Introduction
Thyroid cancer incidence is rapidly increasing in
USA, with the estimated annual diagnoses and mortalities in 2017
being 56,870 and 2010, respectively (1). Thyroid cancer is generally classified as
papillary, follicular and anaplastic carcinomas. Amongst these
classifications, anaplastic thyroid cancer (ATC) was reported in
2012 to account for 1–2% of all thyroid tumor cases in the US
(2). It is characterized by
aggressive and local invasion, and frequent distant metastasis.
Currently, available therapies for ATC include chemotherapy,
radiotherapy and surgery (2).
However, ATC remains one of the most fatal cancer types, with a
mean survival time of 6 months (2).
Therefore, it is crucial to identify the molecular etiology and
molecular mechanisms underlying the progression and metastasis in
ATC, and thus improve the therapeutic strategies and prognosis.
Previously, microRNA (miRNA), a series of small,
highly-conserved, non-coding RNA molecules 18–25 nucleotides in
length, are known to activate or inhibit the progression of various
cancer types and have been proposed as novel targets for anticancer
therapies (3). Of all miRNAs, >50%
are known to be involved in human tumorigenesis by directly
targeting oncogenes or tumor suppressor genes (3). For instance, miR-125b inhibits the tumor
growth by directly targeting phosphatidylinositol-4.5-biphosphate
3-kinase catalytic subunit δ in ATC (4). Additionally, miR-4295 serves as an
oncogene and promotes cell proliferation and invasion in ATC via
cyclin dependent kinase inhibitor 1A (5). Previously, a number of studies have
reported that miR-544 was significantly downregulated or
upregulated in a number of human cancer types (6–10).
However, miR-544 may serve as an oncogene or tumor suppressor
depending on the type of tissue and the context in which it is
expressed (6–10). In ATC, the expression pattern,
biological roles and potential molecular mechanism of miR-544
remain largely unknown.
In the present study, the expression level of
miR-544 was firstly detected in ATC tissues and cell lines.
Secondly, the biological function of miR-544 in ATC proliferation,
migration and invasion in vitro and in vivo was
investigated. Finally, Yin Yang-1 (YY1) was identified as a direct
target of miR-544. The results revealed that targeting the
miR-544/YY1 axis may represent a promising therapeutic strategy for
ATC treatment.
Materials and methods
Cell culture and tissue
collections
The ATC cell lines (SW1736, KAT-18 and 8305C) and
immortal thyroid cell line Nthy-ori3-1 were purchased from the
American Type Culture Collection (Manassas, VA, USA). The cell
lines were authenticated using short-tandem repeat profiling, which
was performed by BMR Genomics (Padova, Italy). The cells were
maintained in Dulbecco's modified Eagle's medium (DMEM) with 10%
fetal bovine serum (FBS) (both from HyClone; GE Healthcare Life
Sciences, Logan, UT, USA). Cells were incubated in a humidified
atmosphere containing 5% CO2 and humidified sphere of
95% humidity at 37°C. Human ATC specimens and their adjacent normal
thyroid tissues (40 pairs) were collected from 15 males and 25
female patients (mean, 62 years; range, 34–72 years) who underwent
surgery between January 2016 and July 2017, according to an
approved human protocol at the Yantai Laiyang Central Hospital
(Yantai, China) and were used to detect the expression of miR-544
and mRNA expression of YY1. The present study was approved by the
Ethics Committee of Yantai Laiyang Central Hospital. Written
informed consent was obtai ned from every patient.
Cell transfection
The miR-544 mimic (sense,
5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACGAACTTT-3′) and the
negative control (miR-NC; sense, 5′-ACUACUGAGUGACAGUAGA-3′) were
purchased from Ambion (Thermo Fisher Scientific, Inc., Waltham, MA,
USA). The inhibitor control (anti-miR-NC;
5′-CAGUACUUUUGUGUAGUACAA-3′) was purchased from Ambion (Thermo
Fisher Scientific, Inc.). The miR-544 inhibitor was obtained from
Guangzhou RiboBio Co., Ltd. (Guangzhou, China). The following
sequences were used: 5′-CUUGUUAAAAAGCAGAUUCU-3′. The small RNAs,
including small interfering (si)-YY1 and sicontrol, were obtained
from Santa Cruz (Santa Cruz Biotechnology, Inc., Dallas, TX, USA).
The small RNA sequences are as follows:
siYY1-5′-GACGACUACAUUGAACAATT-3; negative control
RNA-5′-UUCUCCGAACGUGUCACGUTT-3. YY1 overexpression plasmid was
achieved using pcDNA3.1/YY1 transfection. A total of 10 nmol of
YY1-pcDNA3.1 was transfected into the cells. Phblv-u6-puro vectors
was purchased from Han Heng Biotechnology Co., Ltd. (Shanghai,
China). For cell transfection, SW1736 and 8305C cells
(2×105) were seeded in six-well plates and cultured
until 60% confluency was reached. Transfection was performed with
Lipofectamine® 2000 Reagent (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. The
transfection mixture was replaced in a medium containing 10% FBS
following 6–8 h and after 48 h the transfection efficiency was
detected by reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) and western blot.
Cell Counting Kit-8 (CCK-8)
A CCK-8 assay (Beyotime Institute of Biotechnology,
Haimen, China) was used to detect the cell viability. Briefly,
SW1736 and 8305C cells were seeded into 96-well plates
(1×103/well) in DMEM containing 10% FBS for 0, 24, 48,
72 h at 37°C. At the indicated time 10 µl CCK-8 was added to each
well. After incubation for 3 h at room temperature, the absorbance
of each well was measured using Multiskan MK3.
Colony formation assay
For the colony formation assay, 4×102
SW1736 and 8305C cells were seeded in 6-well plates separately.
After 10 days, the cells were washed with PBS three times, fixed
with 4% paraformaldehyde for 15 min and stained with 0.5% crystal
violet at room temperature (Beyotime Institute of Biotechnology).
The clone number (cell population >50) was counted using a CKX41
light microscope (magnification, ×10).
Dual-luciferase assay
The fragment of the YY1 3′-untranslated region (UTR)
containing the miR-544 predicted binding sequences (predicted by
targetscan 7.1) or the mutant sequences was synthesized by Shanghai
GenePhama Co., Ltd., and then cloned into a psiCHECK-2 vector
(Promega Corporation, Madison, WI, USA). For convenience, the
psiCHECK-2-YY1-3 tUTR-wild-type (Wt) and the psiCHECK-2-YY1-3
tUTR-wild-type (Mut) were designated as Wt and Mut, respectively.
SW1736 cells were co-transfected with the aforementioned luciferase
reporter vector with miR-544 mimic or miR-NC. 8305C cells were
co-transfected with the aforementioned luciferase reporter vector
with miR-544 inhibitor or anti-miR-NC. Cell transfection were
performed using Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. Cells
were cultured 37°C for 24 h and luciferase activities were analyzed
using the Dual-Luciferase Reporter Assay system. Results were
normalized to Renilla luciferase activity.
Wound-healing assay
The wound-healing assay was performed to assess cell
migration. In brief, SW1736 and 8305C cells seeded in six-well
plates (8×105 cells/well) and cultured until they
reached 90% confluency at 37°C. The cell monolayer was subsequently
scraped with a sterile 200 µl micropipette tip to create separate
wounds, and the wells were washed three times with PBS to remove
cell debris. Finally, the cells were cultured at 37°C and observed
at 0 and 24 h using a CKX41 light microscope (magnification,
×10).
Transwell and Matrigel assays
Migration and invasion abilities were analyzed using
Transwell chambers (Corning Incorporated, Corning, NY, USA). For
the invasion assays, the Transwell inserts were coated with 50 µl
Matrigel (BD Biosciences, Franklin Lakes, NJ, USA). For the assays,
5×104 SW1736 and 8305C cells were resuspended in 0.1 ml
serum-free DMEM and added to the Transwell inserts. DMEM with 10%
FBS was added to the lower wells. After incubation for 8 h
(migration) or 12 h (invasion), the cells on the upper surface of
the membrane were removed, and the cells on the lower surface were
fixed with 10% methanol at room temperature for 15 min and stained
with 0.1% crystal violet (Beyotime Institute of Biotechnology) at
room temperature for 5 min and then counted under a light
microscope (10 × magnification).
Total mRNA extraction RT-qPCR
The SW1736 and 8305C cells were dissolved in
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) to extract the total mRNA. Following spectrophotometric
quantification, cDNA was synthesized using an iScript cDNA
Synthesis kit (Takara Biotechnology Co., Ltd., Dalian, China),
according to the manufacturer's protocol. Subsequently, qPCR was
performed using SYBR® Premix Ex Taq™ (Takara
Biotechnology Co., Ltd.), according to the manufacturer's protocol.
For detecting the expression of YY1, β-actin acted as internal
control. For detecting the expression of miR-544, U6 served as
internal control. The thermocycling conditions were as follows:
95°C for 10 min followed by 50 cycles of 95°C for 10 sec, 55°C for
10 sec, 72°C for 5 sec; 99°C for 1 sec; 59°C for 15 sec; 95°C for 1
sec; followed by cooling to 40°C. The relative expression level of
miR-544 and YY1 was calculated as the inverse log of ∆∆Cq and
normalized to the reference (11).
The primers used for amplification were: YY1, forward,
5′-CCCCGGCAAGTGTGAGTGAA-3′, and reverse,
5′-ACAAGCCAAGGTCCTGCTGG-3′; β-actin, forward,
5′-GATCATTGCTCCTCCTGAGC-3′, and reverse,
5′-ACTCCTGCTTGCTGATCCAC-3′; miR-544, forward,
5′-GCCCGATTCTGCATTTTTAGC-3′, and reverse
5′-CGGGCTAAGACGTAAAAACG-3′; and U6, forward,
5′-TGCGGGTGCTCGCTTCGCAGC-3′, and reverse,
5′-CCAGTGCAGGGTCCGAGGT-3′.
RNA immunoprecipitation
RNA immunoprecipitation assays were performed using
an Imprint RNA Immunoprecipitation kit (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany), according to the manufacturer's protocol with
the protein argonaute-2 (AGO2; dilution, 1:100; cat. no. 2897; Cell
Signaling Technology, Inc., Danvers, MA, USA) or IgG antibodies
(dilution, 1:100; cat. no. A6066; Sigma-Aldrich; Merck KGaA) for 6
h at 4°C. The expression of miR-544 and the YY1 mRNA levels in the
immunoprecipitates were analyzed with RT-qPCR analysis, according
to the aforementioned protocol.
Western blot analysis
Cells were lysed using radioimmunoprecipitation
assay buffer (Beijing Solarbio Bioscience and Technology Co., Ltd.,
Beijing, China) and protein was quantified using a bicinchoninic
acid protein assay kit (Thermo Fisher Scientific, Inc.). Identical
amounts of proteins (20 µg) from the lysates of the SW1736 and
8305C cells were subjected to electrophoresis through 10% SDS-PAGE
and were transferred onto polyvinylidene difluoride membranes
(Invitrogen; Thermo Fisher Scientific, Inc.). The membranes were
blocked with 5% skimmed milk for 30 min at 4°C and incubated with
primary antibodies for YY1 (dilution, 1:200; cat. no. ab12132;
Abcam, Cambridge, MA, USA), E-cadherin (dilution, 1:500; cat. no.
sc-8426; Santa Cruz Biotechnology, Inc.), N-cadherin (dilution,
1:500; cat. no. sc-8424; Santa Cruz Biotechnology, Inc.) or β-actin
(dilution, 1:500; cat. no. AF0003; Beyotime Institute of
Biotechnology) overnight at 4°C, followed by incubation with
horseradish peroxidase-conjugated goat anti-rabbit antibody
(dilution, 1:2,000; cat. no. sc-2004; Santa Cruz Biotechnology,
Inc.) at room temperature for 2 h. Signals were detected with
Enhanced Chemiluminescent Plus (Beyotime Institute of
Biotechnology), according to the manufacturer's protocols. The
relative protein levels were calculated based on β-actin as the
loading control.
Xenografted tumor model
A total of 24 female, 4-week old BALB/c nude mice
(weight, range; 20–25 g) purchased from Jilin University
(Changchun, China) were inoculated subcutaneously with
1×106 SW1736 cells infected with lentiviral-miR-544
mimics or lentiviral-miR-NC in the dorsal flank and maintained in a
specific-pathogen free environment with constant humidity (45–50%)
and constant temperature (25–27°C) under a 12/12 h light/dark cycle
with free access to food and water. Measurements were obtained from
the tumors every 5 days, including the greatest tumor length and
width measurements with calipers, and then the tumor volumes were
analyzed. Tumor volume was calculated using the equation:
Volume=(length × width2)/2. After 30 days, the animals
were sacrificed by cervical dislocation, and tumors were excised
and weighed. The short hairpin (sh) YY1 and shControl transfected
into the SW1736 cells was carried out as previously reported. To
determine the function of YY1 in the growth of tumors in
vivo, stable lentiviral siYY1- and siControl-SW1736 cells
(1×106) were subcutaneously injected into the dorsal
flank of athymic nude mice (n=6/group), studied as aforementioned
statement. All animal studies were approved by the Animal Care and
Welfare Committee of Yantai Laiyang Central Hospital.
Bioinformatics analysis
The prediction of the YY1 3′-UTR as a miR-544
binding target was determined using TargetScan 7.1 software
(www.targetscan.org) (12). TargetScan is online software, that
predicts biological targets of miRNA by searching for the presence
of conserved 8mer, 7mer, and 6mer sites that match the seed region
of each miRNA.
Statistical analysis
Data are expressed as mean ± standard error.
Statistical analysis was conducted using SPSS 13.0 software (SPSS,
Inc., Chicago, IL, USA). Statistical significance was determined
through unpaired Student's t-tests or one-way analysis of variance
with post-hoc Tukey's test. P<0.05 was considered to indicate a
statistically significant difference.
Results
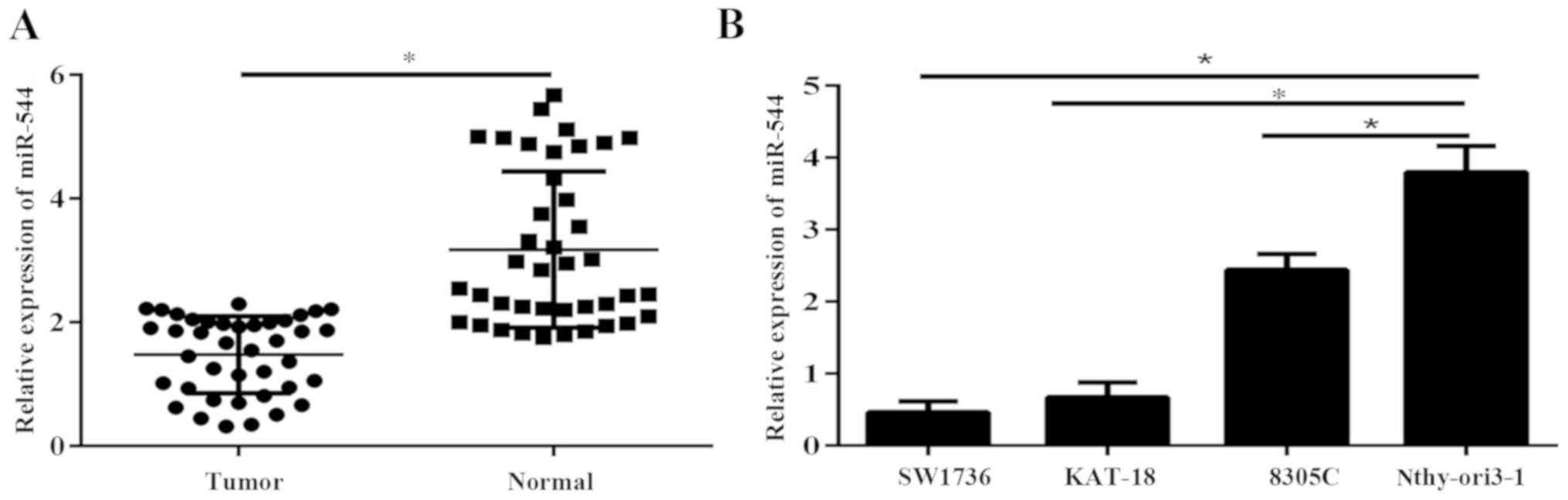
miR-544 is downregulated in ATC
tissues and cell lines
Expression of miR-544 was determined in 40 ATC
samples and adjacent normal thyroid tissues via RT-qPCR. The
results demonstrated that miR-544 was significantly downregulated
in ATC tissues, compared with normal thyroid tissues (P<0.05;
Fig. 1A). The expression level of
miR-544 was significantly reduced in ATC cell lines, compared with
the immortal thyroid cell line Nthy-ori3-1 (P<0.05; Fig. 1B).
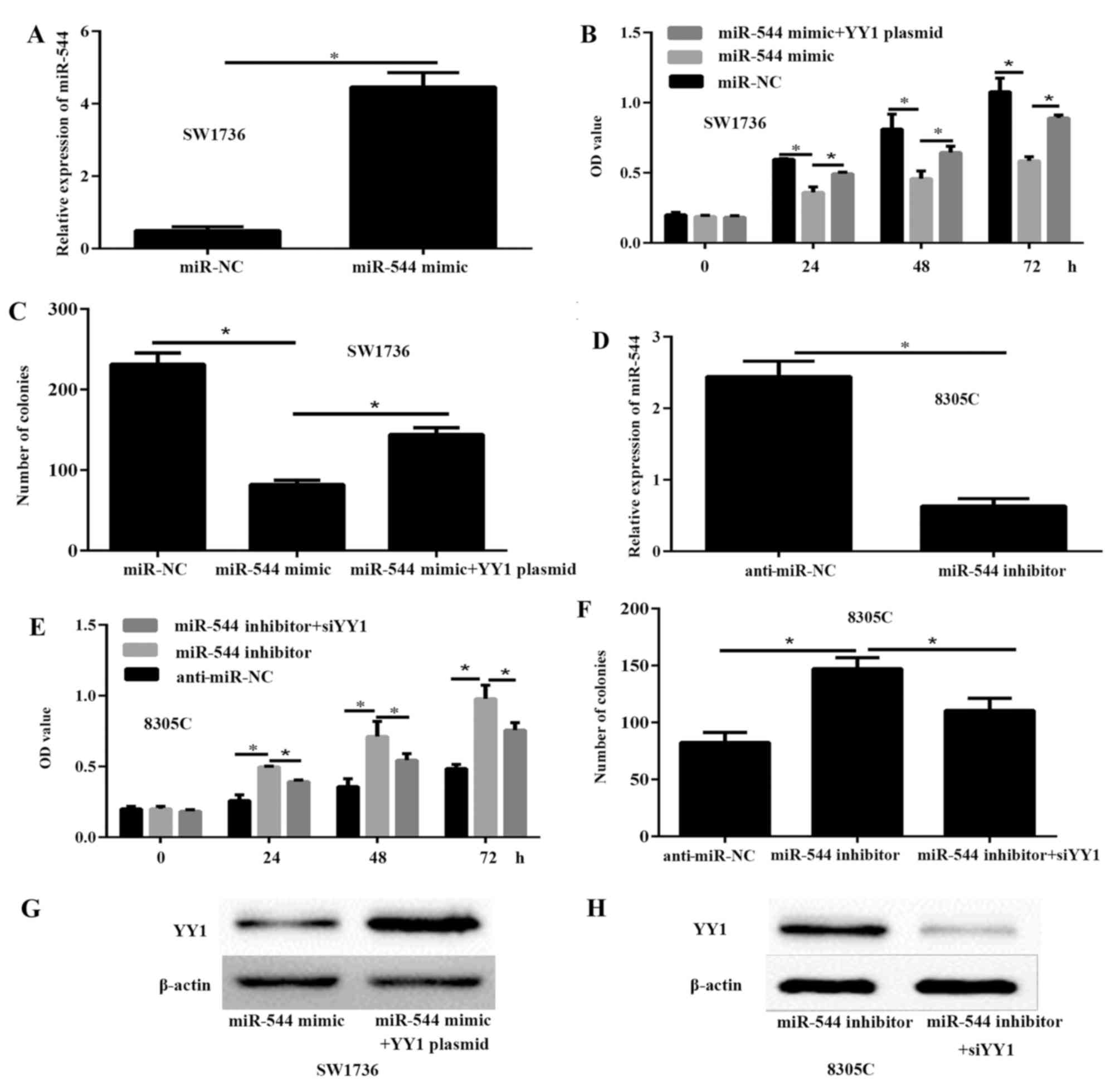
miR-544 inhibits the viability and
proliferation of ATC cells in vitro
To investigate the potential functions of miR-544 in
ATC, SW1736 cells were transfected with 50 nM miR-544 mimic or
miR-NC to significantly increase the expression of miR-544
(Fig. 2A). The results of the CCK-8
(Fig. 2B) and colony formation assays
(Fig. 2C) demonstrated that the
viability and proliferation of the SW1736 cells transfected with
miR-544 mimic were significantly decreased, compared with SW1736
cells transfected with miR-NC. To further investigate the effects
of miR-544 on ATC progression, 50 nM miR-544 inhibitor was
transfected into the 8305C cells. The RT-qPCR results demonstrated
that the expression of miR-544 was significantly decreased in the
8305C cells compared with the anti-miR-NC cells (P<0.05;
Fig. 2D). The CCK-8 (Fig. 2E) and colony formation assays
(Fig. 2F) indicated that miR-544
knockdown significantly enhanced the viability and proliferation of
8305C cells, compared with the anti-miR-NC cells (P<0.05). These
data demonstrated that miR-544 inhibited the viability and
proliferation of ATC cells. To further demonstrate the association
of miR-544 and YY1 in ATC, miR-544 mimic or miR-544 mimic and YY1
plasmid were transfected into SW1736 cells, and miR-544 inhibitor
or miR-544 inhibitor and siYY1 into 8305C cells. The results of the
western blot analysis indicated that YY1 was significantly
upregulated in SW1736 cells transfected with miR-544 mimic and YY1
plasmid compared to miR-544 mimic group (Fig. 2G). In addition, the expression of YY1
was decreased in 8305C cells transfected with miR-544 inhibitor and
siYY1 compared to miR-544 inhibitor (Fig.
2H).
miR-544 inhibits the migration and
invasion of ATC cells
To investigate the effect of miR-544 on the
metastatic ability of ATC cells, wound-healing, migration and
invasion assays were performed in SW1736 and 8305C cells. The
wound-healing assays demonstrated that the miR-544-overexpressing
cells had significantly decreased migration, compared with the
SW1736 cells transfected with miR-NC (P<0.05; Fig. 3A). By contrast, the miR-544-silenced
8305C cells had significantly enhanced migration capability,
compared with the 8305C cells transfected with anti-miR-NC
(P<0.05; Fig. 3B). Consistent with
the wound-healing assay, the Transwell and Matrigel assays also
demonstrated that miR-544 restoration significantly decreased the
migration and invasion capabilities of SW1736 cells (Fig. 3C and D), whereas miR-544 inhibition
significantly enhanced the migration and invasion capabilities of
8305C cells (P<0.05; Fig. 3E and
F).
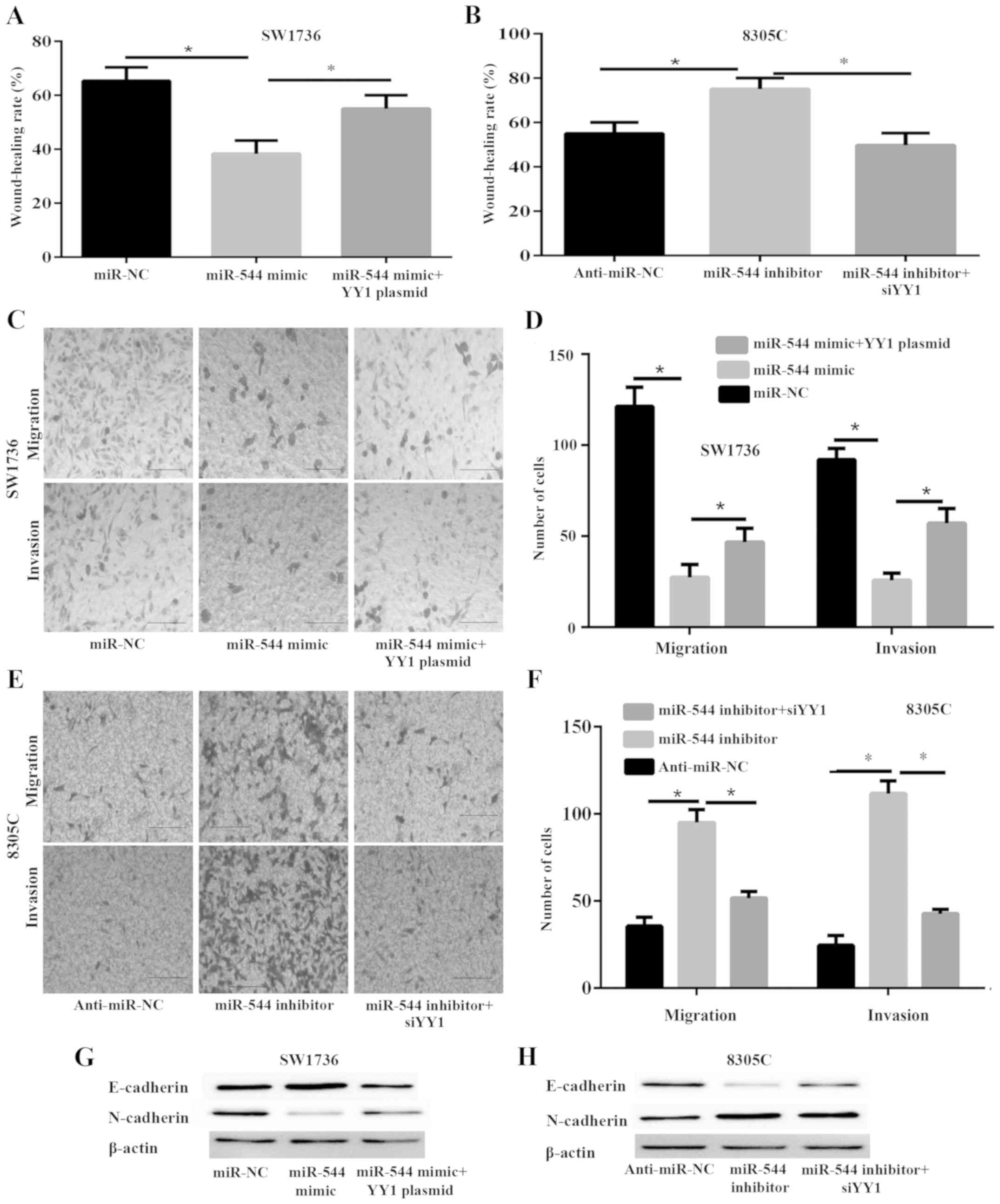 | Figure 3.miR-544 inhibits metastasis and
epithelial-mesenchymal transition of anaplastic thyroid cancer
cells via the expression of YY1. SW1736 cells were transfected with
miR-NC, miR-544 mimic or miR-544 mimic and YY1 plasmid, while 8305C
cells were transfected with anti-miR-NC, miR-544 inhibitor or
miR-544 inhibitor and si-YY1 for 48 h prior to analysis. (A)
wound-healing assay was used to analyze the migration capability of
SW1736 cells. (B) Wound-healing assay was used to analyze the
migration capability of 8305C cells. Transwell migration and
Matrigel invasion assays were used to detect the migration and
invasion capability, respectively. (C) Images (captured at ×100
magnification) of migration and invasion of SW1736 cells in the
lower chamber (scale bars, 50 µm). (D) Quantification of migration
and invasion of SW1736 cells in the lower chamber (scale bars, 50
µm). (E) ages (captured at ×100 magnification) of migration and
invasion of 8305C cells in the lower chamber (scale bars, 50 µm).
(F) Quantification of migration and invasion of 8305C cells in the
lower chamber (scale bars, 50 µm). (G) The protein expression of
E-cadherin and N-cadherin in SW1736 cells was detected with a
western blot analysis assay (H) The protein expression of
E-cadherin and N-cadherin in 8305C cells was detected with a
western blot analysis assay. *P<0.05. YY1, Yin Yang-1; miR,
microRNA; miR-NC, miR-negative control; anti-miR-NC, inhibitor
control. |
Epithelial-mesenchymal transition (EMT) is a
potential mechanism for tumor cell metastasis (13). To investigate whether miR-544 affects
EMT, the expression of the epithelial marker E-cadherin and
mesenchymal marker N-cadherin were measured in SW1736 and 8305C
cells by western blot analysis. The results demonstrated that
miR-544 mimic notably enhanced the expression of E-cadherin and
downregulated that of N-cadherin in SW1736 cells (Fig. 3G), whereas miR-544 inhibitor notably
decreased the expression of E-cadherin and upregulated that of
N-cadherin in 8305C cells (Fig.
3H).
YY1 is a direct target of miR-544
To determine the underlying molecular mechanisms by
which miR-544 suppresses ATC cancer growth, a search was conducted
to determine candidate targets of miR-544 that may serve a role in
ATC progression with and TargetScan 7.1. Among the candidates, YY1,
which is a pro-oncogene gene that is frequently upregulated in
various cancer types, was predicted to be a miR-544 target and
selected for further experimental verification. The predicted
interaction between miR-544 and the target site in the YY1 3′-UTR
is shown in Fig. 4A. Dual-luciferase
reporter assays were conducted to investigate whether miR-544
targets YY1 by binding to its 3′-UTR. SW1736 cells were
co-transfected with the Wt or Mut reporter vector and miR-544 mimic
or miR-NC. The results demonstrated that luciferase activities were
significantly decreased in the SW1736 cells transfected with the Wt
reporter vector and miR-544 mimic but not in the cells with the
mutant reporter vector compared with the transfected cells with the
miR-NC (P<0.05; Fig. 4B).
Additionally, inhibition of miR-544 significantly increased the
luciferase activity of the plasmid transfected with the WT 3′-UTR
of YY1 in 8305C cells compared with the transfected cells with the
anti-miR-NC (P<0.05; Fig. 4C).
Furthermore, the western blot assay demonstrated that miR-544
overexpression decreased the protein expression of YY1 in SW1736
cells compared with the transfected cells with the miR-NC (Fig. 4D), whereas the opposite is true in
8305C cells transfected with inhibitor compared with the
transfected cells with the anti-miR-NC (Fig. 4E). To further confirm the interaction
between miR-544 and YY1 3′-UTR, RNA immunoprecipitation assays were
performed. In the RNA extracted from the precipitated AGO2 protein,
it was possible to detect the miR-544 and YY1 3′UTR with notable
enrichment, compared with IgG in SW1736 and 8305C cells (Fig. 4F and G), indicating that miR-544 and
the YY1 3′-UTR existed in a RNA-induced silencing complex. These
data demonstrate that miR-544 directly targets YY1 by binding to
its 3′-UTR region in ATC cells.
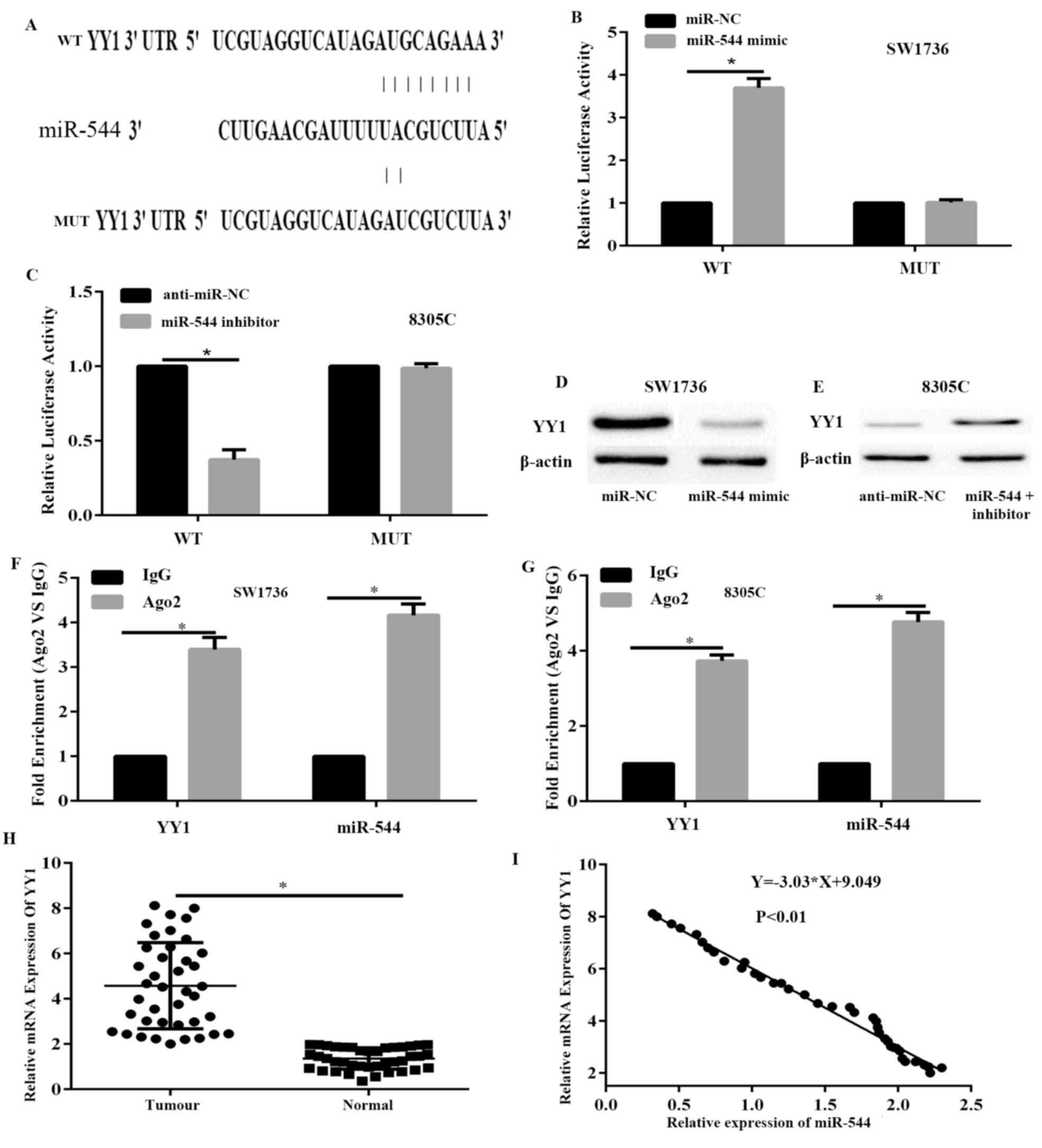 | Figure 4.YY1 is a direct target of miR-544. (A)
Sequence alignment of human miR-544 with 3′-UTR of YY1, as
predicted by TargetScan. A luciferase assay in (B) SW1736 and (C)
8305C cells co-transfected with indicated miRNA mimic or miRNA
inhibitor and luciferase reporter plasmids. Expression levels of
YY1 were detected in (D) SW1736 and (E) 8305C cells with a western
blot analysis assay. The association between miR-544/YY1 and AGO2
was analyzed. SW1736 and 8305C cellular lysates were used for RNA
immunoprecipitation with an AGO2 antibody. RT-qPCR was used to
detect the expression of miR-544 and YY1 in SW1736 cells (F). qPCR
was used to detect the expression of miR-544 and YY1 in 8305C cells
(G). miR-544 expression data was normalized to U6 small RNA
expression. All data of YY1 expression were normalized to β-actin
mRNA expression levels. (H) The mRNA expression levels of YY1 were
significantly increased in ATC tissues, compared with normal
tissues, as assessed with RT-qPCR. (I) Correlation between miR-544
expression levels and YY1 mRNA levels in ATC tissues was analyzed
using Spearman's correlation analysis. *P<0.05. YY1, Yin Yang-1;
miR, microRNA; miR-NC, miR-negative control; anti-miR-NC, inhibitor
control; Mut, mutated; Wt, wild-type; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; ATC,
anaplastic thyroid cancer. |
Subsequently, the mRNA expression levels of YY1 were
examined in ATC and adjacent normal tissues via RT-qPCR. The
results demonstrated that the expression level of YY1 was
significantly upregulated in ATC tissues, compared with adjacent
normal tissues (P<0.05; Fig. 4H).
Additionally, Spearman's correlation analysis indicated that the
expression levels of miR-544 were significantly inversely
correlated with YY1 mRNA in ATC tissues (P<0.05; Fig. 4I). Overall, these data demonstrate
that miR-544 directly targets YY1 by binding to its 3′-UTR region
in ATC cells.
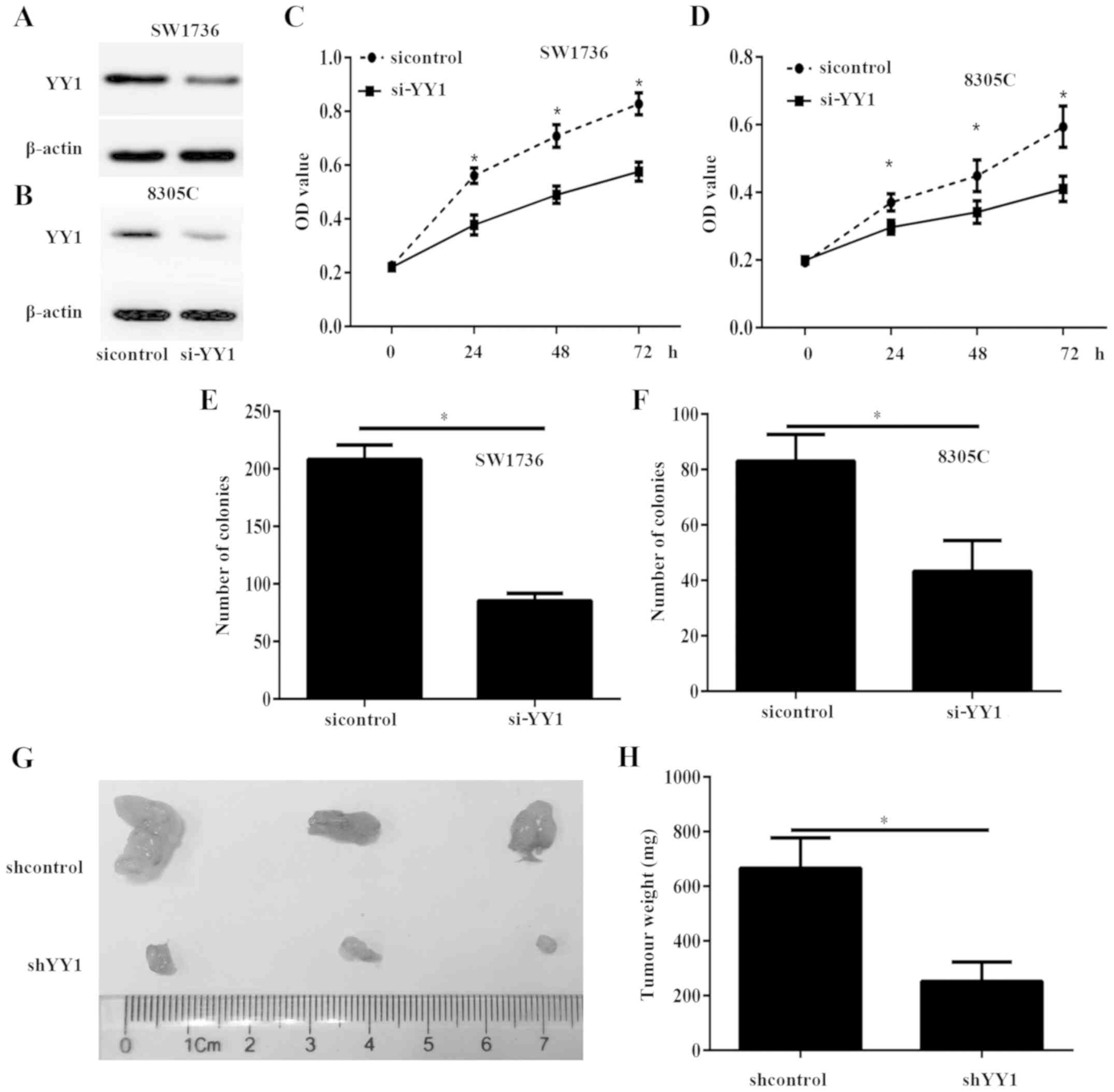
YY1 silencing inhibits ATC cell growth
in vitro and in vivo
To investigate the potential functions of YY1 in
ATC, SW1736 and 8305C cells with were transfected with si-YY1 or
sicontrol to decrease the expression of YY1 (Fig. 5A and B). The CCK-8 assay results
demonstrated that the viability of SW1736 and 8305C cells
transfected with si-YY1 was significantly decreased, compared with
the sicontrol cells (P<0.05; Fig. 5C
and D). The colony formation assay indicated that the colony
numbers in the SW1736 and 8305C cells transfected with si-YY1 were
significantly decreased, compared with those in the sicontrol cells
(P<0.05; Fig. 5E and F). To
confirm the aforementioned data in vivo, tumor xenograft
mouse models were produced. Nude mice were subcutaneously implanted
with SW1736 cells stably transfected with lentiviral shYY1 and
shControl. After 30 days, all mice were sacrificed and the tumor
xenograft was obtained (Fig. 5G). The
tumor weight significantly decreased in the shYY1 group, compared
with the shControl group (P<0.05; Fig.
5H).
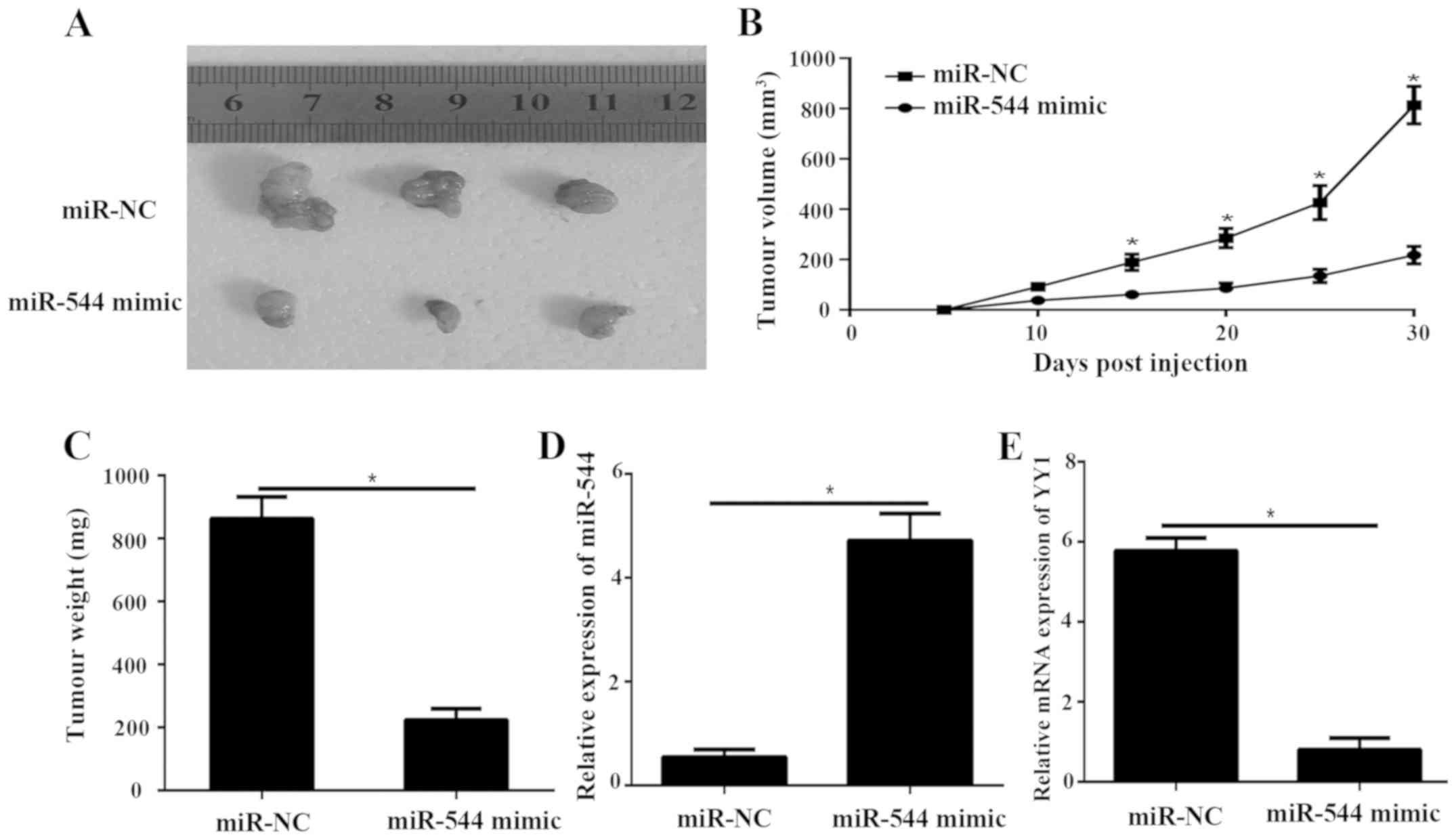
miR-544 suppresses tumor growth in
nude mice by downregulating YY1
To confirm the aforementioned data in vivo,
tumor xenograft mouse models were produced. Nude mice were
subcutaneously implanted with SW1736 cells transfected with
lentiviral miR-544 and miR-NC. After 30 days, all mice were
sacrificed, and the tumor xenograft was obtained (Fig. 6A). The tumor volume miR-544 in mimic
group was dramatically smaller compared with the miR-NC group
(P<0.05; Fig. 6B). The tumor
weight (Fig. 6C) were significantly
decreased in the miR-544 mimic group, compared with the miR-NC
group (P<0.05; Fig. 6C). The
results of RT-qPCR demonstrated that the expression level of
miR-544 in the miR-544 mimic tumor group was significantly
decreased, compared with the miR-NC group (P<0.05; Fig. 6C and D). The mRNA level of YY1 was
significantly decreased in the tumor-transfected miR-544 mimic
compared to miR-NC group (P<0.05; Fig.
6E).
miR-544 promoted ATC progression via
YY1
Finally, whether miR-544 promotes ATC progression
via YY1 was investigated. The protein expression level of YY1 was
detected by western blot assay (Fig. 2G
and H). The results of the CCK-8 (P<0.05; Fig. 2B), colony formation assay (Fig. 2C; P<0.05), and Transwell migration
and Matrigel invasion assays (P<0.05; Fig. 3C and D) demonstrated that YY1
overexpression significantly reversed the inhibitory effects of
miR-544 on the viability, proliferation and metastasis of SW1736
cells compared with the miR-544 mimic group (P<0.05), whereas
YY1 silencing partially reversed the promotion effects of the
miR-544 inhibitor of the viability (P<0.05; Fig. 2E), proliferation (P<0.05; Fig. 2F), and metastasis (P<0.05; Fig. 3E and F) of 8305C cells compared with
the miR-544 inhibitor group (P<0.05). Furthermore, western blot
analysis indicated that YY1 partially reverses the inhibitory
effect of EMT induced by miR-544 mimic in SW1736 cells (Fig. 3G); however, YY1 silencing can
partially reverse the inhibitory effect of EMT induced by miR-544
inhibitor in 8305C cells (Fig. 3H).
These data demonstrate that miR-544 inhibits tumor progression via
YY1 in ATC.
Discussion
Previous studies demonstrated that the abnormal
expression of miR-544 is associated with glioma, osteosarcoma,
cervical cancer, breast cancer and gastric cancer (6–10).
However, the expression and function of miR-544 in ATC tissues
remains unclear. In the present study, it was indicated that
miR-544 is downregulated in ATC and that it may act as a novel
tumor suppressor in ATC. Furthermore, upregulation of miR-544
significantly suppressed tumor growth, proliferation and
metastasis, while the miR-544 inhibitor enhanced the tumor
progression. Additionally, it was demonstrated that miR-544 acted
as a tumor suppresser via directly targeting YY1.
YY1 is a conserved multifunctional protein belonging
to the GLI-Kruppel family, and it binds to the promoter of other
genes primarily through the C-terminal zinc finger region (14,15). As a
transcriptional regulator, YY1 is involved in multiple biological
functions of cells, including embryonic formation, cell
proliferation, apoptosis, DNA repair and differentiation via
upregulating or downregulating different genes (16). It has been reported that YY1 is
overexpressed and serves as oncogene in numerous tumor types,
including breast, ovary, colon, prostate, gastric and laryngeal
cancer (17–21). However, YY1 may serve an inhibitory
role in pancreatic cancer (22). A
recent study demonstrated that the expression of YY1 was
significantly upregulated in differentiated thyroid and anaplastic
cancer (23). However, the function
of YY1 in ATC has not been well illustrated. In the present study,
consistent with previous study, it was also determined that YY1 has
a high expression level in ATC tissues. Additionally, YY1 silencing
suppressed cell viability and proliferation, and significantly
decreased the migration and invasion, of ATC cells. As a key
transcription factor, YY1 regulates its target gene transcription
in a complex manner. YY1 can activate the transcription of
oncogenes and tumor suppressors in different cancer tissues
(14–23). However, the mechanism regulating YY1
has not been well illustrated. In the present study, YY1 was
identified as a direct target of miR-544 by dual-luciferase
reporter assays. Furthermore, Transwell migration and invasion
assay indicated that the most important effect exerted by miR-544
on ATC cells invasion and migration, which was partially reversed
when co-transfected with the YY1 plasmid. These results
demonstrated that YY1 was a functional target gene of miR-544 in
ATC. It is notable that each miRNA can regulate dozens of genes and
multiple miRNAs may regulate the identical gene, which affect the
activities of whole pathways. Therefore, the possibility that other
target genes may also be involved in the suppressive effects of
miR-544 cannot be excluded.
In conclusion, it was determined that miR-544 was
significantly downregulated in ATC. Furthermore, it was
demonstrated, for the first time, that the role of the miR-544/YY1
axis is to regulate ATC proliferation and metastasis. This novel
miR-544/YY1 association provides a new insight into the mechanisms
underlying ATC development, and indicates that targeting the
miR-544/YY1 axis may represent a promising therapeutic strategy for
ATC treatment.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
FW and BS contributed to study design, statistical
analysis, data interpretation, manuscript preparation and the
literature search. ZL performed the experiments and data
collection. All authors have read and approved the final version of
the manuscript.
Ethics approval and consent to
participate
This study was approved by the Ethics Committee of
Yantai Laiyang Central Hospital (Yantai, China). Written informed
consent was obtained from every patient. All animal studies were
approved by the Animal Care and Welfare Committee of Yantai Laiyang
Central Hospital.
Patient consent for publications
All patients provided written informed consent prior
to participation in the present study and consent for the
publication of the present study.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
O'Neill JP and Shaha AR: Anaplastic
thyroid cancer. Oral Oncol. 49:702–706. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bu Q, You F, Pan G, Yuan Q, Cui T, Hao L
and Zhang J: MiR-125b inhibits anaplastic thyroid cancer cell
migration and invasion by targeting PIK3CD. Biomed Pharmacother.
88:443–448. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shao M, Geng Y, Lu P, Xi Y, Wei S, Wang L,
Fan Q and Ma W: miR-4295 promotes cell proliferation and invasion
in anaplastic thyroid carcinoma via CDKN1A. Biochem Biophys Res
Commun. 464:1309–1313. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhu Z, Wang S, Zhu J, Yang Q, Dong H and
Huang J: MicroRNA-544 down-regulates both Bcl6 and Stat3 to inhibit
tumor growth of human triple negative breast cancer. Biol Chem.
397:1087–1095. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jin S, Dai Y, Li C, Fang X, Han H and Wang
D: MicroRNA-544 inhibits glioma proliferation, invasion and
migration but induces cell apoptosis by targeting PARK7. Am J
Transl Res. 8:1826–1837. 2016.PubMed/NCBI
|
|
8
|
Mao L, Zhang Y, Deng X, Mo W, Yu Y and Lu
H: Transcription factor KLF4 regulates microRNA-544 that targets
YWHAZ in cervical cancer. Am J Cancer Res. 5:1939–1953.
2015.PubMed/NCBI
|
|
9
|
Ma R, Zhang G, Wang H, Lv H, Fang F and
Kang X: Downregulation of miR-544 in tissue, but not in serum, is a
novel biomarker of malignant transformation in glioma. Oncol Lett.
4:1321–1324. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhi Q, Guo X, Guo L, Zhang R, Jiang J, Ji
J, Zhang J, Zhang J, Chen X, Cai Q, et al: Oncogenic miR-544 is an
important molecular target in gastric cancer. Anticancer Agents Med
Chem. 13:270–275. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
Elife. (4)2015.
|
|
13
|
Brabletz T, Kalluri R, Nieto MA and
Weinberg RA: EMT in cancer. Nat Rev Cancer. 18:128–134. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shi Y, Seto E, Chang LS and Shenk T:
Transcriptional repression by YY1, a human GLI-Kruppel-related
protein and relief of repression by adenovirus E1A protein. Cell.
67:377–388. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gordon S, Akopyan G, Garban H and Bonavida
B: Transcription factor YY1: Structure, function, and therapeutic
implications in cancer biology. Oncogene. 25:1125–1142. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bonavida B and Kaufhold S: Prognostic
significance of YY1 protein expression and mRNA levels by
bioinformatics analysis in human cancers: A therapeutic target.
Pharmacol Ther. 150:149–168. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Seligson D, Horvath S, Huerta-Yepez S,
Hanna S, Garban H, Roberts A, Shi T, Liu X, Chia D, Goodglick L and
Bonavida B: Expression of transcription factor Yin Yang 1 in
prostate cancer. Int J Oncol. 27:131–141. 2005.PubMed/NCBI
|
|
18
|
Kang W, Tong JH, Chan AW, Zhao J, Dong Y,
Wang S, Yang W, Sin FM, Ng SS, Yu J, et al: Yin Yang 1 contributes
to gastric carcinogenesis and its nuclear expression correlates
with shorter survival in patients with early stage gastric
adenocarcinoma. J Transl Med. 12:802014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Allouche A, Nolens G, Tancredi A,
Delacroix L, Mardaga J, Fridman V, Winkler R, Boniver J, Delvenne P
and Begon DY: The combined immunodetection of AP-2alpha and YY1
transcription factors is associated with ERBB2 gene overexpression
in primary breast tumors. Breast Cancer Res. 10:R92008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chinnappan D, Xiao D, Ratnasari A, Andry
C, King TC and Weber HC: Transcription factor YY1 expression in
human gastrointestinal cancer cells. Int J Oncol. 34:1417–1423.
2009.PubMed/NCBI
|
|
21
|
Qu SY, Sun YY, Li YH, Xu ZM and Fu WN: YY1
directly suppresses MYCT1 leading to laryngeal tumorigenesis and
progress. Cancer Med. 6:1389–1398. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu D, Zhang J, Wu Y, Shi G, Yuan H, Lu Z,
Zhu Q, Wu P, Lu C, Guo F, et al: YY1 suppresses proliferation and
migration of pancreatic ductal adenocarcinoma by regulating the
CDKN3/MdM2/P53/P21 signaling pathway. Int J Cancer. 142:1392–1404.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Arribas J, Castellví J, Marcos R, Zafón C
and Velázquez A: Expression of YY1 in differentiated thyroid
cancer. Endocr Pathol. 26:111–118. 2015. View Article : Google Scholar : PubMed/NCBI
|