Introduction
B-cell acute lymphoplastic leukemia (B-ALL) is a
heterogeneous disease, however from the genetic point of view it is
the result of block differentiation and clonal proliferation of
lymphoid B-cell progenitors in bone marrow, blood and
extramedullary sites. B-ALL is the most common cancer in children,
represents 80% of ALL cases and is the leading cause of
cancer-related death in children (1,2).
A key role for B-ALL pathogenesis has been
attributed to recurrent cytogenetic and molecular abnormalities
which can be detected in ~60% of ALL cases. A diagnostic, treatment
and prognostic significance is known for example for translocation
t(12;21)(p13;q22) leading to the ETV6/RUNX1 gene-fusion,
carriers of which are more likely to be cured than those with a
translocation BCR/ABL1 or KMT2A/AFF1. Besides,
high-hyperdiploidy (51–67 chromosomes), encompassing 25% of
childhood B-ALL, has been associated with good survival and
favorable outcome. Controversely, hypodiploidy (<44 chromosomes)
has been linked to unfavorable prognosis (2,3). Complex
karyotypes (CKs) are rare events in children with B-ALL and
typically involve three to five or even more chromosomal
abnormalities and usually result in activation of oncogenes or
deletion of tumor suppression genes. CKs are found in ~5% of ALL
cases and are also associated with an adverse outcome (2,4).
Accordingly it is not surprising that copy number
alterations (CNAs) play a fundamental role in ALL development and
progression, being present as deletion or duplication of genes
involved in lymphoid development, cell cycle regulation and
apoptosis, such as EBF1, PAX5, IKZF1 and VPREB1. Such
genetic markers were already integrated into risk stratification
systems, after their validation in large clinical cohorts (5–10).
Consequently, B-ALL is one of the genetically best
characterized malignancies and the success of curing pediatric ALLs
has increased dramatically reaching ~85%. Nonetheless, cryptic
structural chromosomal abnormalities were and are still a challenge
in cytogenetic diagnostics of B-ALL (2,10).
In the present study, a comprehensive analysis of a
childhood B-ALL case was done using high resolution molecular
approaches, revealing a cryptic complex karyotype with involvement
of FUS, NR3C1 and VPREB1 genes.
Case report
Clinical description
A 4-year-old female was admitted to the Institute of
Mother and Child of Serbia ‘Dr. Vukan Cupic’ because of bruisings
and splenomegaly. At admission, the white blood cell (WBC) count
was 229×109/l with of 92% lymphocytes, the hemoglobin was 103 g/l,
the platelets were 52×109/l and red blood cells (RBC) count was
3.78×106/mm3. Serum uric acid (UA) value was
529 µmol/l (normal 150–450), and lactate-dehydrogenase (LDH) value
was 6,579 IU/l (normal 120–250).
Bone marrow aspirate showed infiltration with more
than 90% of lymphoblasts, FAB L1 60% and L2 30%. Periodic
acid-Schiff stain (PAS) and Sudan Black B were negative. Flow
cytometric (FCM) analysis characterized the expression of a variety
of B-cell-specific antigens and were positive for CD10 89%, CD19
93%, CD45 74%, iCD79a 94%, iIgM 68%, sIgλ 18% and were negative for
CD34, iCD3, sCD3, CD7, CD8, CD13, CD15, sIgκ and MPO. These
findings were consistent with B-ALL.
Unfortunately, during induction therapy (ALL IC-BFM
2009), a diagnosis of sepsis was established with capillary leak
syndrome and cardiopulmonary failure, resulting in a fatal outcome.
So, it was not possible to perform a control bone marrow aspiration
and determine whether a remission was achieved or not.
Chromosome analysis
Banding cytogenetic analyses was performed on
unstimulated bone marrow aspirate according to standard procedures
(11). A total of 20 metaphases were
available for cytogenetic evaluation and analyzed on a banding
level of 200 bands per haploid karyotype (12).
Molecular genetics
Total RNA was extracted from bone marrow cells,
using phenol chloroform extraction protocol (13). cDNA was prepared from 1 µg of total
RNA with the High Capacity Reverse Transcription Kit (Applied
Biosystems; Thermo Fisher Scientific, Inc.). Screening for
BCR/ABL1 (p190, p210), MLL/AF4,
ETV6/RUNX1 and PBX1/E2A fusion transcripts
were performed according to BIOMED-1 Concerted Action program
(BMH-CMT 94–1675), using Maxima Hot Start Taq DNA polymerase
(Thermo Fisher Scientific, Inc.) (14).
Molecular cytogenetics
Fluorescence in situ hybridization (FISH) was
done according to standard procedures and/or according to
manufacturer's instructions. Homemade probes and probe sets were
applied as follows: i) BAC (bacterial artificial chromosome) clones
of interest were identified through the Human Genome Browser
Database of the Genome Bioinformatics Group at the University of
California at Santa Cruz (http://genome.ucsc.edu/) and Ensembl Genome Data
Resources of the Sanger Institute Genome Database (http://www.ensembl.org/). DNA probes (Table I) obtained from Resources Center were
labeled by PCR with SpectrumGreen, SpectrumOrange or TexasRed-dUTP
and applied in two- or three-color FISH-approaches. ii) Chromosome
specific high resolution array-proven multicolor-banding (aMCB)
probes sets for #5, #9, #16, #19 and X were used. In combination
with whole chromosome painting (WCP) for chromosomes 5, 6, 9, 16,
17, 18, 19, 20, 21, 22, X were applied as well (15,16).
 | Table I.Locus specific probes used for
FISH. |
Table I.
Locus specific probes used for
FISH.
| Probe | Cytoband | Position
[NCBI36/hg18] | Genes/locus | Result (signals
on…) |
|---|
| RP11-434D11 | 5q23.2 |
chr5:126,045,879-126,232,850 | n.d. | der(5) |
| RP11-114H7 | 5q23.3 |
chr5:130,306,745-130,460,728 | n.d. | der(5) |
| LSI EGR1/D5S721,
D5S23 (Vysis) | 5q31.2 |
chr5:137,829,080-137,832,903 | EGR1 | der(5) |
| LSI PDGFRB DCBAP
(Vysis) | 5q33.1 |
chr5:149,473,595-149,515,615 | PDGFRB | der(9) |
| 5qTEL (Vysis) | 5q35.3 |
chr5:180,510,748-180,711,420 | D5S2907 | der(9) |
| 9pTEL (Vysis) | 9p24.3 |
chr9:131,486-331,767 |
305J7-T7 | der(5) |
| POSEIDON™ MLL/MLLT3
DCDFP (Kreatech) | 9p21.3 |
chr9:20,331,667-20,612,514 | MLLT3 | der(5) |
| SPEC CDKN2A/CEN9
DCP (Zytovision) | 9p21.3 |
chr9:21,957,751-21,965,038 |
CDKN2A/B | biallelic deletion
(9)(p21.3p21.3) |
| LSI BCR/ABL1 DCDFP
(Vysis) | 9q34.12 |
chr9:132,579,089-132,752,883 | ABL1 | der(9) |
| 9qTEL (Vysis) | 9q34.3 |
chr9:140,000,171-140,200,260 | D9S325 | der(9) |
| SPEC BIRC3/MALT1
DCDFP (Zytovision) | 11q22.3 |
chr11:101,693,404-101,713,675 | BIRC3 | #11 |
| RP11-142A12 | 16p12.1 |
chr16:26,595,070-26,757,869 | n.d | der(16) |
| RP11-147I4 | 16p12.1 |
chr16:27,071,343-27,211,332 | KDM8,
NSMCE1 | der(16) |
| RP11-159J3 | 16p11.2 |
chr16:28,012,684-28,171,002 | XPO6 | der(16) |
| SPEC FUS DCBAP
(Zytovision) | 16p11.2 |
chr16:31,098,954-31,110,600 | FUS | signal split on
der(16) and der(19) |
| SPEC TP53/ATM DCP
(Zytovision) | 17p13.1 |
chr17:7,512,445-7,531,588 | TP53 | #17 |
| RP11-21J15 | 19q13.31 |
chr19:49,726,602-49,900,222 | CEACAM22P,
IGSF23, PVR, CEACAM19 | der(19) |
| RP11-84C16 | 19q13.31 |
chr19:49,089,546-50,686,777 | BCL3 | der(19) |
| RP11-492P7 | 19q13.32 |
chr19:51,987,986-52,169,539 | AP2S,
ARHGAP35 | der(19) |
| SPEC 19q13/19p13
DCP (Zytovision) | 19q13.32 |
chr19:52,477,388-53,038,398 | GLTSCR1,
GLTSCR2, CRX | der(19) |
| RP11-3N16 | 19q13.32 |
chr19:53,269,619-53,431,708 | PLA2G4C, LIG1,
CARD8 | der(19) |
| RP11-264M8 | 19q13.33 |
chr19:54,767,615-54,925,355 | PRRG2, PRRG2,
IRF3, BCL2L12, PRMT1, CPTC1 | der(16) |
Additionally, commercially available probes were
applied: ZytoLight®SPEC CDKN2A/CEN9 (in 9p21.3
and 9p11q11 dual color probe), ZytoLight®SPEC
BIRC3/MALT1 Dual Color Dual Fusion Probe (in 11q22.2 and 18q21.32),
ZytoLight®SPEC FUS (16p11.2 Break Apart Probe)
all from ZytoVision GmbH; LSI EGR1/D5S23, D5S721 (in 5q31 and
5p15.2), LSI PDGFRB (5q32-q33 Break apart probe), subtelomeric
probe for 5qter (D5S2907), LSI ABL1/BCR (in 9q34.1 and 22q11.2 dual
color probe), LSI p53/ATM (in 17p13.1 and 11q22.3 dual color probe)
all from Vysis (Abbott GmbH & Company, KG) and POSEIDON
MLL/MLLT3 (in 11q23 and 9p21 dual color probe, Kreatech
Diagnostics).
A total of 10–15 metaphase spreads were analyzed and
(where applicable) for interphase-FISH analysis, 200 interphase
nuclei were examined, using a fluorescence microscope
(AxioImager.Z1 mot; Zeiss) equipped with appropriate filter sets to
discriminate between a maximum of five fluorochromes and the
counterstain 4,6-diamidino-2-phenylindole (DAPI). Image capturing
and processing were carried out using an ISIS imaging system
(MetaSystems).
DNA isolation
Genomic DNA was extracted from bone marrow cells by
Puregene DNA Purification Kit (Gentra Systems). DNA concentration
was determined by a NanoDrop spectrophotometer. The quality of DNA
was checked using agarose gel electrophoresis. DNA-samples
extracted from fixed cells of 2 healthy males and 2 healthy females
by the same method were used as reference samples.
High resolution array-comparative
genomic (aCGH)
aCGH was performed using Agilent SurePrint G3 Human
Genome microarray 180 K (Agilent Technologies), an oligonucleotide
microarray containing approximately 180,000 probes 60-mer with a 17
kb average probe spacing. Genomic DNA of patient was co-hybridized
with a male control DNA (Agilent Technologies). Labeling was
performed using Agilent Genomic DNA enzymatic labeling kit (Agilent
Technologies) according to the manufacturers' instructions. After
hybridization, the aCGH slide was scanned on an Agilent scanner,
processed with Feature Extraction software (v10.7) and results were
analyzed using Cytogenomics (v2.9.1.3) using ADM2 as aberration
algorithm.
Results
GTG-banding at low resolution revealed a female
karyotype as 46,XX,t(X;19)(q13;q13.3),der(9)[10]/46,XX[10] (result
not shown). Molecular analyses of RNA/cDNA did not detect any hint
on presence of tested fusion genes BCR/ABL1 (p190,
p210), MLL/AF4, ETV6/RUNX1 or
PBX1/E2A (results not shown). Application of
molecular cytogenetics approaches including WCPs, aMCB probe sets
and locus specific probes revealed the karyotype 46, XX,
der(5)t(5;9)(pter->q32::p21.3->pter),
der(9)(9qter->9q33.3::9p21.3->9q33.3::5q32->5qter),t(16;19)(p11.2;
q13.33) in the aberrant clone. Overall, the present case had
genetic changes involving four chromosomes and five break events
(Fig. 1).
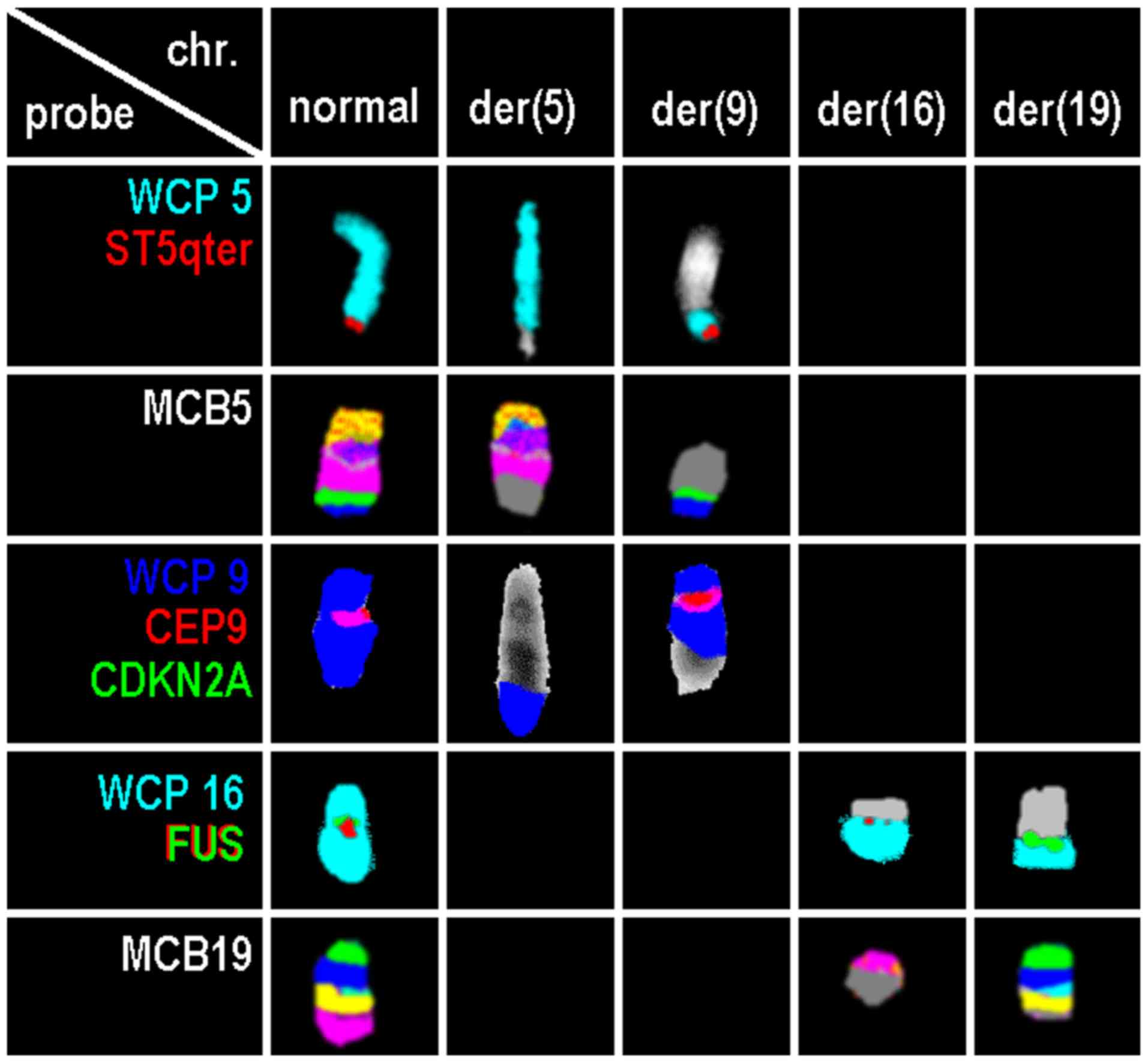 | Figure 1.Results of normal and derivative
chromosomes 5, 9, 16 and 19 for probes or probe sets. Probes or
probe sets are indicated in the first column. chr., chromosome;
der, derivative; WCP, whole chromosome probe; CDKN2A,
cyclin-dependent kinase inhibitor 2A; CEP, centromeric probe; chr,
chromosome; der, derivative chromosome; FUS, fused in sarcoma; MCB,
multicolor banding; qter, long arm terminal; ST, subtelomeric
probe. |
The breakpoint for translocation t(16;19) covered
FUS gene in 16p11.2 (Fig. 1).
Using locus specific probes, the breakpoints at 5q32, 9p21.3, 9q33
and 19q13.33 could be narrowed down, as well (Table I). For the 19q13.33, the break was
shown to be located between the positions:
Chr19:53,431,708-54,767,615 (NCBI36/hg18); 48 OMIM genes are
located there (Table II).
| Table II.Summary of CNAs detected by
array-comparative genomic hybridization. |
Table II.
Summary of CNAs detected by
array-comparative genomic hybridization.
| Chromosome
(alteration U or R) | Cytobands | NCBI36/hg18 | Size of imbalance
(Mb) | Online Mendelian
Inheritance in Man genes |
|---|
| 5 (U) |
del(5)(q31.3q32) |
chr5:142,130,476-145,871,262 | 3.74 | ARHGAP26, NR3C1,
HMHB1, YIPF5, KCTD16, PRELID2, GRXCR2, SH3RF2, PLAC8L1, LARS,
RBM27, POU4F3, TCERG1 |
| 9 (R) |
del(9)(p21.3p21.3) |
chr9:21,212,342-22,632,472 | 1.42 | IFNA17, IFNA14,
IFNA22P, IFNA5, KLHL9, IFNA6, IFNA13, IFNA2, IFNA8, IFNA1, MIR31HG,
IFNE, MIR31, MTAP, CDKN2A-DT, CDKN2A, CDKN2B-AS1, CDKN2B,
DMRTA1 |
| 9 (U) |
del(9)(q33.3q33.3) |
chr9:124,813,871-125,060,804 | 0.24 | RABGAP1, GPR21,
MIR600HG, STRBP |
| 22 (U) |
del(22)(q11.22q11.22) |
chr22:20,811,200-21,422,368 | 0.61 | VPREB1, BMS1P20,
ZNF280B, ZNF280A, PRAME, LL22NC03-63E9.3, POM121L1P,
GGTLC2 |
Array-CGH revealed four additional small genomic
imbalances: A loss of 1.42 Mb in the region of 9p21.3p21.3 between
the positions 21,212,342 and 22,632,472 (NCBI36/hg18) with
biallalic deletion of CDKN2A and CNKN2B and confirmed
by locus specific FISH probe result (Fig. 1).
Additional genomic imbalances revealed by array-CGH
were losses in: 5q31.3-q32 at positions 142,130,476-145,871,262
including loss of NR3C1; 9q33.2-q33.3 at positions
124,813,871 and 125,060,804 (no specific candidate gene
identified), and 22q11.22 at positions 20,811,200 and 21,422,368
including loss of VPREB1 (Table
II).
Discussion
In pediatric B-ALL, chromosomal rearrangements play
a crucial role in leukemogenesis by contributing to inactivation of
tumor suppressor genes and/or dysregulated expression of oncogenes
and the generation of novel gene fusions. Thus, chromosomal
break-events can be hints on genes with biologic function in the
leukemogenesis. Here we showed involvement of Fused in Sarcoma
(FUS) gene located in chromosome 16p11.2 in childhood B-ALL.
Interestingly, very little is known about the oncogenic role of
FUS gene in ALL (17).
FUS gene was first identified as an oncogene in human myxoid
liposarcomas, being activated by translocations (18). FUS gene encodes an RNA-binding
protein (takes part in transcription initiation by binding RNA pol
II and some transcription factors), the C-terminal end of which is
involved in protein and RNA binding and which appears to be
involved in transcriptional activation with its N-terminal end. FUS
is a nucleoprotein that functions in DNA and RNA metabolism,
including DNA repair, and the regulation of transcription, RNA
splicing and export to the cytoplasm (19,20).
In acute leukemia, FUS gene rearrangements
have been identified in both childhood and adults: The recurrent
translocation t(16;21)(p11;q22)/FUS-ERG was yet seen in
approximately 80 cases of acute myeloid leukemia (AML). It occurs
in <0.5% of AML cases and is associated with poor prognosis and
high risk of relapse (21,22). In ALL, the translocation
t(16;21)(p11;q22)/FUS-ERG is very rare and only identified
in 15 patients, including 10 adults and five children (17,22). In
addition, the translocation t(9;16)(q34;p11.2) FUS/SET has
been detected in one T-ALL patient (23).
To the best of our knowledge, the balanced
translocation t(16;19)(p11.2;q13.3)?/FUS has not yet been
reported according to the Mitelman database and the literature.
This translocation seems to create a yet unknown fusion gene
between FUS gene on 16p11.2 and a gene on 19q13.33. In
19q13.33 region 48 OMIM genes are located, including the
EMP3 gene (epithelial membrane protein 3); the translocation
t(11;19)(q23;q13)CEP164/EMP3 has been detected previously in only
one T-ALL patient. However in gene-rich subband 19q13.33, it is
difficult to determine which one might have provided a fusion with
or strong promoter for FUS gene dysregulate expression.
Overall, FUS gene at 16p11.2 is considered as one of the
fusion products being critical for leukaemogenic process as a sole
abnormality (2,17,22–25). On
the other hand, an unbalanced translocation in this case involves
three other breakpoints 5q32, 9p21.3 and 9q33. Participation of
cancer-related oncogenes PDGFRB in 5q32, MLLT3 in
9p21.3, and ABL1 in 9q34.13 could be excluded (Table I), however, loss of two tumor
suppressor genes in 9p21 was identified.
Interestingly, the unbalanced translocation involves
simultaneously interstitial deletions and complex rearrangements.
Tumor suppressor genes CDKN2A and CDKN2B at 9p21,
which encode p14/ARF and p16/INK4A (both encoded by CDKN2A)
and p15/INK4B (encoded by CDKN2B) proteins have a role in
cell growth regulation and apoptosis. Thus, here observed
homozygous deletion represents a marker of high risk of relapse and
poor prognosis in children B-ALL as reported in previous studies
(26–28).
NR3C1 gene in 5q31.3, encoding DNA- and
hormone-binding domains of the human glucocorticoid receptor, can
function both as a transcription factor that binds to
glucocorticoid response elements in promoters of glucocorticoid
responsive genes and activate their transcription, and as a
regulator of other transcription factors. Interestingly, loss of
NR3C1 gene is associated with glucocorticoid resistance,
which is a key component of childhood ALL therapy. Besides,
deletion of NR3C1 gene is rare in B-ALL and significantly
associated with clinical high risk of relapse and an inferior
outcome, where it correlated to high rate of induction failure and
death (2,7,29).
Furthermore, VPREB1 gene deletion at 22q11.2
has been currently reported in association with poor prognosis and
high risk of relapse in both children and adult B-ALL.
VPREB1 gene encodes the iota polypeptide chains, which
associate with the Ig-mu chain to form a molecular complex that
expressed on the surface of pre-B cells. VPREB1 gene is
located within the IGL@ locus among the variable
immunoglobulin (Ig) segments, upstream from the VJ junction and is
an essential gene in B-cell development and differentiation due to
its role as part of the surrogate light chain in the pre-B cell
receptor (pre-BCR). A recent report by Mangum et al
(10) described a new mechanism of
B-ALL development emerging by loss of VPREB1 gene during
B-cell maturation. In this context, loss of VPREB1 gene
contributes to leukemogenesis as a result of failure to form a
viable surrogate light chain in the pre-BCR in both human and mice
due to block the pro- to pre-B-cell transition in the bone marrow
with a decrease in circulating mature B-cells. Besides,
VPREB1 gene deletion was also identified in specific B-ALL
subtypes including ETV6-RUNX1 and associate with higher WBC
count as also in our case, BCR-ABL1 and TCF3-HLF
(6,8,10,30,31).
In conclusion, further studies are required to
establish whether such potentially poor outcome due to the
combination of the here reported aberrations is linked directly to
glucocorticoid resistance or whether it is part of a broader drug
resistance profile. We suggest NR3C1 and VPREB1 genes
as excellent candidates for further studying B-ALL leukemogenesis
to improve treatment strategies in pediatric B-ALL. Overall, we
provided evidence, that comprehensive molecular analyses are
urgently necessary for the detection of cryptic rearrangements and
identification of genes involved in B-ALL formation and
progression.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
MAKO, BM and FAS performed the FISH studies. JBM,
IMC and MAKO performed array comparative genomic hybridization
analyses and interpretation. NL performed the reverse transcription
PCR. MĐ, ZZ, DV and GS provided B-ALL-case, including clinical
data, and performed banding cytogenetic data interpretation and
made decisions on what tests should be performed in their lab.
MAKO, RA and TL developed the study, made decisions on what tests
should be performed, were involved in overall data interpretation
and drafted the study. All authors read and approved the study.
Ethics approval and consent to
participate
As the case was identified during routine
diagnostics, ethical approval and consent to participate is not
applicable, except for the agreement that diagnostics was performed
to solve the clinical question.
Patient consent for publication
Informed consent was provided by the patient's
parent.
Competing interests
BM works at Zytovison, the company providing one of
the FISH-probes used. All other authors declare that they have no
competing interests.
References
|
1
|
Malouf C and Ottersbach K: Molecular
processes involved in B cell acute lymphoblastic leukaemia. Cell
Mol Life Sci. 75:417–446. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tasian SK and Hunger SP: Genomic
characterization of paediatric acute lymphoblastic leukaemia: An
opportunity for precision medicine therapeutics. Br J Haematol.
176:867–882. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Moorman AV: New and emerging prognostic
and predictive genetic biomarkers in B-cell precursor acute
lymphoblastic leukemia. Haematologica. 101:407–416. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Moorman AV, Harrison CJ, Buck GA, Richards
SM, SeckerWalker LM, Martineau M, Vance GH, Cherry AM, Higgins RR,
Fielding AK, et al: Karyotype is an independent prognostic factor
in adult acute lymphoblastic leukemia (ALL): Analysis of
cytogenetic data from patients treated on the Medical Research
Council (MRC) UKALLXII/Eastern Cooperative Oncology Group (ECOG)
2993 trial. Blood. 109:3189–3197. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hamadeh L, Enshaei A, Schwab C, Alonso CN,
Attarbaschi A, Barbany G, den Boer ML, Boer JM, Braun M, Dalla
Pozza L, et al: Validation of the United Kingdom copy-number
alteration classifier in 3239 children with B-cell precursor ALL.
Blood Adv. 3:148–157. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ribera J, Zamora L, Morgades M, Mallo M,
Solanes N, Batlle M, Vives S, Granada I, Juncà J, Malinverni R, et
al: Copy number profiling of adult relapsed B-cell precursor acute
lymphoblastic leukemia reveals potential leukemia progression
mechanisms. Genes Chromosomes Cancer. 56:810–820. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Irving JA, Enshaei A, Parker CA, Sutton R,
Kuiper RP, Erhorn A, Minto L, Venn NC, Law T, Yu J, et al:
Integration of genetic and clinical risk factors improves
prognostication in relapsed childhood B-cell precursor acute
lymphoblastic leukemia. Blood. 128:911–922. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Öfverholm I, Tran AN, Olsson L,
Zachariadis V, Heyman M, Rudd E, Syk Lundberg E, Nordenskjöld M,
Johansson B, Nordgren A and Barbany G: Detailed gene dose analysis
reveals recurrent focal gene deletions in pediatric B-cell
precursor acute lymphoblastic leukemia. Leuk Lymphoma.
57:2161–2170. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Othman MA, Melo JB, Carreira IM, Rincic M,
Glaser A, Grygalewicz B, Gruhn B, Wilhelm K, Rittscher K, Meyer B,
et al: High rates of submicroscopic aberrations in karyotypically
normal acute lymphoblastic leukemia. Mol Cytogenet. 8:452015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mangum DS, Downie J, Mason CC, Jahromi MS,
Joshi D, Rodic V, Müschen M, Meeker N, Trede N, Frazer JK, et al:
VPREB1 deletions occur independent of lambda light chain
rearrangement in childhood acute lymphoblastic leukemia. Leukemia.
28:216–220. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Claussen U, Michel S, Mühlig P, Westermann
M, Grummt UW, Kromeyer-Hauschild K and Liehr T: Demystifying
chromosome preparation and the implications for the concept of
chromosome condensation during mitosis. Cytogenet Genome Res.
98:136–146. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shaffer LG, McGowan-Jordan J and Schmid M:
An International System for Human Cytogenetic Nomenclature. Karger;
Basel: pp. 2108–2109. 2013
|
|
13
|
Liu X and Harada S: RNA isolation from
mammalian samples. Curr Protoc Mol Biol. Chapter 4:Unit 4.16. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
van Dongen JJ, Macintyre EA, Gabert JA,
Delabesse E, Rossi V, Saglio G, Gottardi E, Rambaldi A, Dotti G,
Griesinger F, et al: Standardized RT-PCR analysis of fusion gene
transcripts from chromosome aberrations in acute leukemia for
detection of minimal residual disease. Report of the BIOMED-1
Concerted Action: Investigation of minimal residual disease in
acute leukemia. Leukemia. 13:1901–1928. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Weise A, Mrasek K, Fickelscher I, Claussen
U, Cheung SW, Cai WW, Liehr T and Kosyakova N: Molecular definition
of high-resolution multicolor banding probes: First within the
human DNA sequence anchored FISH banding probe set. J Histochem
Cytochem. 56:487–493. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liehr T: Fluorescence In Situ
Hybridization (FISH)-Application Guide. Springer Verlag; Berlin:
2017
|
|
17
|
Coccé MC, Alonso CN, Rossi J, Felice MS,
Gitter MR and Gallego MS: A case of pediatric ALL with
t(16;21)(p11.2;q22) and FUS-ERG rearrangement. Blood Res. 50:55–58.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Aman P, Ron D, Mandahl N, Fioretos T, Heim
S, Arheden K, Willén H, Rydholm A and Mitelman F: Rearrangement of
the transcription factor gene CHOP in myxoid liposarcomas with
t(12;16)(q13;p11). Genes Chromosomes Cancer. 5:278–285. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Naro C, Bielli P, Pagliarini V and Sette
C: The interplay between DNA damage response and RNA processing:
The unexpected role of splicing factors as gatekeepers of genome
stability. Front Genet. 6:1422015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou Y, Liu S, Oztürk A and Hicks GG:
FUS-regulated RNA metabolism and DNA damage repair: Implications
for amyotrophic lateral sclerosis and frontotemporal dementia
pathogenesis. Rare Dis. 2:e295152014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Noort S, Zimmermann M, Reinhardt D,
Cuccuini W, Pigazzi M, Smith J, Ries RE, Alonzo TA, Hirsch B,
Tomizawa D, et al: Prognostic impact of t(16;21)(p11;q22) and
t(16;21)(q24;q22) in pediatric AML: A retrospective study by the
I-BFM Study Group. Blood. 132:1584–1592. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zerkalenkova E, Panfyorova A, Kazakova A,
Baryshev P, Shelihova L, Kalinina I, Novichkova G, Maschan M,
Maschan A and Olshanskaya Y: Molecular characteristic of acute
leukemias with t(16;21)/FUS-ERG. Ann Hematol. 97:977–988. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Atak ZK, Gianfelici V, Hulselmans G, De
Keersmaecker K, Devasia AG, Geerdens E, Mentens N, Chiaretti S,
Durinck K, Uyttebroeck A, et al: Comprehensive analysis of
transcriptome variation uncovers known and novel driver events in
T-cell acute lymphoblastic leukemia. PLoS Genet. 9:e10039972013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mitelman F, Johansson B and Mertens F:
Mitelman Database of Chromosome Aberrations and Gene Fusions in
Cancer. National Cancer Institute; Bethesda, MD: April 18–2019
|
|
25
|
Liu Y, Easton J, Shao Y, Maciaszek J, Wang
Z, Wilkinson MR, McCastlain K, Edmonson M, Pounds SB, Shi L, et al:
The genomic landscape of pediatric and young adult T-lineage acute
lymphoblastic leukemia. Nat Genet. 49:1211–1218. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kathiravan M, Singh M, Bhatia P, Trehan A,
Varma N, Sachdeva MS, Bansal D, Jain R and Naseem S: Deletion of
CDKN2A/B is associated with inferior relapse free survival in
pediatric B cell acute lymphoblastic leukemia. Leuk Lymphoma.
60:433–441. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Agarwal M, Bakhshi S, Dwivedi SN, Kabra M,
Shukla R and Seth R: Cyclin dependent kinase inhibitor 2A/B gene
deletions are markers of poor prognosis in Indian children with
acute lymphoblastic leukemia. Pediatr Blood Cancer. 65:e270012018.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Braun M, Pastorczak A, Fendler W, Madzio
J, Tomasik B, Taha J, Bielska M, Sedek L, Szczepanski T, Matysiak
M, et al: Biallelic loss of CDKN2A is associated with poor response
to treatment in pediatric acute lymphoblastic leukemia. Leuk
Lymphoma. 58:1162–1171. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Donner KM, Hiltunen TP, Jänne OA, Sane T
and Kontula K: Generalized glucocorticoid resistance caused by a
novel two-nucleotide deletion in the hormone-binding domain of the
glucocorticoid receptor gene NR3C1. Eur J Endocrinol. 168:K9–K18.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Eswaran J, Sinclair P, Heidenreich O,
Irving J, Russell LJ, Hall A, Calado DP, Harrison CJ and Vormoor J:
The pre-B-cell receptor checkpoint in acute lymphoblastic
leukaemia. Leukemia. 29:1623–1631. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mårtensson IL and Ceredig R: Review
article: Role of the surrogate light chain and the pre-B-cell
receptor in mouse B-cell development. Immunology. 101:435–441.
2000. View Article : Google Scholar : PubMed/NCBI
|














