for 75–80% of all lung carcinoma cases. The
prognosis of NSCLC is unsatisfactory despite the advanced
progression in therapeutic strategies. Invasiveness and metastasis
are the major reasons for the poor prognosis of NSCLC. Chemotherapy
based on platinum-containing drugs is preferred for advanced NSCLC
patients (3). Cisplatin (DDP) is the
most common first-line platinum-based chemotherapy drug. DDP could
inhibit DNA replication, affect cell transcription and translation,
and promote apoptosis of tumor cells. However, some tumor patients
experience DDP resistance during their chemotherapy treatment.
Treatment failure of DDP-based chemotherapy remarkably affects the
survival of tumor patients (4,5). Thus,
uncovering the mechanism underlying DDP resistance in tumors would
improve the therapeutic efficacy of NSCLC.
As an important member of the non-coding RNA family,
long non-coding RNA (lncRNA) regulates gene expression at
epigenetic, transcriptional, and post-transcriptional levels,
thereby affecting the progression of NCSLC (6). With the advances in high-throughput
sequencing technology and bioinformatics analysis methods, a great
number of lncRNAs with potential biological values in tumors have
been identified. lncRNAs are capable of alleviating or inducing
drug-resistance in tumors through a number of different mechanisms
(7–9).
AFAP1-AS1 is a newly discovered lncRNA that encodes
an antisense product of the AFAP1 gene. Studies have shown that
AFAP1-AS1 exerts a carcinogenic role in numerous types of tumors.
In lung carcinoma, AFAP1-AS1 is highly expressed and accelerates
invasiveness and metastasis by inhibiting AFAP1 level (10). Upregulated level of AFAP1-AS1 in
liver cancer is correlated to poor prognosis of affected patients
through activating RhoA/Rac2 pathway (11). AFAP1-AS1 is upregulated in gastric
cancer, which stimulates proliferation and suppresses apoptosis
through PTEN/p-AKT pathway (12).
Additionally, AFAP1-AS1 is significantly upregulated in esophageal
adenocarcinoma tissues and cells. Knockdown of AFAP1-AS1 in
esophageal adenocarcinoma attenuates the proliferative, migratory
and invasive abilities, whereas induces apoptosis of tumor cells
(13). However, the biological
function of AFAP1-AS1 in DDP-resistant NSCLC has not been
reported.
Patients and methods
Sample collection
Thirty-four tumor tissues of NSCLC patients were
surgically resected from January 2011 to December 2017 in Linyi
Central Hospital (Linyi, China). The patients included in the study
were 23 males and 11 females between 54 and 78 years of age.
According to the DDP resistance, the NSCLC tissues were divided
into the DDP and NG group. Samples were immediately preserved in
liquid nitrogen within 15 min ex vivo. The study was
approved by the Ethics Committee of Linyi Central Hospital. Signed
written informed consents were obtained from the patients and/or
guardians.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from NSCLC tissues using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.), and was quantified using NanoDrop 2000 spectrophotometer
(NanoDrop Technologies; Thermo Fisher Scientific, Inc.). Total RNA
was reverse transcribed into complementary deoxyribonucleic acid
(cDNA) at 37°C for 15 min using PrimeScript RT reagent (Takara
Biotechnology Co., Ltd.). The obtained cDNA was further amplified
by quantitative PCR using SYBR® Premix Ex Taq™ (Takara
Biotechnology Co., Ltd.). The reaction system volume was 25 µl in
total. The following thermocycling conditions were used for PCR:
pre-denaturation at 95°C for 5 min, denaturation at 95°C for 30
sec, annealing at 60°C for 45 sec, extension at 72°C for 3 min, for
35 cycles, and then extension at 72°C for 5 min. PCR products were
stored at 4°C. The relative levels were quantitatively analyzed
using the 2−∆∆Cq method (14). Glyceraldehyde 3-phosphate
dehydrogenase (GAPDH) was used as internal reference. Primer
sequences were as follows: AFAP1-AS1 forward, TCGCTCAATGGAGTGACGGCA
and reverse, CGGCTGAGACCGCTGAGAACT; GAPDH forward, GGA
GCGAGATCCCTCCAAAAT and reverse, GGCTGTTGT CATACTTCTCATGG; U6
forward, AAAATATGGAACGC TTCACGAATTTG and reverse, CTCGCTTCGGCAGCA
CATATACT.
Cell culture
A549 cells (cat. no. SCSP-503) and DDP-resistant
cell line A549/DDP (cat. no. SCSP-524) were provided by the
Shanghai Cell Bank (http://www.cellbank.org.cn/). A549 cells were cultured
in RPMI-1640 medium (HyClone; GE Healthcare Life Sciences)
containing 10% fetal bovine serum (FBS) (Gibco; Thermo Fisher
Scientific, Inc.). A549/DDP cells were cultured in medium
containing 1 µmol/l DDP to maintain drug resistance.
Cell transfection
Cells in the logarithmic growth phase were subjected
to transfection at confluency of 70–80%. Transfection vectors and
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) were respectively diluted in Opti-MEM (Shanghai
GeneChem Co., Ltd.). After mixture and incubation for 20 min, the
cells were plated in each well. si-AFAP1-AS1 at the concentration
of 50 nM was added to each well and then incubated for 48 h.
Complete fresh medium was replaced after 6 h. The transfection
plasmids were provided by Shanghai GenePharma Co., Ltd.
Cell Counting Kit-8 (CCK-8)
Cells were seeded in a 96-well plate with
2.0×103 cells/well and incubated with 0, 0.5, 1, 1.5, 2
and 2.5 mg/ml DDP, respectively. Each experiment was performed in
triplicate. Absorbance at 450 nm was recorded using CCK-8 kit
(Dojindo Molecular Technologies, Inc.), the viability curve was
plotted and IC50 was calculated.
Colony formation assay
Cells were seeded in a 6-well plate with 500
cells/well and incubated for 2–3 weeks. Subsequently, the cells
were fixed in 100% methanol at 20°C and dyed with 0.5% crystal
violet (Sigma-Aldrich; Merck KGaA) at 20°C for 20 min. Colonies of
>50 µm in size were counted by Quantity One software (Bio-Rad
Laboratories, Inc.). Colonies were finally observed under a light
microscope (BX-42; Olympus Corporation) and calculated
(magnification, ×10).
Cell cycle determination
Transfected cells for 48 h were washed with pre-cold
PBS and fixed in pre-cold ethanol overnight. Subsequently, the
cells were subjected to incubation at 20°C for 10 min with 10 µl of
propidium iodide (PI) and RNase A in the dark for 1 h. Cell cycle
was determined using a flow cytometer (FACSCalibur; BD
Biosciences). Data were obtained and analyzed using the CellQuest
Pro software (version 3.3; Becton, Dickinson and Company).
Cell apoptosis determination
Cells were resuspended in 500 µl of binding buffer
at the dose of 1–5×105/ml. Subsequently, the cells were
subjected to incubation with 5 µl of Annexin V-FITC and 5 µl of PI
in the dark at 20°C for 30 min. Apoptosis was determined using flow
cytometry within half an hour. Flow cytometer (FACSCalibur) was
used for analysis. Data were obtained and analyzed using the
CellQuest Pro software (version 3.3).
Western blot analysis
The cells were lysed using a cell lysis buffer (cat.
no. QC25-05099; Shanghai Qincheng Biotechnology, Co.). Total
protein was extracted from cells using radioimmunoprecipitation
assay and was quantified using the bicinchoninic acid (both from
Beyotime Institute of Biotechnology) method. A total of 30 µg of
protein was added per lane for electrophoresis. The extracted
protein was separated using a 10% SDS-PAGE gel. After transferred
onto polyvinylidene fluoride membranes (EMD Millipore), the protein
was blocked in 5% skim milk at 4°C for 2 h, incubated with primary
antibodies at 4°C overnight and secondary antibodies at 20°C for 2
h. Bands were visualized by an enhanced chemiluminescence detection
kit (Amersham; GE Healthcare) and analyzed by ImageJ Software
(National Institutes of Health). Rabbit polyclonal Akt antibody
(dilution, 1:500; cat. no. ab8805), rabbit monoclonal p-Akt
antibody (dilution, 1:500; cat. no. ab81283), rabbit polyclonal
E-cadherin antibody (dilution, 1:500; cat. no. ab15148), rabbit
polyclonal N-cadherin antibody (dilution, 1:500; cat. no. ab18203),
rabbit monoclonal snail antibody (dilution, 1:500; cat. no.
ab216347), rabbit polyclonal vimentin antibody (dilution, 1:500;
cat. no. ab137321), rabbit polyclonal GAPDH antibody (dilution,
1:500; cat. no. ab37168) and secondary goat anti-rabbit (HRP) IgG
antibody (dilution, 1:2,000; cat. no. ab6721) were all purchased
from Abcam. GAPDH was used as the reference protein.
Transwell assay
Cell density was adjusted to 1–5×105/ml.
Suspension (200 µl/well) was applied in the upper chamber of the
Matrigel-coated (diluted in serum-free medium with 1:10) Transwell
chamber (EMD Millipore). In the bottom chamber, 500 µl of medium
containing 10% FBS were applied. The cells on the top surface of
the membrane were removed by a cotton swab after a 24-h incubation
at 37°C. After incubation for 48 h, the invasive cells were fixed
in 4% paraformaldehyde, dyed with crystal violet for 2 min at 37°C
and counted using a light microscope (BX-42; Olympus Corporation).
Penetrating cells were counted in ten randomly selected fields per
sample (magnification, ×10). Transwell migration assay was
conducted following the same procedures except for Matrigel
pre-coating.
Subcellular distribution analysis
Cytoplasmic and nuclear RNA was extracted using the
PARIS kit (Invitrogen; Thermo Fisher Scientific, Inc.) and then
subjected to RT-qPCR. U6 and GAPDH were the internal references of
nucleus and cytoplasm, respectively.
RNA immunoprecipitation (RIP)
Cells were treated following the manufacturer's
instructions of Millipore Magna RIP™ RNA-Binding Protein
Immunoprecipitation kit (EMD Millipore). Cell lysate was incubated
with rabbit monoclonal enhancer of zeste homolog 2 (EZH2) antibody
(dilution, 1:500; cat. no. ab191080; Abcam) or anti-IgG antibody at
4°C for 6 h. A protein-RNA complex was captured and digested with
0.5 mg/ml proteinase K containing 0.1% SDS to extract RNA. The
magnetic beads were repeatedly washed with RIP washing buffer to
remove non-specific adsorption as much as possible. Finally, the
extracted RNA was subjected to mRNA level determination using
RT-qPCR.
Statistical analysis
SPSS 19.0 software (IBM Corp.) was used for data
analysis. Data were expressed as the mean ± standard deviation.
Intergroup comparisons were made using t-test. Comparisons between
multiple groups were made using one-way ANOVA followed by the Least
Significant Difference post hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
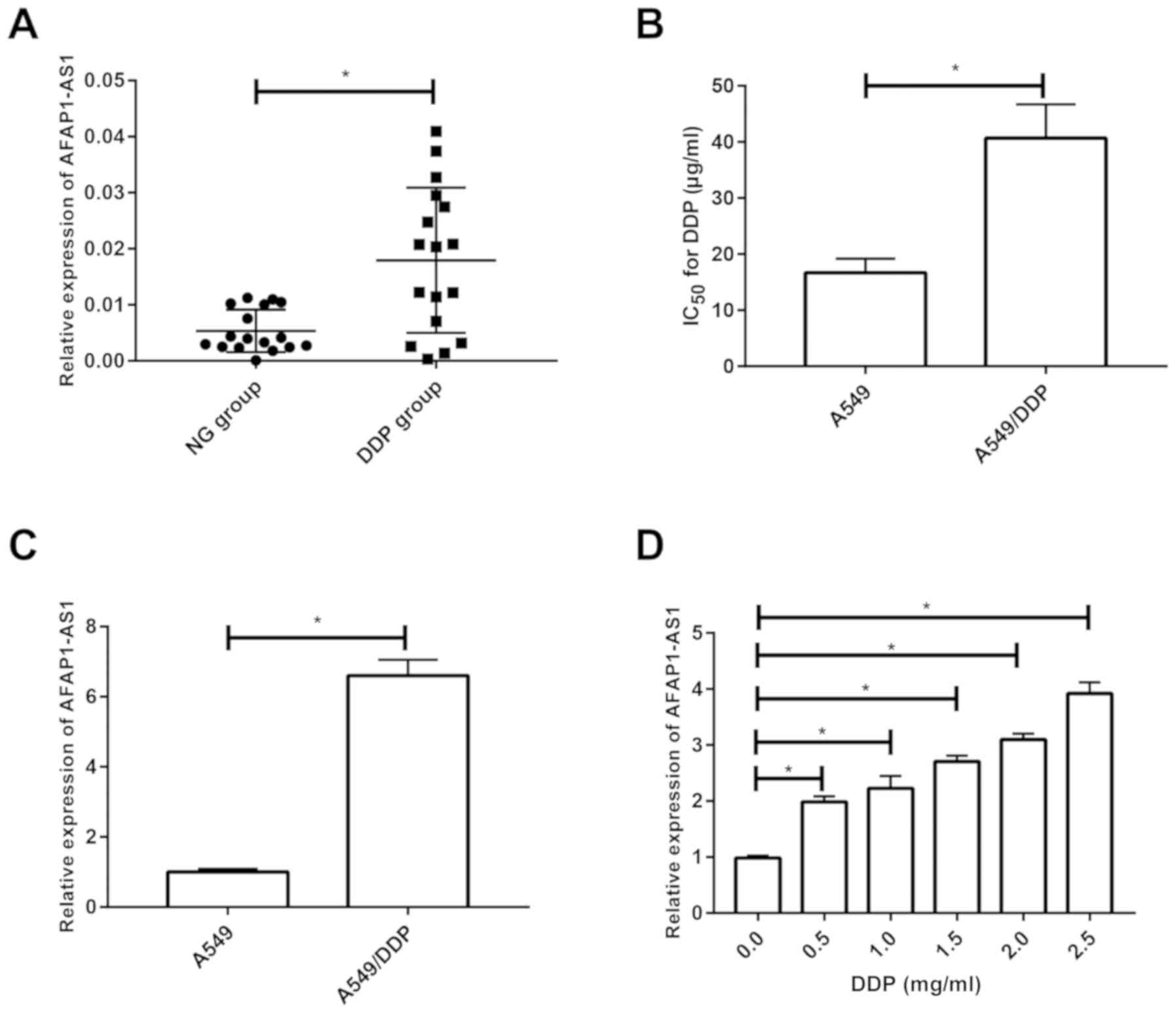
AFAP1-AS1 is upregulated in
DDP-resistant NSCLC patients and cell lines
Compared with the NSCLC patients who did not receive
DDP-based chemotherapy, those with DDP resistance presented higher
level of AFAP1-AS1 (Fig. 1A). CCK-8
assay revealed a higher IC50 for DDP in A549/DDP cells
compared with that in A549 cells, verifying the cytotoxicity in
DDP-resistant A549 cell line (Fig.
1B). Similarly, AFAP1-AS1 expression was much higher in
A549/DDP cells than that in the parental cells (Fig. 1C). A549 cells were cultured in medium
containing 0, 0.5, 1, 1.5, 2 and 2.5 mg/ml DDP for 24 h. RT-qPCR
revealed that the AFAP1-AS1 expression gradually increased in a
dose-dependent manner, indicating the involvement of AFAP1-AS1 in
DDP-resistant NSCLC (Fig. 1D).
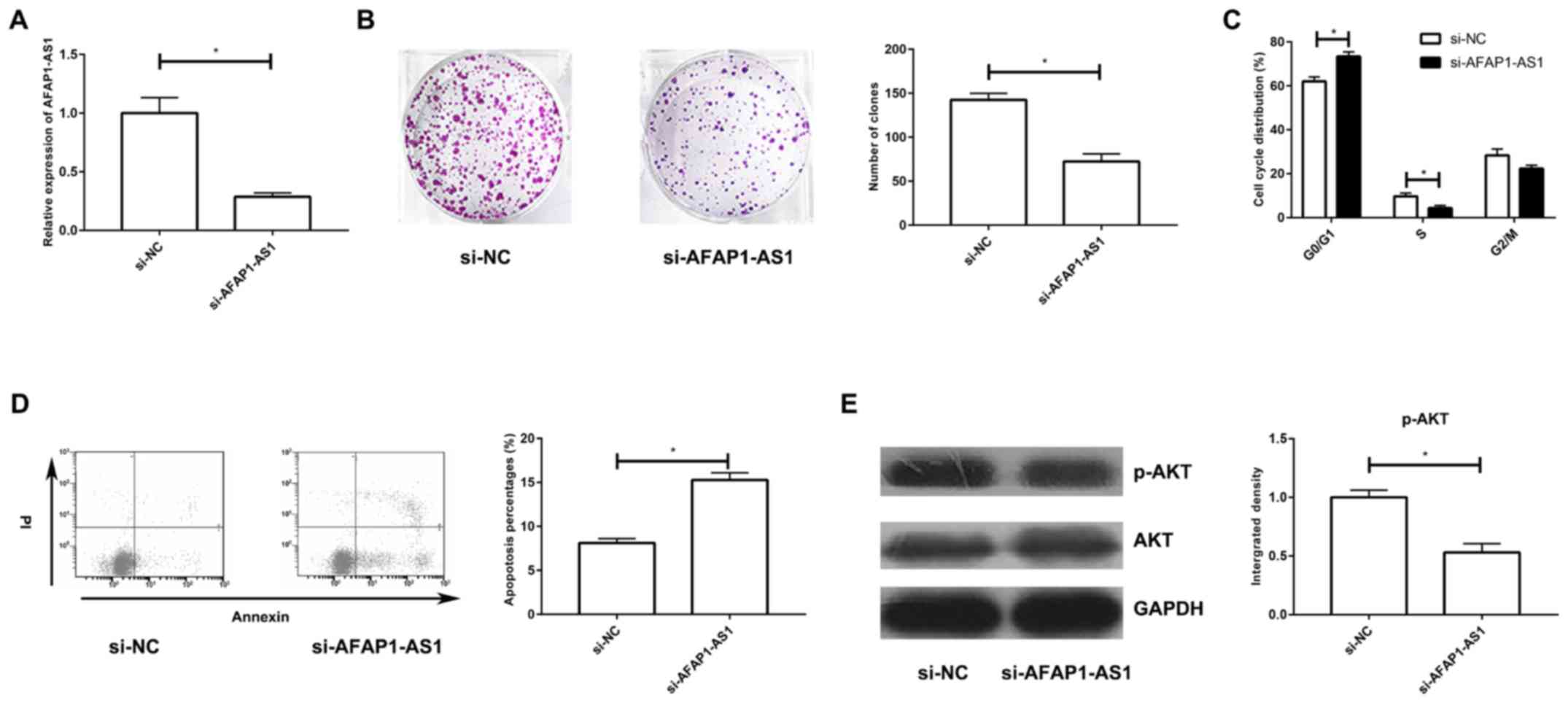
Silencing of AFAP1-AS1 suppresses the
proliferative ability and cell cycle progression, and induces
apoptosis in A549/DDP cells through the PI3K/AKT pathway
To uncover the biological function of AFAP1-AS1 in
DDP-resistant NSCLC, si-AFAP1-AS1 was constructed and its
transfection efficacy in A549/DDP cells was tested (Fig. 2A). Transfection with si-AFAP1-AS1
markedly decreased the number of colonies in A549/DDP cells and
arrested the cells in G0/G1 phase (Fig.
2B and C). Moreover, silencing of AFAP1-AS1 greatly enhanced
the apoptotic rate in DDP-resistant cells (Fig. 2D). Western blot analysis revealed the
downregulation of p-AKT in A549/DDP cells transfected with
si-AFAP1-AS1, confirming the inhibited PI3K/AKT pathway (Fig. 2E).
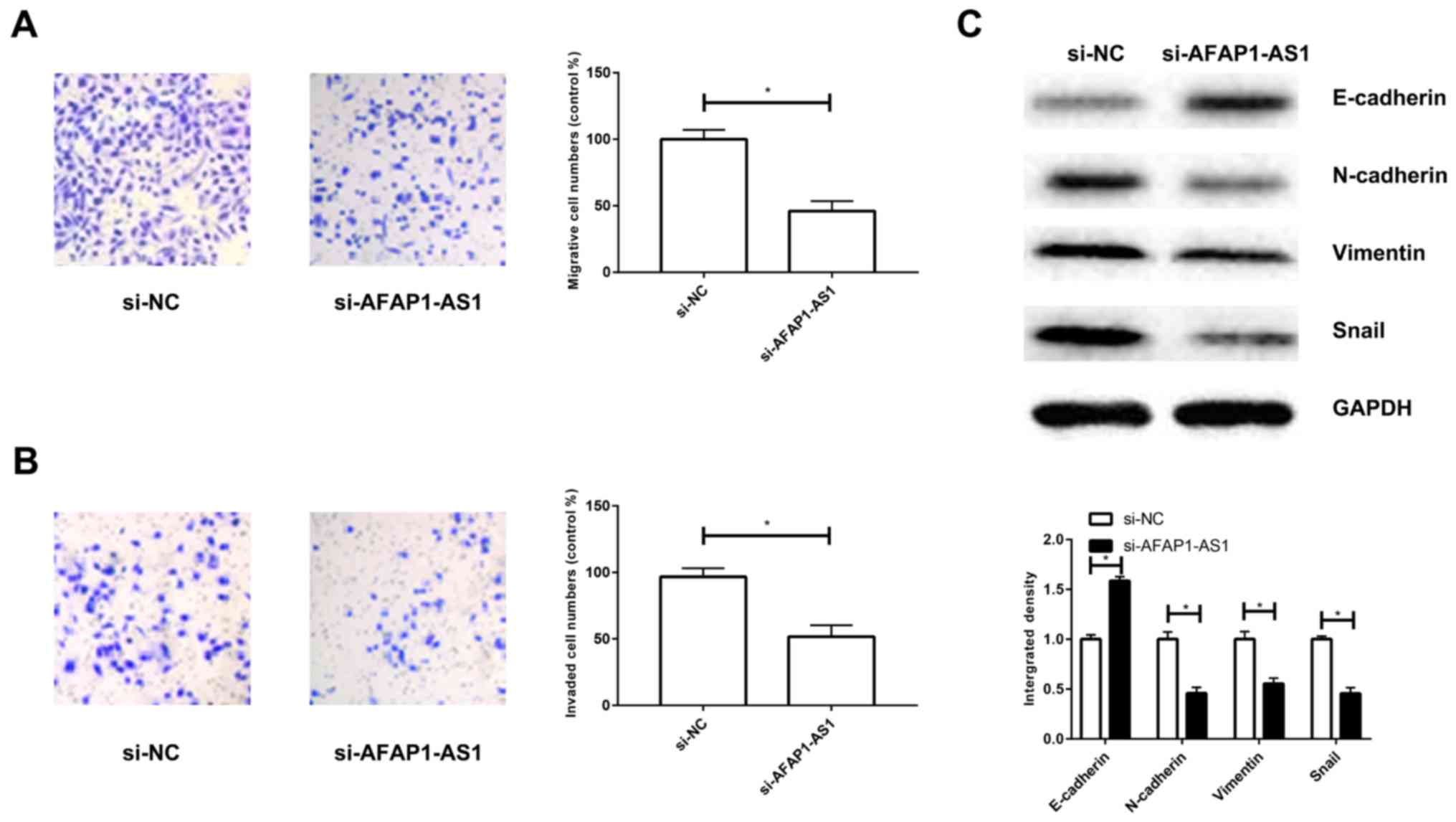
Silencing of AFAP1-AS1 suppresses
migratory and invasive abilities of A549/DDP cells by inhibiting
epithelial-mesenchymal transition (EMT)
Transfection with si-AFAP1-AS1 attenuated the
migratory and invasive abilities of A549/DDP cells (Fig. 3A and B). Since EMT is vital in tumor
metastasis, key proteins related to EMT were examined. Silencing of
AFAP1-AS1 upregulated E-cadherin; however, downregulated
N-cadherin, vimentin and snail (Fig.
3C). These results suggest that knockdown of AFAP1-AS1 markedly
inhibits EMT in DDP-resistant NSCLC.
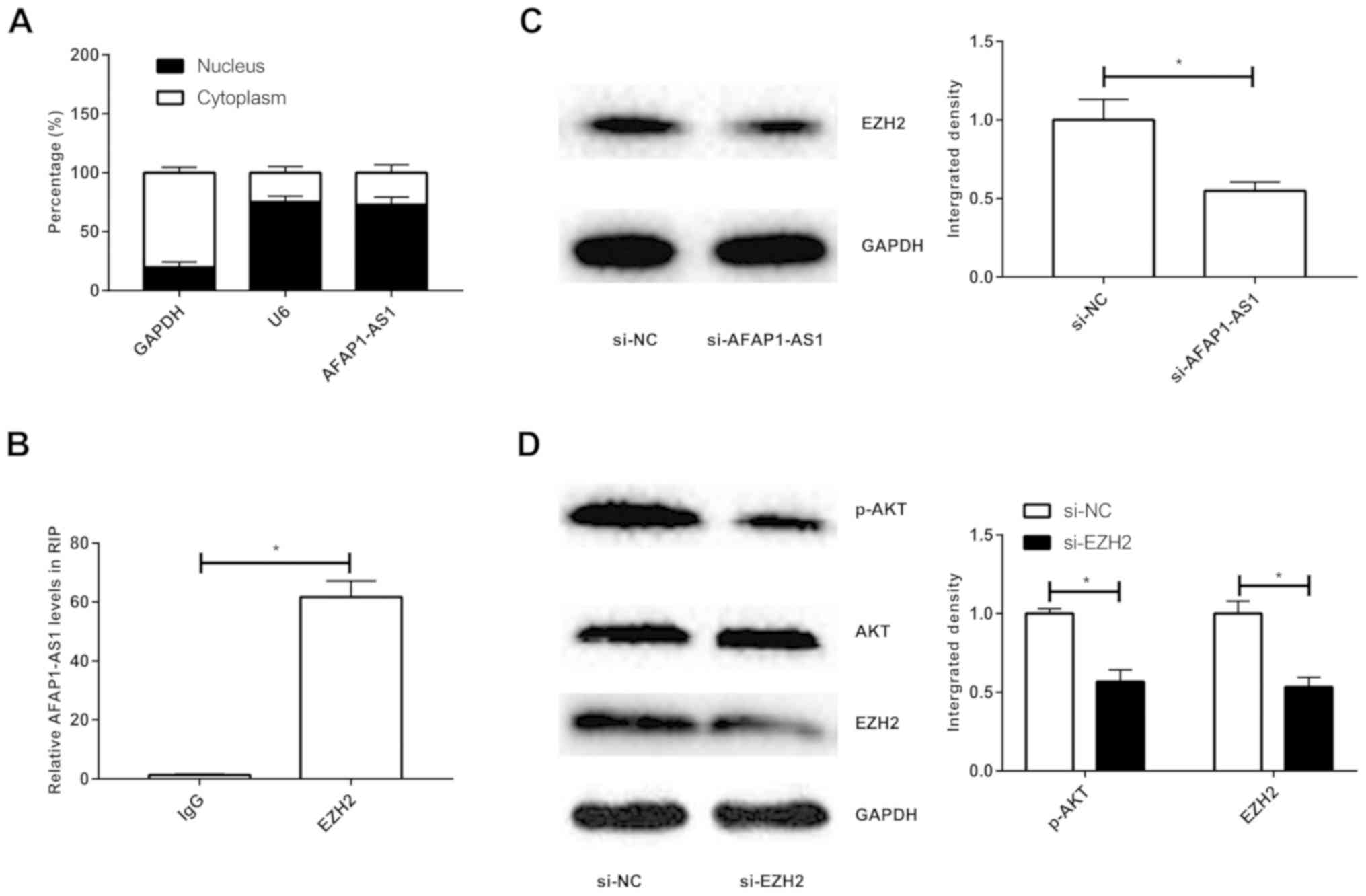
AFAP1-AS1 activates PI3K/AKT pathway
through interacting with EZH2
Subcellular distribution analysis revealed that
AFAP1-AS1 was mainly expressed in nucleus (Fig. 4A). A previous study has presented
that EZH2 induces the activation of PI3K/AKT pathway in NSCLC
(15). Herein, RIP assay revealed
higher enrichment of AFAP1-AS1 in anti-EZH2 compared with that in
anti-IgG, confirming the interaction between AFAP1-AS1 and EZH2
(Fig. 4B). Transfection with
si-AFAP1-AS1 sufficiently downregulated the protein level of EZH2,
presenting a negative association (Fig.
4C). After transfection with si-EZH2, the protein level of
p-AKT was remarkably downregulated, confirming the inhibited
PI3K/AKT pathway (Fig. 4D).
Discussion
In the present study, abnormally expressed AFAP1-AS1
was found to be related to DDP resistance in NSCLC. AFAP1-AS1 level
remained higher in DDP-resistant NSCLC tissues and cell lines.
Silencing of AFAP1-AS1 suppressed proliferative, migratory and
invasive abilities of A549/DDP cells through regulating EMT and
PI3K/AKT pathway.
EMT is the process where glandular epithelial cells
transform to mesenchymal cells. EMT enhances the migratory and
invasive abilities, and attenuates aging and apoptosis of cells,
thus improving the immunosuppression. EMT is a vital mechanism
leading to distant metastasis of tumors. Once EMT occurs,
downregulated E-cadherin disrupts the cell conjunction between
adjacent epithelial cells, dislocates in situ tumor cells
and thereafter results in tumor metastasis (16,17). A
variety of lncRNAs have been found to induce EMT in human tumors.
lncRNA XIST induces EMT in osteosarcoma through targeting
miR-195-5p/YAP axis (18). lncRNA
XIST also stimulates the progression of EMT in NSCLC via regulating
miR-367/miR-141-ZEB2 axis (19).
lncRNA DNM3OS has been reported to participate in the EMT of
ovarian cancer (20). Upregulation
of DNM3OS stimulates the metastatic capacity of ovarian cancer
cells and predicts a poor prognosis (20). In colorectal cancer, lncRNA N-BLR
promotes EMT and metastasis of tumor cells by mediating EMT-related
transcriptional factor Zeb1 (21).
In the present study, silencing of AFAP1-AS1 upregulated
E-cadherin, whereas downregulated N-cadherin, vimentin and snail,
suggesting that AFAP1-AS1 markedly induces EMT in DDP-resistant
NSCLC.
Transfection of A549/DDP cells with si-AFAP1-AS1
downregulated p-AKT level, indicating the involvement of PI3K/AKT
pathway in DDP-resistant NSCLC. PI3K/AKT pathway is involved in
regulating multiple cellular behaviors and tumor progression.
PI3K/AKT pathway could be influenced by multiple factors in tumor
diseases. For example, miR-221 activates PI3K/AKT pathway by
negatively regulating PTEN, thereby mediating gefitinib sensitivity
in cervical cancer (22). miR-146b
aggravates the progression of thyroid cancer via targeting PI3K/AKT
pathway (23). Recent studies have
reported that numerous lncRNAs could be able to mediate PI3K/AKT
pathway and thereafter aggravate tumor progression as oncogenes.
For example, lncRNA ASAP1-IT1 attenuates malignant phenotypes of
NSCLC through targeting PTEN/AKT pathway (24). The growth of osteosarcoma is
stimulated by lncRNA UCA1 through activating PTEN/AKT pathway
(25). The proliferation and EMT of
NSCLC cells are promoted through lncRNA FAL1-induced activation of
PTEN/AKT pathway (26). The results
of the present study demonstrated that AFAP1-AS1 robustly interacts
with EZH2 to activate PI3K/AKT pathway.
This study only constructed the knockdown model of
AFAP1-AS1 in A549/DDP cells. In future research, an overexpression
model of AFAP1-AS1 using lentiviruses or overexpression plasmids is
required to confirm the conclusions of this study.
In conclusion, AFAP1-AS1 accelerates the
proliferative and metastatic abilities of the A549/DDP cells,
whereas inhibits the apoptosis of the A549/DDP cells by interacting
with EZH2 to activate PI3K/AKT pathway; thus, inducing DDP
resistance in NSCLC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YL and XW designed the study and performed the
experiments, YL and QH acquired the data, XW and QH analyzed the
data, YL and XW prepared the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
Linyi Central Hospital (Linyi, China). Signed written informed
consents were obtained from the patients and/or guardians.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhao P, Dai M, Chen W and Li N: Cancer
trends in China. Jpn J Clin Oncol. 40:281–285. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zappa C and Mousa SA: Non-small cell lung
cancer: Current treatment and future advances. Transl Lung Cancer
Res. 5:288–300. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xiong Y, Huang BY and Yin JY:
Pharmacogenomics of platinum-based chemotherapy in non-small cell
lung cancer: Focusing on DNA repair systems. Med Oncol. 34:482017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tièche CC, Peng RW, Dorn P, Froment L,
Schmid RA and Marti TM: Prolonged pemetrexed pretreatment augments
persistence of cisplatin-induced DNA damage and eliminates
resistant lung cancer stem-like cells associated with EMT. BMC
Cancer. 16:1252016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhan Y, Zang H, Feng J, Lu J, Chen L and
Fan S: Long non-coding RNAs associated with non-small cell lung
cancer. Oncotarget. 8:69174–69184. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pan JJ, Xie XJ, Li X and Chen W: Long
non-coding RNAs and drug resistance. Asian Pac J Cancer Prev.
16:8067–8073. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang H, Guan Z, He K, Qian J, Cao J and
Teng L: LncRNA UCA1 in anti-cancer drug resistance. Oncotarget.
8:64638–64650. 2017.PubMed/NCBI
|
|
9
|
Loewen G, Jayawickramarajah J, Zhuo Y and
Shan B: Functions of lncRNA HOTAIR in lung cancer. J Hematol Oncol.
7:902014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zeng Z, Bo H, Gong Z, Lian Y, Li X, Li X,
Zhang W, Deng H, Zhou M, Peng S, et al: AFAP1-AS1, a long noncoding
RNA upregulated in lung cancer and promotes invasion and
metastasis. Tumour Biol. 37:729–737. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang JY, Weng MZ, Song FB, Xu YG, Liu Q,
Wu JY, Qin J, Jin T and Xu JM: Long noncoding RNA AFAP1-AS1
indicates a poor prognosis of hepatocellular carcinoma and promotes
cell proliferation and invasion via upregulation of the RhoA/Rac2
signaling. Int J Oncol. 48:1590–1598. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Guo JQ, Li SJ and Guo GX: Long noncoding
RNA AFAP1-AS1 promotes cell proliferation and apoptosis of gastric
cancer cells via PTEN/p-AKT pathway. Dig Dis Sci. 62:2004–2010.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu W, Bhagat TD, Yang X, Song JH, Cheng Y,
Agarwal R, Abraham JM, Ibrahim S, Bartenstein M, Hussain Z, et al:
Hypomethylation of noncoding DNA regions and overexpression of the
long noncoding RNA, AFAP1-AS1, in Barrett's esophagus and
esophageal adenocarcinoma. Gastroenterology. 144:956–966.e4. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Geng J, Li X, Zhou Z, Wu CL, Dai M and Bai
X: EZH2 promotes tumor progression via regulating VEGF-A/AKT
signaling in non-small cell lung cancer. Cancer Lett. 359:275–287.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Brabletz T, Kalluri R, Nieto MA and
Weinberg RA: EMT in cancer. Nat Rev Cancer. 18:128–134. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Roche J: The epithelial-to-mesenchymal
transition in cancer. Cancers (Basel). 10(pii): E522018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang C, Wu K, Wang S and Wei G: Long
non-coding RNA XIST promotes osteosarcoma progression by targeting
YAP via miR-195-5p. J Cell Biochem. 119:5646–5656. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li C, Wan L, Liu Z, Xu G, Wang S, Su Z,
Zhang Y, Zhang C, Liu X, Lei Z, et al: Long non-coding RNA XIST
promotes TGF-β-induced epithelial-mesenchymal transition by
regulating miR-367/141-ZEB2 axis in non-small-cell lung cancer.
Cancer Lett. 418:185–195. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mitra R, Chen X, Greenawalt EJ, Maulik U,
Jiang W, Zhao Z and Eischen CM: Decoding critical long non-coding
RNA in ovarian cancer epithelial-to-mesenchymal transition. Nat
Commun. 8:16042017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rigoutsos I, Lee SK, Nam SY, Anfossi S,
Pasculli B, Pichler M, Jing Y, Rodriguez-Aguayo C, Telonis AG,
Rossi S, et al: N-BLR, a primate-specific non-coding transcript
leads to colorectal cancer invasion and migration. Genome Biol.
18:982017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Du J, Wang L, Li C, Yang H, Li Y, Hu H, Li
H and Zhang Z: MicroRNA-221 targets PTEN to reduce the sensitivity
of cervical cancer cells to gefitinib through the PI3K/Akt
signaling pathway. Tumour Biol. 37:3939–3947. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ramírez-Moya J, Wert-Lamas L and
Santisteban P: MicroRNA- 146b promotes PI3K/AKT pathway
hyperactivation and thyroid cancer progression by targeting PTEN.
Oncogene. 37:3369–3383. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang L, Shi SB, Zhu Y, Qian TT and Wang
HL: Long non-coding RNA ASAP1-IT1 promotes cell proliferation,
invasion and metastasis through the PTEN/AKT signaling axis in
non-small cell lung cancer. Eur Rev Med Pharmacol Sci. 22:142–149.
2018.PubMed/NCBI
|
|
25
|
Li T, Xiao Y and Huang T: HIF-1α-induced
upregulation of lncRNA UCA1 promotes cell growth in osteosarcoma by
inactivating the PTEN/AKT signaling pathway. Oncol Rep.
39:1072–1080. 2018.PubMed/NCBI
|
|
26
|
Pan C, Yao G, Liu B, Ma T, Xia Y, Wei K,
Wang J, Xu J, Chen L and Chen Y: Long noncoding RNA FAL1 promotes
cell proliferation, invasion and epithelial-mesenchymal transition
through the PTEN/AKT signaling axis in non-small cell lung cancer.
Cell Physiol Biochem. 43:339–352. 2017. View Article : Google Scholar : PubMed/NCBI
|