Introduction
Gastric cancer (GC) is the fifth most frequent
cancer (1,033,701 new cases in 2018) and the third leading cause of
cancer-associated mortality 782,685 deaths in 2018) worldwide
(1). Incidence rates are the highest
in East Asia, particularly in Japan and China (1). Despite comprehensive use of surgical
treatment, chemotherapy, radiotherapy and molecular-targeted
treatment, the 5-year survival rate of patients with GC remains
<20% in China (2–4). Further investigation is therefore
required to fully elucidate the molecular mechanisms underlying the
progression of GC and to identify novel therapeutic targets.
Autophagy is a highly conserved catabolic process in
which cytoplasmic material and organelles are delivered to
lysosomes for degradation. Autophagy is a multi-step and complex
process that is regulated by ~30 autophagy-associated proteins and
multiple signaling pathways, including AMPK, PERK (5,6).
Dysregulation of autophagy has been detected in numerous human
diseases, including cancer (7). In
addition, increasing evidence indicates that autophagy serves a
crucial role in tumor cell proliferation and differentiation, and
that it affects the efficacy of anti-tumor drugs (8,9).
Although a number of studies have investigated autophagy in GC
(10–15), the precise role of autophagy in the
development of GC remains to be completely identified.
Long non-coding RNAs (lncRNAs) serve crucial roles
in various biological processes, including immune responses,
angiogenesis, cell proliferation, differentiation, apoptosis and
autophagy (16–18). Previous studies have also reported
the abnormal expression and roles of lncRNAs in cancer (19–21).
Investigation of the tumor-associated molecular mechanisms of
lncRNAs may therefore contribute to the prevention and treatment of
cancer. Metastasis-associated lung adenocarcinoma transcript 1
(MALAT1) is a lncRNA discovered in 2003 that is closely
associated with the clinical outcome and progression of lung cancer
(22). MALAT1 expression has
been reported to be significantly upregulated in lung cancer,
hepatocellular carcinoma, bladder cancer and other types of
malignancy (23–25). A recent study reported that
MALAT1 may be used as a diagnostic marker of GC metastasis
(26). However, the precise
mechanism of MALAT1 in the development of GC remains not
fully understood.
MicroRNA (miR)-204 is a well-studied tumor
suppressor, which is commonly downregulated in breast and prostate
cancer, renal cell carcinoma and GC (27–32).
Numerous studies have indicated that miR-204 can repress the
development of GC (31,33,34).
Furthermore, loss of miR-204 leads to upregulated expression of
transient receptor potential melastatin 3 (TRPM3), which stimulates
oncogenic autophagy by regulating microtubule-associated protein 1
light chain 3α (MAP1LC3A, also known as LC3A) and LC3B and promotes
cancer growth (33,35). However, in cholangiocarcinoma and
lung cancer, miR-204 is negatively regulated by MALAT1
(36,37).
The present study aimed to clarify the association
between MALAT1, miR-204, and GC progression, and to
investigate the underlying molecular mechanism of GC.
Materials and methods
Clinical samples
A total of 57 human GC tissues and corresponding
non-cancerous adjacent tissues (5 cm from the edge of tumor) were
collected from 18 women and 39 men who received surgery at Jiangyin
Hospital Affiliated to Nantong University (Jiangyin, China) between
September 2017 and June 2018. The mean age of patients was 56.3
years (age range, 37–72 years). All patient materials were obtained
with written informed consent in accordance with the requirements
of the Clinical Research Ethics Committees of Jiangyin Hospital
Affiliated to Nantong University; The experimental protocols were
approved by the Research Ethics Committee in Jiangyin Hospital
Affiliated to Nantong University (approval no. JY1608AC97). All
methods used in this study were in accordance with the approved
guidelines.
Cell lines and transfection
The human normal gastric epithelial cell line GES-1
and the GC cell line MKN45 were purchased from the American Type
Culture Collection. The gastric adenocarcinoma-derived circulating
tumor cell lines CTC141 and CTC105 (38) were kindly donated by the Laboratory
of Stem Cell Biology of Sichuan University (Chengdu, China). All
cells were cultured in RPMI-1640 medium (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% fetal bovine serum (Gibco;
Thermo Fisher Scientific, Inc.) and 1% penicillin-streptomycin
(Sigma-Aldrich; Merck KGaA) and placed at 37°C in a humidified
incubator containing 5% CO2. Cells were passaged every
2–3 days.
For the transfection, the small interfering (si)RNA
were obtained from GenSript. Sequences for the siRNAs were designed
as follows: Negative control siRNA (si-NC),
5′-CUUGCCUGGACCAGCUUAAdTdT-3′; si-MALAT1−1,
5′-CAGCCCGAGACTTCTGTAAdTdT-3′; and si-MALAT2,
5′-AGCCCGAGACTTCTGTAAAdTdT-3′. The pcDNA vector and MALAT1
overexpression plasmid (pcDNA-MALAT1) were obtained from
Nanjing KGI Biological Technology Development Co., Ltd. miR-204
mimic (cat. no. miR10022693-1-5) and mimic control were purchased
from Guangzhou RiboBio Co., Ltd. Transcription efficiency was
evaluated by quantitative PCR.
CTC 141 and CTC105 cells were transiently
transfected with siRNA (100 nM/1×105 cells), plasmid (1
µg/1×105 cells) or mimic (100 nM/1×105 cells)
using Lipofectamine® 3000 (12 µl/1×105 cells;
Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. Following 48 h transfection, cells
were subjected to experiments. The transcription efficiency was
evaluated by quantitative real-time PCR.
MTT assay
CTC 141 and CTC105 cell proliferation was measured
with MTT assay (Sigma-Aldrich). Cells were seeded into 96-well
plates (3×103 cells/well) for 24, 48 or 72 h and were
treated for 3 h with 2 mM 3-methyladenine (3-MA; Selleck, cat. no.
S2767), used as a cell autophagy inhibitor, before assessing cell
proliferation. MTT (20 µl) was then added to each well for 4 h at
37°C. Cell viability was assessed by detecting absorbance at 450 nm
using a microplate reader (Thermo Fisher Scientific, Inc.). All
tests were performed in quadruplicate.
Colony formation assay
Transfected CTC105 cells were plated in a 6-well
plate (200 cells/well) and cultured for 14 days under standard
culture conditions. Cell culture medium was replaced every 2–3
days. Cells were fixed with methanol for 20 min, and stained with
0.5% crystal violet for 5 min at room temperature. Stained colonies
(cells number >30) were imaged using a Nikon camera (Nikon,
Tokyo, Japan). Each experiment was performed at least three
times.
Immunofluorescence staining
Paraffin-embedded GC tissues were collected and
fixed with 10% formalin overnight at room temperature. Section (0.5
µm thick) were dehydrated by increasing ethanol gradient (50, 70,
85, 95 and 100%) at room temperature for 2 h, deparaffinized with
dimethylbenzene at room temperature for 15 min and rehydrated by
decreasing gradient of ethanol (100, 95, 85 and 75%) for 5 min at
room temperature. Tissue sections were incubated in 0.5 M sodium
citrate buffer (JRDUN Biotechnology, Co., Ltd.) at 37°C for 15 min
and heated in a microwave at 92–98°C for 20 min for antigen
retrieval. Tissue sections were permeabilized with 0.1% Triton
X-100 (Sangon Biotech Co. Ltd.) in PBS for 25 min at room
temperature, and blocked with 2% bovine serum albumin
(Sigma-Aldrich) in PBS for 1 h at room temperature. Tissue sections
were then incubated with anti-LC3B (Cell Signaling Technology,
Inc.; cat. no. 83506; 1:200) and anti-Ki67 antibodies (Cell
Signaling Technology, Inc.; cat. no. 9449; 1:300) at 4°C overnight.
Tissue sections were then incubated with fluorescein
isothiocyanate-labeled secondary anti-mouse antibody (Cell
Signaling Technology Europe, B.V.; cat. no. 4410; 1:500) for 2 h at
room temperature. Cell nuclei were stained with DAPI (1 µg/ml;
Sigma-Aldrich; Merck KGaA) for 5 min at room temperature. Stained
sections were observed under a CX41RF fluorescence microscope
(Olympus Corporation) with ×200 magnification.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
TRIzol® reagent (Takara Biotechnology
Co., Ltd.) was used to extract total RNA from gastric tissues and
cells. cDNAs were generated using the PrimeScript™ RT reagent kit
(Takara Biotechnology Co., Ltd.) according to the manufacturer's
instructions. SYBR® Premix Ex Taq (Takara Biotechnology
Co., Ltd.) was used for qPCR according to the manufacturer's
instructions. The thermocycling conditions were as follows: 95°C
for 10 sec, followed by 40 cycles of 95°C for 5 sec, 60°C for 15
sec, and 72°C for 30 sec, the final extension was 72°C for 5 min.
Primer sequences used for qPCR were as follows: MALAT1
forward, 5′-AGCGGAAGAACGAATGTAAC-3′ and reverse,
5′-GAACAGAAGGAAGAGCCAAG-3′; LC3B forward,
5′-GATGTCCGACTTATTCGAGAGC-3′ and reverse,
5′-TTGAGCTGTAAGCGCCTTCTA-3′; TRPM3 forward,
5′-ATACCCAGCACCAAAGACC-3′ and reverse 5′-TCTGAAGCACGGAGATACTG-3′;
and GAPDH forward, 5′-TGAACGGGAAGCTCACTGG-3′ and reverse,
5′-TCCACCACCCTGTTGCTGTA-3′. The relative expressions levels were
normalized to the endogenous control GAPDH and calculated
using the 2−ΔΔCq method (39).
To detect miRNA-204, reverse transcription and qPCR
were performed using a Bulge-Loop™ miRNA qPCR Primer Set for
hsa-miR-204 (Guangzhou RiboBio Co., Ltd.) and U6 snRNA (Guangzhou
RiboBio Co., Ltd.) according to the manufacturer's instructions and
as previously described (40). U6
served as an internal control.
Western blotting
Total cellular proteins from CTC105 and CTC141 cells
were extracted using radioimmunoprecipitation assay buffer
(Auragene). Protein concentration was determined with bicinchoninic
acid assay (Thermo Fisher Scientific, Inc.). Proteins (20 µg) were
separated by 12% SDS-PAGE and transferred onto polyvinylidene
difluoride membranes. Membranes were blocked with 5% skimmed milk
at room temperature for 1 h and incubated with primary antibodies
against p62 (cat. no. 39749; 1:2,000; Cell Signaling Technology,
Inc.), LC3B (cat. no. 3868; 1:1,000; Cell Signaling Technology,
Inc.), Ki67 (cat. no. 13110; 1:5,000; Cell Signaling Technology,
Inc.), β-actin (cat. no. 4970; 1:1,000; Cell Signaling Technology,
Inc.) and TRPM3 (cat. no. ab56171; 1:1,000; Abcam) at room
temperature for 2 h. Bands were detected using enhanced
chemiluminescence substrate (Applygen Technologies, Inc.) according
the manufacturer's protocol. Protein quantification was performed
by ImageJ software (National Institutes of Health).
Statistical analysis
Data are presented as the mean ± standard deviation.
SPSS 17.0 statistical software (SPSS, Inc.) was used for
statistical analyses. Comparison between groups was performed with
paired Student's t-test or analysis of variance followed by
Holm-Sidak's or Dunnett's multiple comparisons test (GraphPad
Software, Inc.). The expression correlation was analyzed using
Pearson's correlation test. P<0.05 was considered to indicate a
statistically significant difference.
Results
MALAT1 is associated with autophagy
activation in GC tissues
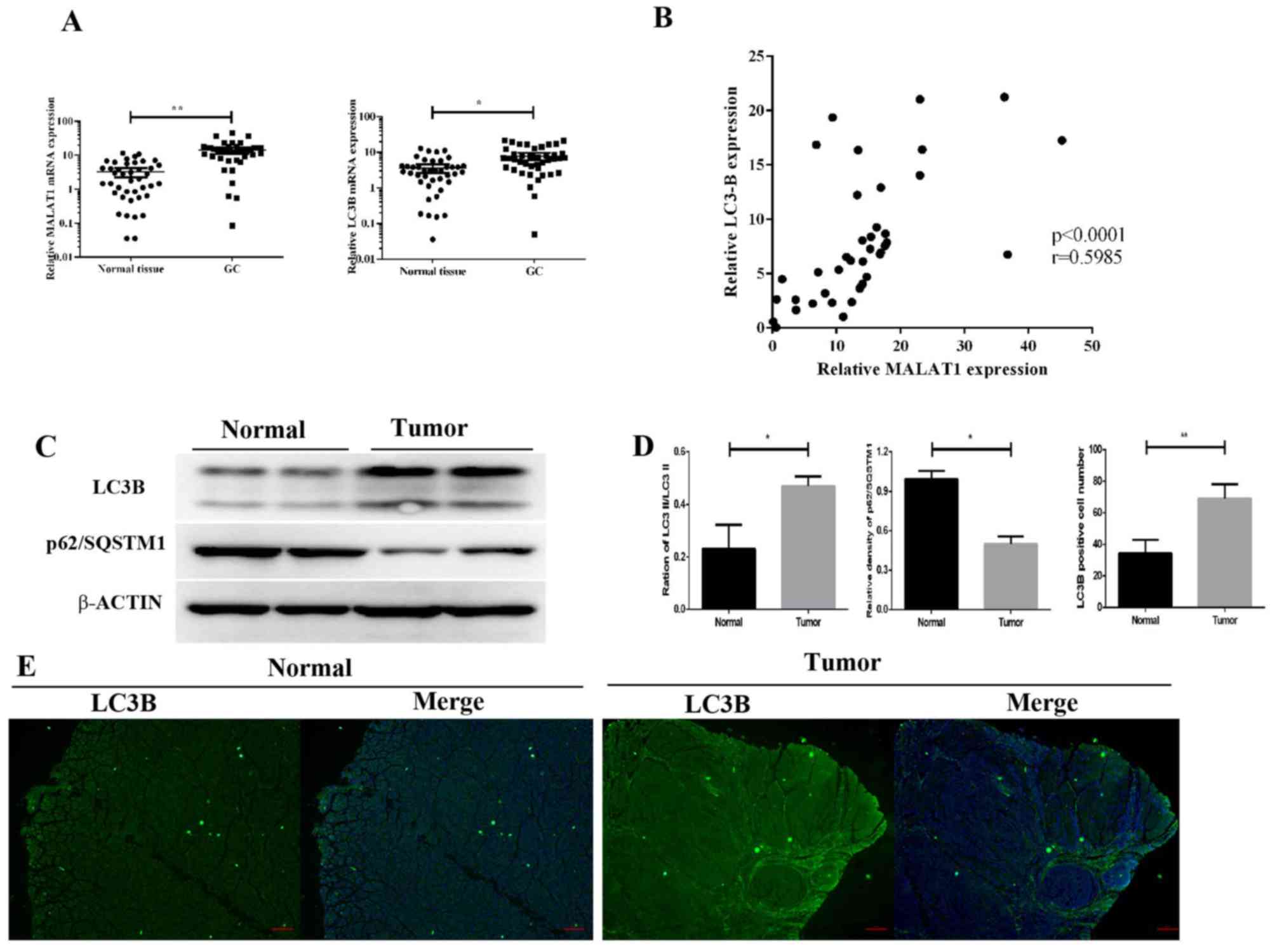
To examine the role of MALAT1 in GC and to
identify potential molecular events, RT-qPCR was used to detect
MALAT1 expression levels in 57 GC and paired non-tumorous
tissues. The results demonstrated that MALAT1 expression was
significantly increased in GC tissues compared with paired
non-tumorous tissues (P<0.01; Fig.
1A). As a structural protein of autophagosome membranes
(41), LC3B mRNA level was
upregulated in GC tumors (P<0.05; Fig. 1A). Furthermore, the mRNA level of
LC3B was positively correlated with MALAT1. As a
marker of autophagy, the ratio of LC3-II/LC3-I was also increased
in GC tissues compared with the control group (Fig. 1C and D). The results of the western
blotting analysis demonstrated that p62 expression was lower in GC
tissues compared with the control group (Fig. 1C and D). In addition, fluorescence
microscopy revealed that the number of FITC-LC3 puncta was higher
in GC tissues compared with controls (Fig. 1D and E). These results demonstrated
that upregulated MALAT1 levels were associated with
increased expression of autophagy markers in GC tissues.
MALAT1 enhances GC cell
proliferation
To examine whether MALAT1 may be associated
with GC cell proliferation, Ki67 expression levels were assessed in
clinical specimens by immunofluorescence analysis. Ki67 expression
levels in GC tissues were demonstrated to be higher compared with
the control group (Fig. 2A and B),
which indicated increased proliferation in GC samples.
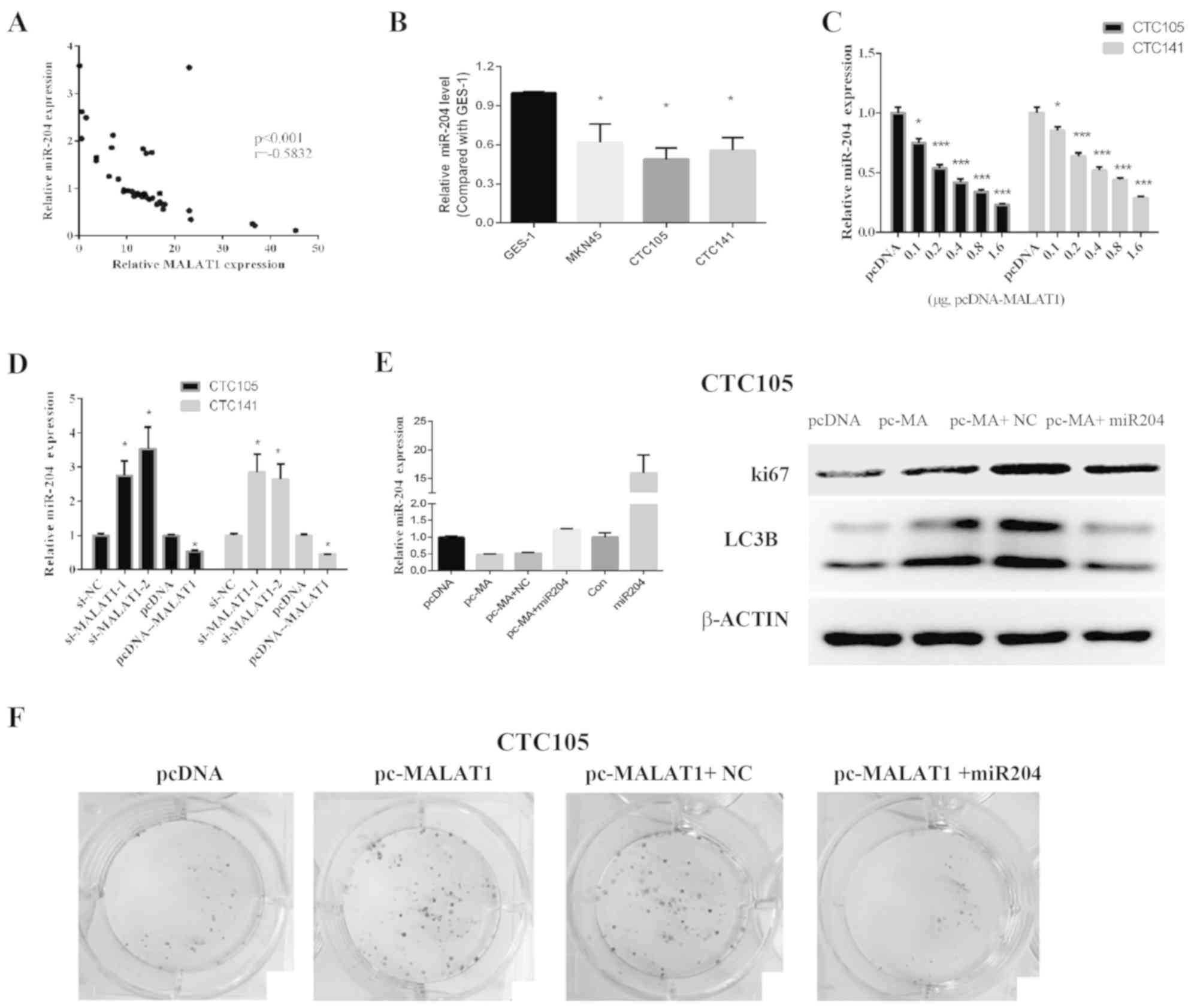
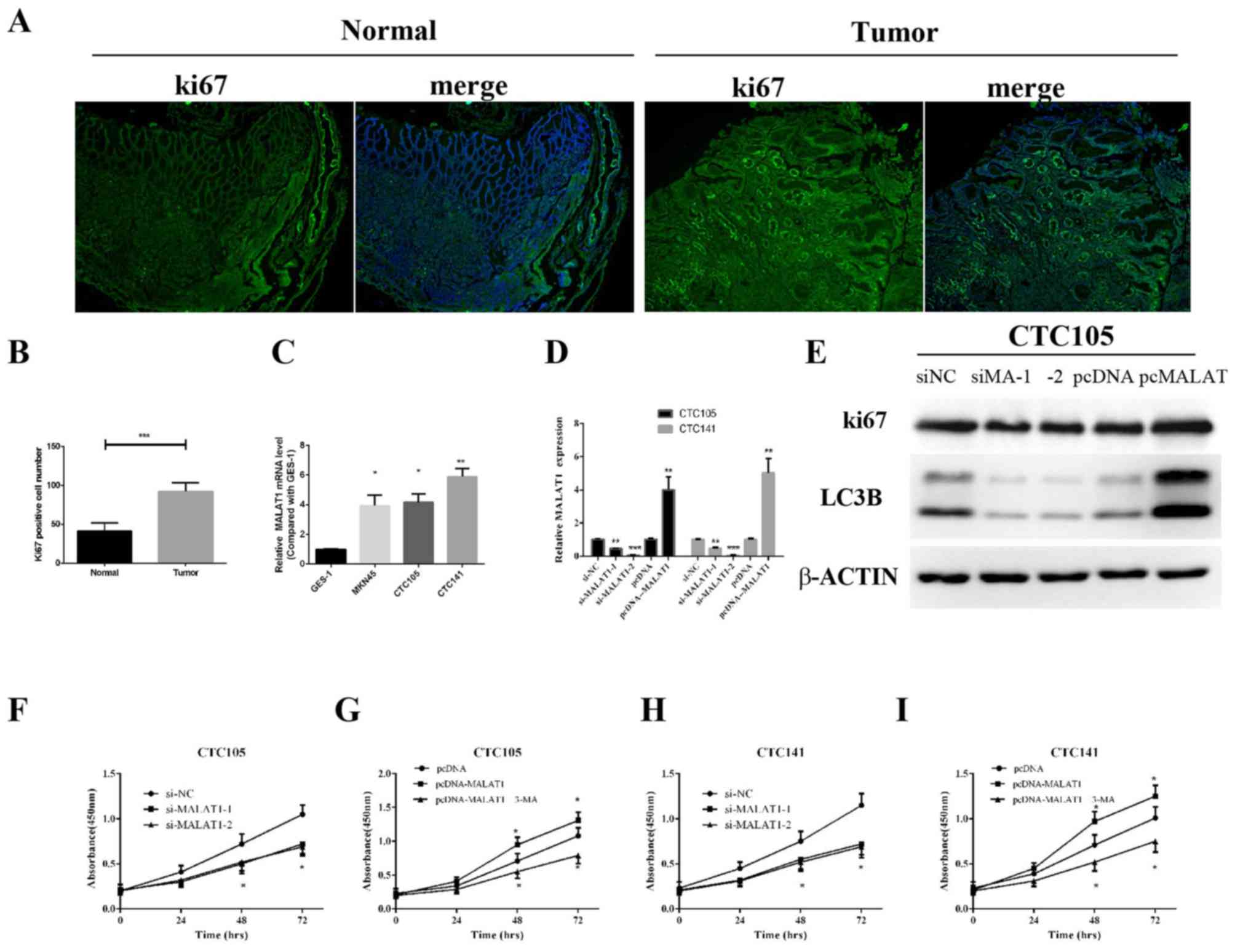 | Figure 2.MALAT1 promotes GC cell
proliferation. (A and B) Immunofluorescence detection of Ki-67 in
GC and non-tumorous tissues. (C) MALAT1 expression levels in
the normal human gastric epithelial cell line GES-1 and human GC
cell lines MKN45, CTC105 and CTC141 were measured by RT-qPCR. (D
and E) CTC105 and CTC141 cell lines were transfected with the
MALAT1 overexpression vector, MALAT1 siRNAs or the
indicated controls and examined by (D) RT-qPCR and (E) western
blotting. (F-I) CTC141 and CTC105 cells were transfected with (F
and H) MALAT1 siRNAs, (H and I) the MALAT1
overexpression vector or the indicated controls, and MTT assays
were performed at 0, 24, 48 and 72 h, with or without 3-MA (2 mM, 3
h). *P<0.05, **P<0.01 and ***P<0.001 vs. the corresponding
control. MA, MALAT1; MALAT1, metastasis-associated
lung adenocarcinoma transcript 1; NC, negative control; RT-qPCR,
reverse transcription-quantitative polymerase chain reaction; si,
small interfering. |
MALAT1 expression levels in GC cells lines
were evaluated, and RT-qPCR results demonstrated that MALAT1
expression was significantly upregulated in CTC105 and CTC141 cells
compared with normal gastric epithelial GES-1 cells (Fig. 2C). To evaluate the function of
MALAT1 in GC-derived circulating cells, MALAT1
overexpression plasmid and siRNAs targeting MALAT1 were
used. MALAT1 mRNA expression increase in cells transfected
with the MALAT1 overexpression plasmid and decrease in
si-MALAT1- and si-MALAT2-transfected cells were
confirmed (Fig. 2D). Similar results
were obtained by western blotting in CTC105 cells (Fig. 2E).
MALAT1 downregulation significantly decreased
CTC105 and CTC141 cell proliferation compared with controls
(Fig. 2F and H), whereas upregulated
MALAT1 enhanced CTC105 and CTC141 cell proliferation, and
the increased proliferation was inhibited by the autophagy
inhibitor, 3-methyladenine (3-MA) (Fig.
2G and I). These results demonstrated that MALAT1 may
increase GC cell proliferation through autophagy.
MALAT1 regulates miR-204 in GC
A previous study reported that MALAT1 exerted
its effects by binding and inhibiting miR-204 (36). The results from the present study
demonstrated that miR-204 and MALAT1 expression levels were
negatively correlated in GC tissues (Fig. 3A), which suggested that MALAT1
may inhibit miR-204 expression in GC. In addition, miR-204
expression levels in CTC105 and CTC141 cells were significantly
lower compared with those in GES-1 cells (Fig. 3B), and the levels or miR-204
decreased following MALAT1 overexpression (Fig. 3C). Furthermore, following
MALAT1 downregulation in CTC105 and CTC141 cells, miR-204
expression level was increased, whereas miR-204 expression level
was decreased in CTC105 and CTC141 cells overexpressing
MALAT1 (Fig. 3D).
Furthermore, MALAT1 overexpression increased
LC3B and Ki67 expression in CTC105 cells, which was abolished
following miR-204 mimic transfection (Fig. 3E). In addition, colony formation
assays indicated that MALAT1 promoted GC cell proliferation
and that miR-204 rescued the effect of MALAT1 on cancer
cells (Fig. 3F). These results
suggested that MALAT1 may induce miR-204 downregulation and
that MALAT1-induced cell proliferation and autophagy marker
upregulation was blocked by miR-204. These results suggested that
MALAT1 may work by negatively regulating miR-204.
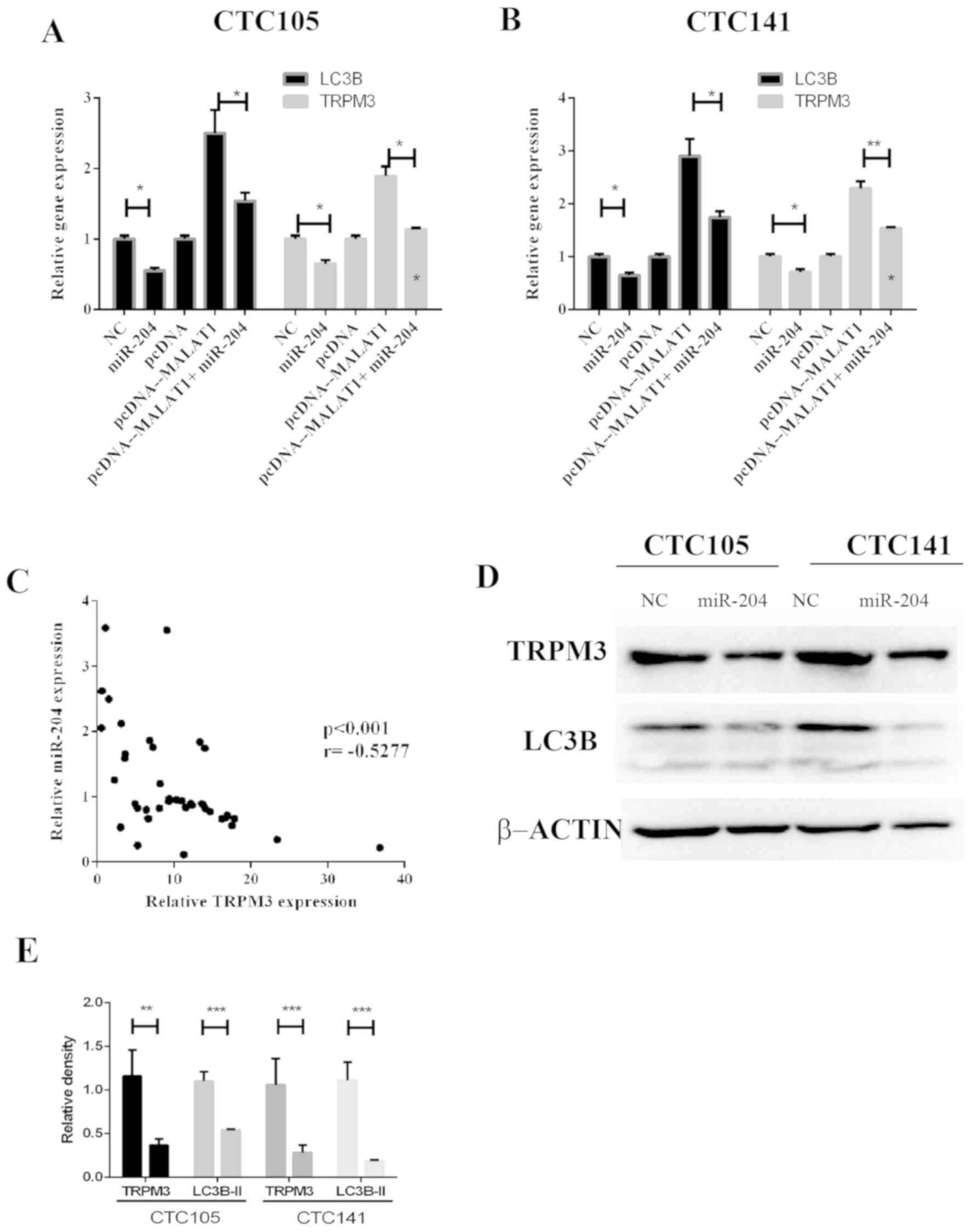
miR-204 inhibits MALAT1-induced
regulation of LC3B and TRPM3 in GC
The regulatory effects of MALAT1 and miR-204
on TRPM3, which is an autophagy activator, were evaluated.
The results demonstrated that MALAT1 increased LC3B
and TRPM3 mRNA expression levels, whereas miR-204
co-expression eliminated these effects in CTC105 (Fig. 4A) and CTC141 (Fig. 4B) cell lines. In addition,
TRPM3 and miR-204 expression levels were negatively
correlated GC tissues (Fig. 4C).
Western blotting demonstrated that LC3B and TRPM3 protein levels
were significantly decreased in CTC105 and CTC141 cells transfected
with the miR-204 mimic (Fig. 4D and
E).
Discussion
Numerous studies have demonstrated that lncRNAs
serve crucial roles in the development of various types of cancer
(16–21). However, the function of lncRNAs in GC
remains unclear. Identification of cancer-associated lncRNAs and
their targets is therefore critical for understanding their roles
in tumorigenesis and for the development of novel targets for GC
therapy. The present study investigated the role of lncRNA
MALAT1 in GC and its potential underlying mechanisms.
MALAT1 has recently been proposed as a marker
of GC (26). Previous studies have
reported that MALAT1 promotes GC cell proliferation and
metastasis (42,43). In addition, MALAT1 serves a
crucial role in the regulation of autophagy, and its overexpression
is associated with chemoresistance (44). Although these studies indicate that
MALAT1 is involved in GC progression and
autophagy-associated chemoresistance, the correlations between
MALAT1, autophagy and proliferation in vivo remain
unclear.
In the present study, the correlation between
MALAT1 expression levels and autophagy activation in GC cell
lines and tissues and the association between MALAT1
expression and GC proliferation were examined. The results
demonstrated that MALAT1 upregulation was associated with
increased autophagy in GC tissues. MALAT1 expression levels
were significantly increased in GC tissues, and LC3B
expression was proportional to the level of MALAT1. In
addition, expression levels of LC3B were also significantly higher
in GC tissues compared with non-tumor tissues. In the pcDNA
negative control group, LC3B was downregulated compared with the
siNC group, which may be due to endotoxin effect following plasmid
extraction that may have affected cell autophagy (45). However, this did not affect our
conclusion.
However, p62 expression was lower in GC tissues
compared with the control group, p62, which is a
scaffolding/adaptor protein, is involved in numerous physiological
processes, including inflammation, mitosis and autophagy and is a
crucial factor during tumorigenesis (46,47).
These findings demonstrated that upregulated MALAT1 was
associated with increased autophagy activation in GC. The results
observed in GC cell lines were consistent with these findings. In
addition, the present study demonstrated that MALAT1
overexpression increased GC cell proliferation.
As a target of MALAT1, miR-204 prevents tumor
development (33,36,37,48,49) and
regulates TRPM3-induced oncogenic autophagy (50,51). The
results of the present study revealed that the miR-204 mimic
reversed the effects of MALAT1 overexpression on LC3B
and TRPM3 in GC cells, which indicated that MALAT1
may exert its roles in GC by negatively regulating miR-204.
Furthermore, the results suggested that miR-204 may regulate the
expression of the autophagy markers LC3B and TRPM3 in GC cells. The
results of the present study demonstrated that MALAT1
overexpression resulted in decreased miR-204 levels, and that
MALAT1 and miR-204 expression levels were negatively
correlated in GC tissues, which further indicated that
MALAT1 may negatively regulate miR-204 in GC cells. Although
the MALAT1/miR-204/LC3B axis has been reported to
regulate autophagy in myocardial ischemia-reperfusion injury
(52), the effect of the
microenvironment on cell autophagy through this axis remains
unclear. Preliminary data from a mouse model of GC indicated that
this axis was involved in Helicobacter pylori-induced GC and
regulated the infection by causing autophagy (data not shown).
Identifying how the MALAT1/miR-204/LC3B axis
regulates the H. pylori-induced infection and tumorigenesis
through autophagy will be the aim of our further study.
The present study explored the molecular mechanism
of MALAT1 in the occurrence and development of GC. The
results may provide potential novel targets for the development of
molecular targeted therapy of GC.
Acknowledgements
The authors would like to thank Dr Xianming Mo
(Laboratory of Stem Cell Biology of Sichuan University) for
providing the cancer cell lines CTC105 and CTC141.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant. no. 81602636), the Wuxi
Health and Family Planning Commission Youth Research Foundation
(grant no. Q201720) and the Li Jie-shou Gut Barrier Foundation
(grant no. LJS-201708).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
GS, ZZ and WZ analyzed and interpreted the patient
data. GS, ZZ and GH performed the cellular and molecular
experiments, LZ, WL, CX and XZ analyzed the data. WZ and CX were
major contributors in writing the manuscript. All authors read and
approved the final version of the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Research
Ethics Committee of Jiangyin Hospital Affiliated to Nantong
University (approval no. JY1608AC97). All patients provided written
informed consent for the present study.
Patients consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Russo AE and Strong VE: Gastric cancer
etiology and management in Asia and the West. Annu Rev Med.
70:353–367. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Irino T, Takeuchi H, Terashima M, Wakai T
and Kitagawa Y: Gastric cancer in Asia: Unique features and
management. Am Soc Clin Oncol Educ Book. 37:279–291. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen L and Zhang K: Surgical treatment for
metastatic gastric cancer. Zhonghua Wei Chang Wai Ke Za Zhi.
20:731–734. 2017.(In Chinese). PubMed/NCBI
|
|
5
|
Li S and Le W: An insight review of
autophagy biology and neurodegenerative diseases: Machinery,
mechanisms and regulation. Sci China Life Sci. 60:1457–1459. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yin Z, Pascual C and Klionsky DJ:
Autophagy: Machinery and regulation. Microb Cell. 3:588–596. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fulda S: Targeting autophagy for the
treatment of cancer. Biol Chem. 399:673–677. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tian Y, Xu H, Farooq AA, Nie B, Chen X, Su
S, Yuan R, Qiao G, Li C, Li X, et al: Maslinic acid induces
autophagy by down-regulating HSPA8 in pancreatic cancer cells.
Phytother Res. 32:1320–1331. 2018. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu J, Feng L, Zhang H, Zhang J, Zhang Y,
Li S, Qin L, Yang Z and Xiong J: Effects of miR-144 on the
sensitivity of human anaplastic thyroid carcinoma cells to
cisplatin by autophagy regulation. Cancer Biol Ther. 19:484–496.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Song J, Zhou Y, Gong Y, Liu H and Tang L:
Rottlerin promotes autophagy and apoptosis in gastric cancer cell
lines. Mol Med Rep. 18:2905–2913. 2018.PubMed/NCBI
|
|
11
|
Liu D, Gao M and Zhao S: Autophagy as a
novel strategy for treatment of gastric cancer: A hypothesis. Med
Sci Monit. 19:794–796. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li LQ, Pan D, Zhang SW, Xie DY, Zheng XL
and Chen H: Autophagy regulates chemoresistance of gastric cancer
stem cells via the Notch signaling pathway. Eur Rev Med Pharmacol
Sci. 22:3402–3407. 2018.PubMed/NCBI
|
|
13
|
Bai XY, Liu YG, Song W, Li YY, Hou DS, Luo
HM and Liu P: Anticancer activity of tetrandrine by inducing
pro-death apoptosis and autophagy in human gastric cancer cells. J
Pharm Pharmacol. 70:1048–1058. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fan H, Jiang M, Li B, He Y, Huang C, Luo
D, Xu H, Yang L and Zhou J: MicroRNA-let-7a regulates cell
autophagy by targeting Rictor in gastric cancer cell lines MGC-803
and SGC-7901. Oncol Rep. 39:1207–1214. 2018.PubMed/NCBI
|
|
15
|
Li B, Wang W, Li Z, Chen Z, Zhi X, Xu J,
Li Q, Wang L, Huang X, Wang L, et al: MicroRNA-148a-3p enhances
cisplatin cytotoxicity in gastric cancer through mitochondrial
fission induction and cyto-protective autophagy suppression. Cancer
Lett. 410:212–227. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee J, Giordano S and Zhang J: Autophagy,
mitochondria and oxidative stress: Cross-talk and redox signalling.
Biochem J. 441:523–540. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fang Y and Fullwood MJ: Roles, functions,
and mechanisms of long non-coding RNAs in cancer. Genomics
Proteomics Bioinformatics. 14:42–54. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sun T: Long noncoding RNAs act as
regulators of autophagy in cancer. Pharmacol Res. 129:151–155.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang F, Bi J, Xue X, Zheng L, Zhi K, Hua J
and Fang G: Up-regulated long non-coding RNA H19 contributes to
proliferation of gastric cancer cells. FEBS J. 279:3159–3165. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang Y, Zhu M, Sun Y, Li W, Wang Y and Yu
W: Upregulation of lncRNA CASC2 suppresses cell proliferation and
metastasis of breast cancer via inactivation of the TGF-β signaling
pathway. Oncol Res. 27:379–387. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tsai KW, Lo YH, Liu H, Yeh CY, Chen YZ,
Hsu CW, Chen WS and Wang JH: Linc00659, a long noncoding RNA, acts
as novel oncogene in regulating cancer cell growth in colorectal
cancer. Mol Cancer. 17:722018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ji P, Diederichs S, Wang W, Böing S,
Metzger R, Schneider PM, Tidow N, Brandt B, Buerger H, Bulk E, et
al: MALAT-1, a novel noncoding RNA, and thymosin beta4 predict
metastasis and survival in early-stage non-small cell lung cancer.
Oncogene. 22:8031–8041. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ma J, Wu K, Liu K and Miao R: Effects of
MALAT1 on proliferation and apo- ptosis of human non-small cell
lung cancer A549 cells in vitro and tumor xenograft growth in vivo
by modulating autophagy. Cancer Biomark. 22:63–72. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li C, Cui Y, Liu LF, Ren WB, Li QQ, Zhou
X, Li YL, Li Y, Bai XY and Zu XB: High expression of long noncoding
RNA MALAT1 indicates a poor prognosis and promotes clinical
progression and metastasis in bladder cancer. Clin Genitourin
Cancer. 15:570–576. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang CJ, Shi SB, Tian J, Xu J and Niu ZX:
lncRNA MALAT1, HOTTIP and PVT1 as predictors for predicting the
efficacy of GEM based chemotherapy in first-line treatment of
pancreatic cancer patients. Oncotarget. 8:95108–95115.
2017.PubMed/NCBI
|
|
26
|
Xia H, Chen Q, Chen Y, Ge X, Leng W, Tang
Q, Ren M, Chen L, Yuan D, Zhang Y, et al: The lncRNA MALAT1 is a
novel biomarker for gastric cancer metastasis. Oncotarget.
7:56209–56218. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shuai F, Wang B and Dong S: MicroRNA-204
inhibits the growth and motility of colorectal cancer cells by
downregulation of CXCL8. Oncol Res. 26:1295–1305. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shu L, Zhang Z and Cai Y: MicroRNA-204
inhibits cell migration and invasion in human cervical cancer by
regulating transcription factor 12. Oncol Lett. 15:161–166.
2018.PubMed/NCBI
|
|
29
|
Shu Y, Ren L, Xie B, Liang Z and Chen J:
MiR-204 enhances mitochondrial apoptosis in doxorubicin-treated
prostate cancer cells by targeting SIRT1/p53 pathway. Oncotarget.
8:97313–97322. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shen SQ, Huang LS, Xiao XL, Zhu XF, Xiong
DD, Cao XM, Wei KL, Chen G and Feng ZB: MiR-204 regulates the
biological behavior of breast cancer MCF-7 cells by directly
targeting FOXA1. Oncol Rep. 38:368–376. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Canu V, Sacconi A, Lorenzon L, Biagioni F,
Lo Sardo F, Diodoro MG, Muti P, Garofalo A, Strano S, D'Errico A,
et al: MiR-204 down-regulation elicited perturbation of a gene
target signature common to human cholangiocarcinoma and gastric
cancer. Oncotarget. 8:29540–29557. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li T, Pan H and Li R: The dual regulatory
role of miR-204 in cancer. Tumour Biol. 37:11667–11677. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shrestha S, Yang CD, Hong HC, Chou CH, Tai
CS, Chiew MY, Chen WL, Weng SL, Chen CC, Chang YA, et al:
Integrated microRNA-mRNA analysis reveals miR-204 inhibits cell
proliferation in gastric cancer by targeting CKS1B, CXCL1 and
GPRC5A. Int J Mol Sci. 19(pii): E872017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen X, Liu XS, Liu HY, Lu YY and Li Y:
Reduced expression of serum miR-204 predicts poor prognosis of
gastric cancer. Genet Mol Res. 15:2016.
|
|
35
|
Ying Z, Li Y, Wu J, Zhu X, Yang Y, Tian H,
Li W, Hu B, Cheng SY and Li M: Loss of miR-204 expression enhances
glioma migration and stem cell-like phenotype. Cancer Res.
73:990–999. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li J, Wang J, Chen Y, Li S, Jin M, Wang H,
Chen Z and Yu W: LncRNA MALAT1 exerts oncogenic functions in lung
adenocarcinoma by targeting miR-204. Am J Cancer Res. 6:1099–1107.
2016.PubMed/NCBI
|
|
37
|
Tan X, Huang Z and Li X: Long non-coding
RNA MALAT1 interacts with miR-204 to modulate human hilar
cholangiocarcinoma proliferation, migration, and invasion by
targeting CXCR4. J Cell Biochem. 118:3643–3653. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen T, Yang K, Yu J, Meng W, Yuan D, Bi
F, Liu F, Liu J, Dai B, Chen X, et al: Identification and expansion
of cancer stem cells in tumor tissues and peripheral blood derived
from gastric adenocarcinoma patients. Cell Res. 22:248–258. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhao W, Chang C, Cui Y, Zhao X, Yang J,
Shen L, Zhou J, Hou Z, Zhang Z, Ye C, et al: Steroid receptor
coactivator-3 regulates glucose metabolism in bladder cancer cells
through coactivation of hypoxia inducible factor 1α. J Biol Chem.
289:11219–11229. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yin JJ, Liang B and Zhan XR: MicroRNA-204
inhibits cell proliferation in T-cell acute lymphoblastic leukemia
by down-regulating SOX4. Int J Clin Exp Pathol. 8:9189–9195.
2015.PubMed/NCBI
|
|
41
|
Bolanos JM, Moran AM, da Silva CM, Dávila
MP, Muñoz PM, Aparicio IM, Tapia JA, Ferrusola CO and Peña FJ:
During cooled storage the extender influences processed autophagy
marker light chain 3 (LC3B) of stallion spermatozoa. Anim Reprod
Sci. 145:40–46. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang J, Su L, Chen X, Li P, Cai Q, Yu B,
Liu B, Wu W and Zhu Z: MALAT1 promotes cell proliferation in
gastric cancer by recruiting SF2/ASF. Biomed Pharmacother.
68:557–564. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Qi Y, Ooi HS, Wu J, Chen J, Zhang X, Tan
S, Yu Q, Li YY, Kang Y, Li H, et al: MALAT1 long ncRNA promotes
gastric cancer metastasis by suppressing PCDH10. Oncotarget.
7:12693–12703. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
YiRen H, YingCong Y, Sunwu Y, Keqin L,
Xiaochun T, Senrui C, Ende C, XiZhou L and Yanfan C: Long noncoding
RNA MALAT1 regulates autophagy associated chemoresistance via
miR-23b-3p sequestration in gastric cancer. Mol Cancer. 16:1742017.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chung KW, Kim KM, Choi YJ, An HJ, Lee B,
Kim DH, Lee EK, Im E, Lee J, Im DS, et al: The critical role played
by endotoxin-induced liver autophagy in the maintenance of lipid
metabolism during sepsis. Autophagy. 13:1113–1129. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Moscat J and Diaz-Meco MT: p62 at the
crossroads of autophagy, apoptosis, and cancer. Cell.
137:1001–1004. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Moscat J and Diaz-Meco MT: p62: A
versatile multitasker takes on cancer. Trends Biochem Sci.
37:230–236. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Liu Z, Long J, Du R, Ge C, Guo K and Xu Y:
MiR-204 regulates the EMT by targeting snai1 to suppress the
invasion and migration of gastric cancer. Tumour Biol.
37:8327–8335. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Hou Z, Xu X, Zhou L, Fu X, Tao S, Zhou J,
Tan D and Liu S: The long non-coding RNA MALAT1 promotes the
migration and invasion of hepatocellular carcinoma by sponging
miR-204 and releasing SIRT1. Tumour Biol. 39:10104283177181352017.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hall DP, Cost NG, Hegde S, Kellner E,
Mikhaylova O, Stratton Y, Ehmer B, Abplanalp WA, Pandey R, Biesiada
J, et al: TRPM3 and miR-204 establish a regulatory circuit that
controls oncogenic autophagy in clear cell renal cell carcinoma.
Cancer Cell. 26:738–753. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Cost NG and Czyzyk-Krzeska MF: Regulation
of autophagy by two products of one gene: TRPM3 and miR-204. Mol
Cell Oncol. 2:e10027122015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wang S, Yu W, Luo X, Chen J and Deng F:
MALAT1/miR-204/LC3-II: A potential regulated axis of autophagy in
myocardial ischemia-reperfusion injury. Int J Cardiol. 277:2222019.
View Article : Google Scholar : PubMed/NCBI
|