Introduction
Papillary thyroid carcinoma (PTC) is the most
frequently diagnosed type of thyroid cancer, accounting for >80%
of all cases of thyroid cancer in 2010 worldwide (1). Thyroid cancer affects ~4% of all
patients with cancer in 2010 worldwide (2). However, the incidence rates of this
disease have increased >3-fold over the past three decades
(3). Therefore, thyroid cancer is
predicted to be a major type of malignancy in the near future. PTC
is usually treated with radioiodine ablation and surgery, and
treatment outcomes are usually satisfactory (4). At present, >95% of patients with PTC
live >5 years after diagnosis (4). However, on occasion, certain aggressive
forms of PTC can invade the lymph nodes, invade local or distant
tissues, or even differentiate into lethal thyroid types of cancer
(5), leading to poor survival
rates.
Long non-coding RNAs (lncRNAs), ncRNAs of >200
nucleotides in length, form a group of functional non-protein
coding RNA transcripts (6). lncRNAs
were originally considered ‘noise’, although accumulating evidence
over the past three decades has demonstrated that they are critical
determinants in both physiological processes and pathological
conditions, such as the occurrence and development of cancer
(7–9). Therefore, regulation of lncRNA
expression may provide new insights into the treatment or even
prevention of cancer (10). However,
the clinical application of lncRNAs is limited by the unknown
function of the majority of lncRNAs. It has been previously
reported that pre-ribosomal RNA antisense transcript (PAPAS) is
able to repress ribosomal RNA (rRNA) synthesis (11). Our previous transcriptome analysis
(12) revealed that PAPAS was
downregulated in PTC tissues, and its expression levels were
inversely associated with lncRNA HOXA transcript at the distal tip
(HOTTIP), which is an oncogenic lncRNA in PTC (12). The present study assessed the
involvement of PAPAS in PTC and investigated its association with
HOTTIP.
Materials and methods
Patients
The present study enrolled 58 patients (21 males and
37 females) with PTC from the Qingdao Municipal Hospital (Qingdao,
China), between January 2015 and January 2018. The mean age of the
patients was 45.2±5.2 years, (age range; 32–58 years). The
inclusion criteria for the patients were as follows: i) Patients
with PTC that had been diagnosed by imaging and histopathological
examinations; ii) had no previous history of malignancy; iii) the
patient was diagnosed with PTC, but had received no previous
therapy; and iv) had normal thyroid function. The exclusion
criteria for the patients were as follows: i) Patients had multiple
clinical disorders diagnosed in addition to PTC; and ii) had
received any form of treatment. Based on the American Joint
Committee on Cancer system (13),
there were 15, 16, 13 and 14 cases at stage I, II, III and IV,
respectively, in the present study. There were 26 cases at high
grade and 32 cases at low grade (14). A total of 20 patients with thyroid
goiter (8 males and 12 females; 34–56 years; 45.8±5.8 years) were
also selected during the same time period at the aforementioned
hospital. All patients provided written informed consent prior to
the start of the study. The present study was approved by the
Ethics Committee of Qingdao Municipal Hospital.
Specimens and cell lines
Thyroid biopsy was performed on all patients in
order to collect specimens of both tumor tissues and adjacent
(within 2 cm from the tumor tissues) healthy tissues. All specimens
were confirmed by three experienced pathologists at the
aforementioned hospital (blinded). Thyroid biopsies were also
obtained from the 20 patients with thyroid goiter. All thyroid
biopsies from patients with thyroid goiter were stored in a liquid
nitrogen sink at specimen library of the Qingdao Municipal
Hospital.
IHH-4 and HTH-83 PTC cell lines and Nthy-ori 3–1
thyroid follicular epithelial cell line were used in the present
study to perform all the in vitro cell experiments. Cells
were bought from the American Type Culture Collection. DMEM
supplemented with 100 mg/ml penicillin G, 10% FBS (Sigma-Aldrich;
Merck KGaA), and 100 U/ml streptomycin (Invitrogen; Thermo Fisher
Scientific, Inc.) was used to culture cells in an incubator at 37°C
with 5% CO2 to reach about 80% confluence
Total RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA in both tissue specimens and in
vitro cells were extracted using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to
manufacturer's instructions. Tumor and adjacent healthy tissues
were frozen in liquid nitrogen (−196°C) prior to the addition of
the TRIzol reagent. Following RNA extraction, SuperScript III
Reverse Transcriptase kit (Thermo Fisher Scientific, Inc.) was used
to perform the reverse transcription to synthesize cDNA through the
following thermal conditions: 55°C for 30 min and 80°C for 10 min.
All PCR reaction mixtures were prepared using the SYBR Green Master
mix (Bio-Rad Laboratories, Inc.). 18S rRNA was used as the
endogenous control. The 2−ΔΔCq method (15) was used to process the data. The
primer sequences used were as follows: PAPAS forward,
5′-ATGGGGCCAAGATTGTGTCT-3′ and reverse, 5′-AGACACAATCTTGGCCCCAT-3′;
HOTTIP forward, 5′-AAGGCGGTTTTACATACTGGTC-3′ and reverse,
5′-TAGCACCTGTAGTTGCCCATTCC-3′; 18S rRNA forward,
5′-GGCCCTGTAATTGGAATGAG-3′ and reverse,
5′-CCAAGATCCAACTACGAGCTT-3′. The thermocycling conditions were as
follows: 95°C for 5 min, followed by 40 cycles of 95°C for 10 sec
and 55°C for 40 sec.
Vector constructions and cell
transfection
pcDNA3.1 vectors expressing PAPAS and HOTTIP were
constructed by Sangon Biotech Co., Ltd. An empty vector (pcDNA3.1)
was used as negative control (NC). NC small interfering (si)RNA
(5′-UUCUCCGAACGUGUCACGUUU-3′) and HOTTIP siRNA
(5′-GCCGCCGUGUCCACCGGCAGCU-3′) were also obtained from Sangon
Biotech Co., Ltd. IHH-4 and HTH-83 PTC cells were harvested once
they reached 70–80% confluence, and Lipofectamine 2000 (Thermo
Fisher Scientific, Inc.) was used to transfect 10 nM plasmid or 40
nM siRNA into 106 cells. The cells were harvested 24 h
post-transfection prior to subsequent experiments.
Cell proliferation analysis
The effects of transfections on cell proliferation
were analyzed using a Cell Counting Kit-8 (CCK-8; Sigma-Aldrich;
Merck KGaA) assay according to manufacturer's instructions. Single
cell suspensions were prepared using DMEM and cell concentration
was diluted to 4×103 cells/ml. Cell culture was then
performed using a 96-well plates (100 µl per well). The plates were
incubated in an incubator at 37°C with 5% CO2, followed
by the addition of 10 µl CCK-8 solution every 24 h for 96 h. Cells
were then cultured for an additional 4 h, followed by the addition
of 10 µl DMSO. Finally, optical density values at 450 nm were
measured to analyze cell proliferation.
Statistical analysis
All statistical analysis was performed using
GraphPad Prism 6 software (GraphPad Software, Inc.). Experiments
were repeated in triplicate, and the results are presented as the
mean ± SD. Differences were assessed using one-way ANOVA followed
by Tukey's test. Associations between the expression levels of two
genes were analyzed by linear regression analysis. P<0.05 was
considered to indicate a statistically significant difference.
Results
Expression levels of PAPAS and HOTTIP
are altered in tumor tissues of patients with PTC
Differential gene expression provides strong
evidence for the involvement of certain genes in various
pathological processes. Therefore, the present study investigated
the expression of PAPAS and HOTTIP in both tumor and adjacent
healthy tissues via RT-qPCR. Compared with the adjacent healthy
tissues, expression levels of PAPAS were significantly lower in
tumor tissues and thyroid biopsies from patients with thyroid
goiter (P<0.05; Fig. 1A). By
contrast, lncRNA HOTTIP levels were upregulated in tumor tissues
and thyroid biopsies form thyroid goiter patients than in adjacent
healthy tissues (P<0.05; Fig.
1B). However, no significant differences were observed between
tumor tissues and thyroid biopsies from patients with thyroid
goiter.
PAPAS and HOTTIP are inversely
associated in PTC
Correlations between PAPAS and HOTTIP were analyzed
by linear regression analysis. It was observed that PAPAS and
HOTTIP were inversely and significantly associated in tumor tissues
(P<0.0001; Fig. 2A). However, the
expression levels of PAPAS and HOTTIP in adjacent healthy tissues
were not significantly associated (P=0.1420; Fig. 2B), indicating that PAPAS and HOTTIP
were associated specifically under pathological conditions.
PAPAS is an upstream negative
regulator of HOTTIP in PTC cell lines
Expression levels of PAPAS and HOTTIP in IHH-4,
HTH-83 and Nthy-ori 3–1 cell lines were measured by RT-qPCR.
Compared with Nthy-ori 3-1 cells, PAPAS was downregulated
(P<0.05; Fig. S1A) and HOTTIP
was upregulated (P<0.055; Fig.
S1B) in IHH-4 and HTH-83 cells. In order to further investigate
the interactions between PAPAS and HOTTIP, PAPAS and HOTTIP vectors
were transfected into the IHH-4 and HTH-83 cell lines. The RT-qPCR
results revealed that PAPAS and HOTTIP were overexpressed at 24 h
after transfection in the IHH-4 and HTH-83 cell lines (P<0.05;
Fig. 3A). Compared with the
untransfected (C) and NC groups, cells overexpressing PAPAS
exhibited significantly downregulated expression levels of HOTTIP
(P<0.05; Fig. 3B). However, the
expression level of PAPAS was not significantly altered in cells
overexpressing HOTTIP (Fig. 3C).
HOTTIP siRNA silencing was achieved in both IHH-4 and HTH-83 cells
(P<0.05; Fig. S2A). Compared
with the C and NC groups, HOTTIP siRNA silencing failed to
significantly affect the expression of PAPAS in these cells
(Fig. S2B).
Expression levels of PAPAS and HOTTIP
are affected by clinical stage
Comparison among patients with different clinical
stages revealed that expression levels of PAPAS decreased
significantly along with the increase in clinical staging
(P<0.05; Fig. 4A). In contrast,
HOTTIP expression level increased significantly with the increase
of clinical staging (P<0.05; Fig.
4B). The present results indicated that PAPAS and HOTTIP are
involved in the progression of PTC.
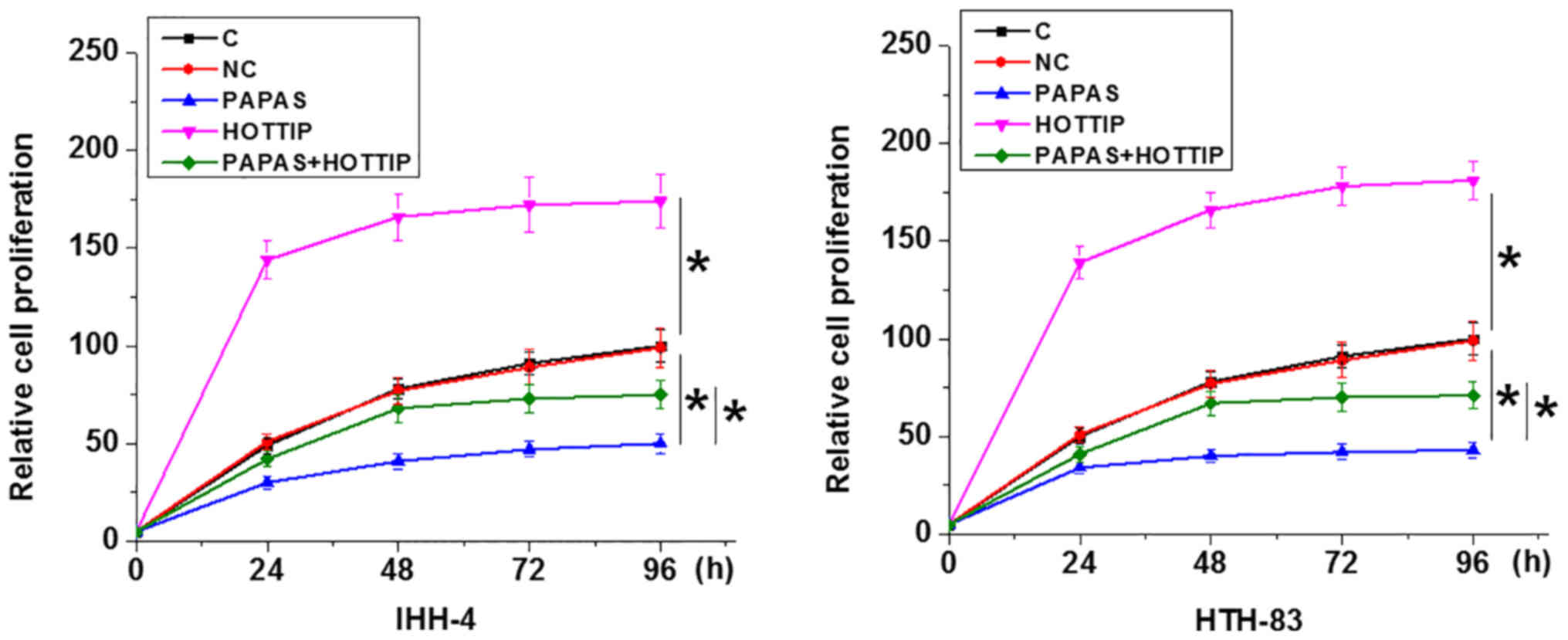
PAPAS regulates PTC cell proliferation
through HOTTIP
Compared with C and NC groups, PAPAS overexpression
led to decreased cell proliferation, whereas HOTTIP overexpression
increased the proliferation of PTC cells, and HOTTIP overexpression
attenuated the effects of PAPAS overexpression in both IHH-4 and
HTH-83 cells (P<0.05; Fig.
5).
Discussion
Although PAPAS has been reported to play a critical
role in rRNA synthesis (11), its
role in human disease remains unknown. The present study revealed
that PAPAS was downregulated in PTC tissues and PTC cell lines, and
played a tumor suppressive role in this disease. However, HOTTIP
was upregulated in both PTC tissues and PTC cell lines. In
addition, the role of PAPAS in PTC was found to be mediated by the
downregulation of oncogenic HOTTIP.
Accumulating evidence demonstrated that HOTTIP
promotes the development and progression of different types of
cancer, such as pancreatic cancer and esophageal cancer (16,17).
HOTTIP is involved in cancer development and progression primarily
by regulating cancer cell behaviors via its interactions with
downstream tumor suppression of oncogenic pathways, such as HOXA13
and the epithelial-to-mesenchymal transition pathways (16,17). In
a recent study, Yuan et al (12) reported that HOTTIP was upregulated in
PTC and was able to promote cancer cell proliferation, invasion and
migration through the downregulation of microRNA-637, which
inhibits cancer development (12).
Consistent with previous results (12), the present study observed the
upregulated HOTTIP in PTC tissues, and the overexpression of HOTTIP
led to significantly increased cancer cell proliferation.
Therefore, the present study further confirmed the oncogenic role
of HOTTIP in PTC.
To the best of our knowledge, the present study is
the first to report the tumor suppressive role of PAPAS in PTC.
lncRNAs participate in cancer biology, primarily by interacting
with downstream oncogenes or tumor suppressors (18,19).
Notably, the present study indicated that PAPAS could serve a role
in cancer development acting as an upstream inhibitor of HOTTIP in
PTC. However, the interaction between PAPAS and HOTTIP was found to
be likely indirect, due to the lack of association between the
expression levels of these two lncRNAs in tumor adjacent healthy
tissues. HOTTIP overexpression only partially attenuated the
inhibitory effects of PAPAS overexpression on cancer cell
proliferation. Therefore, it is likely that PAPAS may interact with
multiple downstream effectors to achieve a fine regulation of PTC
cell behaviors.
However, the present study is limited by the small
sample size. In addition, studies with in vivo animal models
are required in order to further validate the conclusions from the
present study. However, the present study provided evidence of the
existence of the interactions between different lncRNAs in cancer
biology. The present results provide new insights into cancer
biology. However, the molecular mechanisms underlying the
interactions between lncRNAs are unclear.
PAPAS and HOTTIP were also dysregulated in thyroid
goiter. Therefore, the interaction between PAPAS and HOTTIP is
unlikely to be cancer-specific. Future studies should aim to
investigate the involvement of PAPAS and HOTTIP in other types of
thyroid disease.
In conclusion, PAPAS was downregulated in PTC and
may inhibit PTC cell proliferation by downregulating HOTTIP.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JX and JL designed the experiments. JX and ZB
performed experiments. GX and YG analyzed the data. JL drafted the
manuscript. All authors approved the manuscript.
Ethics approval and consent to
participate
The present study was approved by Ethics Committee
of Qingdao Municipal Hospital (Qingdao, China). All patients
provided written informed consent prior to the study start.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lloyd RV, Buehler D and Khanafshar E:
Papillary thyroid carcinoma variants. Head Neck Pathol. 5:51–56.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
LiVolsi VA: Papillary thyroid carcinoma An
update. Mod Pathol. 24 (Suppl 2):S1–S9. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cancer Genome Atlas Research
NetworkIntegrated genomic characterization of papillary thyroid
carcinoma. Cell. 159:676–690. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Morris LG, Shaha AR, Tuttle RM, Sikora AG
and Ganly I: Tall-cell variant of papillary thyroid carcinoma: A
matched-pair analysis of survival. Thyroid. 20:153–158. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jendrzejewski J, Thomas A, Liyanarachchi
S, Eiterman A, Tomsic J, He H, Radomska HS, Li W, Nagy R, Sworczak
K and de la Chapelle A: PTCSC3 is involved in papillary thyroid
carcinoma development by modulating S100A4 gene expression. J Clin
Endocrinol Metab. 100:E1370–E1377. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fatica A and Bozzoni I: Long non-coding
RNAs New players in cell differentiation and development. Nat Rev
Genet. 15:7–21. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Spizzo R, Almeida MI, Colombatti A and
Calin GA: Long non-coding RNAs and cancer A new frontier of
translational research. Oncogene. 31:4577–4587. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gutschner T and Diederichs S: The
hallmarks of cancer A long non-coding RNA point of view. RNA Biol.
9:703–719. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shi X, Sun M, Liu H, Yao Y and Song Y:
Long non-coding RNAs A new frontier in the study of human diseases.
Cancer Lett. 339:159–166. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Qi P and Du X: The long non-coding RNAs, a
new cancer diagnostic and therapeutic gold mine. Mod Pathol.
26:155–165. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhao Z, Sentürk N, Song C and Grummt I:
lncRNA PAPAS tethered to the rDNA enhancer recruits
hypophosphorylated CHD4/NuRD to repress rRNA synthesis at elevated
temperatures. Genes Dev. 32:836–848. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yuan Q, Liu Y, Fan Y, Liu Z, Wang X, Jia
M, Geng Z, Zhang J and Lu X: LncRNA HOTTIP promotes papillary
thyroid carcinoma cell proliferation, invasion and migration by
regulating miR-637. Int J Biochem Cell Biol. 98:1–9. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim TH, Kim YN, Kim HI, Park SY, Choe JH,
Kim JH, Kim JS, Oh YL, Hahn SY, Shin JH, et al: Prognostic value of
the eighth edition AJCC TNM classification for differentiated
thyroid carcinoma. Oral Oncol. 71:81–86. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hakala T and Kholová I: Changes in
classification of follicular thyroid cancers. Thyroid. 26:8662016.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li Z, Zhao X, Zhou Y, Liu Y, Zhou Q, Ye H,
Wang Y, Zeng J, Song Y, Gao W, et al: The long non-coding RNA
HOTTIP promotes progression and gemcitabine resistance by
regulating HOXA13 in pancreatic cancer. J Transl Med. 13:842015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen X, Han H, Li Y, Zhang Q, Mo K and
Chen S: Upregulation of long noncoding RNA HOTTIP promotes
metastasis of esophageal squamous cell carcinoma via induction of
EMT. Oncotarget. 7:84480–84485. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Schmitt AM and Chang HY: Long noncoding
RNAs in cancer pathways. Cancer Cell. 29:452–463. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huarte M: The emerging role of lncRNAs in
cancer. Nat Med. 21:1253–1261. 2015. View
Article : Google Scholar : PubMed/NCBI
|