Introduction
Lung cancer is one of the malignant tumors that
seriously threaten health and lives. The morbidity and mortality
rates of lung cancer rank first worldwide (1). Detection rate of early-stage lung
cancer is extremely low owing to the atypical symptoms and signs
(2). Non-small cell lung cancer
(NSCLC), includes squamous cell carcinoma and adenocarcinoma,
accounting for >80% of lung cancer cases (3). Current treatments for NSCLC, such as
surgery, radiotherapy and chemotherapy have advanced greatly. The
5-year survival of NSCLC, however, is as low as 15% (4). It is urgent to uncover the pathogenesis
of NSCLC, thus developing sensitive and effective approaches for
clinical treatment.
Long noncoding RNAs (lncRNAs) can not encode
proteins and are over 200 nucleotides in length. Functionally,
lncRNAs are closely linked with gene expression, embryonic
development, and systematic metabolism. Recently, vital roles of
lncRNAs in human diseases have been identified. They are capable of
regulating cellular behaviors, drug resistance and
epithelial-mesenchymal transition (EMT) (5–7).
Plasmacytoma variant translocation 1 (PVT1) is a recently
discovered specific lncRNA located on the human chromosome 8q24,
with a full length of 1716 bp. PVT1 is the downstream gene of
proto-oncogene MYC, and its transcription products are intronic
transcripts (8). PVT1 is upregulated
in many types of tumors, and closely related to tumor prognosis
(9,10).
MicroRNAs (miRNAs) are small noncoding RNAs 22–24 nt
in length. They are of significance in maintaining metabolic
functions. Through binding 3′-untranslated region (3′-UTR) of
target genes, miRNAs degrade them or suppress their translation,
thus downregulating target gene expression (11). Abnormally expressed miRNAs are
involved in tumorigenesis (12). It
is reported that miRNA-526b is downregulated in NSCLC.
Overexpression of miRNA-526b remarkably inhibits lung cancer cell
proliferation (13).
EZH2 locates on the human chromosome 7q35-7q36. It
spans 76,939 bp, and contains 20 exons and 19 introns (14). EZH2 participates in the histone
deacetylase (HDAC) process by interacting with DNA
methyltransferases (DNMT), which can affect the activity of
H3K27me3 (15). In breast cancer and
prostate cancer, EZH2 influences tumor progression by regulating
tumor growth and metastasis (16).
This study investigated the function of PVT1/miRNA-526b/EZH2
regulatory loop in the progression of NSCLC.
Patients and methods
Sample collection
NSCLC tissues and adjacent normal tissues were
collected from NSCLC patients undergoing radical resection in
Hainan Provincial People's Hospital (Haikou, China) from May 2016
to December 2018. All tissues were pathologically confirmed. None
of patients had preoperative antitumor treatment. This study was
approved by the Ethics Committee of Hainan Provincial People's
Hospital. Signed informed consents were obtained from all
participants before the study.
Cell culture and transfection
A549 cells were cultured in Dulbeccos modified eagle
medium (DMEM) containing 10% fetal bovine serum (FBS; both from
HyClone), 100 µg/ml penicillin and 0.1 mg/ml streptomycin, at 37°C,
in 5% CO2 incubator. Cells pre-seeded in a 24-well plate
were cultured at 70% confluence and transfected using Lipofactamine
2000 (Invitrogen; Thermo Fisher Scientific, Inc.). Six hours later,
complete medium was replaced. Sequences of transfection vectors
were as follows: Si-PVT1, forward, 5′-GCUUCUCCUGUUGCUGCATT-3′ and
reverse, 5′-UAGCAGCAACAGGAGAAGCTT-3′; Si-NC, forward,
5′-GCUACGAUCUGCCCAAGAUTT-3′ and reverse,
5′-AUCUUAGGCAFGAUCGUCGCTT-3′.
Quantitative real-time polymerase
chain reaction (qRT-PCR)
Extraction of total RNA in cells was performed using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.). RNAs
were subjected to reverse transcription. The extracted cDNA was
applied for PCR using SYBR-Green method. Primer sequences were as
follows: PVT1, forward, 5′-TGAGAACTGTCCTTACGTGACC-3′ and reverse,
5′-AGAGCACCAAGACTGGCTCT-3′; glyceraldheyde 3-phosphate
dehydrogenase (GAPDH), forward, 5′-GCACCGTCAAGGCTGAGAAC-3′ and
reverse, 5′-TGGTGAAGACGCCAGTGGA-3′; miRNA-526b, forward,
5′-CTTGCTTGGAAGGGGCATGCA-3′ and reverse,
5′-GTCGTATCCAGTGCAGGGTCCGAGGTA-3′; U6, forward,
5′-AGAAAATCTGCGCTTGGTCGTCC-3′ and reverse, 5′-TAG
CCGTGATATCGATGTAGCAA-3′. EZH2, forward,
5′-CGGGGTACCGAGTCATACTTGTGAAG-3′ and reverse,
5′-GCACTCGAGCCTGTTTTTGTTTGATG-3′.
Cell counting kit-8 (CCK-8)
Cells were seeded in a 96-well plate with
3×103 cells per well and cultured overnight. Absorbance
(A) at 450 nm was recorded at the appointed time using the CCK-8
kit (Dojindo Laboratories) for depicting the viability curves.
Wound closure assay
Cells were inoculated in a 25 mm2 culture
dish in serum-free medium. After cell adherence, a 10 µl pipette
tip was utilized for scratching an artificial wound. After 24-h
culture, wound closure was observed and captured (magnification,
×40).
Dual-luciferase reporter gene
assay
Wild-type and mutant-type vectors were constructed
based on the binding sites in the promoter regions of the genes.
Cells were co-transfected with wild-type/mutant-type vectors and
miRNA-526b mimic/control for 48 h. Then, cells were lysed for
determining luciferase activity.
Statistical analysis
Statistical Product and Service Solutions (SPSS)
22.0 (IBM Corp.) was used for data analyses. Data were expressed as
mean ± standard deviation. Intergroup differences were analyzed by
the t-test. Spearman correlation test was conducted to analyze the
relationship between PVT1 and miRNA-526b in NSCLC tissues.
P<0.05 was considered as statistically significant.
Results
Upregulation of PVT1 in NSCLC
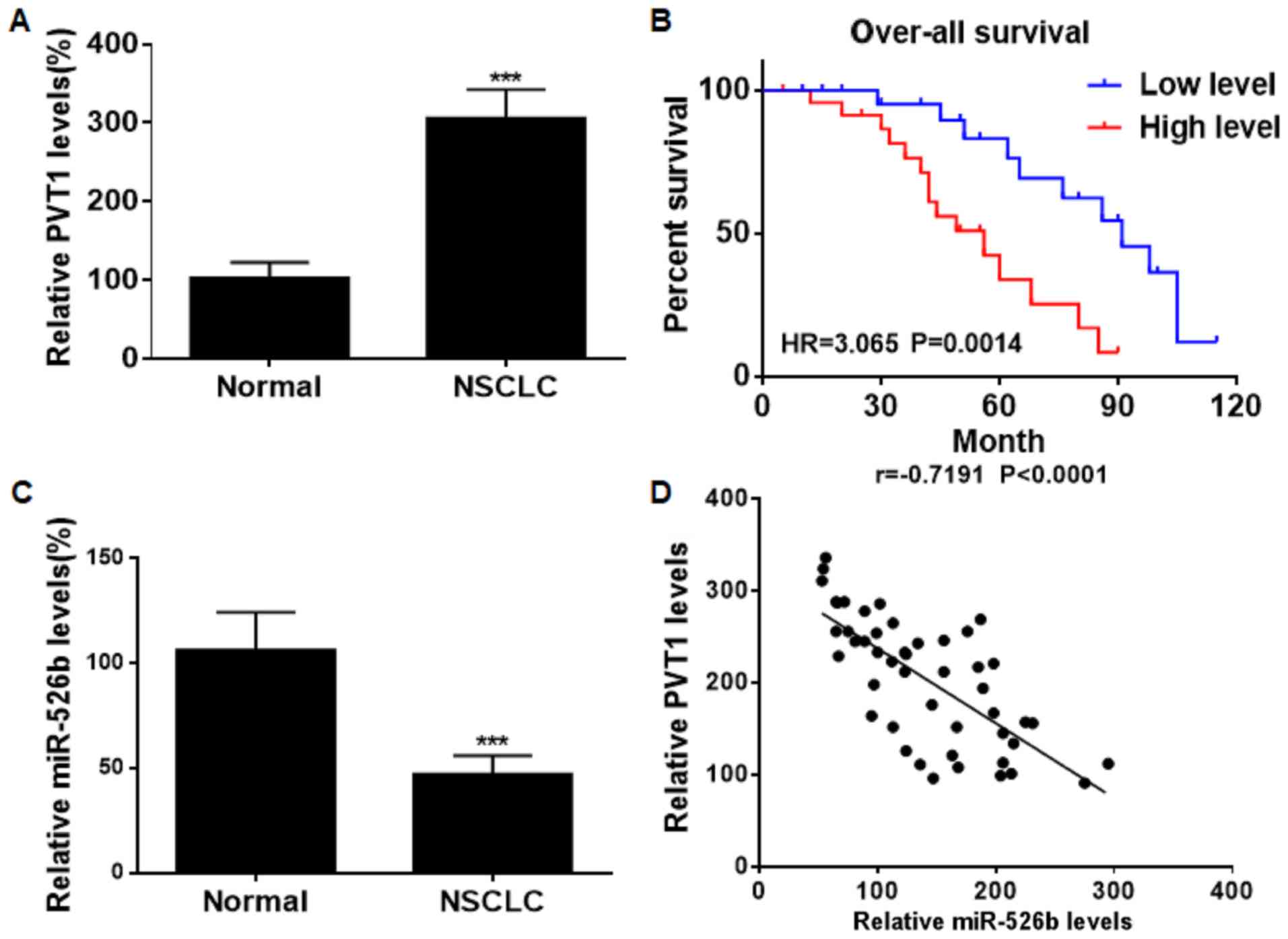
PVT1 was found to be upregulated in NSCLC tissues
relative to normal ones (Fig. 1A).
Kaplan-Meier analyses revealed that NSCLC patients expressing high
level of PVT1 suffered worse prognosis than those with low level
(Fig. 1B). On the contrary,
miRNA-526b was downregulated in NSCLC tissues (Fig. 1C). Spearman correlation test showed a
negative relationship between levels of PVT1 and miRNA-526b in
NSCLC tissues (Fig. 1D).
Silence of PVT1 attenuates viability
and migration in NSCLC
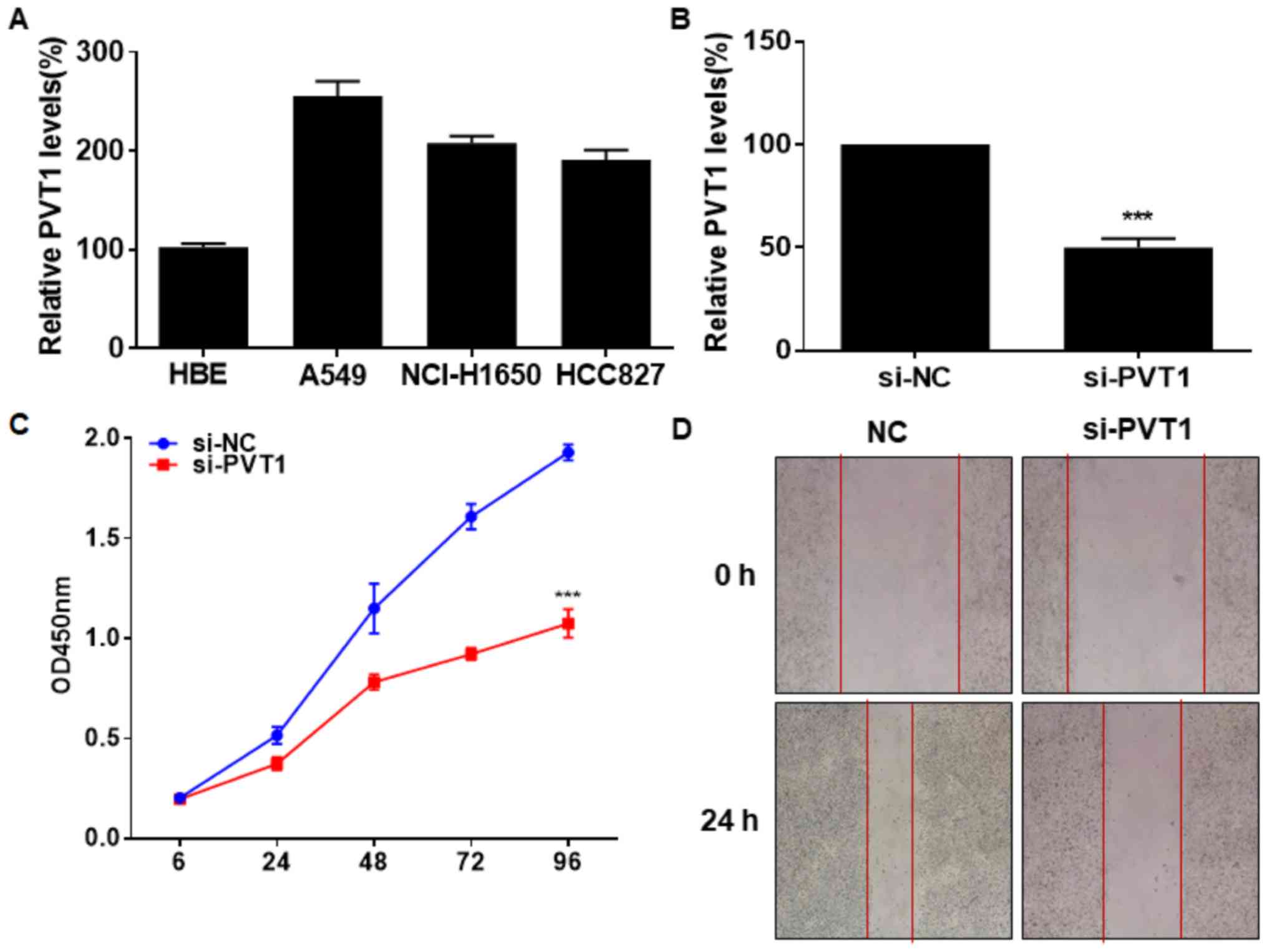
In vitro level of PVT1 was identically
upregulated in lung cancer cells and normal bronchial epithelial
cells (Fig. 2A). We constructed
si-PVT1 and tested its transfection efficacy in A549 cells
(Fig. 2B). CCK-8 assay indicated an
inhibited viability in A549 cells transfected with si-PVT1
(Fig. 2C). Silence of PVT1 also
reduced the wound closure percentage in A549 cells, suggesting
suppressed migratory ability (Fig.
2D). It is believed that PVT1 stimulated NSCLC to proliferate
and migrate.
PVT1 influences proliferative and
migratory abilities of NSCLC by targeting miRNA-526b
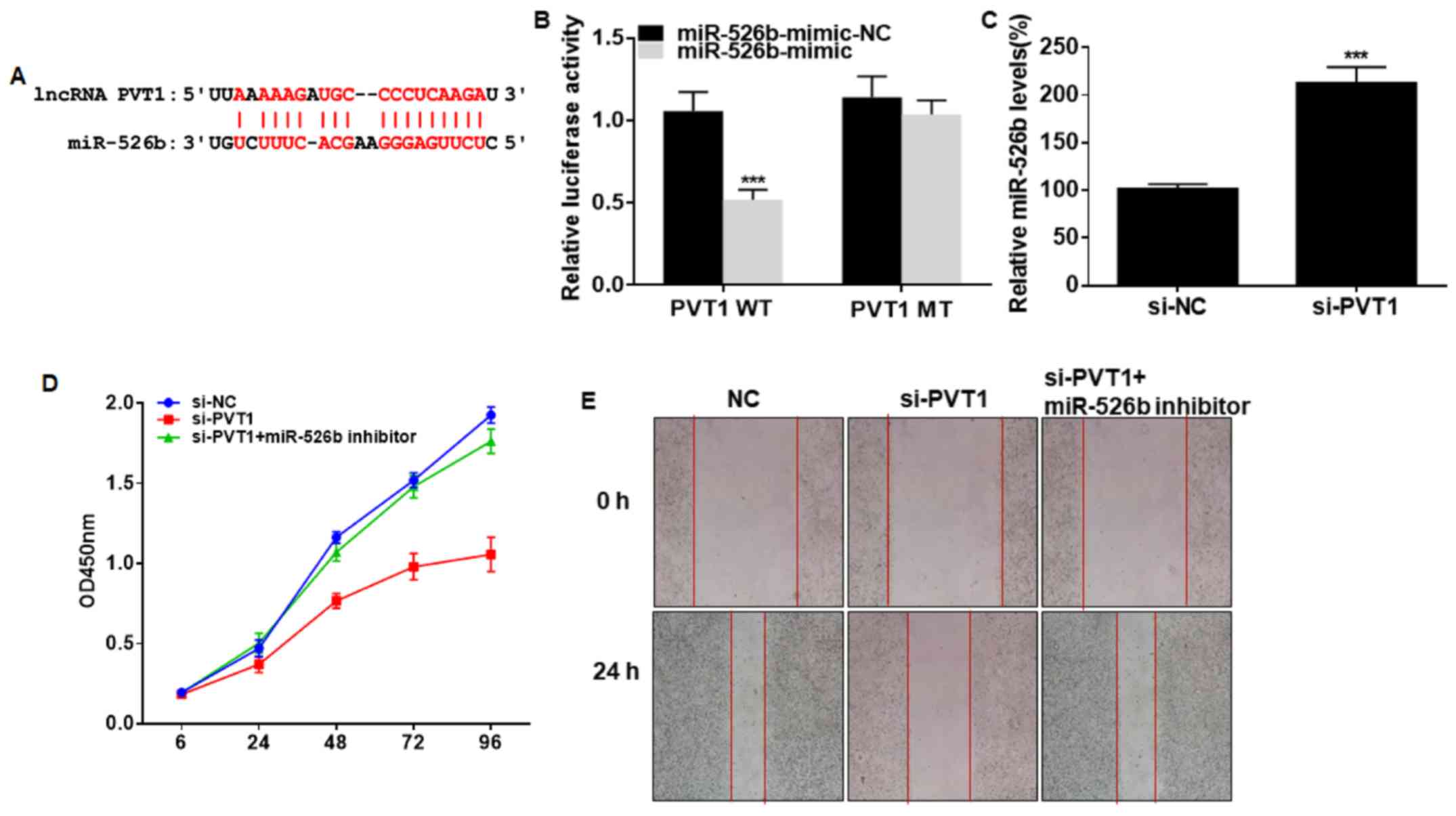
Potential binding sites were predicted in promoter
regions of PVT1 and miRNA-526b (Fig.
3A). Overexpression of miRNA-526b quenched the luciferase
activity in cells transfected with PVT1 WT, verifying the binding
between PVT1 and miRNA-526b (Fig.
3B). By transfection of si-PVT1, miRNA-526b level in A549 cells
was remarkably upregulated (Fig.
3C). Interestingly, the reduced viability and wound closure
percentage in A549 cells with PVT1 knockdown were partially
reversed after silencing of miRNA-526b (Fig. 3D and E). Hence, miRNA-526b was
responsible for the progression of NSCLC regulated by PVT1.
miRNA-526b influences proliferative
and migratory abilities of NSCLC by downregulating EZH2
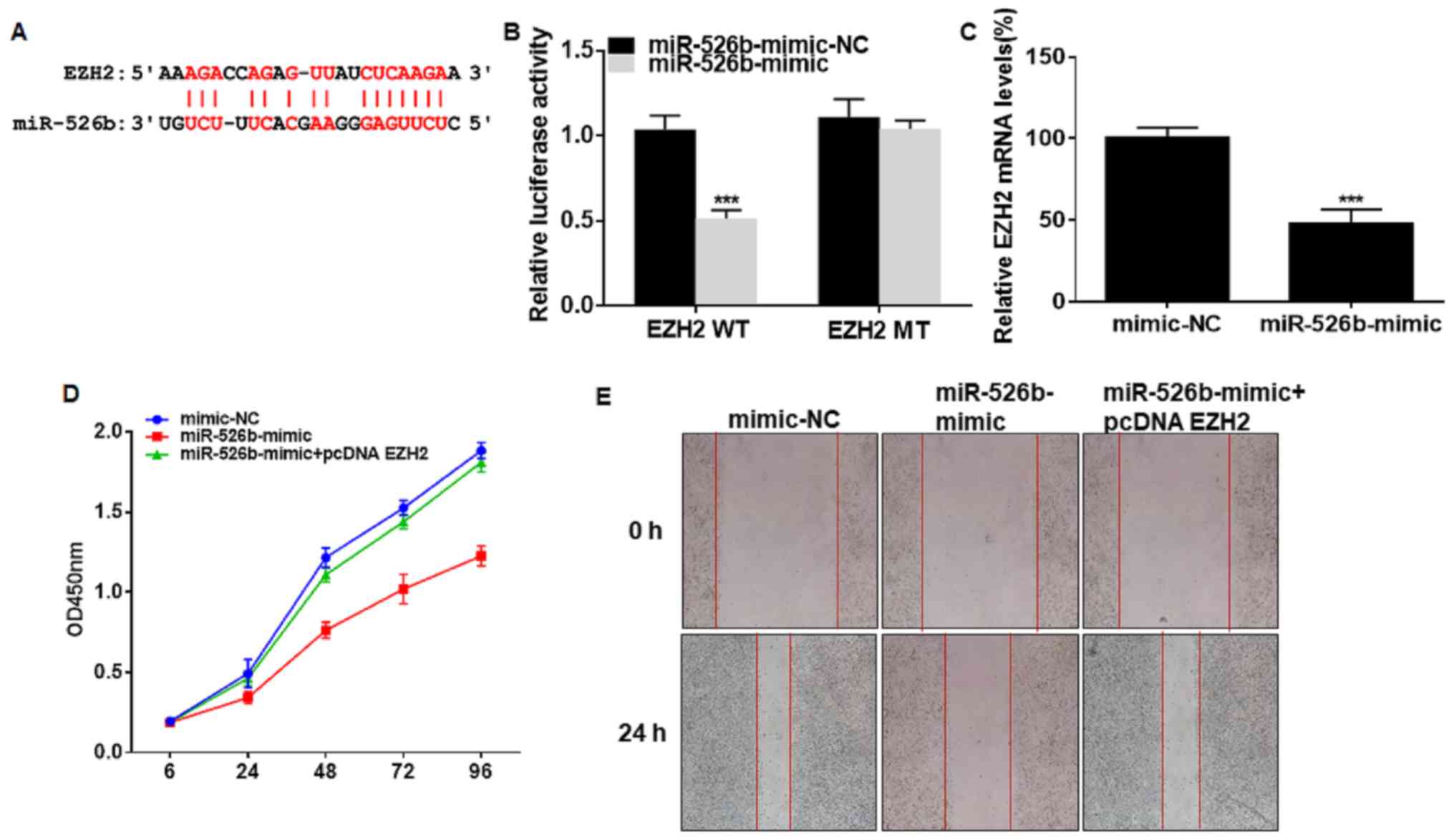
The presence of binding sites were predicted in
promoter regions of EZH2 and miRNA-526b (Fig. 4A). Dual-luciferase reporter gene
assay verified the online prediction that EZH2 was the downstream
target of miRNA-526b (Fig. 4B). EZH2
level was downregulated in A549 cells overexpressing miRNA-526b
(Fig. 4C). Transfection of
miRNA-526b mimic suppressed viability and wound closure percentage
in A549 cells, and were reversed by co-transfection of pcDNA EZH2
(Fig. 4D and E). As a result, EZH2
was involved in miRNA-526b-regulated progression of NSCLC.
Discussion
NSCLC is a fatal malignancy with high morbidity and
mortality rates. Most NSCLC patients miss the chance for surgery
due to the atypical symptoms in the early stage (17). Once tumor metastasis occurs, the
NSCLC patients cannot undergo radical resection. The prognosis of
NSCLC is very poor, leading to extremely high mortality (2). Target inhibition of metastasis is of
significance to improve the clinical outcome of advanced NSCLC
patients. Accumulating evidence has proposed that dysregulated
lncRNAs are closely related with the occurrence and progression of
NSCLC. These lncRNAs may be utilized as diagnostic, therapeutic and
prognostic markers for NSCLC (18,19). Zhu
et al (20) pointed out that
linc00312 is downregulated in NSCLC and exerts a tumor-suppression
effect. UCA1 is identified to be highly expressed in NSCLC tissues
(21). Silence of UCA1 suppresses
the malignant proliferative ability of NSCLC. Our findings
uncovered that PVT1 was upregulated in NSCLC tissues and cells.
Knockdown of PVT1 remarkably attenuated proliferative and migratory
abilities of lung cancer cells.
Regulatory mechanisms of lncRNAs in mediating cell
behavior mainly include: i) lncRNAs regulate promoter transcription
of target genes; ii) lncRNAs regulate activities of RNA polymerase
II; iii) lncRNAs and transcripts of target genes contribute to the
formation of complementary double strands; iv) lncRNAs regulate
protein activities by binding to them; v) lncRNAs alter the
intracellular localization of target proteins. Recently, a novel
regulatory mechanism was proposed. lncRNAs can silence target
miRNA-mediated gene expression through sponging these miRNAs, that
is, the ceRNA hypothesis (22).
Previous studies have discovered many miRNAs that are related to
NSCLC metastasis (23). Hence, we
speculated that PVT1 may serve as a ceRNA to target certain miRNAs,
thus influencing the progression of NSCLC. Through bioinformatics
prediction and dual-luciferase reporter gene assay verification,
miRNA-526b was confirmed to be the downstream target of PVT1.
Functional experiments confirmed that miRNA-526b was necessary for
PVT1-regulated NSCLC progression.
Upregulation of EZH2 in tumor diseases markedly
stimulates proliferative rate of tumor cells. During B cell
differentiation in lymphoid tissues, EZH2 exerts a vital function
(24). EZH2 is also a prognostic
marker. High level of EZH2 predicts worse prognosis in breast
cancer and bladder cancer (25). A
relevant study demonstrated that overexpression of miR-138 induced
apoptosis and arrested cell cycle in lung cancer cells by
downregulating EZH2 (26). In this
study, the presence of binding sites between EZH2 and miRNA-526b
was identified. Importantly, the attenuated viability and wound
closure ability in A549 cells overexpressing miRNA-526b were
reversed after EZH2 overexpression.
Collectively, PVT1/miRNA-526b/EZH2 regulatory loop
was discovered accelerating the malignant progression of NSCLC via
influencing tumor cell proliferation and migration. Our findings
provide new directions for clinical treatment of NSCLC.
In conclusion, PVT1 is upregulated in NSCLC, and
predicts a poor prognosis. PVT1 accelerates the progression of
NSCLC via targeting miRNA-526b/EZH2 regulatory loop.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors contributions
CQ, SL and SY designed the study and performed the
experiments, CQ and DS collected the data, SL and SY analyzed the
data, CQ, SL and SY prepared the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
This study was approved by the Ethics Committee of
Hainan Provincial People's Hospital (Haikou, China). Signed
informed consents were obtained from the patients and/or
guardians.
Patient consent for publication
Not applicable.
Competing interests
The authors declare they have no competing
interests.
References
|
1
|
Molina JR, Yang P, Cassivi SD, Schild SE
and Adjei AA: Non-small cell lung cancer: Epidemiology, risk
factors, treatment, and survivorship. Mayo Clin Proc. 83:584–594.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Benderli Cihan Y: ALK inhibitors and
cranial radiotherapy in brain metastasis from NSCLC. J BUON.
23:15582018.PubMed/NCBI
|
|
3
|
Yang B, Zheng D, Zeng Υ, Qin A, Gao J and
Yu G: Circulating tumor cells predict prognosis following
secondline AZD 9291 treatment in EGFR-T790M mutant non-small cell
lung cancer patients. J BUON. 23:1077–1081. 2018.PubMed/NCBI
|
|
4
|
Hu Z, Chen J, Tian T, Zhou X, Gu H, Xu L,
Zeng Y, Miao R, Jin G, Ma H, et al: Genetic variants of miRNA
sequences and non-small cell lung cancer survival. J Clin Invest.
118:2600–2608. 2008.PubMed/NCBI
|
|
5
|
Liu JY, Yao J, Li XM, Song YC, Wang XQ, Li
YJ, Yan B and Jiang Q: Pathogenic role of lncRNA-MALAT1 in
endothelial cell dysfunction in diabetes mellitus. Cell Death Dis.
5:e15062014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cheng N, Li X, Zhao C, Ren S, Chen X, Cai
W, Zhao M, Zhang Y, Li J, Wang Q, et al: Microarray expression
profile of long non-coding RNAs in EGFR-TKIs resistance of human
non-small cell lung cancer. Oncol Rep. 33:833–839. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Vikram R, Ramachandran R and Abdul KS:
Functional significance of long non-coding RNAs in breast cancer.
Breast Cancer. 21:515–521. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tseng YY, Moriarity BS, Gong W, Akiyama R,
Tiwari A, Kawakami H, Ronning P, Reuland B, Guenther K, Beadnell
TC, et al: PVT1 dependence in cancer with MYC copy-number increase.
Nature. 512:82–86. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ding C, Yang Z, Lv Z, Du C, Xiao H, Peng
C, Cheng S, Xie H, Zhou L, Wu J, et al: Long non-coding RNA PVT1 is
associated with tumor progression and predicts recurrence in
hepatocellular carcinoma patients. Oncol Lett. 9:955–963. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang YR, Zang SZ, Zhong CL, Li YX, Zhao SS
and Feng XJ: Increased expression of the lncRNA PVT1 promotes
tumorigenesis in non-small cell lung cancer. Int J Clin Exp Pathol.
7:6929–6935. 2014.PubMed/NCBI
|
|
11
|
Volinia S, Calin GA, Liu CG, Ambs S,
Cimmino A, Petrocca F, Visone R, Iorio M, Roldo C, Ferracin M, et
al: A microRNA expression signature of human solid tumors defines
cancer gene targets. Proc Natl Acad Sci USA. 103:2257–2261. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pekarsky Y, Santanam U, Cimmino A,
Palamarchuk A, Efanov A, Maximov V, Volinia S, Alder H, Liu CG,
Rassenti L, et al: Tcl1 expression in chronic lymphocytic leukemia
is regulated by miR-29 and miR-181. Cancer Res. 66:11590–11593.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang ZY, Fu SL, Xu SQ, Zhou X, Liu XS, Xu
YJ, Zhao JP and Wei S: By downregulating Ku80, hsa-miR-526b
suppresses non-small cell lung cancer. Oncotarget. 6:1462–1477.
2015.PubMed/NCBI
|
|
14
|
Tsang DP and Cheng AS: Epigenetic
regulation of signaling pathways in cancer: Role of the histone
methyltransferase EZH2. J Gastroenterol Hepatol. 26:19–27. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tonini T, DAndrilli G, Fucito A, Gaspa L
and Bagella L: Importance of Ezh2 polycomb protein in tumorigenesis
process interfering with the pathway of growth suppressive key
elements. J Cell Physiol. 214:295–300. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Riquelme E, Behrens C, Lin HY, Simon G,
Papadimitrakopoulou V, Izzo J, Moran C, Kalhor N, Lee JJ, Minna JD,
et al: Modulation of EZH2 expression by MEK-ERK or PI3K-AKT
signaling in lung cancer is dictated by different KRAS oncogene
mutations. Cancer Res. 76:675–685. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang J and Li H: CircRNA circ_0067934
silencing inhibits the proliferation, migration and invasion of
NSCLC cells and correlates with unfavorable prognosis in NSCLC. Eur
Rev Med Pharmacol Sci. 22:3053–3060. 2018.PubMed/NCBI
|
|
18
|
Yang J, Lin J, Liu T, Chen T, Pan S, Huang
W and Li S: Analysis of lncRNA expression profiles in non-small
cell lung cancers (NSCLC) and their clinical subtypes. Lung Cancer.
85:110–115. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wei MM and Zhou GB: Long non-coding RNAs
and their roles in non-small-cell lung cancer. Genomics Proteomics
Bioinformatics. 14:280–288. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhu Q, Lv T, Wu Y, Shi X, Liu H and Song
Y: Long non-coding RNA 00312 regulated by HOXA5 inhibits tumour
proliferation and promotes apoptosis in non-small cell lung cancer.
J Cell Mol Med. 21:2184–2198. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nie W, Ge HJ, Yang XQ, Sun X, Huang H, Tao
X, Chen WS and Li B: lncRNA-UCA1 exerts oncogenic functions in
non-small cell lung cancer by targeting miR-193a-3p. Cancer Lett.
371:99–106. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chan JJ and Tay Y: Noncoding RNA: RNA
regulatory networks in cancer. Int J Mol Sci. 19:192018. View Article : Google Scholar
|
|
23
|
Legras A, Pécuchet N, Imbeaud S, Pallier
K, Didelot A, Roussel H, Gibault L, Fabre E, Le Pimpec-Barthes F,
Laurent-Puig P, et al: Epithelial-to-mesenchymal transition and
microRNAs in lung cancer. Cancers (Basel). 9:92017. View Article : Google Scholar
|
|
24
|
Viré E, Brenner C, Deplus R, Blanchon L,
Fraga M, Didelot C, Morey L, Van Eynde A, Bernard D, Vanderwinden
JM, et al: The Polycomb group protein EZH2 directly controls DNA
methylation. Nature. 439:871–874. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Martinez-Garcia E and Licht JD:
Deregulation of H3K27 methylation in cancer. Nat Genet. 42:100–101.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang H, Zhang H, Zhao M, Lv Z, Zhang X,
Qin X, Wang H, Wang S, Su J, Lv X, et al: MiR-138 inhibits tumor
growth through repression of EZH2 in non-small cell lung cancer.
Cell Physiol Biochem. 31:56–65. 2013. View Article : Google Scholar : PubMed/NCBI
|