Introduction
Epithelial ovarian cancer (EOC) is the most lethal
type of gynecological malignancy and ranks as the 5th leading cause
of cancer-associated mortality among women in the USA (1), accounting for 22,240 new cases and
14,070 deaths in 2018 (2). Despite
intensive treatment options, including debulking surgery, platinum
and taxane-based chemotherapy and targeted therapy, such as
poly(ADP-ribose) polymerase inhibitors (PARPis), angiogenesis
inhibitors and immunotherapy agents, the overall 5-year survival
rate for all types of EOC is relatively low (47.4%) and has
remained stagnant for >2 decades (3). Additionally, ~60% of patients with EOC
possess distant metastases at initial diagnosis, and the 5-year
survival for these patients is considerably lower at 26% (2). There are a number of reasons for these
poor survival outcomes, including the absence of reliable and
accurate screening tests and limited effectiveness of current
chemotherapies (1,2). Cumulative toxicity, cross-resistance to
chemotherapies and compromised quality of life are additional
serious clinical challenges for patients with EOC. Therefore,
developing more effective and less toxic therapeutic strategies
that target the fundamental vulnerabilities of EOC is required to
improve patients' outcomes and quality of life.
PARPis are a new class of oncology drugs that are
transforming the management of EOC (4). PARPis exert anti-cancer properties by
trapping PARP on DNA at the sites of single-strand breaks, which
leads to DNA repair defects and the generation of DNA DSBs that
require homologous recombination (HR) mediated by BRCA1, BRCA2 and
other proteins (such as ATM, ATR, RAD51, CHK1 and FANCA) (5–7).
Therefore, BRCA1/2 mutant or HR-deficient cells are exceptionally
sensitive to PARP inhibition (7–9), and the
combination of two genetic deficiencies (e.g., BRCA1/2 and PARP)
leads to synthetic lethality in cancer cells. Based on the
promising clinical efficacy and the manageable toxicity profile of
PARPis in patients with advanced EOC (10–13),
three PARPis (olaparib, rucaparib and niraparib) were approved by
the U.S. Food and Drug Administration, either as a monotherapy
(olaparib and rucaparib) for women with heavily pretreated germline
BRCA-mutated (gBRCAm) EOC or as a maintenance therapy (olaparib,
rucaparib and niraparib) for women with platinum-sensitive
recurrent EOC regardless of BRCA or HR-deficiency (HRD) status. In
addition, a recent Phase III multicenter study (NCT01844986)
(12) revealed that olaparib
maintenance monotherapy significantly improved progression-free
survival times in women newly diagnosed with advanced ovarian
cancer who harbored a BRCA1/2 mutation (14). However, new challenges have arisen
for PARPi therapy. Mutations in BRCA1 or BRCA2 occur in <20% of
patients with EOC (16% germline and 4% somatic) (15,16). The
majority of patients with EOC are BRCA1/2 wild-type carriers who
respond much less favorably to PARPis, limiting the clinical
efficacy and utility of PARPis. Additionally, although several
clinical studies have shown that some non-gBRCAm or patients with
HRD negative cancer can benefit from PARPis (13,17),
developing effective prediction tools independent of HRD is
difficult, due to the lack of predictive biomarkers, which makes
patient selection challenging. The combination of a PARPis and
chemotherapy have yet to show significant clinical benefits, and
enhanced myelosuppression, as the main dose-limiting toxicity, has
been observed (18,19), which may limit future combinatorial
use of these two types of therapy. As such, developing novel PARPi
combination therapies with a broad efficacy and low toxicity is one
potential direction for improving treatment of patients with
EOC.
Previously, it has been shown that ascorbate
(vitamin C) when used in high intravenous doses (IVC), has
potential as a therapeutic agent for the treatment of a variety of
different types of cancer (20–25).
High-dose IVC, in contrast to oral doses, establishes
pharmacological concentrations in the millimolar range in tissues,
and selectively kills cancer cells by generating
H2O2 in the extracellular fluid, while
leaving healthy cells unharmed (21–23). The
exquisite selectivity of pharmacological ascorbate suggests a low
toxicity of IVC treatment. Multiple early phase clinical trials in
patients with solid or hematological malignancies, where IVC was
used alone or in combination with conventional chemotherapies or
radiation therapy, demonstrated that IVC was safe, well tolerated
and did not increase the toxicities of standard therapies (20,25–28). The
authors of the present study first demonstrated a notable decrease
in chemo-associated toxicities by adding IVC to standard
carboplatin/paclitaxel chemotherapy in patients with stage III or
IV EOC (20). In addition, the
preliminary clinical benefits in prolonged relapse time and/or
tumor responses by adding IVC to standard chemo- or radiation
therapy has been demonstrated (20,25).
By generating H2O2,
pharmacological ascorbate damages DNA and preferentially kills
cancer cells (20,29). Therefore, it was hypothesized that
the combination of pharmacological ascorbate and PARPis may enhance
DNA repair deficiency, and thus enhance the therapeutic effect of
either agent alone against EOC, regardless of BRCA status. In the
present study, the DNA damage response (DDR) induced by
pharmacological ascorbate in ovarian cancer cells bearing wild-type
BRCA1/2 was characterized, and the efficacy and feasibility of the
combination treatment of pharmacological ascorbate and the PARPi,
olaparib, in preclinical models of EOC harboring wild-type BRCA
were investigated.
Materials and methods
Cell culture and reagents
Human EOC cell lines OVCAR8 and SHIN3 were kindly
provided by Dr Peter Eck (University of Manitoba, Manitoba, Canada)
and OVCAR3, OVCAR5, OVCAR10, SKOV3, A2780 and HIO-80 (an
immortalized, nontumorigenic human ovarian epithelium cell line)
were kindly provided by Dr Thomas Hamilton, or were derived by Dr
Andrew K. Godwin, both of the Fox Chase Cancer Center
(Philadelphia, USA). SHIN3 cells were maintained in DMEM
supplemented with 10% FBS, (both Sigma-Aldrich; Merck KGaA) and 1%
penicillin-streptomycin. HIO-80 was cultured in M199/MCDB105 medium
(1:1, v/v) containing 4% FBS, insulin (0.3 U/ml) and 2 mM
L-glutamine. The remaining cell lines were cultured in PRMI-1640
medium supplemented with 10% FBS and 1% penicillin-streptomycin.
All cells were cultured at 37°C with 5% CO2 and 85–95%
humidity. Cell line authentication was carried out by the Clinical
Molecular Oncology Laboratory of University of Kansas Medical
Center (Kansas, USA) using multiplex short tandem repeat DNA
profiling.
L-Ascorbic Acid (Thermo Fisher Scientific, Inc.) was
prepared as 1 M stock solutions in sterile water, with sodium
hydroxide added drop-wise to adjust the pH to 7.0. Aliquots were
stored at −80°C and thawed for single use. Catalase (Sigma-Aldrich;
Merck KGaA) was prepared in distilled water at 10,000 units/ml, and
was used at a working concentration of 600 units/ml. Olaparib and
veliparib were obtained from Selleck Chemicals and were prepared in
dimethyl sulfoxide (DMSO) and diluted with cell culture media to
working concentrations, for the in vitro experiments. For
the in vivo experiments, olaparib was dissolved in PBS
containing 10% 2-hydroxy-propyl-betacyclodextrin (Sigma-Aldrich;
Merck KGaA). All other reagents and chemicals were obtained from
Thermo Fisher Scientific, Inc., unless specifically indicated.
BRCA1/2 mutation analysis
The BRCA1/2 wild-type status was reported previously
(30) for all the EOC cell lines
used in the present study except SHIN3. The genomic DNA of SHIN3
cells was extracted using a Blood & Cell Culture DNA Mini kit
(Qiagen GmbH). The largest and functionally most important exon
(exon 11) of both BRCA1 (3,630 bp) and BRCA2 (5,018 bp) was
amplified from the genomic DNA template using PCR as previously
described (31). The PCR amplicons
were submitted to Genewiz, Inc. for DNA sequencing. The primer
sequences are provided in Table SI.
The thermocycling conditions and Taq enzyme used were as previously
described (31). DNA sequences were
analyzed using the DNASTAR analysis package (version 8.1; DNASTAR,
Inc.). Both nucleic acid and amino acid sequences were aligned
using BioEdit (version 7.2) (32).
MTT assay
Cells were seeded at a density of 1×104
cells per well in a 96 well plate, and incubated overnight. Cells
were then exposed to a serial dilution of ascorbate (0–3.5 mM),
olaparib (0–1,000 µM in SHIN3 cells; 0–800 µM in OVCAR5 cells) and
veliparib (0–1,000 µM in SHIN3 cells; 0–800 µM in OVCAR5 cells), or
treatment combinations and incubated for 24 or 48 h. In the drug
combination groups, either olaparib or veliparib was added 15 min
prior to ascorbate treatment. Following treatment, the culture
medium was replaced with fresh, drug-free medium, and cells were
incubated with MTT for 4 h. Formazan crystals were dissolved using
DMSO and the absorbance at 492 nm was measured on a Synergy™ 4
Hybrid microplate reader (BioTek Instruments, Inc.). The half
maximal inhibitory concentration (IC50) was determined
using a non-linear regression analysis to fit the data to the
log10 [inhibitor] compared with a normalized response
with a variable slope model.
Different concentrations of ascorbate (ranging from
0–5 mM) were used to avoid drawing conclusions from a single
particular concentration. Concentration at IC50 or a
concentration range including the IC50 were used. If the
treatment time was <48 h, concentrations >IC50
were used, with additional multiple concentrations including at
least one close to or lower than the IC50. The in
vitro concentration ranges used in the present study are easily
achievable in patients by intravenous ascorbate infusion (26).
Poly(ADP-ribose) (PAR) level
measurement
PAR levels were measured using a HT PARP in
vivo Pharmacodynamic assay II (Trevigen, Inc.), and normalized
to the protein contents. Protein concentrations of cell lysates
were measured using a Pierce bicinchoninic acid assay kit (Thermo
Fisher Scientific, Inc.).
Western blot analysis
Cells were lysed in ice-cold
radioimmunoprecipitation buffer (Thermo Fisher Scientific, Inc.),
supplemented with cOmplete™ Mini Protease Inhibitor Cocktail
Tablets (Sigma-Aldrich, Merck KGaA) and Halt™ Phosphatase Inhibitor
Cocktail (Thermo Fisher Scientific, Inc.). Protein concentration
was determined using the Bradford Protein Assay Kit (Bio-Rad,
Inc.). A total of 60 µg protein/lane was resolved on the 4–20%
Mini-PROTEAN TGX™ Precast gels (Bio-Rad, Inc.) and transferred onto
polyvinylidene difluoride (PVDF) membranes (Bio-Rad, Inc.). The
membranes were blocked using 5% skim milk in TBST (20 mM Tris_HCl,
pH 7.4, 150 mM NaCl, 0.1% Tween 20) for 1 h at 4°C, followed by
incubation at 4°C overnight with specific antibodies against
H2AX (1:500; Cell Signaling Technology, Inc.; cat. no.
7631); p-H2AXSer139 (1:1,000; Cell Signaling
Technology, Inc.; cat no. 9718); ATM (1:1,000; Cell Signaling
Technology, Inc.; cat. no. 2873); p-ATMSer1981 (1:500;
Cell Signaling Technology, Inc.; cat. no. 13050); BRCA1 (1:1,000;
Cell Signaling Technology, Inc.; cat. no. 14823); BRCA2 (1:1,000;
R&D Systems, Inc.; cat. no. MAB2476); RAD51 (1:1,000; Cell
Signaling Technology, Inc.; cat. no. 8875); Ku70 (1:1,000; Cell
Signaling Technology, Inc.; cat. no. 4588); Ku80 (1:1,000; Cell
Signaling Technology, Inc.; cat. no. 2,180);
p-DNA-PKcsThr2609 (1:350; Thermo Fisher Scientific,
Inc.; cat. no. PA5-12913); DNA-PKcs (1:4,000; Santa Cruz; cat. no.
sc-9051); β-actin (1:5,000; Thermo Fisher Scientific, Inc.; cat.
no. MA5-15739); or vinculin (1:1,000; Cell Signaling Technology,
Inc.; cat. no. 13901). Target proteins were visualized using
horseradish peroxidase (HRP)-conjugated goat-anti-rabbit IgG
(1:5,000; Cell Signaling Technology, Inc.; cat. no. 7,074) or
HRP-conjugated horse-anti-mouse IgG (1:5,000; Cell Signaling
Technology, Inc.; cat. no. 7076) for 1 h at room temperature with
Pierce™ ECL Plus Western blotting substrate (Thermo Fisher
Scientific, Inc.). Each western blot analysis was performed.
b-actin or vinculin were used as the loading controls. In vivo
xenograft mouse model. All procedures were performed in
accordance with a protocol (ACUP #2018-2443) approved by the
Institutional Animal Care and Use Committee of the University of
Kansas Medical Center (Kansas, USA). The intraperitoneal (i.p.)
tumor xenografts were established via i.p. injection of
2×106 SHIN3 cells suspended in 200 µl PBS in fifty
4–6-week-old female athymic NCr-nu/nu mice (20–25 g body weight;
National Cancer Institute). A total of 2 weeks after cell
injection, mice were randomly grouped as follows: i) Control group;
i.p. injection of saline solution osmotically equivalent to
ascorbate twice daily and olaparib's solvent (PBS containing 10%
2-hydroxy-propyl-betacyclodextrin) once daily in volumes equivalent
to the olaparib treated group; ii) ascorbate group, i.p. injection
of ascorbate at 4 g/kg twice daily; iii) olaparib group, i.p.
injection of olaparib at 50 mg/kg once daily; and iv) combination
of ascorbate and olaparib, which were prepared and administered in
the same manner as individual drug treatments. After 25 days of
treatment, all mice were euthanized by CO2 inhalation in
a closed chamber (20% volume/min) followed by bilateral thoracotomy
as approved in the protocol, and gross necropsy was performed with
tumor weights and ascites volumes measured, and the number of tumor
cells in the ascetic fluids counted. The total tumor burden of each
mouse was indicated as total tumor weight at the end of experiment.
The liver, kidney and spleen from each group were subjected to
histopathological analysis using hematoxylin and eosin (H&E)
staining, on 4-µm tissue sections with an automated procedure, as
previously reported (33).
Statistical analysis
All statistical analyses were performed using
GraphPad Prism version 5.0 (GraphPad Software, Inc.). Multiple
comparisons between groups were performed using a one-way ANOVA
with a post-hoc Turkey's test with a family-wise error rate of
0.05. Adjusted P<0.05 was considered to indicate a statistically
significant difference.
Results
Pharmacological ascorbate induces
H2O2-dependent cytotoxicity in BRCA1/2
wild-type EOC cells
Exon 11 is the largest and most functionally
important exon. To the best of our knowledge, the current study was
the first to sequence the BRCA1 and 2 genes (both of
exon 11) of SHIN3 cells. As indicated in Fig. S1, a single variant of A→G at codon
349 of BRCA1 was detected, which is not known to have a
functional outcome or be associated with breast or ovarian cancer.
In addition, two variants were detected at codon 2660 A→G and codon
4560 G→C, in exon 11 of BRCA2, which do not result in amino
acid changes. The present results indicate that no functional
mutations were detected in exon 11 of BRCA1 and 2 in
SHIN3 cells. A panel of BRCA1/2 wild-type human ovarian cancer cell
lines (A2780, OVCAR10, OVCAR3, OVCAR5, SKOV3, OVCAR8 and SHIN3)
(34), and an immortalized,
non-tumorigenic human ovarian epithelium cell line (HIO-80) were
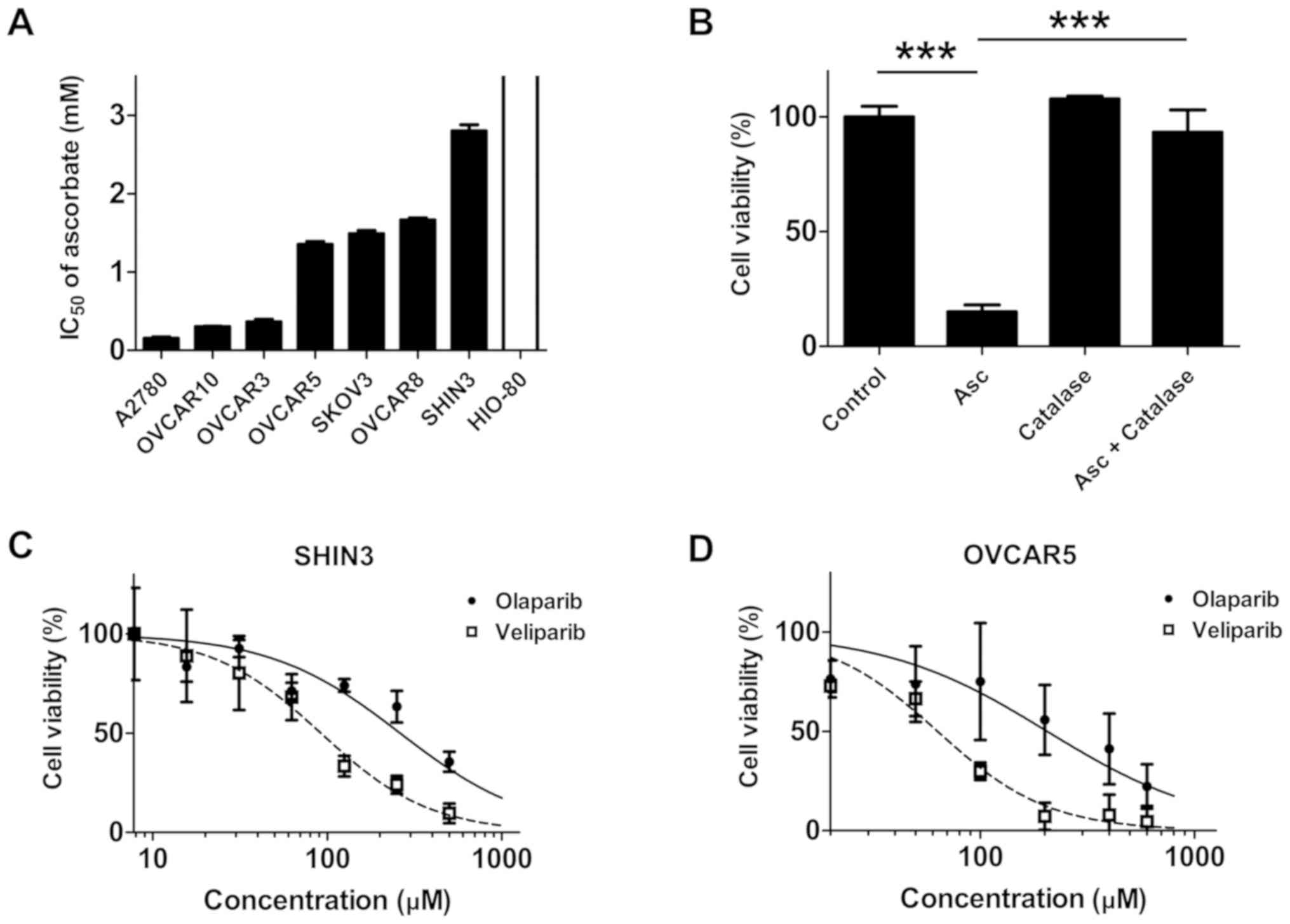
then screened for sensitivity to ascorbate. As presented in
Fig. 1A and Table SII, the IC50 values of
ascorbate in the ovarian cancer cells ranged from 0.15–2.80 mM,
which is readily achievable by i.v. ascorbate infusion (23,26). In
contrast, HIO-80 cells were resistant to ascorbate treatment in the
tested concentration range (0.0–3.5 mM), with an IC50
value >3.5 mM (Fig. 1A). When
catalase, a H2O2 scavenger, was added to the
culture medium, SHIN3 cells were protected from the cytotoxic
effects of ascorbate (P<0.001; Fig.
1B), suggesting that the ascorbate cytotoxicity was mediated
through H2O2, consistent with previously
published studies (21,23).
The cell lines exhibiting the highest levels of
resistance to ascorbate (SHIN3) and a moderately resistant cell
line OVCAR5 were selected as representative cell lines, and their
sensitivity to the PARPis olaparib and veliparib. In agreement with
their BRCA wild-type status, both SHIN3 and OVCAR5 cells exhibited
resistance to the PARPi treatments, as the concentrations of PARP
required to exert inhibitory effects on cell viability were
particularly high (35) (Fig. 1C and D; Table SII).
Pharmacological ascorbate in
combination with PARPis synergistically inhibits the growth of
BRCA1/2 wild-type EOC cells
Since ascorbate-induced H2O2
damage to DNA in EOC cells (20) and
PARPis impair DNA damage repair, it was hypothesized that the
combination of pharmacological ascorbate and PARPis may enhance DNA
repair deficiency and improve therapeutic efficacy against EOC. In
order to verify this hypothesis, the effects of the combination
treatment of pharmacological ascorbate and PARPi olaparib or
veliparib in SHIN3 and OVCAR5 cells were determined. Treatment with
olaparib (20 µM) or veliparib (20 µM) for 24 and 48 h,
respectively, minimally affected the cell viability of SHIN3 and
OVCAR5 (Fig. 2A and B). The combined
treatment of pharmacological ascorbate with either olaparib or
veliparib significantly decreased cell viability compared with
either single drug treatment and vehicle control, in both SHIN3 and
OVCAR5 cells (Fig. 2A and B). PAR
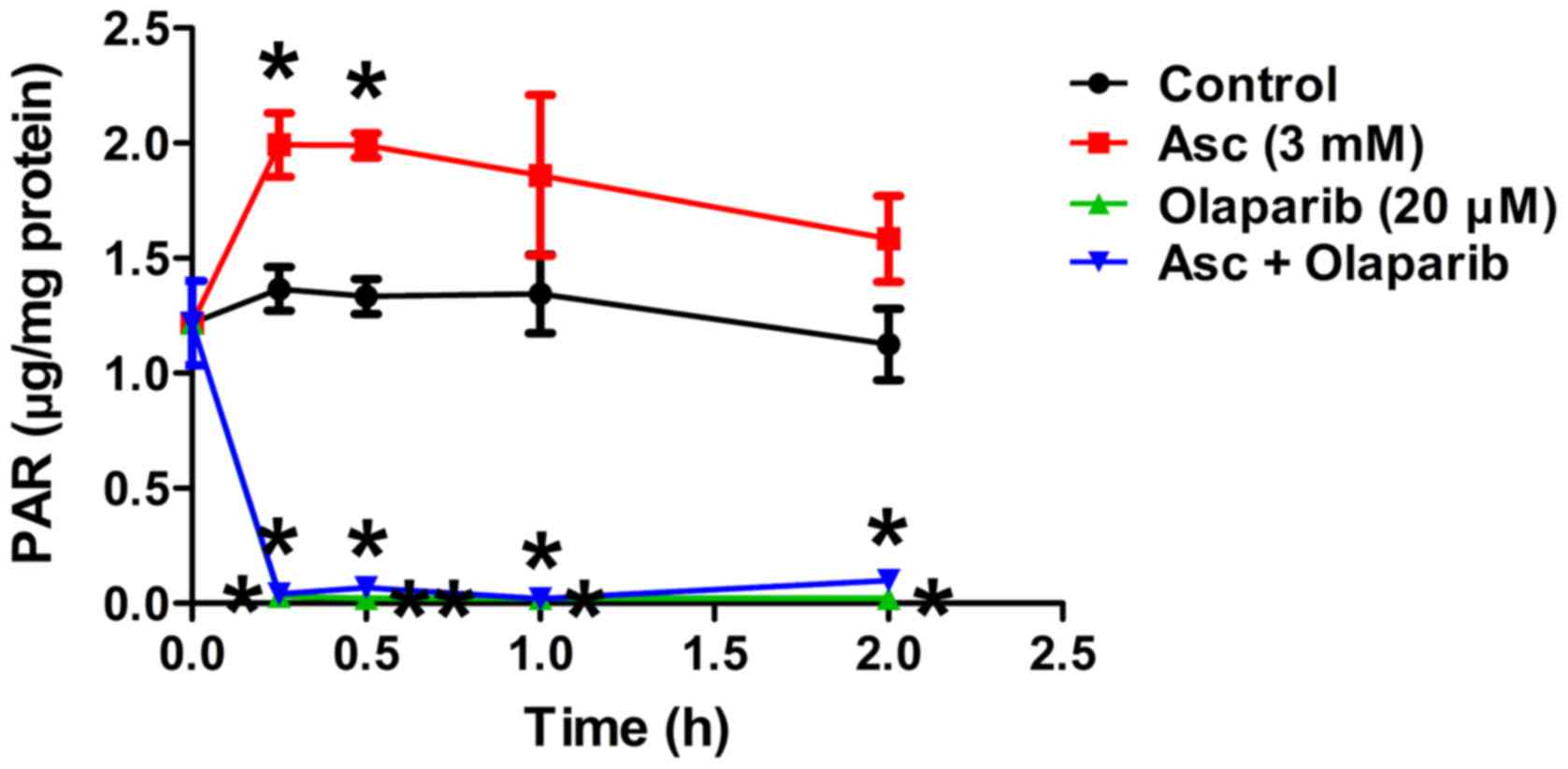
levels were significantly increased by pharmacological ascorbate in
SHIN3 cells compared with the control, as early as 15 min
(P<0.05; Fig. 3), suggesting that
PARP was activated as a cellular response to the
H2O2-induced DNA damage mediated by
ascorbate. Treatment with olaparib significantly decreased PAR
levels in the presence of ascorbate compared with the control
(P<0.05; Fig. 3), suggesting that
PARP-mediated DNA repair was inhibited. Taken together, these
results suggest that when PARP activity is inhibited,
pharmacological ascorbate significantly potentiates cell death,
potentially through enhanced DNA damage in BRCA1/2 wild-type EOC
cells.
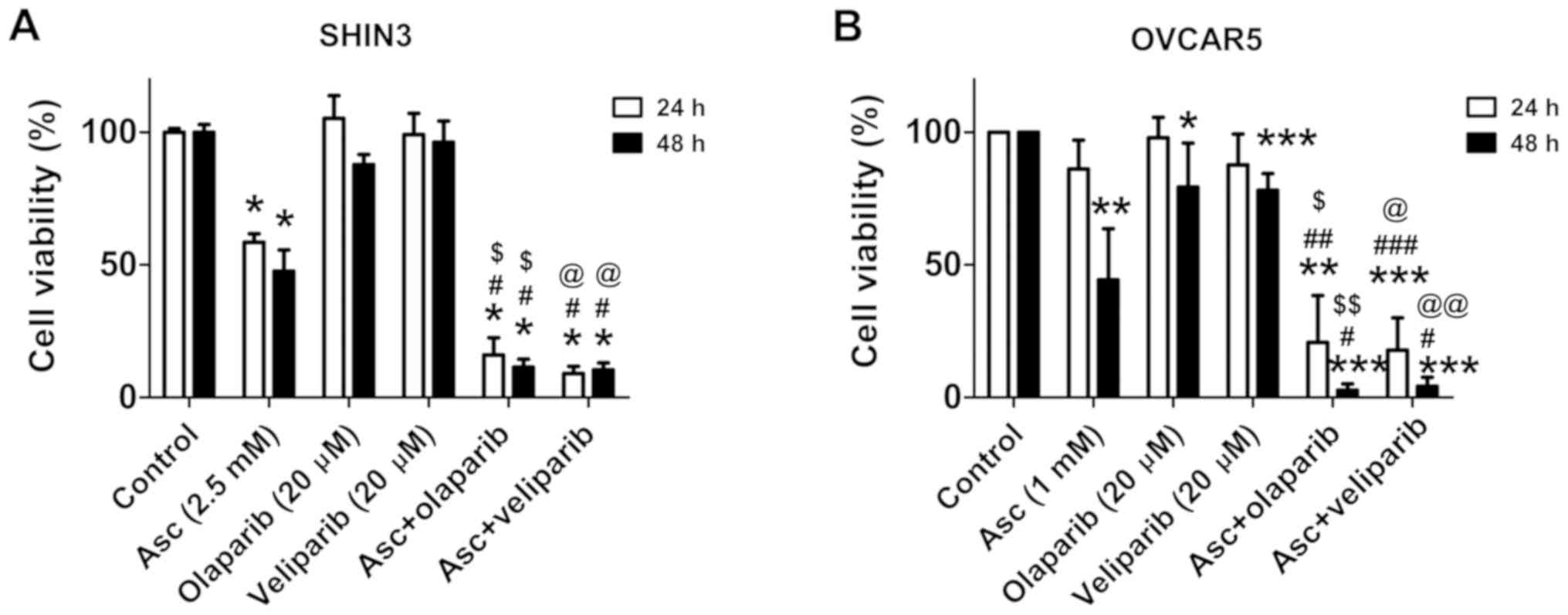 | Figure 2.Combining pharmacological ascorbate
and PARPis synergistically inhibits the viability of BRCA1/2
wild-type EOC cells. Cell viability of (A) SHIN3 (5 independent
experiments each performed in triplicate) or (B) OVCAR cells (4
independent experiments each performed in triplicate) following
treatment with vehicle, 2.5 mM ascorbate, 20 µM olaparib, 20 µM
veliparib, ascorbate + olaparib or ascorbate + veliparib for 24 and
48 h. Data are expressed as the mean ± standard deviation.
*P<0.05, **P<0.01, ***P<0.001 vs. control;
#P<0.05, ##P<0.01,
###P<0.001 vs. Asc; $P<0.05,
$$P<0.01 vs. olaparib; @P<0.05,
@@P<0.01 vs. veliparib. EOC, epithelial ovarian
cancer; PARPi, poly(ADP-ribose) polymerase inhibitor; Asc,
pharmacological ascorbate. |
Pharmacological ascorbate inhibits HR
repair of DNA DSBs in BRCA1/2 wild-type EOC cells
As pharmacological ascorbate induced
H2O2 and caused DNA damage in cancer cells
(20), the effects of
pharmacological ascorbate on DDR in BRCA1/2 wild-type EOC cells was
determined in the present study, with a focus on the HR and
non-homologous end joining (NHEJ) signaling pathways. Treatment
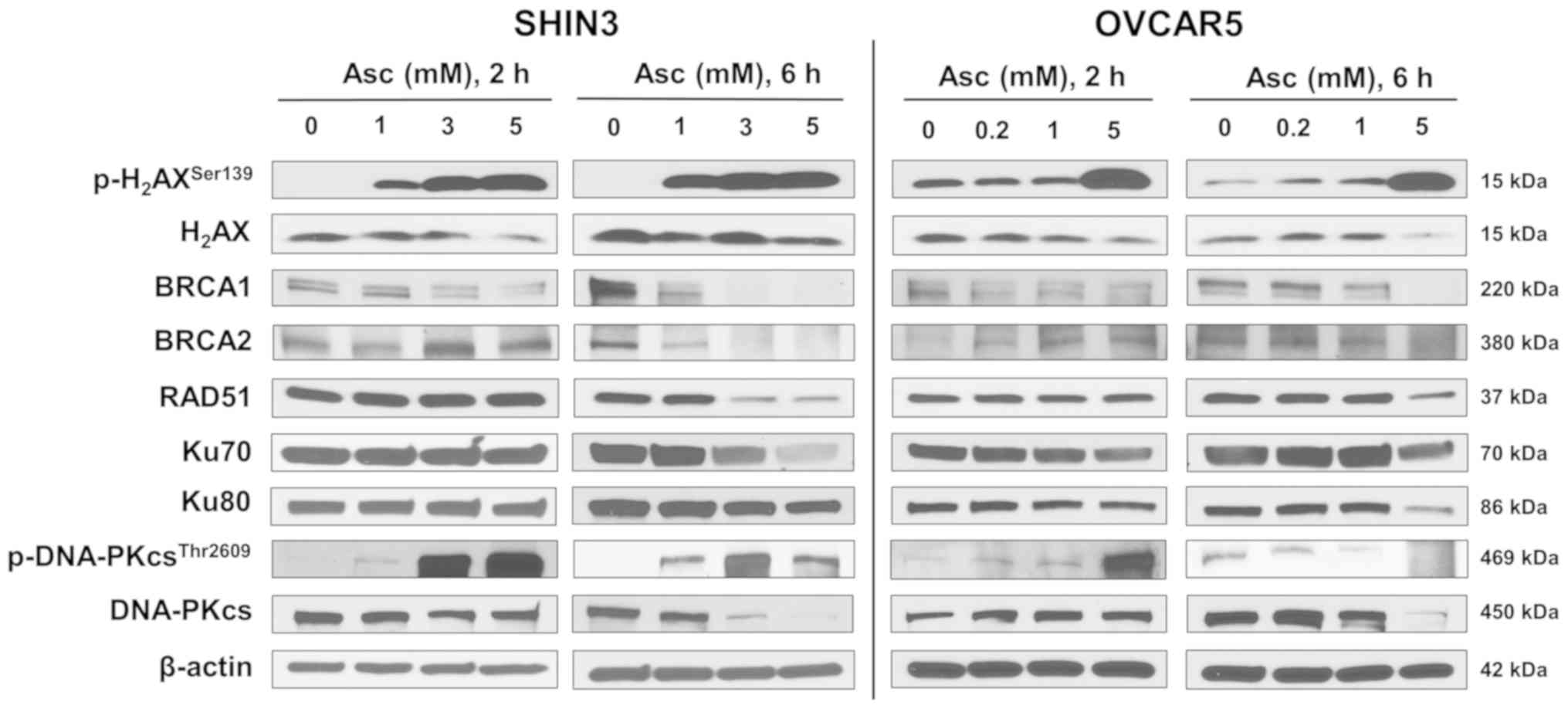
with pharmacological ascorbate for 2 and 6 h both notably increased
p-H2AXSer139 levels in both SHIN3 and OVCAR5
cells, a marker of DNA DSBs (Fig.
4). Expression of BRCA1 was downregulated in both tested cell
lines 2 h post-ascorbate treatment and further decreased 6 h post
treatment (Fig. 4). Notably, the
expression of BRCA2 and RAD51 was slightly increased in SHIN3 cells
2 h post treatment, followed by decreases 6 h post treatment
(Fig. 4), suggesting that
pharmacological ascorbate transiently activated HR repair machinery
as part of the stress response, and then inhibited the HR repair
machinery. Similar patterns of changes in the expression of BRCA2
and RAD51 were observed in OVCAR5 cells (Fig. 4). Together, these data suggest that
pharmacological ascorbate inhibited HR DNA DSBs repair pathway by
decreasing the expression of BRCA1, BRCA2 and RAD51 in BRCA1/2
wild-type EOC cells.
The NHEJ pathway was also investigated. The data
revealed that pharmacological ascorbate treatment minimally
affected the expression of Ku70 and Ku80 in SHIN3 cells within the
first 2 h. After 6 h of treatment, a dose-dependent decrease in the
expression of Ku70 and Ku80 was observed following ascorbate
treatment (Fig. 4). Ascorbate
treatment also decreased the expression of Ku70 and Ku80 in a
dose-dependent manner in OVCAR5 cells at either 2 or 6 h of
treatment (Fig. 4). Ascorbate
treatment increased the levels of p-DNA-PKcsThr2609, a
product of activated NHEJ, in SHIN3 and OVCAR5 cells after of 2 h
treatment as part of the stress responses. After 6 h,
p-DNA-PKcsThr2609 levels were decreased in OVCAR5 cells,
but not in SHIN3 cells (Fig. 4).
However, the expression of total DNA-PKcs was decreased by
ascorbate in a dose-dependent manner in SHIN3 cells after both 2
and 6 h treatment. In OVCAR5 cells DNA-PKcs was first upregulated
(2 h) and then downregulated (6 h) following ascorbate treatment
(Fig. 4). These data suggest that
the regulation of pharmacological ascorbate on the NHEJ DNA repair
proteins (Ku70, Ku80 and DNA-PKcs) was cell-line dependent with an
overall tendency of inhibition.
Combination of pharmacological
ascorbate and olaparib enhances DNA DSBs
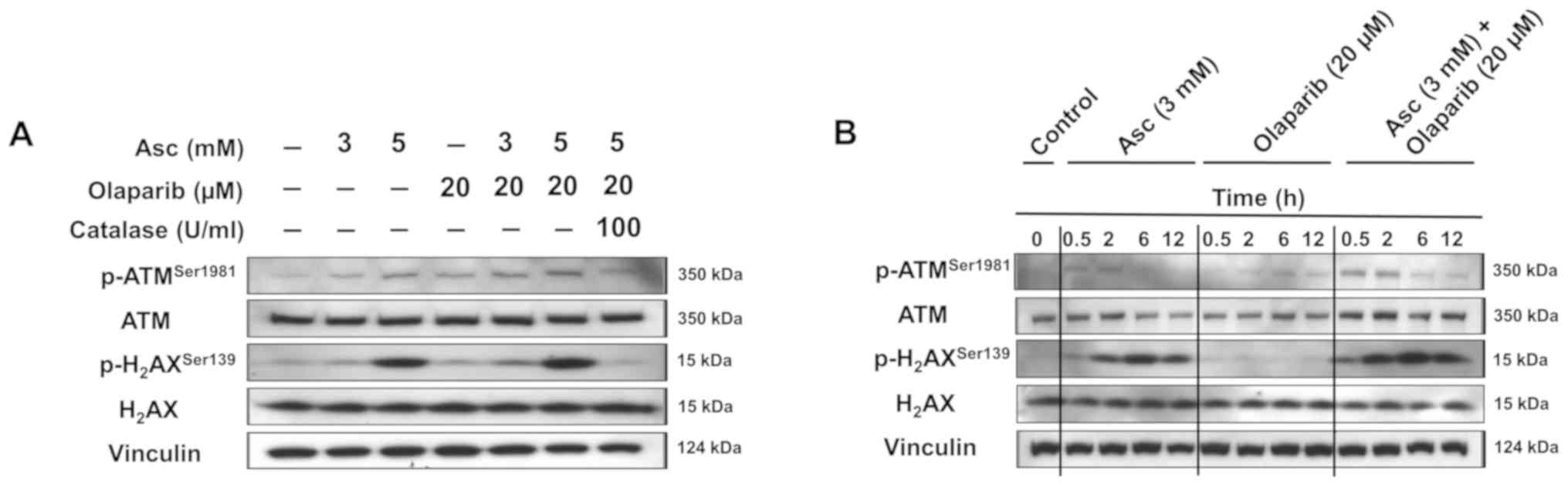
As presented in Fig.
5A, treatment with ascorbate alone or olaparib alone increased
the expression of p-ATMSer1981, an early marker of
oxidative DNA damage, and the expression of
p-H2AXSer139, a marker of DNA DSBs, in a
dose-dependent manner in SHIN3 cells treated for 2 h. The
combination treatment of ascorbate and olaparib further enhanced
the expression of p-ATMSer1981 and
p-H2AXSer139 compared with both drugs alone.
Treatment with catalase decreased the levels of
p-ATMSer1981 and p-H2AXSer139 when
it was added prior to treatment with olaparib and ascorbate,
suggesting that catalase protects cells from oxidative DNA damage.
Similar observations were also observed in OVCAR5 cells (Fig. S2).
In addition, the time-dependent expression levels of
p-ATMSer1981 and p-H2AXSer139 were
detected following incubation with 3 mM ascorbate in SHIN3 cells
(Fig. 5B). Consistently, the
combination treatment of ascorbate and olaparib further increased
the expression of p-ATMSer1981 and
p-H2AXSer139 compared with either single
agent treatment alone at the same time points (Fig. 5B).
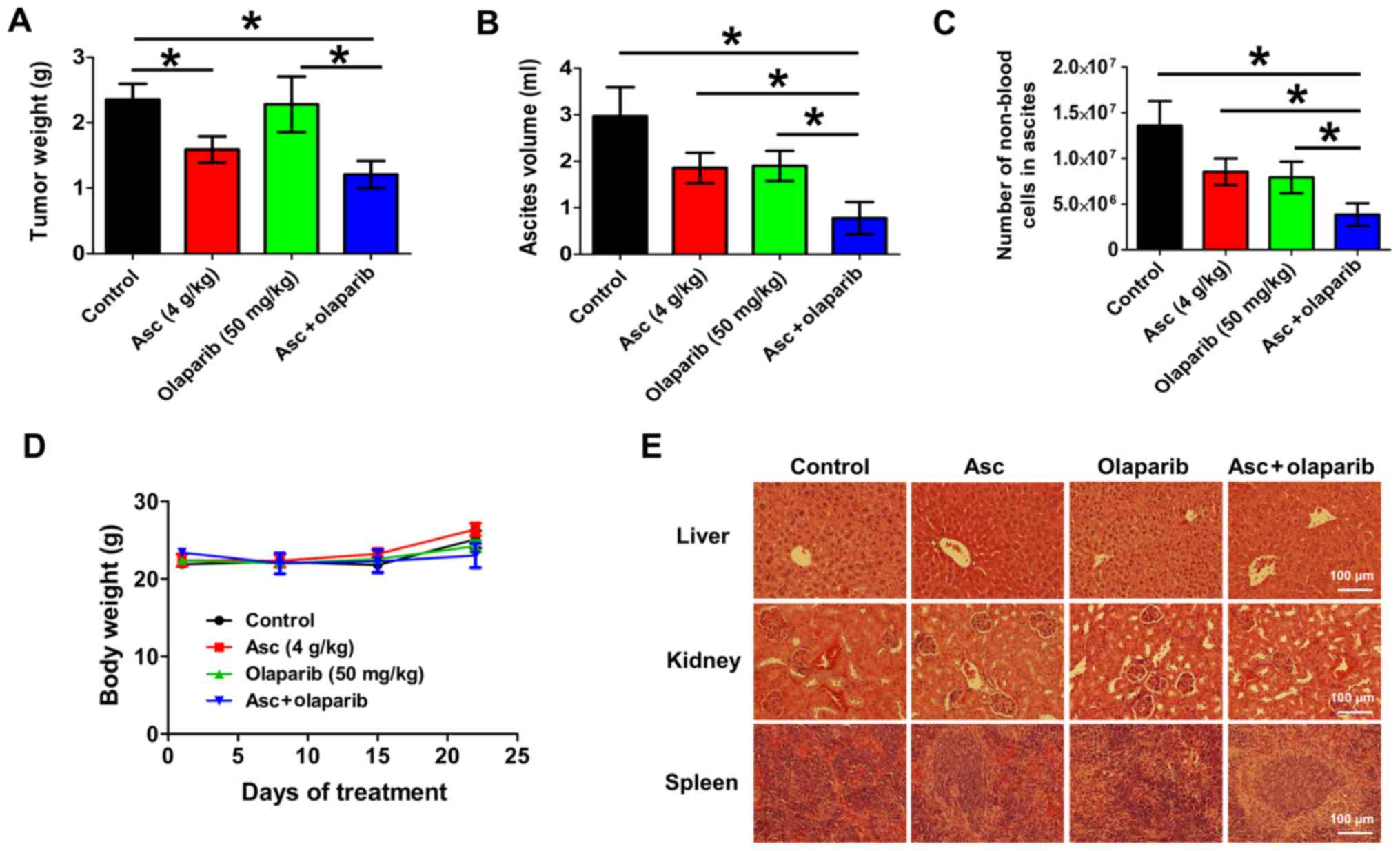
Combination treatment of
pharmacological ascorbate and olaparib significantly decreases
tumor and ascites burden in vivo
In order to assess the therapeutic efficacy of
combination pharmacological ascorbate and olaparib in vivo,
a validated xenograft mouse model of ovarian cancer was used
(20). This model mimics the later
stages of EOC with clinical observations of abdominal tumor
dissemination and ascites formation in patients. After 25 days of
treatment, ascorbate (4 g/kg) alone significantly decreased tumor
weight by 32.5% compared with the vehicle control (P<0.05;
Fig. 6A). Olaparib (50 mg/kg) alone
had minimal effects (P>0.05) on the tumor weight of these BRCA
wild-type xenografts (Fig. 6A). The
combination treatment of ascorbate and olaparib significantly
decreased tumor weight by 48.6% compared with the vehicle control
(P<0.05; Fig. 6A). Treatment with
ascorbate or olaparib alone did not significantly decrease the
ascites volumes or number of tumor cells in the ascites at the end
of treatment, the combination treatment resulted in a 73.8%
decrease in ascites volume (P<0.05; Fig. 6B) and a 71.7% decrease in the number
of tumor cells in the ascetic fluids (P<0.05; Fig. 6C) relative to the vehicle control.
The decrease was significant compared with either treatment alone.
All treatments were well tolerated and did not result in weight
loss (Fig. 6D). H&E staining
demonstrated no pathological changes in the livers, kidneys or
spleens of the animals in all treatment groups, suggesting that
ascorbate, olaparib and the combination treatment were of low
toxicity at the tested concentrations (Fig. 6E).
Discussion
BRCA1/2 mutations occur in <20% of patients with
EOC, and it is hypothesized that this mutation limits the clinical
efficacy of PARPis (15,16). However, PARPis are used in certain
patients with EOC regardless of BRCA status (36,37).
Currently, three PARPis are used as standard treatment for EOC, and
two of these, olaparib and rucaparib, are used for treatment of
recurrent BRCA mutant ovarian cancer, as well as maintenance
therapy in platinum-sensitive relapsed EOC regardless of BRCA or
HRD status (36,37). Niraparib, the third clinically used
PARPi, is indicated for maintenance, and used irrespective of BRCA
status (38). There is an unmet
clinical need to decrease the resistance and enhance the efficacy
of PARPis.
In order to expand the applicability and increase
the efficacy of PARPis, the addition of pharmacological ascorbate
to treatment with PARPis in BRCA wild-type EOC models was assessed
for several reasons: i) A previous study demonstrated that
pharmacological ascorbate decreased the toxicities of conventional
chemotherapies (20); ii)
pharmacological ascorbate produces peroxide and damages DNA, thus
could work synergistically with an inhibitor of the DNA repair
machinery, as presented in the present study; and iii) ascorbate
influenced the homologous recombination pathway, and thus could
decrease resistance to PARPis, also demonstrated in the present
study. Therefore, the present study highlights the potential of
combining pharmacological ascorbate and PARPis and how it may
benefit a broader population of patients with EOC, even with
wild-type BRCA. A clinical proof-of-concept study is required as
the next step to further investigate this potential.
The present study demonstrates the preclinical
efficacy and feasibility of combining pharmacological ascorbate and
PARPis for treating BRCA wild-type ovarian cancer. In vitro,
the combination synergistically induced the death of BRCA wild-type
EOC cells; in vivo, the combination treatment significantly
decreased tumor and ascites burdens without inducing any toxicity.
The present study demonstrates that pharmacological ascorbate
potentiates the therapeutic efficacy of olaparib in BRCA wild-type
EOC by inducing HR deficiency or a ‘BRCAness’ phenotype. Therefore,
the combination used in the present study is a novel and
potentially promising therapeutic option for treating patients with
EOC, particularly those who do not respond to PARPis alone. Such a
strategy could be applied to a variety of heterogeneous and
hard-to-treat malignancies, including breast, pancreatic and
prostate cancer, where BRCA1, BRCA2 or other HR repair proteins are
instrumental in the repair of DNA DSBs and the potential of PARPis
has not yet been fully exploited (39).
Consistent with previous studies (22,29,40), the
data in the present study demonstrated that treatment with
pharmacological ascorbate resulted in the production of
H2O2, which damages DNA, leading to PARP
activation, and this was impaired by the PARPis. The oxidative
stress induced by pharmacological ascorbate caused excessive DNA
DSBs in BRCA1/2 wild-type EOC cells within the first 6 h of
treatment. The concentration and time ranges are clinically
relevant to those of i.v. ascorbate infusion (26). Pharmacological ascorbate is
selectively lethal to cancer cells, but not normal cells, which is
partially attributed to the increased intracellular levels of
reactive oxygen species (ROS) and the decreased ability to
metabolize H2O2 in cancer cells compared with
normal cells (41,42). The ROS-induction mechanism of high
dose ascorbate provides the first rationale for combining ascorbate
with a PARPi.
Ascorbate inhibited DNA repair enzymes, which
provides another rationale for combined treatment with PARPi. HR
and NHEJ are the two primary DNA DSBs repair pathways in eukaryotic
cells (43). HR is a
template-directed DNA repair with high-fidelity, which is crucial
for the maintenance of both telomere integrity and genomic
stability; whereas NHEJ is an error-prone DNA repair process, which
does not use a complementary template and can introduce deleterious
mutations during repair (43). The
results of the present study suggest that pharmacological ascorbate
suppressed the expression of HR repair proteins BRCA1, BRCA2 and
RAD51, leading to HR deficiency. In addition, ascorbate influences
the NHEJ pathway, impeding both HR and NHEJ pathways. Patel et
al (44) and Do et al
(45) reported that with PARP
inhibition, HR-deficient cancer cells upregulated the NHEJ as an
alternative DNA repair pathway. As pharmacological ascorbate
impedes both HR and NHEJ, adding it to a PARPi can further promote
genomic instability and enhance cytotoxicity.
Patients with EOC with wild-type BRCA are
considerably less responsive to PARPis compared with carriers of
germline or somatic BRCA mutations (13,17,46).
Consistent with the clinical observations, the present study
demonstrated that BRCA wild-type EOC cells are not sensitive to
olaparib, both in vitro and in vivo. Addition of
pharmacological ascorbate to olaparib overcomes the resistance to
olaparib and significantly decreased tumor burden. These results
highlight the potential of a novel clinical solution for patients
with EOC who do not benefit from PARPis alone.
It is well-documented that high-dose IVC is well
tolerated with minimal toxicity in humans (26–28,47). A
previous Phase I/IIa clinical trials in patients with EOC (20) and pancreatic cancer (40), together with other trials (25,27,28),
have consistently demonstrated that adding IVC to standard
treatments (chemotherapy or radiation therapy) is safe, well
tolerated, feasible and potentially effective. Addition of IVC to
carboplatin and paclitaxel chemotherapy substantially decreased
side-effects in patients with stage III or IV EOC (20). Potential survival benefits of IVC
were reported in combination with chemotherapy or
radio-chemotherapy in the treatment of pancreatic cancer,
glioblastoma multiforme and advanced-stage non-small cell lung
cancer (25,28). Several randomized Phase II trials
evaluating the efficacy of IVC are underway (48).
A limitation of the present study lies in the cell
line-derived xenograft, which may not be representative of the
heterogeneous nature of tumors in patients. Due to the
idiosyncratic characteristics of different tumors from different
patients, the present study now warrant testing this novel drug
combination in multiple patient-derived xenograft mouse models,
which will provide more relevant data for future clinical trials.
Nevertheless, the results of the present study highlight new
opportunities for the translational studies of pharmacological
ascorbate in combination with PARPis for treating patients with
EOC, regardless of BRCA or HRD status.
In conclusion, the present study demonstrated that
the combination treatment of pharmacological ascorbate with PARPis
had the potential to provide therapeutic benefits to patients with
ovarian cancer who do not respond to PARPis alone. The advantages
of this combination therapy lie in the potentially broad
applicability, improved efficacy and low toxicity.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Dr. Peter Eck
(University of Manitoba; Manitoba, Canada) for providing the SHIN3
cell line, Ms. Min Yang (University of Kansas Medical Center;
Kansas City, USA) for helping to generate part of the western
blotting data, Dr. Kishore Polireddy and Dr. Thuy-Vy Do (University
of Kansas Medical Center; Kansas City, USA) for assisting with
exploratory experiments, Dr. Chunhua Li and Dr. Nan He (University
of Kansas Medical Center; Kansas City, USA) for DNA sequencing and
sequence analysis, and the staff of the Clinical Molecular Oncology
Laboratory of University of Kansas Medical Center for cell line
authentication.
Funding
The present study was financially supported by a
bridging grant from the University of Kansas Research Institute
(Kansas, USA), and a grant from the University of Kansas Endowment
provided by the GR's Foundation, Mosby Lincoln Foundation, and
Donlan Foundation (Kansas, USA).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
QC, AKG, JAD and YM conceived and designed the
study. YM, PC and DK developed the methodology, performed the
experiments and collected data. YM, PC, QC and DK analyzed,
computed and interpreted the data. YM wrote the manuscript. YM, QC,
AKG, JAD, PC and DK reviewed and revised the manuscript. JAD, AKG
and QC provided administrative, technical and material support. QC
supervised the study.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Trabert B, DeSantis CE, Miller
KD, Samimi G, Runowicz CD, Gaudet MM, Jemal A and Siegel RL:
Ovarian cancer statistics, 2018. CA Cancer J Clin. 68:284–296.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
https://seer.cancer.gov/statfacts/html/ovary.html
|
|
4
|
Taylor KN and Eskander RN: PARP inhibitors
in epithelial ovarian cancer. Recent Pat Anticancer Drug Discov.
13:145–158. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Stecklein SR, Kumaraswamy E, Behbod F,
Wang W, Chaguturu V, Harlan-Williams LM and Jensen RA: BRCA1 and
HSP90 cooperate in homologous and non-homologous DNA
double-strand-break repair and G2/M checkpoint activation. Proc
Natl Acad Sci USA. 109:13650–13655. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bryant HE and Helleday T: Inhibition of
poly (ADP-ribose) polymerase activates ATM which is required for
subsequent homologous recombination repair. Nucleic Acids Res.
34:1685–1691. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
McCabe N, Turner NC, Lord CJ, Kluzek K,
Bialkowska A, Swift S, Giavara S, O'Connor MJ, Tutt AN, Zdzienicka
MZ, et al: Deficiency in the repair of DNA damage by homologous
recombination and sensitivity to poly(ADP-ribose) polymerase
inhibition. Cancer Res. 66:8109–8115. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bryant HE, Schultz N, Thomas HD, Parker
KM, Flower D, Lopez E, Kyle S, Meuth M, Curtin NJ and Helleday T:
Specific killing of BRCA2-deficient tumours with inhibitors of
poly(ADP-ribose) polymerase. Nature. 434:913–917. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Farmer H, McCabe N, Lord CJ, Tutt AN,
Johnson DA, Richardson TB, Santarosa M, Dillon KJ, Hickson I,
Knights C, et al: Targeting the DNA repair defect in BRCA mutant
cells as a therapeutic strategy. Nature. 434:917–921. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kim G, Ison G, McKee AE, Zhang H, Tang S,
Gwise T, Sridhara R, Lee E, Tzou A, Philip R, et al: FDA approval
summary: Olaparib monotherapy in patients with deleterious germline
BRCA-mutated advanced ovarian cancer treated with three or more
lines of chemotherapy. Clin Cancer Res. 21:4257–4261. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pujade-Lauraine E, Ledermann JA, Selle F,
Gebski V, Penson RT, Oza AM, Korach J, Huzarski T, Poveda A,
Pignata S, et al: Olaparib tablets as maintenance therapy in
patients with platinum-sensitive, relapsed ovarian cancer and a
BRCA1/2 mutation (SOLO2/ENGOT-Ov21): A double-blind, randomised,
placebo-controlled, phase 3 trial. Lancet Oncol. 18:1274–1284.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Coleman RL, Oza AM, Lorusso D, Aghajanian
C, Oaknin A, Dean A, Colombo N, Weberpals JI, Clamp A, Scambia G,
et al: Rucaparib maintenance treatment for recurrent ovarian
carcinoma after response to platinum therapy (ARIEL3): A
randomised, double-blind, placebo-controlled, phase 3 trial.
Lancet. 390:1949–1961. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mirza MR, Monk BJ, Herrstedt J, Oza AM,
Mahner S, Redondo A, Fabbro M, Ledermann JA, Lorusso D, Vergote I,
et al: Niraparib maintenance therapy in platinum-sensitive,
recurrent ovarian cancer. N Engl J Med. 375:2154–2164. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Moore K, Colombo N, Scambia G, Kim BG,
Oaknin A, Friedlander M, Lisyanskaya A, Floquet A, Leary A, Sonke
GS, et al: Maintenance olaparib in patients with newly diagnosed
advanced ovarian cancer. N Engl J Med. 379:2495–2505. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nielsen FC, van Overeem Hansen T and
Sorensen CS: Hereditary breast and ovarian cancer: New genes in
confined pathways. Nat Rev Cancer. 16:599–612. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Walsh T, Casadei S, Lee MK, Pennil CC,
Nord AS, Thornton AM, Roeb W, Agnew KJ, Stray SM, Wickramanayake A,
et al: Mutations in 12 genes for inherited ovarian, fallopian tube,
and peritoneal carcinoma identified by massively parallel
sequencing. Proc Natl Acad Sci USA. 108:18032–18037. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Swisher EM, Lin KK, Oza AM, Scott CL,
Giordano H, Sun J, Konecny GE, Coleman RL, Tinker AV, O'Malley DM,
et al: Rucaparib in relapsed, platinum-sensitive high-grade ovarian
carcinoma (ARIEL2 Part 1): An international, multicentre,
open-label, phase 2 trial. Lancet Oncol. 18:75–87. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Loibl S, O'Shaughnessy J, Untch M, Sikov
WM, Rugo HS, McKee MD, Huober J, Golshan M, von Minckwitz G, Maag
D, et al: Addition of the PARP inhibitor veliparib plus carboplatin
or carboplatin alone to standard neoadjuvant chemotherapy in
triple-negative breast cancer (BrighTNess): A randomised, phase 3
trial. Lancet Oncol. 19:497–509. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bang YJ, Xu RH, Chin K, Lee KW, Park SH,
Rha SY, Shen L, Qin S, Xu N, Im SA, et al: Olaparib in combination
with paclitaxel in patients with advanced gastric cancer who have
progressed following first-line therapy (GOLD): A double-blind,
randomised, placebo-controlled, phase 3 trial. Lancet Oncol.
18:1637–1651. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ma Y, Chapman J, Levine M, Polireddy K,
Drisko J and Chen Q: High-dose parenteral ascorbate enhanced
chemosensitivity of ovarian cancer and reduced toxicity of
chemotherapy. Sci Transl Med. 6:222ra182014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen Q, Espey MG, Krishna MC, Mitchell JB,
Corpe CP, Buettner GR, Shacter E and Levine M: Pharmacologic
ascorbic acid concentrations selectively kill cancer cells: Action
as a pro-drug to deliver hydrogen peroxide to tissues. Proc Natl
Acad Sci USA. 102:13604–13609. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen Q, Espey MG, Sun AY, Lee JH, Krishna
MC, Shacter E, Choyke PL, Pooput C, Kirk KL, Buettner GR and Levine
M: Ascorbate in pharmacologic concentrations selectively generates
ascorbate radical and hydrogen peroxide in extracellular fluid in
vivo. Proc Natl Acad Sci USA. 104:8749–8754. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen Q, Espey MG, Sun AY, Pooput C, Kirk
KL, Krishna MC, Khosh DB, Drisko J and Levine M: Pharmacologic
doses of ascorbate act as a prooxidant and decrease growth of
aggressive tumor xenografts in mice. Proc Natl Acad Sci USA.
105:11105–11109. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Du J, Martin SM, Levine M, Wagner BA,
Buettner GR, Wang SH, Taghiyev AF, Du C, Knudson CM and Cullen JJ:
Mechanisms of ascorbate-induced cytotoxicity in pancreatic cancer.
Clin Cancer Res. 16:509–520. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Schoenfeld JD, Sibenaller ZA, Mapuskar KA,
Wagner BA, Cramer-Morales KL, Furqan M, Sandhu S, Carlisle TL,
Smith MC, Abu Hejleh T, et al: O2− and
H2O2-mediated disruption of fe metabolism
causes the differential susceptibility of NSCLC and GBM cancer
cells to pharmacological ascorbate. Cancer Cell. 32:2682017.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hoffer LJ, Levine M, Assouline S,
Melnychuk D, Padayatty SJ, Rosadiuk K, Rousseau C, Robitaille L and
Miller WH Jr: Phase I clinical trial of i.v. ascorbic acid in
advanced malignancy. Ann Oncol. 19:1969–1974. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Monti DA, Mitchell E, Bazzan AJ, Littman
S, Zabrecky G, Yeo CJ, Pillai MV, Newberg AB, Deshmukh S and Levine
M: Phase I evaluation of intravenous ascorbic acid in combination
with gemcitabine and erlotinib in patients with metastatic
pancreatic cancer. PLoS One. 7:e297942012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Welsh JL, Wagner BA, van't Erve TJ, Zehr
PS, Berg DJ, Halfdanarson TR, Yee NS, Bodeker KL, Du J, Roberts LJ
II, et al: Pharmacological ascorbate with gemcitabine for the
control of metastatic and node-positive pancreatic cancer (PACMAN):
Results from a phase I clinical trial. Cancer Chemother Pharmacol.
71:765–775. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ma E, Chen P, Wilkins HM, Wang T, Swerdlow
RH and Chen Q: Pharmacologic ascorbate induces neuroblastoma cell
death by hydrogen peroxide mediated DNA damage and reduction in
cancer cell glycolysis. Free Radic Biol Med. 113:36–47. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Stordal B, Timms K, Farrelly A, Gallagher
D, Busschots S, Renaud M, Thery J, Williams D, Potter J, Tran T, et
al: BRCA1/2 mutation analysis in 41 ovarian cell lines reveals only
one functionally deleterious BRCA1 mutation. Mol Oncol. 7:567–579.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
van de Wetering M, Barker N, Harkes IC,
van der Heyden M, Dijk NJ, Hollestelle A, Klijn JG, Clevers H and
Schutte M: Mutant E-cadherin breast cancer cells do not display
constitutive Wnt signaling. Cancer Res. 61:278–284. 2001.PubMed/NCBI
|
|
32
|
Hall TA: BioEdit: A user-friendly
biological sequence alignment editor and analysis program for
windows 95/98/NT. Nucleic Acids Symp Ser. 41:95–98. 1999.
|
|
33
|
Feldman AT and Wolfe D: Tissue processing
and hematoxylin and eosin staining. Methods Mol Biol. 1180:31–43.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Imai S, Kiyozuka Y, Maeda H, Noda T and
Hosick HL: Establishment and characterization of a human ovarian
serous cystadenocarcinoma cell line that produces the tumor markers
CA-125 and tissue polypeptide antigen. Oncology. 47:177–184. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Vaidyanathan A, Sawers L, Gannon AL,
Chakravarty P, Scott AL, Bray SE, Ferguson MJ and Smith G: ABCB1
(MDR1) induction defines a common resistance mechanism in
paclitaxel- and olaparib-resistant ovarian cancer cells. Br J
Cancer. 115:431–441. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Food and Drug Administration, . FDA
approves olaparib tablets for maintenance treatment in ovarian
cancer. https://www.fda.gov/drugs/resources-information-approved-drugs/fda-approves-olaparib-tablets-maintenance-treatment-ovarian-cancerMay
15–2018
|
|
37
|
Food and Drug Administration, . FDA
approves rucaparib for maintenance treatment of recurrent ovarian,
fallopian tube, or primary peritoneal cancer. https://www.fda.gov/drugs/resources-information-approved-drugs/fda-approves-rucaparib-maintenance-treatment-recurrent-ovarian-fallopian-tube-or-primary-peritonealMay
15–2018
|
|
38
|
Ison G, Howie LJ, Amiri-Kordestani L,
Zhang L, Tang S, Sridhara R, Pierre V, Charlab R, Ramamoorthy A,
Song P, et al: FDA approval summary: Niraparib for the maintenance
treatment of patients with recurrent ovarian cancer in response to
platinum-based chemotherapy. Clin Cancer Res. 24:4066–4071. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kamel D, Gray C, Walia JS and Kumar V:
PARP inhibitor drugs in the treatment of breast, ovarian, prostate
and pancreatic cancers: An update of clinical trials. Curr Drug
Targets. 19:21–37. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Polireddy K, Dong R, Reed G, Yu J, Chen P,
Williamson S, Violet PC, Pessetto Z, Godwin AK, Fan F, et al: High
dose parenteral ascorbate inhibited pancreatic cancer growth and
metastasis: Mechanisms and a phase I/IIa study. Sci Rep.
7:171882017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Panieri E and Santoro MM: ROS homeostasis
and metabolism: A dangerous liason in cancer cells. Cell Death Dis.
7:e22532016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Doskey CM, Buranasudja V, Wagner BA,
Wilkes JG, Du J, Cullen JJ and Buettner GR: Tumor cells have
decreased ability to metabolize H2O2:
Implications for pharmacological ascorbate in cancer therapy. Redox
Biol. 10:274–284. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Shibata A: Regulation of repair pathway
choice at two-ended DNA double-strand breaks. Mutat Res.
803-805:51–55. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Patel AG, Sarkaria JN and Kaufmann SH:
Nonhomologous end joining drives poly(ADP-ribose) polymerase (PARP)
inhibitor lethality in homologous recombination-deficient cells.
Proc Natl Acad Sci USA. 108:3406–3411. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Do TV, Hirst J, Hyter S, Roby KF and
Godwin AK: Aurora A kinase regulates non-homologous end-joining and
poly(ADP-ribose) polymerase function in ovarian carcinoma cells.
Oncotarget. 8:50376–50392. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ledermann J, Harter P, Gourley C,
Friedlander M, Vergote I, Rustin G, Scott CL, Meier W,
Shapira-Frommer R, Safra T, et al: Olaparib maintenance therapy in
patients with platinum-sensitive relapsed serous ovarian cancer: A
preplanned retrospective analysis of outcomes by BRCA status in a
randomised phase 2 trial. Lancet Oncol. 15:852–861. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Padayatty SJ, Sun AY, Chen Q, Espey MG,
Drisko J and Levine M: Vitamin C: Intravenous use by complementary
and alternative medicine practitioners and adverse effects. PLoS
One. 5:e114142010. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ngo B, Van Riper JM, Cantley LC and Yun J:
Targeting cancer vulnerabilities with high-dose vitamin C. Nat Rev
Cancer. 19:271–282. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
BRCA1 BRCA1 DNA repair associated [Homo
sapiens (human)]. Gene ID: 672. https://www.ncbi.nlm.nih.gov/gene/?term=NM_007294Updated.
November 25–2019.
|
|
50
|
BRCA2 BRCA2 DNA repair associated [Homo
sapiens (human)]. Gene ID: 675. https://www.ncbi.nlm.nih.gov/gene/?term=NM_000059Updated.
November 18–2019.
|