Introduction
Cervical cancer is the fourth most prevalent cancer
in women worldwide, behind colon, breast and lung cancer, and
represents a leading cause of tumor-associated mortality amongst
women in developing countries (1).
Notably, it is estimated that ~570,000 cervical cancer cases
occurred worldwide in 2018 (2).
Human papillomavirus (HPV) infection is a persistent infection that
is associated with the tumorigenesis of cervical cancer; in
particular, HPV type 16 (HPV16), which is present in >50% of
patients (3). Previous studies have
revealed that the distribution and prevalence of HPV genotypes
differs depending on age cohort (4–6).
Furthermore, HPV is a sexually transmitted infection and 75–80% of
sexually active individuals are likely to be infected in their
lifetime (7). Hence, the prevalence
of both HPV and cervical cancer among women may be associated with
age (8). For example, females aged
25–44 years have a demonstrated ability to clear or prevent HPV
infection and, consequently, the infection rate of HPV in these
women is lower than other age groups (9). However, over the past 10 years the
prevalence of HPV has increased among gynecological outpatients
aged between 20 and 24 years (8).
This may be due to the long-term use of contraceptives, early
coitus and an overall increase in stress levels (10,11).
Therefore, research on differences in cervical cancer between
various age groups may provide clinical guidance to help address
the increasing prevalence of cervical cancer in young women.
There are currently several treatments for patients
with cervical cancer, including radiotherapy, chemotherapy,
interventional therapy and radical hysterectomy, and the overall
survival rate of patients with cervical cancer has improved to a
certain extent (12,13). However, lymphatic metastasis,
postoperative recurrence and other toxic side effects, including
secondary infection, radiation damage and malnutrition all have an
adverse effect on therapeutic outcomes (14). Therefore, it is essential to
investigate novel and effective prognostic indicators and
therapeutic targets to improve cervical cancer therapy (15).
MicroRNAs (miRNAs) are a group of endogenous short
noncoding RNAs consisting of ~22 nucleotides that cause the
degradation of mRNA by targeting the 3′ untranslated region
(16,17). At present, several miRNAs have been
demonstrated to influence the progression of cervical cancer. For
instance, miR-152 expression is upregulated in the peripheral blood
of patients with cervical intraepithelial neoplasia, serving as an
early diagnostic and prognostic biomarker (18). Furthermore, miR-4429 sensitizes
cervical cancer cells to irradiation via the targeting of RAD51
recombinase (19). Additionally,
increasing evidence has demonstrated that the long non-coding
(lnc)RNAs serve crucial roles in the migration, differentiation,
invasion and proliferation of cells (20,21).
Notably, several lncRNAs that regulate various cellular biological
processes and influence the oncogenesis of cervical cancer have
been identified (22). For example,
the lncRNA CRNDE mediates the tumorigenesis of cervical cancer via
inhibition of p53 upregulated modulator of apoptosis expression
(PUMA) (23).
Recently, it has been demonstrated that certain
lncRNAs and mRNA can co-operate with miRNAs, downregulating the
expression of miRNAs by forming a competing endogenous (ce)RNA
network (24). Hence, the present
study hypothesized that lncRNAs may regulate mRNA expression via
co-operation with miRNAs in cervical cancer. In the present study,
the clinical data, RNA sequencing (RNA-seq)-counts and miRNA data
in different age groups were integrated to further investigate the
molecular mechanisms underlying cervical cancer development,
resulting in the identification of several potential therapeutic
targets of cervical cancer.
Materials and methods
Data source and preprocessing
The Cancer Genome Atlas (TCGA; http://cancergenome.nih.gov) is the largest database
of cancer gene information, containing data regarding 29 common
cancer types, 39 cancer subtypes and >14,500 cancer samples. The
data describe various characteristics, including miRNA, RNA-seq,
single nucleotide polymorphisms, copy number variations and
methylation patterns, as well as detailed clinical information. The
University of California Santa Cruz Genome Browser (UCSC;
http://xena.ucsc.edu) (25) serves as a tool to view the recently
assembled human genome and performs standardized processing of data
from TCGA, the International Cancer Genome Consortium (ICGC) and
Therapeutically Applicable Research to Generate Effective
Treatments (TARGET) databases.
In the present study, clinical (n=317),
RNAseq-counts (n=309) and microRNA data (n=312) were downloaded
from TCGA-cervical squamous cell carcinoma (CESC) dataset (26). Subsequently, the data were
pre-processed via matching, selecting and deleting data using the
UCSC Xena platform. The RNAseq data were then divided into 3 groups
according to the patients age: 1–40 years (84 samples), 41–60 years
(157 samples) and >60 years (65 samples). Similarly, the miRNA
data were assigned into 3 groups according to the patients' age (as
aforementioned), and the number of samples in each group was
consistent with the RNAseq data.
TCGA genes re-annotation and data
preprocessing
The HUGO Gene Nomenclature Committee (HGNC;
http://www.genenames.org) is responsible for
providing unique, standard and widespread symbols for human ncRNA
genes, protein-encoding genes and other genes. For each gene in the
human genome, there is a unique ID and symbol listed in the HGNC
database. Gene information of all symbols in the human genome were
retrieved from the HGNC database for further data preprocessing.
mRNAs and lncRNAs were distinguished from RNA-seq data for
subsequent analysis.
The ENSG (European Neuroblastoma Study Group) number
of genes in the RNAseq expression profiles from HGNC corresponded
to the gene symbol (27,28). Genes that did not match were deleted.
The clinical data were used for the subsequent survival analysis if
they matched the RNA-seq expression profile. Analogously, the
miRNAs were used for subsequent analysis if the samples were
matched with RNA-seq expression profile samples and clinical
samples.
RNA-seq Mfuzz time clustering
analysis
mRNA, lncRNA and miRNA expression matrices were
obtained from TCGA database, and the mean of the expression matrix
was calculated according to each age group. mRNA data was
eliminated if the standard deviation (SD) was <0.3; otherwise,
lncRNA and miRNA were eliminated if their SD was <0.1. The Mfuzz
package in R software (version 3.2.3) (https://www.r-project.org) was applied to perform soft
clustering analysis of noise resistance to the degeneration of DEGs
(29). In this process, the Fuzzy
C-Means clustering method was adopted to conduct clustering
analysis based on the variation of time and the change in
expression levels. Finally, multiple clustering results were
obtained and those with consistent expression changes were placed
in the same group. DEmRNAs, DElncRNAs, and DEmiRNAs were identified
in the group according to the difference in expression value
(P<0.05 was used as the cut-off value).
Enrichment analyses
The clusterProfiler (version 3.2.11; http://www.bioconductor.org/packages/release/bioc/html/clusterProfiler.html)
package of R software was used to conduct functional Gene Ontology
(http://geneontology.org) and Kyoto Encyclopedia
of Genes and Genomes (KEGG) pathway (30) functional enrichment analyses for the
DEmRNAs. P<0.05 was considered to indicate a statistically
significant difference.
Protein-protein interaction (PPI)
analysis
The Search Tool for the Retrieval of Interacting
Genes/Proteins (STRING; version 10.0; http://www.string-db.org/) database was used to search
and predict a PPI network. The database can be used for >2,000
species, containing 9.6 million different proteins and 13.8 million
types of protein interactions. Notably, the PPI network included
direct physical interactions as well as indirect functional
correlations (31). According to the
RNA-seq Mfuzz time clustering analysis, mRNAs that were
significantly associated with the age of patients with cervical
cancer were selected and used to analyze the interaction between
the mRNAs identified in the Search Tool for the Retrieval of
Interacting Genes/Proteins (STRING) database (http://www.string-db.org). mRNA data with a standard
deviation value <0.3 was eliminated. P<0.05 was considered to
indicate a statistically significant difference.
Co-expression analysis of DEmRNAs and
DElncRNAs
mRNA and lncRNA expression matrices were obtained
via RNA-seq Mfuzz time clustering analysis and co-expression was
calculated using the Pearson correlation coefficient method
(6). Using DEmRNAs and DElncRNAs
data, the co-expression association was calculated and the
mRNA-lncRNA co-expressed pairs were obtained. P<0.05 and an
absolute correlation coefficient value |r| >0.4 were considered
to indicate a significant correlation. An r-value >0 was
considered to indicate of a positive correlation, whereas r <0
was considered as a representative of a negative correlation. The
correlation strength was highest when the |r|-value was closest to
1.
Prediction of lncRNA and mRNA targeted
by miRNA
miRWalk (32,33), RNA22 (34), miRanda (http://miranda.org.uk), Targetscan (http://www.targetscan.org/vert_72), miRDB
(http://www.mirdb.org) and miRMap tools
(http://mirmap.ezlab.org) were applied to predict
mRNAs targeting miRNA. The miRWalk, miRanda, Targetscan and
RNAhybrid tools were also used for predicting lncRNAs targeting
miRNA. The target mRNAs and lncRNAs were arbitrarily selected if
they were predicted by >6 and >2 retrieval tools among the
aforementioned tools, respectively. The parameter settings in the
miRWalk tool were selected as miRNA and official symbol. Species
was set as human and the database retrieval association was set as
‘OR’ across all databases.
ceRNA network construction and mRNA
enrichment analysis
The miRNA-mRNA, miRNA-lncRNA and co-expressed
lncRNA-mRNA pairs were identified as aforementioned. Subsequently,
lncRNAs and target genes that were associated with the same miRNA
were selected, and the lncRNA-miRNA-mRNA ceRNA network was
constructed using Cytoscape software (version 3.4.0) (35,36).
Thereafter, functional analyses for crucial mRNAs located in the
ceRNA network were conducted as aforementioned. P<0.05 was
considered to indicate a statistically significant difference.
Survival analysis
The optimal cut-off of key genes was obtained
according to the expression value, survival time and survival state
based on Survminer package (version 0.4.2) (https://www.rdocumentation.org/packages/survminer/versions/0.4.2)
of R software. Genes were considered to be upregulated if the
expression value was greater than the optimal cut-off value;
conversely, genes lower than the cut-off value were considered to
be downregulated. Subsequently, the survival analysis and survival
test were performed utilizing survival in the R package (version
2.42–6). The significance cut-off was set as P<0.05. mRNAs with
a significant correlation between survival analysis and prognosis
were selected and the significant results of survival analysis were
plotted on the Kaplan-Meier curve using SPSS software (version
16.0).
Drug association analysis
The prediction of targeted drugs was performed using
the Drug-Gene Interaction (DGIdb) online database (http://www.dgidb.org/search_interactions) based on the
pre-identified mRNAs located in the ceRNA network and using the
following parameter settings, FDA approved and antineoplastic. The
interaction network of drug-target genes was constructed using
Cytoscape software.
Statistical analysis
Statistical analyses were performed using SPSS
software (version 16.0). The Pearson correlation coefficient method
was used to analyze co-expression between DEmRNAs and DElncRNAs.
Data are presented as the mean ± standard deviation. The
independent-samples student t-test was used to compare data between
two groups and the χ2 test was used to compare the count
data groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
Mfuzz time clustering analysis of
cervical cancer progression in different age groups
The design ideas and analysis process are detailed
in Fig. S1. The Mfuzz analysis
results for cervical cancer are presented in Table I. The results show that a total of
14, 11 and 16 clusters were identified for DEmRNA, DElncRNA, and
DEmiRNA data, respectively. There were 269 DEmRNAs; 30 upregulated
[including alcohol dehydrogenase 7 (ADH7)] and 239
downregulated [including vestigial-like family member 3
(VGLL3) and cytochrome P450, family 26, subfamily B,
polypeptide 1 (CYP26B1)]. A total of 274 DElncRNAs were
identified; 131 upregulated (e.g. LINC01133) and 143 downregulated,
and 16 DEmiRNAs were discovered; 8 upregulated (including miR-499a)
and 8 downregulated (including miR-3065 and −330) (Table SI). In particular, ADH7
expression was upregulated in patients >60 years, while
VGLL3 and CYP26B1 expression were downregulated in
the older age groups (Table
SII).
 | Table I.Mfuzz analysis result for cervical
cancer. |
Table I.
Mfuzz analysis result for cervical
cancer.
| Data type | Cluster(s), n | Group | Cluster
numbers | Gene number(s) |
|---|
| DEmRNA | 14 | Upregulated | 1 | 30 |
|
|
| Downregulated | 6 | 239 |
| DElncRNA | 11 | Upregulated | 2 | 131 |
|
|
| Downregulated | 2 | 143 |
| DEmiRNA | 16 | Upregulated | 4 | 8 |
|
|
| Downregulated | 5 | 8 |
PPI network with age of DEmRNAs in
cervical cancer
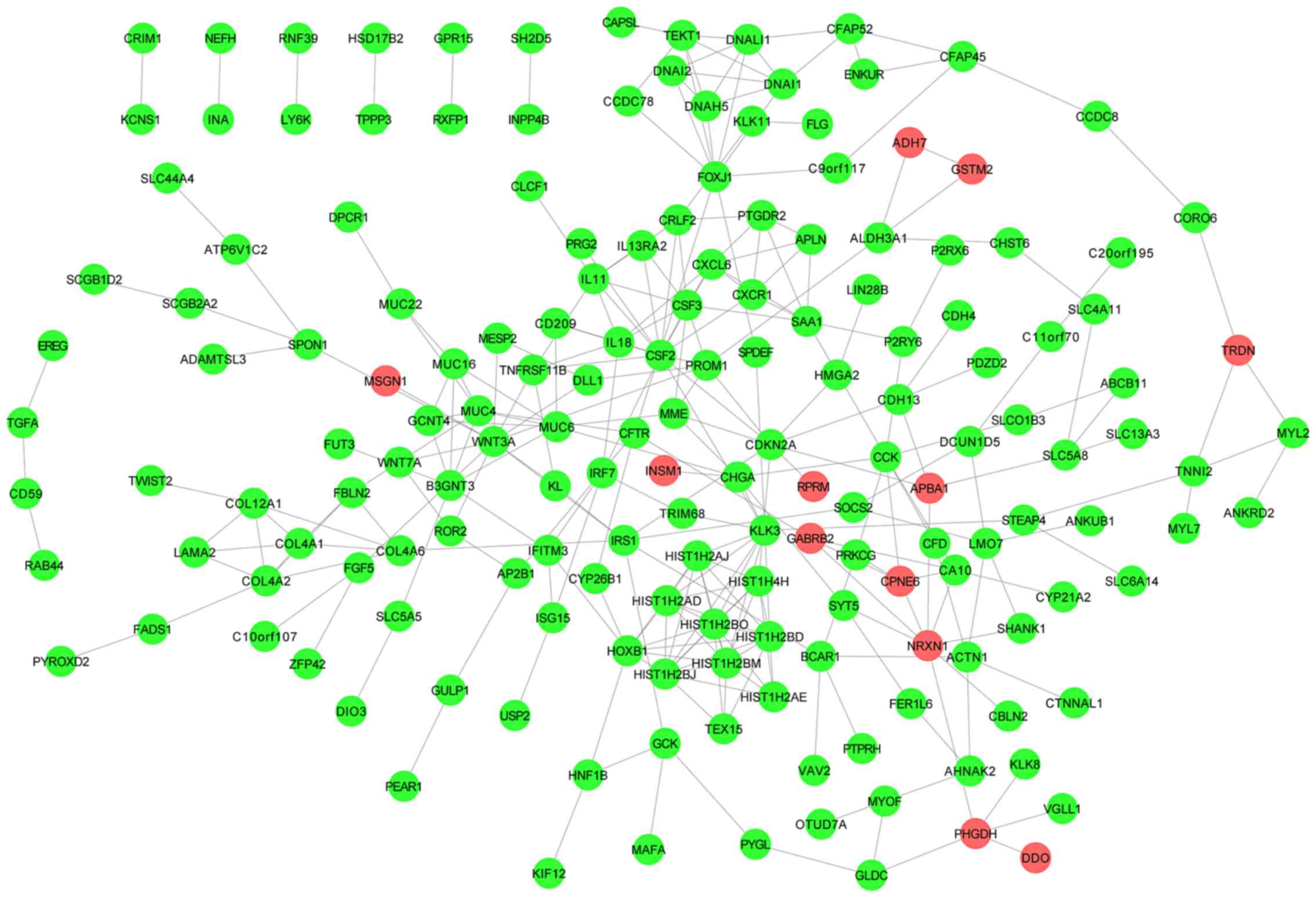
A total of 292 interaction pairs and 173 nodes
identified in the PPI network (Fig.
1). The hub nodes are presented in Table II.
| Table II.Hub nodes in the protein-protein
interaction network. |
Table II.
Hub nodes in the protein-protein
interaction network.
| Gene symbol | Degree | Betweenness | Closeness |
|---|
| CSF2 | 14.0 | 4759.71880 | 0.052211303 |
| KLK3 | 11.0 | 4843.13800 | 0.051845074 |
| WNT3A | 11.0 | 4234.45750 | 0.051328503 |
| HIST1H2BD | 10.0 | 187.71416 | 0.050535080 |
| HIST1H2BJ | 10.0 | 187.71416 | 0.050535080 |
| HIST1H2BO | 10.0 | 187.71416 | 0.050535080 |
| HIST1H2BM | 10.0 | 187.71416 | 0.050535080 |
| FOXJ1 | 9.0 | 3495.36670 | 0.049985297 |
| CDKN2A | 9.0 | 4913.64550 | 0.052372150 |
| HOXB1 | 9.0 | 1055.40840 | 0.050073640 |
Enrichment analyses with age change of
DEmRNAs in cervical cancer
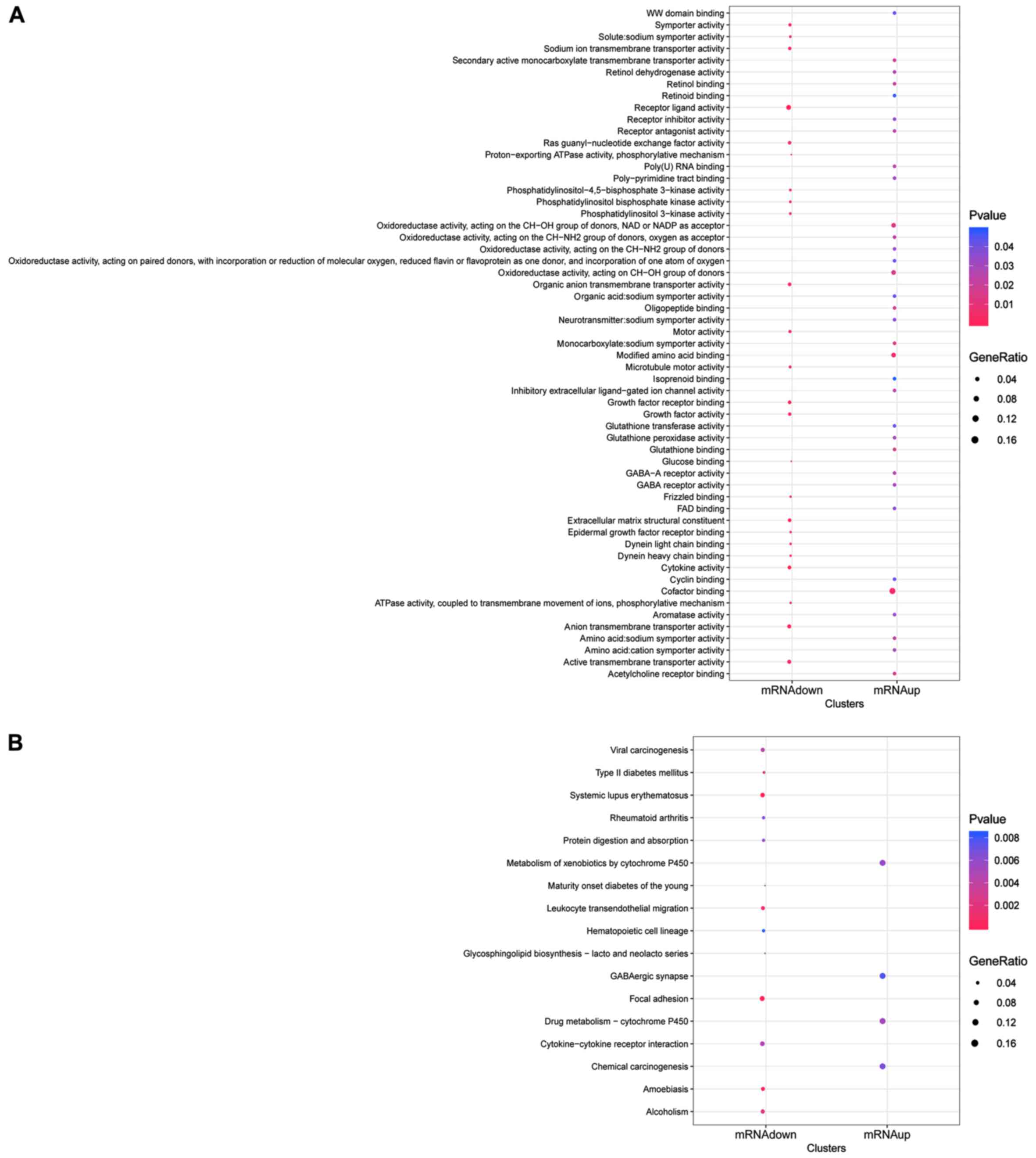
A total of 57 functional terms were obtained for the
DEmRNAs (Fig. 2A). For upregulated
DEmRNAs, the top 10 enriched functions are listed in Table III, such as ‘cofactor binding’,
‘modified amino acid binding’ (eg., ADH7, etc.) and
‘oxidoreductase activity, acting on the CH-OH group of donors, NAD
or NADP as an acceptor’. Similarly, the top 10 enriched functions
for downregulated DEmRNAs included ‘extracellular matrix structural
constituent’, ‘receptor ligand activity’ and ‘growth factor
receptor binding’. DEmRNAs were enriched in 17 KEGG pathways
(Fig. 2B). Upregulated DEmRNAs were
enriched in 4 KEGG pathways; ‘GABAergic synapse’, ‘drug
metabolism-cytochrome P450’, ‘metabolism of xenobiotics by
cytochrome P450’ and ‘chemical carcinogenesis’ (Table IV). Downregulated DEGs were enriched
in 13 KEGG pathways, (Table
IV).
| Table III.GO enrichment analysis for
differentially expressed mRNAs. |
Table III.
GO enrichment analysis for
differentially expressed mRNAs.
| A, Upregulated |
|---|
|
|---|
| Term | Description | P-value | Key genes |
|---|
| GO:0048037 | Cofactor
binding |
4.77×10−03 | CYP4Z1, DDO |
| GO:0072341 | Modified amino acid
binding |
5.07×10−03 | CPNE6, GSTM2 |
| GO:0016616 | Oxidoreductase
activity, acting on the CH-OH group of donors, NAD or NADP as
acceptor |
1.28×10−02 | ADH7, PHGDH |
| GO:0140161 |
Monocarboxylate:sodium symporter
activity |
1.42×10−02 | SLC6A12 |
| GO:0033130 | Acetylcholine
receptor binding |
1.56×10−02 | NRXN1 |
| GO:0043295 | Glutathione
binding |
1.56×10−02 | GSTM2 |
| GO:0016614 | Oxidoreductase
activity, acting on CH-OH group of donors |
1.63×10−02 | ADH7, PHGDH |
| GO:1900750 | Oligopeptide
binding |
1.70×10−02 | GSTM2 |
| GO:0015355 | Secondary active
monocarboxylate transmembrane transporter activity |
1.84×10−02 | SLC6A12 |
| GO:0019841 | Retinol
binding |
2.12×10−02 | ADH7 |
|
| B,
Downregulated |
|
| Term |
Description | P-value | Key
genes |
|
| GO:0005201 | Extracellular
matrix structural constituent |
2.56×10−06 | MUC4, FBLN2 |
| GO:0048018 | Receptor ligand
activity |
2.83×10−05 | IL18, SAA1 |
| GO:0070851 | Growth factor
receptor binding |
1.39×10−04 | CSF2, FGF5 |
| GO:0045504 | Dynein heavy chain
binding |
4.77×10−04 | DNAI2, DNAI1 |
| GO:0005125 | Cytokine
activity |
9.35×10−04 | IL18, WNT7A |
| GO:0008509 | Anion transmembrane
transporter activity |
1.86×10−03 | ABCB11, AQP6 |
| GO:0015081 | Sodium ion
transmembrane transporter activity |
2.44×10−03 | ABCB11, NALCN |
| GO:0045503 | Dynein light chain
binding |
2.44×10−03 | DNAI2, DNAH5 |
| GO:0008083 | Growth factor
activity |
2.61×10−03 | CLCF1, CSF2 |
| GO:0022804 | Active
transmembrane transporter activity |
3.41×10−03 | ATP2C2,
SLC13A3 |
| Table IV.KEGG pathway enrichment analysis for
differentially expressed mRNAs. |
Table IV.
KEGG pathway enrichment analysis for
differentially expressed mRNAs.
| A, Upregulated |
|---|
|
|---|
| Term | Description | P-value | Key genes |
|---|
| hsa00982 | Drug
metabolism-cytochrome P450 |
5.15×10−03 | ADH7, GSTM2 |
| hsa00980 | Metabolism of
xenobiotics by cytochrome P450 |
5.72×10−03 | ADH7, GSTM2 |
| hsa05204 | Chemical
carcinogenesis |
6.63×10−03 | ADH7, GSTM2 |
| hsa04727 | GABAergic
synapse |
7.77×10−03 | GABRB2,
SLC6A12 |
|
| B,
Downregulated |
|
| Term |
Description | P-value | Key
genes |
|
| hsa04510 | Focal adhesion |
4.94×10−05 | VAV2, ACTN1 |
| hsa05322 | Systemic lupus
erythematosus |
5.25×10−05 | HIST1H2AE,
HIST1H2BO |
| hsa05146 | Amoebiasis |
2.19×10−04 | CSF2, PRKCG |
| hsa04670 | Leukocyte
transendothelial migration |
5.99×10−04 | VAV2, ACTN1 |
| hsa05034 | Alcoholism |
2.27×10−03 | HIST1H2AE,
HIST1H2BO |
| hsa04930 | Type II diabetes
mellitus |
2.87×10−03 | MAFA, IRS1 |
| hsa04950 | Maturity onset
diabetes of the young |
4.44×10−03 | MAFA, GCK |
| hsa05203 | Viral
carcinogenesis |
4.47×10−03 | CDKN2A, IRF7 |
| hsa04060 | Cytokine-cytokine
receptor interaction |
4.68×10−03 | IL18, CLCF1 |
| hsa00601 | Glycosphingolipid
biosynthesis-lacto and neolacto series |
4.95×10−03 | FUT3, B3GNT3 |
| hsa04974 | Protein digestion
and absorption |
6.13×10−03 | MME, COL4A6 |
| hsa05323 | Rheumatoid
arthritis |
6.42×10−03 | IL18, CSF2 |
| hsa04640 | Hematopoietic cell
lineage |
8.37×10−03 | CSF2, CSF3 |
ceRNA network construction
In total, 632 mRNA-lncRNA interactions were
identified according to the aforementioned method, including 169
mRNAs and 106 lncRNAs (Table SIII).
A total of 1,841 pairs of miRNA-mRNA interactions (consisting of
1,518 mRNAs and 13 miRNAs) were also identified, followed by a
total of 15 overlapped mRNAs and 7 corresponding miRNAs derived
from the miRNA-mRNA and mRNA-lncRNA interactions. In addition,
90,026 pairs of miRNA-lncRNA interactions were extracted, including
15,069 lncRNAs and 16 miRNAs. Similarly, 102 overlapped lncRNAs and
15 corresponding miRNAs were extracted from those miRNA-lncRNA and
lncRNA-mRNA interactions. Finally, the lncRNA-miRNA-mRNA ceRNA
network was established utilizing those 102 lncRNAs, 15 miRNAs, 15
mRNAs and 522 interaction pairs (Fig.
3).
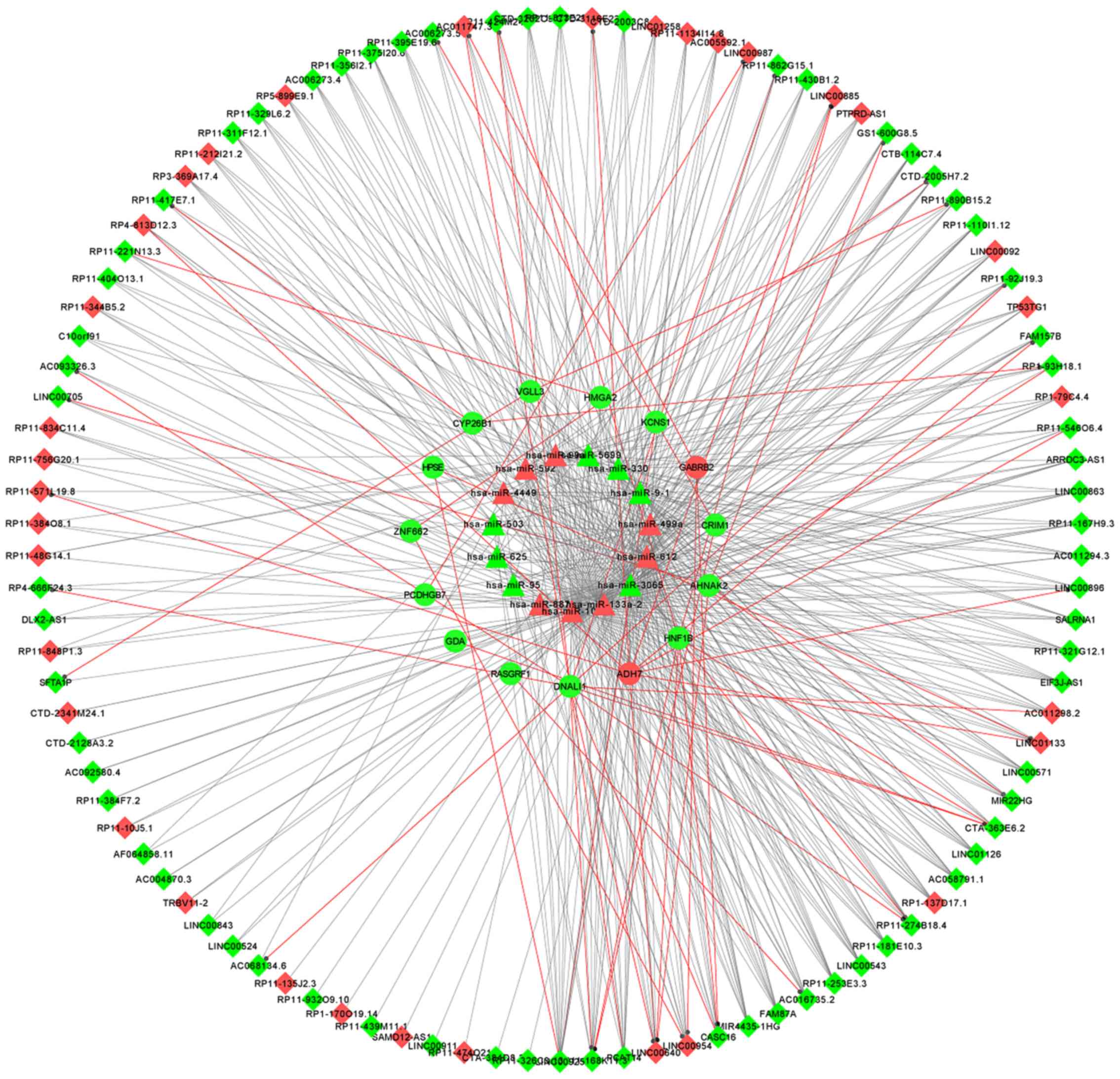 | Figure 3.ceRNA regulatory network comprising
miRNAs, mRNAs and lncRNAs. miRNA-gene, miRNA-lncRNA and
co-expressed lncRNA-mRNA pairs were identified and the
lncRNA-miRNA-mRNA ceRNA network was constructed. Thereafter,
functional analyses for crucial mRNAs located in the ceRNA network
was performed. Red and green colors represent upregulated and
downregulated genes, respectively. Triangles, circles and diamonds
represent miRNAs, mRNAs and lncRNAs, respectively. The red line and
the grey line refer to the pairs of lncRNA-mRNA interactions and
miRNA-mRNA interactions, respectively. ceRNA, competing endogenous
RNA; miRNA, microRNA; lncRNA, long non-coding RNA. |
Enrichment analysis with age change of
crucial mRNAs in cervical cancer
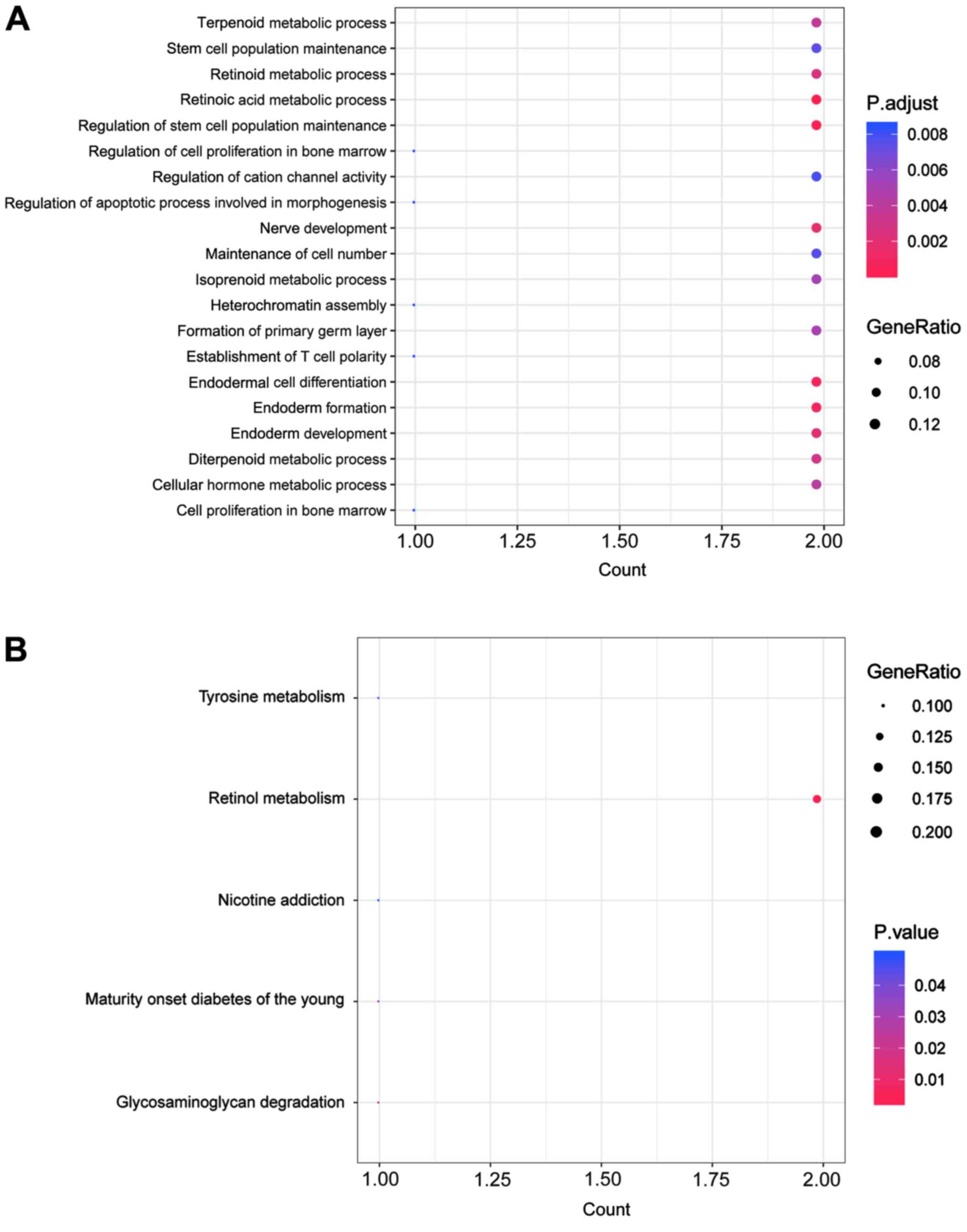
Enrichment analysis for crucial mRNA located in the
ceRNA network was implemented; the results identified 20 functional
terms (Fig. 4A) and 5 KEGG pathways
(Fig. 4B). Here, the top 10 enriched
functions of DEmRNAs are listed in Table
VA, such as the ‘retinoic acid metabolic process’ (e.g.,
ADH7, CYP26B1, etc.), ‘regulation of stem cell population
maintenance’ and ‘endodermal cell differentiation’. In addition,
the top 10 enriched KEGG pathways for DEmRNAs are listed in
Table VB, including ‘retinol
metabolism’, ‘glycosaminoglycan degradation’ and maturity onset
diabetes of the young.
| Table V.Enrichment analysis for crucial
differentially expressed mRNAs. |
Table V.
Enrichment analysis for crucial
differentially expressed mRNAs.
| A, GO analysis |
|---|
|
|---|
| Term | Description | P-value | Count | Key genes |
|---|
| GO:0042573 | Retinoic acid
metabolic process |
1.84×10−04 | 2 | ADH7, CYP26B1 |
| GO:2000036 | Regulation of stem
cell population maintenance |
2.3×10−04 | 2 | HNF1B, HMGA2 |
| GO:0035987 | Endodermal cell
differentiation |
6.83×10−04 | 2 | HNF1B, HMGA2 |
| GO:0001706 | Endoderm
formation |
9.75×10−04 | 2 | HNF1B, HMGA2 |
| GO:0021675 | Nerve
development |
1.90×10−03 | 2 | AHNAK2, GABRB2 |
| GO:0007492 | Endoderm
development |
2.00×10−03 | 2 | HNF1B, HMGA2 |
| GO:0001523 | Retinoid metabolic
process |
2.58×10−03 | 2 | ADH7, CYP26B1 |
| GO:0016101 | Diterpenoid
metabolic process |
3.00×10−03 | 2 | ADH7, CYP26B1 |
| GO:0006721 | Terpenoid metabolic
process |
3.76×10−03 | 2 | ADH7, CYP26B1 |
| GO:0034754 | Cellular hormone
metabolic process |
4.18×10−03 | 2 | ADH7, CYP26B1 |
|
| B, KEGG pathway
analysis |
|
| Term |
Description | P-value | Count | Key
genes |
|
| hsa00830 | Retinol
metabolism |
3.08×10−03 | 2 | ADH7, CYP26B1 |
| hsa00531 | Glycosaminoglycan
degradation |
2.39×10−02 | 1 | HSPE |
| hsa04950 | Maturity onset
diabetes of the young |
3.26×10−02 | 1 | HNF1B |
| hsa00350 | Tyrosine
metabolism |
4.49×10−02 | 1 | ADH7 |
| hsa05033 | Nicotine
addiction |
4.98×10−02 | 1 | GABRB2 |
| hsa00071 | Fatty acid
degradation |
5.46×10−02 | 1 | ADH7 |
| hsa00010 |
Glycolysis/Gluconeogenesis |
8.33×10−02 | 1 | ADH7 |
| hsa00982 | Drug
metabolism-cytochrome P450 |
8.33×10−02 | 1 | ADH7 |
| hsa00982 | Drug
metabolism-cytochrome P450 |
8.80×10−02 | 1 | ADH7 |
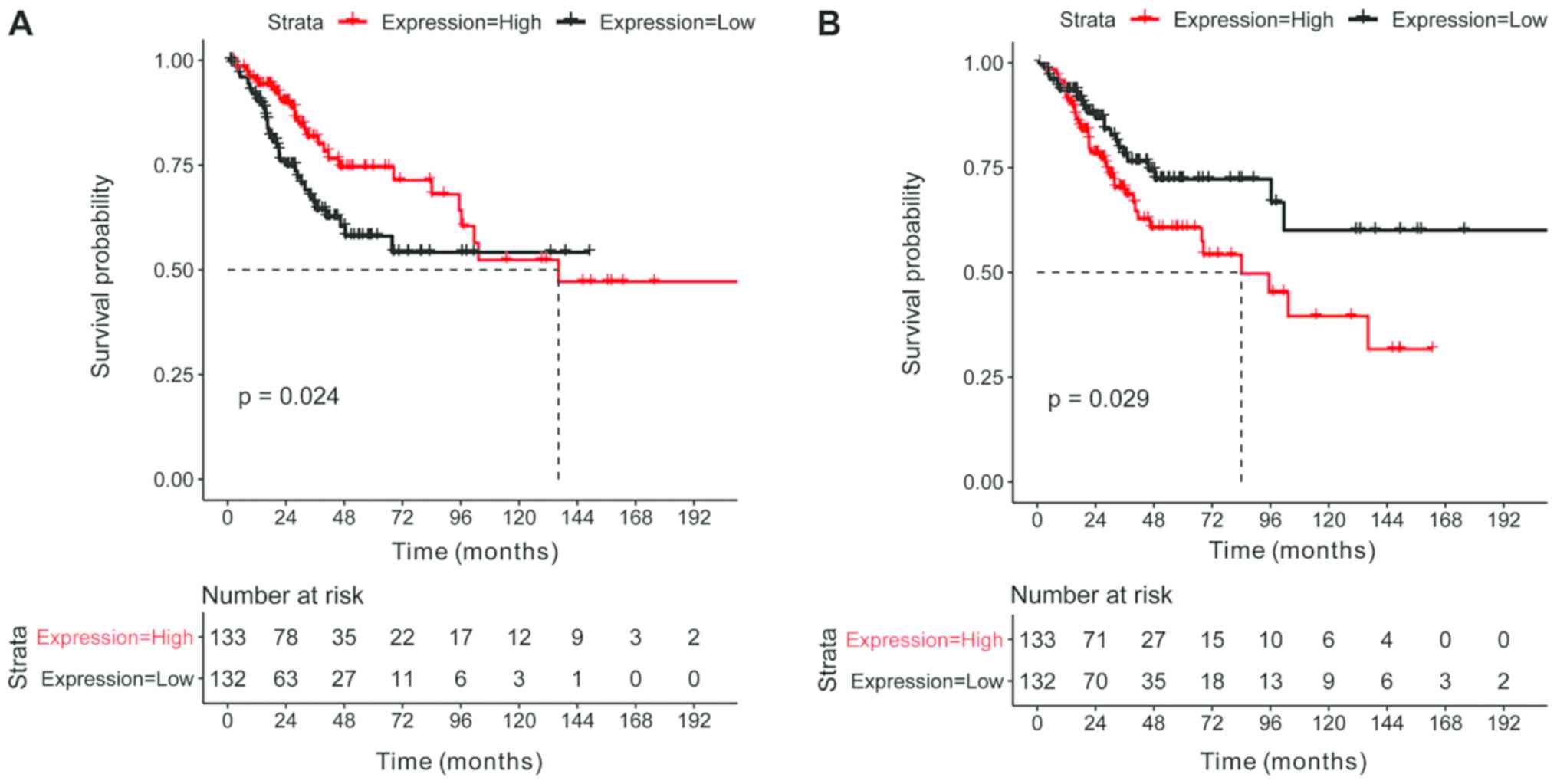
Crucial mRNA survival analysis and
drug association analysis
In total, the survival data of 265 samples were
obtained and the results are presented in Tables VI and SIV. In particular, the crucial mRNAs
ADH7 (Fig. 5A) and
VELL3 (Fig. 5B) were
significantly associated with cervical cancer survival rate
(P<0.05). The prediction of targeted drugs was performed for
ADH7 and VGLL3. While no associated drugs were
revealed to be associated with VGLL3, two drugs
(CHEMBL1161866 and ethanol) were associated with ADH7. This
allows for development of intervention targets for ADH7.
| Table VI.Survival analysis for differentially
expressed mRNAs. |
Table VI.
Survival analysis for differentially
expressed mRNAs.
| Genes | P-value | High median | Low median |
|---|
| ADH7 |
2.44×10−02a | 136.2000000 | N/A |
| AHNAK2 |
5.20×10−01 | 101.5333333 | N/A |
| CRIM1 |
5.15×10−01 | N/A | 101.53333330 |
| CYP26B1 |
7.59×10−01 | 103.2333333 | 101.53333330 |
| DNALI1 |
3.05×10−01 | 136.2000000 | 101.53333330 |
| GABRB2 |
8.53×10−01 | 103.2333333 | 136.20000000 |
| GDA |
6.01×10−01 | 103.2333333 | N/A |
| HMGA2 |
8.94×10−01 | 136.2000000 | 101.53333330 |
| HNF1B |
6.79×10−01 | 103.2333333 | 136.20000000 |
| HPSE |
5.82×10−02 | N/A | 103.23333330 |
| KCNS1 |
2.00×10−01 | N/A | 96.26666667 |
| PCDHGB7 |
5.13×10−01 | 136.2000000 | 101.53333330 |
| RASGRF1 |
6.53×10−01 | NA | 96.26666667 |
| VGLL3 |
2.87×10−02a | 84.0000000 | N/A |
| ZNF662 |
7.24×10−01 | 103.2333333 | 136.2 |
Discussion
Cervical cancer represents a leading cause of
tumor-associated mortality among women, seriously threatening the
health of females (2). It has been
revealed that the prevalence of HPV and cervical cancer among women
may be associated with their age (8). Thus, it is essential to investigate the
molecular mechanism underlying cervical cancer in different ages.
In total, 14 DEmiRs, 11 DElncRNAs and 16 DEmiRNAs were identified
based on the different age groups in the present study.
Additionally, 102 lncRNAs, 15 miRNAs and 15 mRNAs were confirmed in
the ceRNA network. In particular, ADH7 was regulated by
miR-3065, and miR-3065 was associated with LINC01133 in the ceRNA
network. Furthermore, DEmRNAs located in the ceRNA network were
involved in 5 KEGG pathways, such as ‘retinol metabolism’
(ADH7 and CYP26B1). Analogously, DEmRNAs were
enriched in several functions, such as the ‘terpenoid metabolic
process’, ‘stem cell population maintenance’ and ‘retinoic acid
metabolic process’ (ADH7 and CYP26B1). Furthermore,
ADH7 and VGLL3 were significantly associated with
cervical cancer survival rate.
It has been revealed that retinoids have the ability
to regulate cellular differentiation and proliferation, as well as
epithelial-to-mesenchymal transition via specific nuclear
retinoic-acid receptors (37,38).
Retinoids are considered as inhibitors of cervical cancer
progression due to their interaction with the HPV proteins E6 and
E7 (38). Furthermore, Gariglio
et al (39) demonstrated that
retinoid deficiency is associated with cervical squamous
metaplasia. In a recent study, lecithin retinol acyltransferase and
retinol dehydrogenase 12, which are associated with retinol
metabolism, have been identified to suppress the apoptosis,
migration and invasiveness of cervical cancer cells (40). These data from the present study
indicated that genes correlated with retinol metabolism may
influence cervical cancer progression. Whereas CYP26B1 was
downregulated in cervical cancer samples, ADH7 was
upregulated. Notably, in the current study, both CYP26B1 and
ADH7 were revealed to influence the retinol metabolism
pathway and the retinoic acid metabolic process. ADH7 is as
a member of the alcohol dehydrogenase family and, thus, may be
involved in the synthesis of retinoic acid, which is a hormone
crucial for cellular differentiation (41). It has been reported that the
upregulation of CYP26B1 may result in the progression of
cervical malignancies (42).
Following survival analysis, ADH7 was revealed to be
significantly associated with the cervical cancer survival rate.
Therefore, it was speculated that ADH7 and CYP26B1
participate in cervical cancer progression via the regulation of
the retinol metabolism pathway and the retinoic acid metabolic
process. In particular, ADH7 expression levels gradually
increased with an increase in patient age, while CYP26B1
expression gradually declined with a decrease in patient age,
indicating that ADH7 and CYP26B1 are correlated with
the differential progression of cervical cancer between different
age groups.
It has been revealed that miR-499a influences the
tumorigenesis of several different types of cancer, including
breast cancer (43) and
hepatocellular carcinoma (44). In
addition, miR-499a contributes to the migration and proliferation
of hepatitis B virus-associated hepatocellular carcinoma (HCC)
cells (45). Recently, miR-499a-5p
has been observed to promote migration, proliferation and
epithelial-to-mesenchymal transition in metastatic lung cancer cell
lines, via modulation of the mTOR pathway (46); however, the functions of miR-499 in
cervical cancer development have not yet been fully elucidated. In
the present study, miR-299a expression was upregulated in cervical
cancer samples and was revealed to downregulate CYP26B1
expression. Thus, miR-499a may be associated with the tumorigenesis
of cervical cancer via regulation of CYP26B1.
A previous study has suggested that downregulation
of lncRNA WT1 inhibited cervical carcinoma cell migration and
invasiveness via the regulation of miR-330-5p, indicating that
miR-330-5p may influence cervical cancer progression (47). Although, to the best of our
knowledge, there is currently no evidence that vestigial-like
family member 3 (VGLL3) is associated with cervical cancer
progression, the present study indicated that VGLL3 is
regulated by miR-330 and was significantly correlated with cervical
cancer survival rate. Therefore, it is speculated that VGLL3
may influence cervical cancer carcinogenesis via the regulation of
miR-330 expression. Notably, VGLL3 expression gradually
declined with decreasing patient age, suggesting that VGLL3
may be associated with the progression of cervical cancer in
different age groups.
Numerous studies have indicated that the lncRNA
LINC01133 serves as a regulator of various cancer types; for
example, LINC01133 was revealed to suppress the invasiveness,
migration and proliferation of nasopharyngeal carcinoma cells, via
interaction with Y box binding protein 1 (48). Furthermore, Zheng et al
(49) demonstrated that LINC01133
contributes to the progression of HCC via activation of the
PI3K/AKT pathway. It has also been identified that LINC01133 was
upregulated in lung squamous cell cancer cells and was associated
with poorer overall survival outcomes (50). Finally, LINC01133 was determined to
be associated with the survival time of patients with cervical
squamous cell carcinoma (26). In
the present study, LINC01133 was upregulated in cervical cancer
samples and was demonstrated to interact with miR-3065-5p.
Currently, the role of miR-3065 in cervical cancer progression is
yet to be elucidated. However, a previous study indicated that
miR-3065-5p expression is associated with the tumorigenesis of
clear cell renal cell carcinomas (51). Notably, in the present study miR-3065
expression was downregulated in cervical cancer samples and was
revealed to regulate ADH7 expression. Hence, LINC01133 may
serve as a ceRNA, via interaction with miR-3065, to upregulate
ADH7 expression, resulting in the progression of cervical
cancer.
Overall, ADH7, VGLL3, CYP26B1, miR-3065,
miR-330, miR-499a and LINC01133 may influence the progression of
cervical cancer in different age groups. These genes may therefore
represent novel targets for the diagnosis or treatment of patients
with cervical cancer. There limitations of the present study
include insufficient sample size, lack of clinical practice and
lack of research on the mechanism of intermolecular interactions.
For future studies, the predicted results should be validated using
experimental data, and the molecular mechanism of cervical cancer
carcinogenesis requires further clarification.
Supplementary Material
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was funded by Public Welfare
Application Project of Huzhou (grant no. 2016GY25).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
SD, XH, JZ, BX, LX, DG and WZ contributed to the
design of the present study. SD, XH and WZ drafted the initial
manuscript. JZ, BX, LX and DG interpreted the data. All authors
have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
DEmRNAs
|
differentially expressed mRNAs
|
|
ADH7
|
alcohol dehydrogenase 7
|
|
VGLL3
|
vestigial-like family member 3
|
|
CYP26B1
|
cytochrome P450, family 26, subfamily
B, polypeptide 1
|
|
HPV
|
human papillomavirus
|
|
miRNAs
|
microRNAs
|
|
lncRNAs
|
long noncoding RNAs
|
|
ceRNA
|
competing endogenous RNA
|
|
TCGA
|
The Cancer Genome Atlas
|
|
KEGG
|
Kyoto Encyclopedia of Genes and
Genomes
|
|
HCC
|
hepatocellular carcinoma
|
References
|
1
|
Cotto KC, Wagner AH, Feng YY, Kiwala S,
Coffman AC, Spies G, Wollam A, Spies NC, Griffith OL and Griffith
M: DGIdb 3.0: A redesign and expansion of the drug-gene interaction
database. Nucleic Acids Res. 46:D1068–D1073. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
de Lourdes Mora-García M, López-Cisneros
S, Gutiérrez-Serrano V, García-Rocha R, Weiss-Steider B,
Hernández-Montes J, Sánchez-Peña HI, Ávila-Ibarra LR, Don-López CA,
Muñóz-Godínez R, et al: HPV-16 infection is associated with a high
content of CD39 and CD73 ectonucleotidases in cervical samples from
patients with CIN-1. Mediators Inflamm. 7:46516272019.
|
|
4
|
Kjær SK, Munk C, Junge J and Iftner T:
Carcinogenic HPV prevalence and age-specific type distribution in
40,382 women with normal cervical cytology, ASCUS/LSIL, HSIL, or
cervical cancer: What is the potential for prevention? Cancer
Causes Control. 25:179–189. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bruni L, Diaz M, Castellsagué X, Ferrer E,
Bosch FX and de Sanjosé S: Cervical human papillomavirus prevalence
in 5 continents: Meta-analysis of 1 million women with normal
cytological findings. J Infect Dis. 202:1789–1799. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wheeler CM, Hunt WC, Cuzick J, Langsfeld
E, Pearse A, Montoya GD, Robertson M, Shearman CA and Castle PE;
New Mexico HPV Pap Registry Steering Committee, : A
population-based study of human papillomavirus genotype prevalence
in the United States: Baseline measures prior to mass human
papillomavirus vaccination. Int J Cancer. 132:198–207. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhao P, Liu S, Zhong Z, Hou J, Lin L, Weng
R, Su L, Lei N, Hou T and Yang H: Prevalence and genotype
distribution of human papillomavirus infection among women in
northeastern Guangdong province of China. BMC Infect Dis.
18:2042018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Luo G, Sun X, Li M, Liu T, Hu G, He Y, Mao
L, Yan L, Xie L, Zou H and Luo X: Cervical human papillomavirus
among women in Guangdong, China 2008–2017: Implication for
screening and vaccination. J Med Virol. 91:1856–1865. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu C, Zhu X, Kang Y, Cao Y, Lu P, Zhou W,
Zhou H, Zhang Y and Song Y: Epidemiology of humanpapilloma virus
infection among women in Fujian, China. BMC Public Health.
18:952017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhao FH, Tiggelaar SM, Hu SY, Xu LN, Hong
Y, Niyazi M, Gao XH, Ju LR, Zhang LQ, Feng XX, et al: A
multi-center survey of age of sexual debut and sexual behavior in
Chinese women: Suggestions for optimal age of human papillomavirus
vaccination in China. Cancer Epidemiol. 36:384–390. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dillner J, Rebolj M, Birembaut P, Petry
KU, Szarewski A, Munk C, de Sanjose S, Naucler P, Lloveras B, Kjaer
S, et al: Long term predictive values of cytology and human
papillomavirus testing in cervical cancer screening: Joint European
cohort study. BMJ. 337:969–972. 2008. View Article : Google Scholar
|
|
12
|
Biewenga P, van der Velden J, Mol BW,
Stalpers LJ, Schilthuis MS, van der Steeg JW, Burger MP and Buist
MR: Prognostic model for survival in patients with early stage
cervical cancer. Cancer. 117:768–776. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kumar L, Pramanik R, Kumar S, Bhatla N and
Malik S: Neoadjuvant chemotherapy in gynaecological cancers -
Implications for staging. Best Pract Res Clin Obstet Gynaecol.
29:790–801. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li N, Guo X, Liu L, Wang L and Cheng R:
Molecular mechanism of miR-204 regulates proliferation, apoptosis
and autophagy of cervical cancer cells by targeting ATF2. Artif
Cells Nanomed Biotechnol. 47:2529–2535. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao YB, Wang JH, Chen XX, Wu YZ and Wu Q:
Values of three different preoperative regimens in comprehensive
treatment for young patients with stage Ib2 cervical cancer. Asian
Pac J Cancer Prev. 13:1487–1489. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Beresneva EV, Rykov SV, Hodyrev DS,
Pronina IV, Ermilova VD, Kazubskaia TP, Braga EA and Loginov VI:
Methylation profile of group of miRNA genes in clear cell renal
cell carcinoma; involvement in cancer progression. Genetika.
49:366–375, (In Russian). PubMed/NCBI
|
|
17
|
White NM, Khella HW, Grigull J, Adzovic S,
Youssef YM, Honey RJ, Stewart R, Pace KT, Bjarnason GA, Jewett MA,
et al: MiRNA profiling in metastatic renal cell carcinoma reveals a
tumour-suppressor effect for miR-215. Br J Cancer. 105:1741–1749.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang D and Zhang Q: MiR-152 may function
as an early diagnostic and prognostic biomarker in patients with
cervical intraepithelial neoplasia and patients with cervical
cancer. Oncol Lett. 17:5693–5698. 2019.PubMed/NCBI
|
|
19
|
Sun H, Fan G, Deng C and Wu L: MiR-4429
sensitized cervical cancer cells to irradiation by targeting RAD51.
J Cell Physiol. 23:185–193. 2020. View Article : Google Scholar
|
|
20
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sun L, Luo H, Liao Q, Bu D, Zhao G, Liu C,
Liu Y and Zhao Y: Systematic study of human long intergenic
non-coding RNAs and their impact on cancer. Sci China Life Sci.
56:324–334. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Peng L, Yuan X, Jiang B, Tang Z and Li GC:
LncRNAs: Key players and novel insights into cervical cancer.
Tumour Biol. 37:2779–2788. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang JJ and Fan LP: Long non-coding RNA
CRNDE enhances cervical cancer progression by suppressing PUMA
expression. Biomed Pharmacother. 117:1087262019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang XZ, Cheng TT, He QJ, Lei ZY, Chi J,
Tang Z, Liao QX, Zhang H, Zeng LS and Cui SZ: LINC01133 as ceRNA
inhibits gastric cancer progression by sponging miR-106a-3p to
regulate APC expression and the Wnt/β-catenin pathway. Mol Cancer.
17:1262018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Karolchik D, Baertsch R, Diekhans M, Furey
TS, Hinrichs A, Lu YT, Roskin KM, Schwartz M, Sugnet CW, Thomas DJ,
et al: The UCSC genome browser database. Nucleic Acids Res.
31:51–54. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mao X, Qin X, Li L, Zhou J, Zhou M, Li X,
Xu Y, Yuan L, Liu QN and Xing H: A 15-long non-coding RNA signature
to improve prognosis prediction of cervical squamous cell
carcinoma. Gynecol Oncol. 149:181–187. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zerbino DR, Achuthan P, Akanni W, Amode
MR, Barrell D, Bhai J, Billis K, Cummins C, Gall A, Girón CG, et
al: Ensembl 2018. Nucleic Acids Res. 46:754–761. 2018. View Article : Google Scholar
|
|
28
|
Hubbard T, Barker D, Birney E, Cameron G,
Chen Y, Clark L, Cox T, Cuff J, Curwen V, Down T, et al: The
ensembl genome database project. Nucleic Acids Res. 30:38–41. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kumar L and E Futschik M: Mfuzz: A
software package for soft clustering of microarray data.
Bioinformation. 2:5–7. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kanehisa M and Goto S: KEGG: Kyoto
encyclopaedia of genes and genomes. Nucleic Acids Res. 8:27–30.
2000. View Article : Google Scholar
|
|
31
|
Bolstad BM, Irizarry RA, Astrand M and
Speed TP: A comparison of normalization methods for high density
oligonucleotide array data based on variance and bias.
Bioinformatics. 22:185–193. 2003. View Article : Google Scholar
|
|
32
|
Breuer K, Foroushani AK, Laird MR, Chen C,
Sribnaia A, Lo R, Winsor GL, Hancock RE, Brinkman FS and Lynn DJ:
InnateDB: Systems biology of innate immunity and beyond-recent
updates and continuing curation. Nucleic Acids Res. 41:D1228–D1233.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dweep H and Gretz N: MiRWalk2.0: A
comprehensive atlas of microRNA-target interactions. Nat Methods.
12:6972015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Megiorni F, Cialfi S, Dominici C,
Quattrucci S and Pizzuti A: Synergistic post-transcriptional
regulation of the cystic fibrosis transmembrane conductance
regulator (CFTR) by miR-101 and miR-494 specific binding. PLoS One.
6:e266012011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hansen TB, Jensen TI, Clausen BH, Bramsen
JB, Finsen B, Damgaard CK and Kjems J: Natural RNA circles function
as efficient microRNA sponges. Nature. 495:384–388. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Paraskevopoulou MD and Hatzigeorgiou AG:
Analyzing MiRNA-LncRNA interactions. Methods Mol Biol.
1402:271–286. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sporn MB and Roberts AB: Role of retinoids
in differentiation and carcinogenesis. Cancer Res. 43:3034–3040.
1983.PubMed/NCBI
|
|
38
|
Abu J, Batuwangala M, Herbert K and
Symonds P: Retinoic acid and retinoid receptors: Potential
chemopreventive and therapeutic role in cervical cancer. Lancet
Oncol. 6:712–720. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gariglio P, Gutiérrez J, Cortés E and
Vázquez J: The role of retinoid deficiency and estrogens as
cofactors in cervical cancer. Arch Med Res. 40:449–465. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu P, Chen N and Yang X: The effect of
retinol metabolism related genes LRAT/RDH12 in the progress of
squamous cervical cancer cell line. Prog Obstet Gynecol. 2018.
|
|
41
|
Jelski W, Chrostek L, Zalewski B and
Szmitkowski M: Alcohol dehydrogenase (ADH) isoenzymes and aldehyde
dehydrogenase (ALDH) activity in the sera of patients with gastric
cancer. Dig Dis Sci. 53:2101–2105. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Osanai M and Lee GH: Increased expression
of the retinoic acid-metabolizing enzyme CYP26A1 during the
progression of cervical squamous neoplasia and head and neck
cancer. BMC Res Notes. 7:1–9. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kabirizadeh S, Azadeh M, Mirhosseini M,
Ghaedi K and Tanha HM: The SNP rs3746444 within mir-499a is
associated with breast cancer risk in Iranian population. J Cell
Immunother. 2:95–97. 2016. View Article : Google Scholar
|
|
44
|
Wang XH, Wang FR, Tang YF, Zou HZ and Zhao
YQ: Association of miR-149C>T and miR-499A>G polymorphisms
with the risk of hepatocellular carcinoma in the Chinese
population. Genet Mol Res. 13:5048–5054. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Xiang Z, Wang S and Xiang Y: Upregulated
MicroRNA499a by hepatitis B virus induced hepatocellular
carcinogenesis via targeting MAPK6. PLoS One. 9:e1114102014.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
He S, Li Z, Yu Y, Zeng Q, Cheng Y, Ji W,
Xia W and Lu S: Exosomal miR-499a-5p promotes cell proliferation,
migration and EMT via mTOR signaling pathway in lung
adenocarcinoma. Exp Cell Res. 379:203–213. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Cui L, Nai M, Zhang K, Li L and Li R:
lncRNA WT1-AS inhibits the aggressiveness of cervical cancer cell
via regulating p53 expression via sponging miR-330-5p. Cancer Manag
Res. 11:651–667. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhang W, Du M, Wang T, Chen W, Wu J, Li Q,
Tian X, Qian L, Wang Y, Peng F, et al: Long non-coding RNA
LINC01133 mediates nasopharyngeal carcinoma tumorigenesis by
binding to YBX1. Am J Cancer Res. 9:779–790. 2019.PubMed/NCBI
|
|
49
|
Zheng YF, Zhang XY and Bu YZ: LINC01133
aggravates the progression of hepatocellular carcinoma by
activating the PI3K/AKT pathway. J Cell Biochem. 120:4172–4179.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhang J, Zhu N and Chen X: A novel long
noncoding RNA LINC01133 is upregulated in lung squamous cell cancer
and predicts survival. Tumor Biol. 36:7465–7471. 2015. View Article : Google Scholar
|
|
51
|
Müller S and Nowak K: Exploring the
miRNA-mRNA regulatory network in clear cell renal cell carcinomas
by next-generation sequencing expression profiles. Biomed Res Int.
2014:948408. 2014. View Article : Google Scholar : PubMed/NCBI
|