Introduction
Lung cancer is the leading cause of
cancer-associated mortality worldwide, with 80–90% of all lung
cancer patients diagnosed with non-small cell lung cancer (NSCLC)
(1–3).
Great progress has been made in understanding the carcinogenesis of
NSCLC; however, owing to the heterogeneity and genetic instability
of NSCLC cells, therapeutic methods that are being used in the
clinic, including chemotherapy, radiation therapy or targeted
therapy such as epithelial growth factor receptor (EGFR)-tyrosine
kinase inhibitors, often exhibit short-lived treatment response
(4). Novel therapeutic targets or
methods are therefore required.
A previous study revealed that high mobility group
box 3 (HMGB3) overexpression is an independent risk factor for
NSCLC progression, lymph node metastasis and poor survival rates of
patients (5), indicating the
prognostic value and the oncogenic role of HMGB3 overexpression in
NSCLC development. HMGB3 is a less studied member in the
evolutionarily conserved HMGB protein family, alongside HMGB1 and
HMGB2 (6,7). Initially reported to be a modulator
involved in tissue development (8–10) and
hematopoietic stem cell differentiation (11–13), HMGB3
has also been linked to cancer development by certain studies
(14–19); however, its influence on in the cell
biological behaviors of NSCLC cells remains unrevealed.
The present study investigated the role of HMGB3 on
the cell viability and colony forming ability of NSCLC cells by
constructing an A549 cell line that was stably knocked down for
HMGB3 expression using a lentiviral short hairpin RNA (shRNA)
interference technique. The data generated by the present study
clearly demonstrated that HMGB3 is necessary for A549 cell survival
and colony formation, indicating its role in NSCLC progression and
its potential as a novel therapeutic target.
Materials and methods
Cell culture
The human NSCLC A549 cell line was a gift from
Tianjin Medical University Cancer Institute and Hospital (Tianjin,
China). The human NSCLC H1299 cell line was purchased from Chinese
Academy of Sciences Cell Bank (Shanghai, China). A549 and H1299
cells were cultured in RPMI-1640 medium supplemented with 10% fetal
bovine serum, and 100 U/ml of penicillin and streptomycin (all from
Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), in a
humidified incubator at 37°C with 5% CO2. Cells in the
logarithmic growth phase were subject to each experiment at a
density of 0.5–1×106/ml.
HMGB3 knockdown in NSCLC cells by
shRNA transfection
Lentiviral-based shRNA vectors (containing puromycin
and enhanced green fluorescent protein) were constructed by
Genecopoeia, Inc. (Rockville, MD, USA). Specifically, three shRNAs
with different target sequence in the HMGB3 coding DNA sequence and
one non-targeting shRNA as negative control were involved in this
experiment (referred to as shHMGB3-1, −2 and −3, and shNC,
respectively). The stable knockdown of HMGB3 in NSCLC cells (A549
and H1299 cells) was performed by shRNA transfection using an
EndoFectin transfection (Genecopoeia, Inc.) following
manufacturer's protocol. Transfection with HMGB3 (250 µg/ml) was
conducted for 24 h by EndoFectin transfection. Subsequent
experiments were performed after transfection 36 h. Additionally,
positive transfection was determined by detecting the green
fluorescence under fluorescent microscope. The efficiency of HMGB3
knockdown in each cell line transfected with different shRNAs was
evaluated by measuring the mRNA and protein expression level of
HMGB3 by reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) and western blotting, respectively.
Cell viability, apoptosis and colony
formation assay
The viability (at 1, 12 and 24 h) of NSCLC cells
with or without HMGB3 knockdown was compared using Cell Counting
Kit-8 (Beyotime Institute of Biotechnology, Shanghai, China),
following the manufacturer's protocol.
Apoptosis of NSCLC cells with or without HMGB3
knockdown was compared by flow cytometry using a Attune NxT flow
cytometer (Thermo Fisher Scientific, Inc.) and FlowJo software
(version 7.6; FlowJo LLC, Ashland, OR, USA) following staining with
the Annexin V-EGFP Apoptosis Detection kit (Beyotime Institute of
Biotechnology) following the manufacturer's protocol. NSCLC cells
with or without HMGB3 knockdown were cultured to logarithmic phase,
followed by 0.25% trypsin treatment and pipetting to make single
cell suspension in complete culture medium.
Approximately 200 cells in each experimental group
were seeded in 10 ml RPMI-1640 in a cell culture dish and
maintained in a humidified incubator at 37°C with 5% CO2
for 2–3 weeks until macroscopic colonies formed. Following removal
of the culture medium, cells in each culture dish were fixed with
4% paraformaldehyde solution (room temperature) for 15 min followed
by incubation with 1% crystal violet staining solution (20 min at
room temperature). The number of colonies of >50 cells were
counted under low-magnification light microscope (×100), and the
colony forming efficiency of each experimental group was calculated
with the following formula: Clonal formation rate=number of clones
formed/number of inoculated cells ×100%.
RT-qPCR
Total RNA was isolated by TRIzol®,
chloroform and isopropanol (Takara Bio, Inc., Otsu, Japan)
extraction. Human HMGB3 and GAPDH mRNA expression in cells of each
experimental group was evaluated by RT-qPCR using a custom-made
RT-qPCR kit (cat. no. QP001) purchased from Genecopoeia, Inc.,
following the manufacturer's instructions. Briefly, poly-A RNAs in
the cell lysate in each experimental group were first reversely
transcribed to cDNAs, and HMGB3 and GAPDH cDNAs were subject to
amplification by qPCR monitored by SYBRGreen fluorescent dye. GAPDH
was used as reference gene for HMGB3 quantification. HMGB3
expression levels were normalized to levels of GAPDH using the
2−ΔΔCq method (20).
Primers for HMGB3 and GAPDH cDNA were as follows: HMGB3 forward,
5′-GACCAGCTAAGGGAGGCAA-3′ and reverse 5′-ACAGGAAGAATCCAGACGGT-3′;
and GAPDH forward, 5′-CCTGCACCACCAACTGCTTA-3′ and reverse,
5′-GGCCATCCACAGTCTTCTGAG-3′. Thermocycling conditions for
amplifying HMGB3 and GAPDH cDNAs were as follows: Initial
denaturation at 95°C for 10 min, followed by 35 cycles of
denaturation 95°C for 10 sec, 55°C for 20 sec and 72°C for 15
sec.
Western blot analysis
Anti-HMG4 (cat. no. ab201569), anti-β-actin (cat.
no. ab6276) and horseradish peroxidase (HRP)-conjugated anti-mouse
IgG (cat. no. ab6728) were purchased from Abcam (Cambridge, UK).
Antibodies against Bax (cat. no. D2E11), Bcl-2 (cat. no. D55G8),
cleaved caspase-9 (cat. no. D2D4), cleaved caspase-8 (cat. no.
11G10) and anti-biotin (D5A7) rabbit mAb (HRP conjugate; cat. no.
5571S) were purchased from Cell Signaling Technology, Inc.
(Danvers, MA, USA). Antibodies were applied in western blotting
analysis following manufacturer's instructions with minor
modifications. Cells were homogenized in lysis buffer [10 mM Tris
base, pH 7.4; 150 mM NaCl; 1 mM EDTA; 1 mM EGTA; 1% Triton X-100;
0.5% NP-40; and protease and phosphatase inhibitor cocktail (Thermo
Fisher Scientific, Inc.)]. Protein concentrations were determined
using a BCA assay kit (Thermo Fisher Scientific, Inc.) with bovine
serum albumin (Gibco; Thermo Fisher Scientific, Inc.) as a
standard. Briefly, following separation of proteins (20 µl/cell
group) in cell lysates by SDS-PAGE (15%) and transferred onto
nitrocellulose membranes. Following blocking at 4°C overnight with
non-fat milk and preparation with PBS, the membranes were incubated
for 1 h at room temperature with specific primary antibodies
diluted in bovine serum albumin-dissolved Tris-buffered saline
diluent with final dilution concentration of 1:1,000; secondary
antibodies were diluted in the same diluent at 1:10,000 ratio to
probe the specific primary antibodies by incubation for 30 min at
room temperature and colorized using an enhanced chemiluminescent
substrate (Thermo Fisher Scientific, Inc.). Quantified grayscale
analysis of western blotting result was performed using ImageJ
software (version 1.48, National Institutes of Health, Bethesda,
MD, USA); β-actin was used as loading control for each
analysis.
Statistical analysis
According to the recommended protocol by Szklarczyk
et al (21), the proteins
interacting with HMGB3 were investigated using the STRING online
database (search term, HMGB3; https://string-db.org/cgi/input.pl; version 10.5),
with the confidence score set at 0.4. HMGB3-associated protein
analysis was conducted through Gene ontology (GO) enrichment
analysis (http://www.geneontology.org/). Statistical analysis
was performed using SPSS 17 software (SPSS, Inc., Chicago, IL,
USA). Quantitative data were presented as mean ± standard deviation
and analyzed by analysis of one-way analysis o variance. Comparison
between experimental groups and control group in each graph was
performed with Dunnett's multiple comparison test. P<0.05 was
considered to indicate a statistically significant difference.
Results
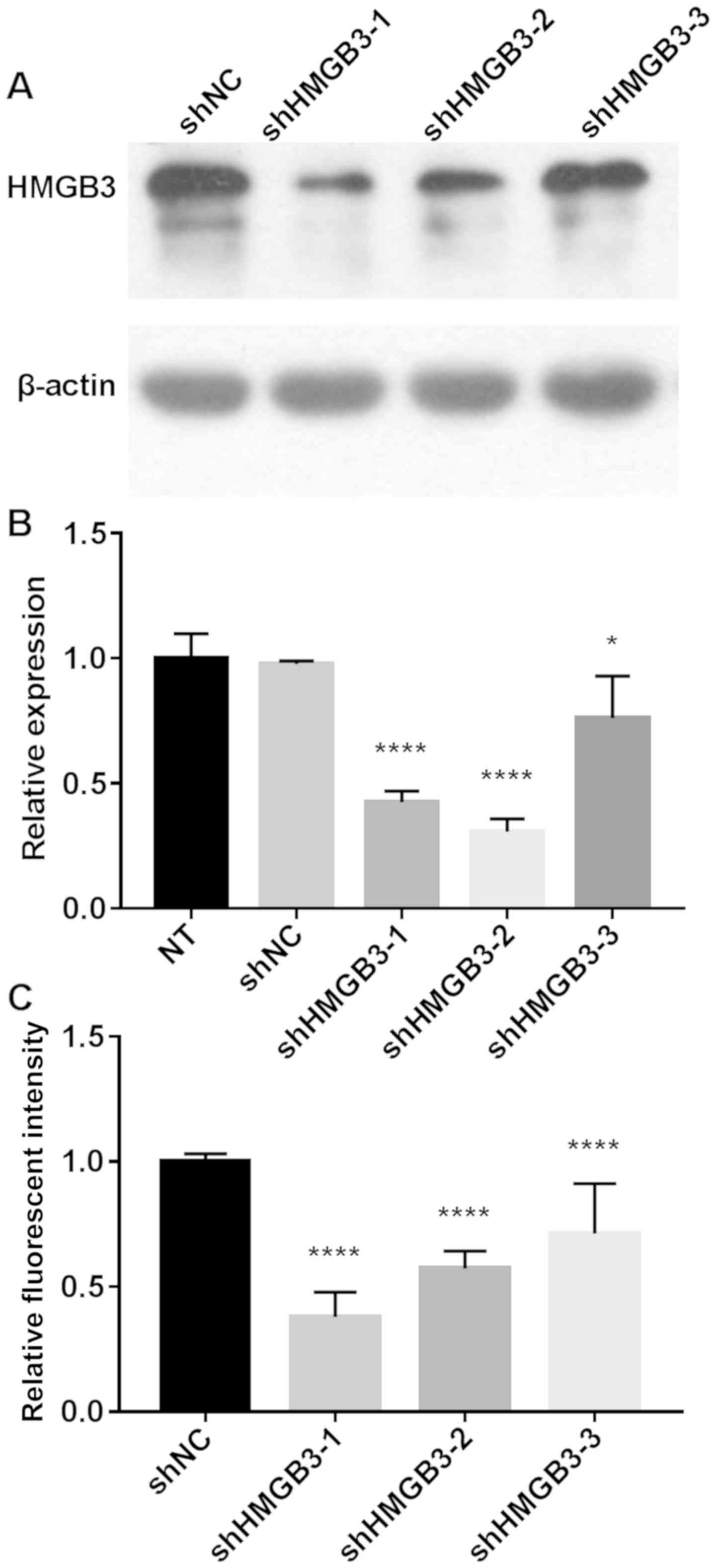
HMGB3 knockdown in NSCLC cells using
shRNA
To investigate the role of HMGB3 on the tumor
biology of human NSCLC A549 and H1299 cell lines with stable
knockdown of HMGB3 were constructed using different shRNAs
(Fig. 1). The data revealed that A549
cells transfected with shHMGB3-1 and shHMGB3-2 exhibited a >50%
decrease in HMGB3 expression at the transcriptional and
translational level (P<0.0001), compared with non-transfected
control cells (NT) or cells transfected with non-targeting shRNA
control (shNC). A549 cells transfected with shHMGB3-3 exhibited a
moderate decrease in HMGB3 expression, whereas those transfected
with shHMGB3-1 exhibited the lowest HMGB3 protein expression. It
was therefore decided to use only the A549 cell line with HMGB3
knockdown achieved using shHMGB3-1 (referred to as shHMGB3
hereafter) for further experimentation. A similar degree of HMGB3
knockdown using shHMGB3-1 was also achieved in H1299 NSCLC cells
(data not shown).
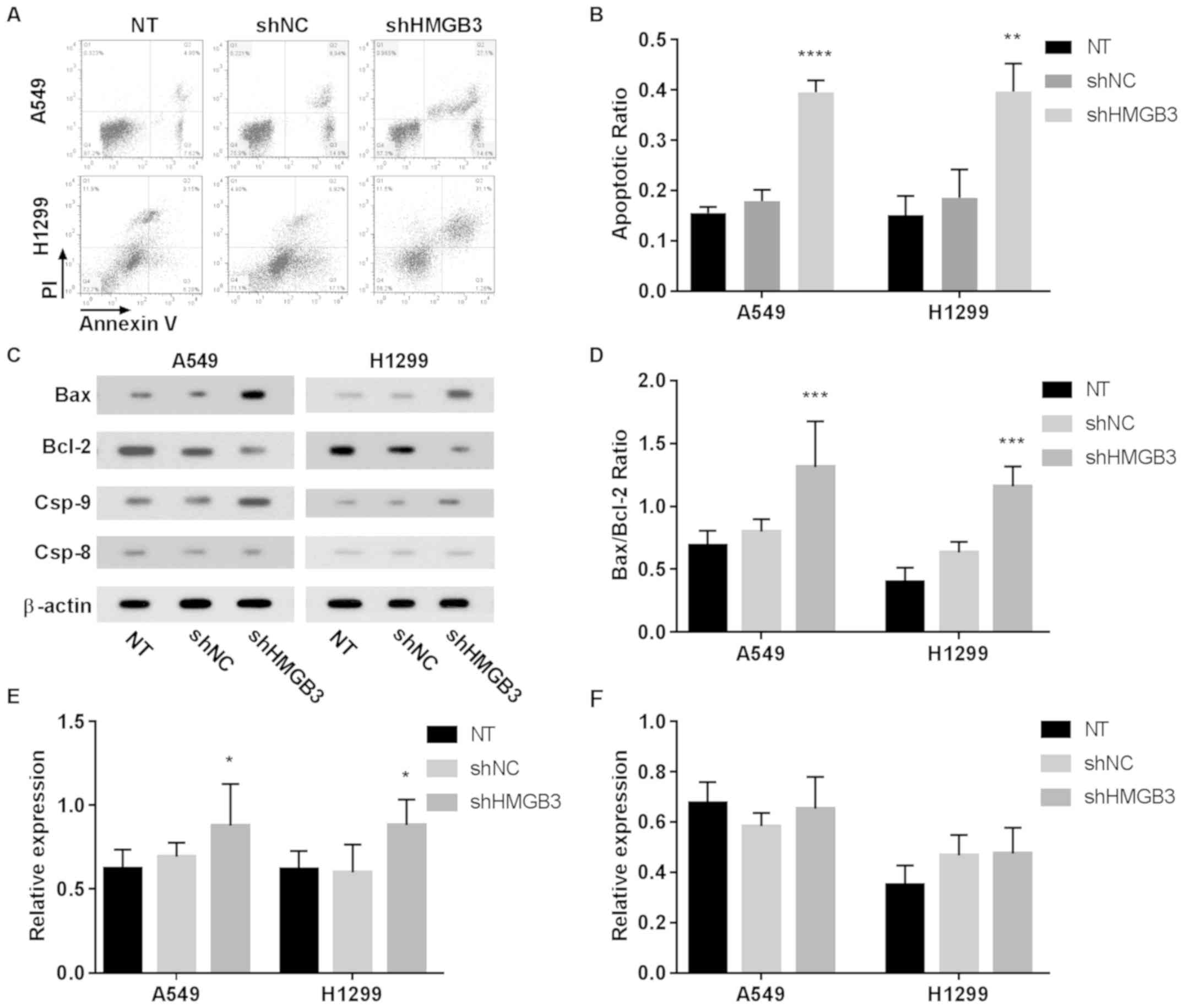
Decrease in cell viability and
increase in apoptosis of NSCLC cells following HMGB3 knockdown
The role of HMGB3 in the self-maintenance of NSCLC
cells was assessed by comparing the viability and apoptosis of A549
or H1299 cells with or without HMGB3 knockdown. The flow cytometry
data revealed a >2-fold increase in cell apoptosis following
HMGB3 knockdown in A549 (P<0.0001) or H1299 (P<0.01) cells
(Fig. 2A and B. These results
indicated that HMGB3 may be vital for the in vitro survival
of A549 or H1299 cells. To investigate the apoptotic inducing
mechanism of HMGB3 knockdown in A549 or H1299 cells, the expression
levels or activation status of certain apoptosis-associated
proteins, namely Bax, Bcl-2, caspase-9 and caspase-8, under
different HMGB3 expression conditions by western blotting (Fig. 2C-F). These data clearly indicated that
HMGB3 knockdown significantly increased the Bax/Bcl-2 ratio by
nearly 2-fold in A549 or H1299 cells (P<0.001), which has been
suggested as a key step during the initiation of apoptosis; HMGB3
knockdown also increased caspase-9 activation in A549 and H1299
cells by ~40% (P<0.05) compared with NT, as evidenced by the
increased caspase-9 cleavage, but not that of caspase-8.
Considering that caspase-9 and caspase-8 belong to the intrinsic
and extrinsic apoptosis pathway, respectively, these data indicated
that HMGB3 knockdown primarily activates the intrinsic apoptosis
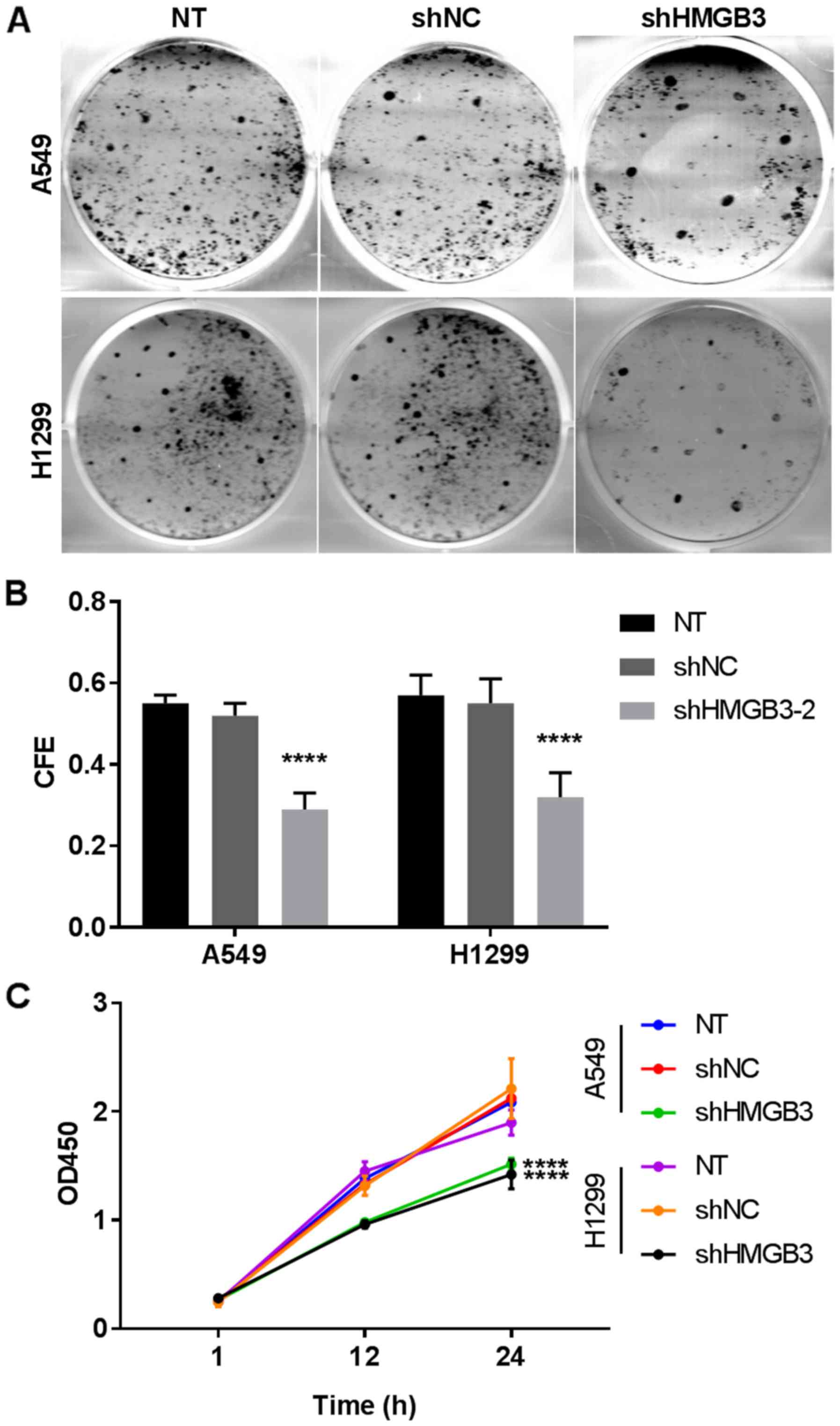
pathway in NSCLC cells in vitro. To investigate the
influence of HMGB3 on the colony formation capacity of NSCLC cells,
the colony-forming efficiency of A549 or H1299 cells with or
without HMGB3 knockdown was assessed (Fig. 3A and B). The data revealed that the
colony forming efficiency of A549 or H1299 cells decreased by
>40% after HMGB3 knockdown (P<0.0001), indicating that it
serves a notable role in the colony formation of NSCLC cells. The
results of the CCK-8 assay exhibited a decrease in cell viability
of almost 50% (P<0.0001) in the two HMGB3-knockdown cell lines,
compared with the NT (Fig. 3C). Cell
viability of A549 or H1299 was decreased following HMGB3
knockdown.
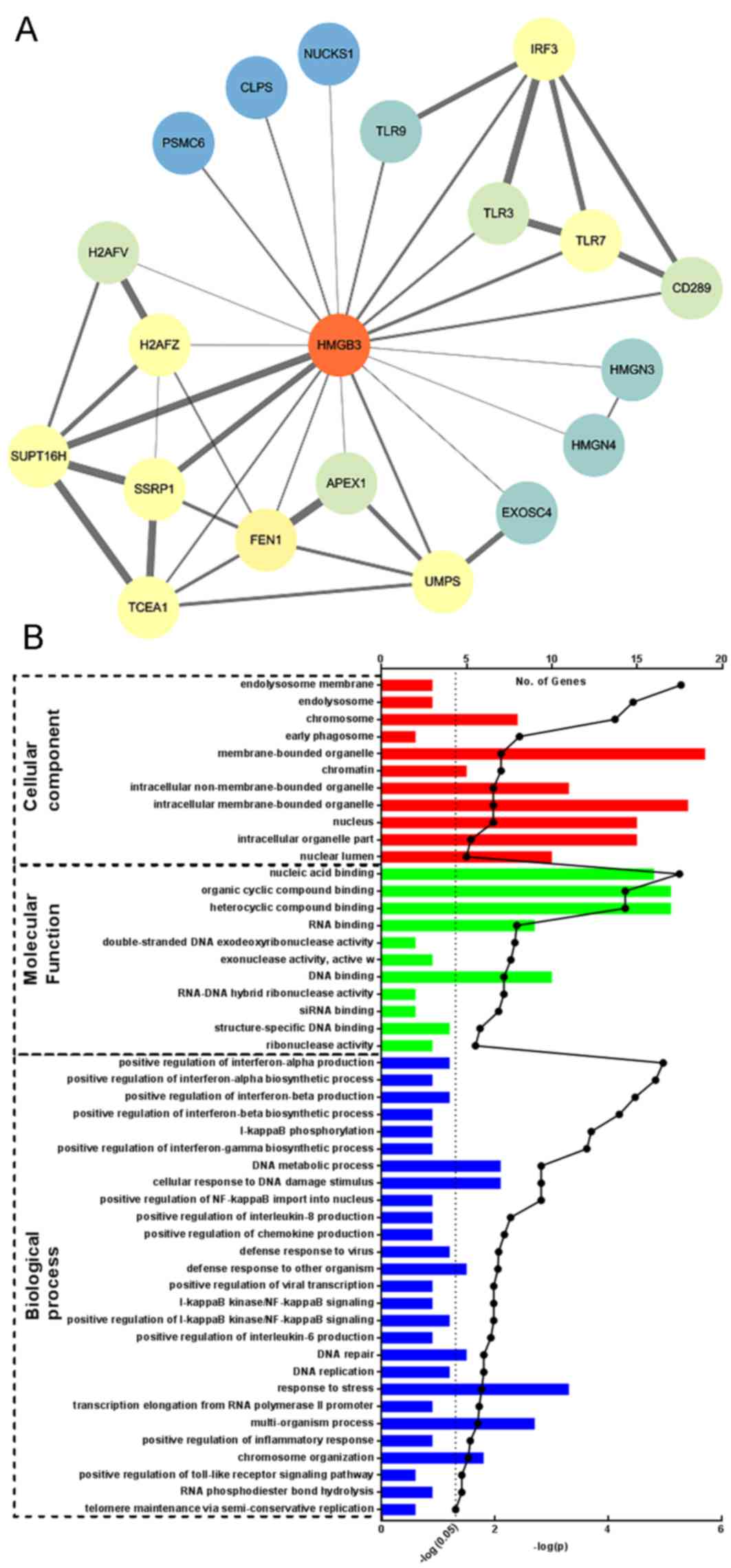
HMGB3 facilitates NSCLC survival in
vitro possibly by interacting with FACT complex protein SSRP1 and
SUPT16H
To investigate the possible mechanism underlying the
supporting effect of HMGB3 on NSCLC survival, the proteins that
have been suggested to interact with HMGB3 were investigated using
the STRING online database. There were 20 proteins retrieved with a
confidence score >0.4 who have either been experimentally
confirmed or presumed by text mining to interact with HMGB3
(Fig. 4A). GO enrichment analysis
revealed that the majority of these proteins were involved in DNA
replication, repair and regulation of mRNA translation, although a
small portion of these proteins belonged to the Toll-like receptor
signaling pathways (Fig. 4B; Table I). Notably, among these proteins FACT
complex subunit SPT16 (SUPT16H) and structure specific recognition
protein 1 (SSRP1) exhibited the highest confidence score. These two
proteins are the two subunits of facilitates chromatin
transcription (FACT) complex, which has been confirmed to be
frequently upregulated in different types of malignant cancer.
These findings indicate that HMGB3 may facilitate NSCLC survival by
interacting with FACT complex proteins SUPT16H and SSRP1 or other
proteins involved in the regulation of DNA metabolism.
 | Table I.Gene Ontology enrichment analysis of
predicted HMGB3 interacting proteins. |
Table I.
Gene Ontology enrichment analysis of
predicted HMGB3 interacting proteins.
| Category | Description | Enriched genes | FDR P-value |
|---|
| Cellular
component | Endolysosome
membrane | TLR3, TLR9, TLR7 |
5.23×10−6 |
|
| Endolysosome | TLR3, TLR9, TLR7 |
3.65×10−5 |
|
| Chromosome | SUPT16H, SSRP1,
H2AFZ, H2AFV, EXOSC4, HMGN4 |
7.63×10−5 |
|
|
| HMGN3, HMGB3 |
|
|
| Early phagosome | TLR9, TLR7 | 0.00371 |
|
| Membrane-bounded
organelle | SUPT16H, APEX1, UMPS,
CLPS, SSRP1, H2AFZ, TLR3, FEN1, H2AFV, IRF3, EXOSC4, HMGN4, HMGN3,
TLR9, NUCKS1, HMGB3, TLR7, PSMC6, TCEA1 | 0.00785 |
|
| Chromatin | H2AFZ, H2AFV,
EXOSC4, HMGN4, HMGN3 | 0.00785 |
|
| Intracellular
non-membrane-bounded organelle | SUPT16H, APEX1,
SSRP1, H2AFZ, FEN1, H2AFV, EXOSC4, HMGN4, HMGN3, HMGB3, TCEA1 | 0.0108 |
|
| Intracellular
membrane-bounded organelle | SUPT16H, APEX1,
UMPS, SSRP1, H2AFZ, TLR3, FEN1, H2AFV, IRF3, EXOSC4, HMGN4, HMGN3,
TLR9, NUCKS1, HMGB3, TLR7, PSMC6, TCEA1 | 0.0108 |
|
| Nucleus | SUPT16H, APEX1,
UMPS, SSRP1, H2AFZ, FEN1, H2AFV, IRF3, EXOSC4, HMGN4, HMGN3,
NUCKS1, HMGB3, PSMC6, TCEA1 | 0.0108 |
|
| Intracellular
organelle part | SUPT16H, APEX1,
SSRP1, H2AFZ, TLR3, FEN1, H2AFV, IRF3, EXOSC4, HMGN4, HMGN3, TLR9,
TLR7, PSMC6, TCEA1 | 0.0268 |
|
| Nuclear lumen | SUPT16H, APEX1,
SSRP1, H2AFZ, FEN1, IRF3, EXOSC4, HMGN3, PSMC6, TCEA1 | 0.0321 |
| Molecular | Nucleic acid
binding function | SUPT16H, APEX1,
SSRP1, H2AFZ, TLR3, FEN1, H2AFV, IRF3, EXOSC4, HMGN4, HMGN3, TLR9,
NUCKS1, HMGB3, TLR7, TCEA1, |
5.57×10−6 |
|
| Organic cyclic
compound binding | SUPT16H, APEX1,
SSRP1, H2AFZ, TLR3, FEN1, H2AFV, IRF3, EXOSC4, HMGN4, HMGN3, TLR9,
NUCKS1, HMGB3, TLR7, PSMC6, TCEA1 |
5.04×10−5 |
|
| Heterocyclic
compound binding | SUPT16H, APEX1,
SSRP1, H2AFZ, TLR3, FEN1, H2AFV, IRF3 EXOSC4, HMGN4, HMGN3, TLR9,
NUCKS1, HMGB3, TLR7, PSMC6, TCEA1 |
5.04×10−5 |
|
| RNA binding | SUPT16H, APEX1,
SSRP1, TLR3, EXOSC4, TLR9, NUCKS1 | 0.00416 |
|
|
| HMGB3, TLR7 |
|
|
| Double-stranded DNA
exodeoxyribonuclease activity | APEX1, FEN1 | 0.00448 |
|
| Exonuclease
activity, active with either ribo-or deoxyribonucleic acids and
producing 5′-phosphomonoesters | APEX1, FEN1,
EXOSC4 | 0.00529 |
|
| DNA binding | APEX1, SSRP1,
H2AFZ, FEN1, H2AFV, IRF3 | 0.00699 |
|
|
| HMGN4, HMGN3,
HMGB3, TCEA1 |
|
|
| RNA-DNA hybrid
ribonuclease activity | APEX1, FEN1 | 0.00699 |
|
| siRNA binding | TLR9, TLR7 | 0.00869 |
|
| Structure-specific
DNA binding | APEX1, H2AFZ, FEN1,
HMGB3 | 0.0185 |
|
| Ribonuclease
activity | APEX1, FEN1,
EXOSC4 | 0.0223 |
| Biological
process | Positive regulation
of interferon-alpha production | TLR3, IRF3, TLR9,
TLR7 |
1.08×10−5 |
|
| Positive regulation
of interferon-α biosynthetic process | TLR3, TLR9,
TLR7 |
1.47×10−5 |
|
| Positive regulation
of interferon-β production | TLR3, IRF3, TLR9,
TLR7 |
3.38×10−5 |
|
| Positive regulation
of interferon-β biosynthetic process | TLR3, TLR9,
TLR7 |
6.41×10−5 |
|
| I-κB
phosphorylation | TLR3, TLR9,
TLR7 |
2.01×10−4 |
|
| Positive regulation
of interferon-γ biosynthetic process | TLR3, TLR9,
TLR7 |
2.41×10−4 |
|
| DNA metabolic
process | SUPT16H, APEX1,
SSRP1, FEN1, EXOSC4, HMGB3, TCEA1 | 0.00154 |
|
| Cellular response
to DNA damage stimulus | SUPT16H, APEX1,
SSRP1, FEN1, IRF3, PSMC6, TCEA1 | 0.00154 |
|
| Positive regulation
of NF-κB import into nucleus | TLR3, TLR9,
TLR7 | 0.00154 |
|
| Positive regulation
of interleukin-8 production | TLR3, TLR9,
TLR7 | 0.00535 |
|
| Positive regulation
of chemokine production | TLR3, TLR9,
TLR7 | 0.00686 |
|
| Defense response to
virus | TLR3, IRF3, EXOSC4,
TLR7 | 0.00869 |
|
| Defense response to
other organism | TLR3, IRF3, EXOSC4,
TLR9, TLR7 | 0.00895 |
|
| Positive regulation
of viral transcription | SUPT16H, SSRP1,
TCEA1 | 0.0106 |
|
| I-κB kinase/NF-κB
signaling | TLR3, TLR9,
TLR7 | 0.0106 |
|
| Positive regulation
of I-κB kinase/NF-κB signaling | TLR3, IRF3, TLR9,
TLR7 | 0.0106 |
|
| Positive regulation
of interleukin-6 production | TLR3, TLR9,
TLR7 | 0.0119 |
|
| DNA repair | SUPT16H, APEX1,
SSRP1, FEN1, TCEA1 | 0.0159 |
|
| DNA
replication | SUPT16H, APEX1,
SSRP1, FEN1, | 0.0159 |
|
| Response to
stress | SUPT16H, APEX1,
SSRP1, TLR3, FEN1, IRF3, EXOSC4, TLR9, TLR7, PSMC6, TCEA1 | 0.0173 |
|
| Transcription
elongation from RNA polymerase II promoter | SUPT16H, SSRP1,
TCEA1 | 0.0191 |
|
| Multi-organism
process | SUPT16H, UMPS,
SSRP1, TLR3, EXOSC4, TLR9, TLR7, PSMC6, TCEA1 | 0.0202 |
|
| Positive regulation
of inflammatory response | TLR3, TLR9,
TLR7 | 0.0275 |
|
| Chromosome
organization | SUPT16H, APEX1,
SSRP1, FEN1, HMGN3, HMGB3 | 0.0298 |
|
| Positive regulation
of Toll-like receptor signaling pathway | TLR3, TLR9 | 0.0387 |
|
| RNA phosphodiester
bond hydrolysis | APEX1, FEN1,
EXOSC4 | 0.0388 |
|
| Telomere
maintenance via semi-conservative replication | APEX1, FEN1 | 0.05 |
Discussion
Several members of the HMGB family have been found
to promote self-renewal and colony formation in stem cells as well
as in cancer cells (22–28), indicating the presence of an
association between these two cell types; however, the detailed
molecular mechanisms underlying their cancer-promoting activity
remain obscure. HMGB3 belongs to the HMGB family, members of which
are proposed to regulate gene transcription by affecting DNA
structure and influencing transcription complex formation or by
altering gene availability (29). On
the basis of previous results, which found that HMGB3
overexpression was closely associated with to the development of
NSCLC, this protein was selected for further investigation.
The present study constructed lentiviral plasmids
encoding three different shRNAs with distinct target sequences
picked from the coding sequence of human HMGB3 gene. Verified by
RT-qPCR and western blotting, shRNA3-1 and shRNA3-2 exhibited
superior suppression of HMGB3 expression than shHMGB3-3, so these
two shRNAs were used to construct stable HMGB3-knockdown A549 cell
lines. The effect of HMGB3 knockdown on the cell viability and
apoptosis was then assessed in A549 and H1299 NSCLC cells. The
CCK-8 assay result revealed that HMGB3 knockdown could
significantly decrease A549 or H1299 cell viability, whereas
annexin V staining followed by flow cytometry analysis demonstrated
that there was an increase in the apoptosis rate of A549 or H1299
cells following HMGB3 knockdown, indicating that HMGB3 is involved
in the in vitro survival of NSCLC cells. Western blot
analysis further indicated that HMGB3 knockdown induces apoptosis
in A549 and H1299 cells, likely by activating the intrinsic
apoptosis pathway. A clonogenic assay is a widely used technique
for evaluating the influence of different experimental conditions,
including compound treatment, UV-irradiation or gene manipulation,
on the reproductive viability (or colony-forming ability) of
adherent cells (30); therefore, this
technique was used to assess the influence of HMGB3 knockdown on
the colony formation of A549 or H1299 cells. The data from this
assay revealed that HMGB3 knockdown significantly decreased the
colony-forming efficiency of these two NSCLC cells. Collectively,
these data indicated that HMGB3 has a pivotal role in NSCLC cell
survival and colony formation.
Oncogenesis is the result of uncontrolled cell
proliferation led by multiple gene deregulation, including the
abnormal activation of oncogenes and deactivation of
tumor-suppressor genes that are involved in cancer initiation,
local infiltration and metastasis (31). HMG proteins are the second most
abundant chromatin proteins, serving notable roles in regulating
chromatin structure and gene expression (32). HMG proteins can be sub-grouped into
HMGA, HMGB and HMGN super-families; among which the HMGB family
proteins contain two HMG-box DNA binding motifs that mediate
specific non-sequential DNA binding and has been identified in
different DNA-binding proteins, including transcriptions factors
(33). Besides, the HMG-box motif has
also been found to involve in protein-protein interactions. HMGB3
is a member of the HMGB super-family (33). Genecards records show that HMGB3
expression is extremely low in lung, liver, brain and urinary
tissues compared with that in other tissues and is upregulated in
the liver cancer HuH-7 and HepG2 cell lines, as well as in the
NSCLC A549 cell line and in 293 cells (https://www.genecards.org/). Initial study on HMGB3
revealed that it participated in regulating the balance between
self-renewal and differentiation in hematopoietic stem cells
(11). The oncogenic potential and
prognostic value of HMGB3 in different cancer types has also been
appreciated (34,35).
The mechanisms of HMGB3 overexpression and promoting
cancer development remain unclear. A study conducted by Zhang et
al (35) proposed that the
WNT/β-catenin pathway, a known oncogenic signaling pathway, could
be activated by HMGB3 overexpression. The results of bioinformatics
analysis performed in the present study by retrieving and analyzing
HMGB3-interacting proteins using the STRING online database
indicated that HMGB3 may interact with the FACT complex proteins
SUPT16H and SSRP1. Previous studies have suggested that the FACT
complex can have an oncogenic role in multiple types of malignant
cancer. For instance, Garcia et al (36) suggested that expression of FACT
complex proteins was associated with cancer cells' poor
differentiation and stemness. Dinant et al (37) reported that SUPT16H facilitates the
repair of DNA damage induced by UV-irradiation and promote mRNA
restart by increasing chromatin dynamics. Dermawan et al
(38) further demonstrated that the
pharmaceutical inhibition of FACT complex enhanced the anticancer
effect of an EGFR/human epidermal growth factor receptor 2
inhibitor on glioblastoma cells. On the basis of these results, we
hypothesize that HMGB3 facilitates NSCLC cell survival and
development, presumably by interacting with and increasing the
function of FACT complex proteins SUPT16H and SSRP1. However, this
hypothesis requires further investigation in future.
Collectively, the results of the present study
provided evidence of the molecular function of HMGB3 in the
initiation and development of NSCLC, contributing to research into
the molecular mechanism of its cellular features and the
development of an early diagnostic method or targeted therapeutic
agents.
Acknowledgements
Not applicable.
Funding
Hebei medical science research project 2015 Key
scientific and technological research plan (grant no.
20150210).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
NS wrote the manuscript. BW and GF participated in
cell culture and cell transfection. LD and SY participated in cell
viability, apoptosis and colony formation assay. WJ and YL
participated in RT-qPCR assay and western blot. NS participated in
statistical analysis and revised the manuscript. All authors read
and approved the final the manuscript.
Ethics approval and consent to publish
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Meza R, Meernik C, Jeon J and Cote ML:
Lung cancer incidence trends by gender, race and histology in the
United States, 1973–2010. PLoS One. 10:e01213232015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Riaz SP, Lüchtenborg M, Coupland VH,
Spicer J, Peake MD and Møller H: Trends in incidence of small cell
lung cancer and all lung cancer. Lung Cancer. 75:280–284. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen Z, Fillmore CM, Hammerman PS, Kim CF
and Wong KK: Non-small-cell lung cancers: A heterogeneous set of
diseases. Nat Rev Cancer. 14:535–546. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Song N, Liu B, Wu JL, Zhang RF, Duan L, He
WS and Zhang CM: Prognostic value of HMGB3 expression in patients
with non-small cell lung cancer. Tumour Biol. 34:2599–2603. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Barreiro-Alonso A, Lamas-Maceiras M,
Rodriguez-Belmonte E, Vizoso-Vázquez Á, Quindós M and Cerdán ME:
High mobility group B proteins, their partners, and other redox
sensors in ovarian and prostate cancer. Oxid Med Cell Longev.
2016:58450612016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Reeves R: High mobility group (HMG)
proteins: Modulators of chromatin structure and DNA repair in
mammalian cells. DNA Repair (Amst). 36:122–136. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Oka T, Sasakawa T, Komori N, Miyamoto K,
Suzuki I, Sassa T and Natori Y: Developmental changes in the
expression of HMG 2a protein. FEBS Lett. 316:20–22. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Oka T, Endo Y, Ito M, Miyamoto K, Sasakawa
T, Suzuki I and Natori Y: Molecular cloning of chick liver HMG 2a
cDNA and developmental expression of HMG 2a mRNA. Biochim Biophys
Acta. 1130:224–226. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gordon JS, Rosenfeld BI, Kaufman R and
Williams DL: Evidence for a quantitative tissue-specific
distribution of high mobility group chromosomal proteins.
Biochemistry. 19:4395–4402. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nemeth MJ, Kirby MR and Bodine DM: Hmgb3
regulates the balance between hematopoietic stem cell self-renewal
and differentiation. Proc Natl Acad Sci USA. 103:13783–13788. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nemeth MJ, Cline AP, Anderson SM,
Garrett-Beal LJ and Bodine DM: Hmgb3 deficiency deregulates
proliferation and differentiation of common lymphoid and myeloid
progenitors. Blood. 105:627–634. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nemeth MJ, Curtis DJ, Kirby MR,
Garrett-Beal LJ, Seidel NE, Cline AP and Bodine DM: Hmgb3: An
HMG-box family member expressed in primitive hematopoietic cells
that inhibits myeloid and B-cell differentiation. Blood.
102:1298–1306. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cai X, Ding H, Liu Y, Pan G, Li Q, Yang Z
and Liu W: Expression of HMGB2 indicates worse survival of patients
and is required for the maintenance of Warburg effect in pancreatic
can. Acta Biochim Biophys Sin (Shanghai). 49:119–127.
2017.PubMed/NCBI
|
|
15
|
Yu JW, Mai W, Cui YL and Kong LY: Genes
and pathways identified in thyroid carcinoma based on
bioinformatics analysis. Neoplasma. 63:559–568. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Guo S, Wang Y, Gao Y, Zhang Y, Chen M, Xu
M, Hu L, Jing Y, Jing F, Li C, et al: Knockdown of high mobility
group-box 3 (HMGB3) expression inhibits proliferation, reduces
migration, and affects chemosensitivity in gastric cancer cells.
Med Sci Monit. 22:3951–3960. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li M, Cai Y, Zhao H, Xu Z, Sun Q, Luo M,
Gu L, Meng M, Han X and Sun H: Overexpression of HMGB3 protein
promotes cell proliferation, migration and is associated with poor
prognosis in urinary bladder cancer patients. Tumour Biol.
36:4785–4792. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gao J, Zou Z, Gao J, Zhang H, Lin Z, Zhang
Y, Luo X, Liu C, Xie J and Cai C: Increased expression of HMGB3: A
novel independent prognostic marker of worse outcome in patients
with esophageal squamous cell carcinoma. Int J Clin Exp Pathol.
8:345–352. 2015.PubMed/NCBI
|
|
19
|
Elgamal OA, Park JK, Gusev Y,
Azevedo-Pouly AC, Jiang J, Roopra A and Schmittgen TD: Tumor
suppressive function of mir-205 in breast cancer is linked to HMGB3
regulation. PLoS One. 8:e764022013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak K J and Schmittgen T D: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Szklarczyk D, Morris JH, Cook H, Kuhn M,
Wyder S, Simonovic M, Santos A, Doncheva NT, Roth A, Bork P, et al:
The STRING database in 2017: Quality-controlled protein-protein
association networks, made broadly accessible. Nucleic Acids Res.
45((D1)): D362–D368. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xian L, Huso T, Belton A, Huso D and Resar
LM: High mobility group A1 chromatin remodeling protein expands the
intestinal stem cell compartment and Paneth cell niche through
Wnt/β-catenin signaling and Sox9. Am Assoc Cancer Res. 76:16–20.
2016.
|
|
23
|
Wu T, Zhang W, Yang G, Li H, Chen Q, Song
R and Zhao L: HMGB1 overexpression as a prognostic factor for
survival in cancer: A meta-analysis and systematic review.
Oncotarget. 7:50417–50427. 2016.PubMed/NCBI
|
|
24
|
Sumter TF, Xian L, Huso T, Koo M, Chang
YT, Almasri TN, Chia L, Inglis C, Reid D and Resar LM: The high
mobility group A1 (HMGA1) transcriptome in cancer and development.
Curr Mol Med. 16:353–393. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Resar L, Xian L, Huso T, Belton A, Cope L
and Huso D: High mobility group A1 chromatin remodeling protein
regulates self-renewal, niche formation and regenerative function
in adult stem cells through wnt/β-catenin signaling. Am Soc
Hematol. 128:26472016.
|
|
26
|
Kalomoiris S, Cicchetto AC, Lakatos K,
Nolta JA and Fierro FA: Fibroblast growth factor 2 regulates high
mobility group A2 expression in human bone marrow-derived
mesenchymal stem cells. J Cell Biochem. 117:2128–2137. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pallante P, Sepe R, Puca F and Fusco A:
High mobility group a proteins as tumor markers. Front Med
(Lausanne). 2:152015.PubMed/NCBI
|
|
28
|
Hayakawa K, Pham LD, Arai K and Lo EH:
High-mobility group box 1: An amplifier of stem and progenitor cell
activity after stroke. Acta Neurochir Suppl. 118:31–38.
2013.PubMed/NCBI
|
|
29
|
Stros M: HMGB proteins: Interactions with
DNA and chromatin. Biochim Biophys Acta. 1799:101–113. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rafehi H, Orlowski C, Georgiadis GT,
Ververis K, El-Osta A and Karagiannis TC: Clonogenic assay:
Adherent cells. J Vis Exp. pii:25732011.
|
|
31
|
Lee EY and Muller WJ: Oncogenes and tumor
suppressor genes. Cold Spring Harb Perspect Biol. 2:a0032362010.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Postnikov YV and Bustin M: Functional
interplay between histone H1 and HMG proteins in chromatin. Biochim
Biophys Acta. 1859:462–467. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Stros M, Launholt D and Grasser KD: The
HMG-box: A versatile protein domain occurring in a wide variety of
DNA-binding proteins. Cell Mol Life Sci. 64:2590–2606. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yamada Y, Nishikawa R, Kato M, Okato A,
Arai T, Kojima S, Yamazaki K, Naya Y, Ichikawa T and Seki N:
Regulation of HMGB3 by antitumor miR-205-5p inhibits cancer cell
aggressiveness and is involved in prostate cancer pathogenesis. J
Hum Genet. 63:195–205. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang Z, Chang Y, Zhang J, Lu Y, Zheng L,
Hu Y, Zhang F and Li X, Zhang W and Li X: HMGB3 promotes growth and
migration in colorectal cancer by regulating WNT/β-catenin pathway.
PLoS One. 12:e01797412017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Garcia H, Miecznikowski JC, Safina A,
Commane M, Ruusulehto A, Kilpinen S, Leach RW, Attwood K, Li Y,
Degan S, et al: Facilitates chromatin transcription complex is an
‘accelerator’ of tumor transformation and potential marker and
target of aggressive cancers. Cell Rep. 4:159–173. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dinant C, Ampatziadis-Michailidis G, Lans
H, Tresini M, Lagarou A, Grosbart M, Theil AF, van Cappellen WA,
Kimura H, Bartek J, et al: Enhanced chromatin dynamics by FACT
promotes transcriptional restart after UV-induced DNA damage. Mol
Cell. 51:469–479. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Dermawan JK, Hitomi M, Silver DJ, Wu Q,
Sandlesh P, Sloan AE, Purmal AA, Gurova KV, Rich JN, Lathia JD, et
al: Pharmacological targeting of the histone chaperone complex FACT
preferentially eliminates glioblastoma stem cells and prolongs
survival in preclinical models. Cancer Re. 76:2432–2442. 2016.
View Article : Google Scholar
|