Introduction
Cervical cancer is the fourth most common cancer of
woman worldwide (1). In 2012, the
incidence rate for cervical cancer was ~7.5/100,000 women worldwide
(2). Currently, human papillomavirus
is observed in nearly all reported cases of cervical cancer
(3), whereby persistent infection
with human papillomavirus is associated with an increased risk of
developing cervical cancer (3).
Surgical removal, chemotherapy, radiotherapy or concurrent
chemoradiation are the common methods used to treat the different
subtypes of cervical cancer (4,5).
Although cisplatin-based chemoradiation yields overall survival
rates of 66%, cisplatin is considered to cause kidney damage
following long-term usage (6). Thus,
the development of novel targets for the treatment of cervical
cancer remains critical.
Tropomodulins (TMOD) are a family of proteins that
cap the pointed end of actin filaments and regulate their
depolymerization (7). There are
currently four confirmed TMODs: TMOD1, 2, 3 and 4 that are involved
in a variety of cellular functions, such as myofibril alignment
(8), neurite extension (9) and endothelial cell migration (10). TMOD1 and TMOD3 demonstrate broad
expression, while TMOD2 and TMOD4 demonstrate specific expression
in the nervous system and skeletal muscle, respectively (7). Due to the role of TMODs in regulating
actin dynamics, TMODs were observed to be associated with the
negative regulation of neurite outgrowth (9) and cell migration (10). As a key member of the TMOD family,
TMOD1 has been extensively researched thus far (7). Despite its involvement in the nervous
system and in cardiac muscle, TMOD1 has been implicated in cancer
development through its function in actin dynamics regulation
(11). High TMOD1 expression has
been reported to restore thymosin β-10 mediated apoptosis in SKOV-3
ovarian cancer cells (12).
Furthermore, TMOD1 has been demonstrated to enhance hematopoietic
reconstitution (13). It has also
been reported that TMOD1 may serve as a target gene for NF-κB and
promote the development of breast cancer via the
NF-κB-TMOD1-β-catenin-MMP13 axis (14). Furthermore, overexpression of TMOD1
has been demonstrated to enhance regional lymph node metastasis in
human oral cancer (15). Recently,
the TMOD1 loci was screened as a potential risk loci in patients
with esophageal adenocarcinoma using a high-density single
nucleotide polymorphism array (16).
TMOD1 has been implicated in several types of
cancer; however, its role in cervical cancer remains unknown. The
present study investigated TMOD1 function in two cervical cancer
cell lines, HeLa and CaSki, assessing its effects on cell motility,
cell proliferation and cell cycle by downregulating or
overexpressing TMOD1 in cervical cancer cells. Furthermore,
clinical data obtained from patients with cervical cancer was
assessed. The results indicate an association between the TMOD1
expression and the development of cervical cancer, which provides
novel evidence on the ambiguous role of TMOD1 in cervical
cancer.
Materials and methods
Plasmids
Non-targeting short hairpin (sh)RNA and specific
shRNA for human TMOD1 were purchased from Sigma-Aldrich (Merck
KGaA). The targeting sequence of TMOD1 shRNA-1 localizes in the
untranslated region of the TMOD1 gene. The TMOD1 overexpression
vector was constructed with the empty vector, CSII-CMV-MCS (RIKEN
BioResource Center) and amplified according to the coding sequences
of the TMOD1 gene. CSII-CMV-MCS was used as a control vector in
overexpression experiments. The TMOD1 overexpression vector was
used as a shRNA resistant TMOD1 expression vector. The sequences of
shRNAs are presented in Table I.
 | Table I.Sequences of shRNAs. |
Table I.
Sequences of shRNAs.
| shRNA clone | Antisense
sequence |
|---|
| Human TMOD1
shRNA-1 |
5′-ATTTGGCAACTTTAATCGAGG-3′ |
| Human TMOD1
shRNA-2 |
5′-ATATTCCGGATATTATTGAGG-3′ |
Cell culture and transfection
The HeLa and CaSki cervical cancer cell lines were
obtained from the American Type Culture Collection. Cells were
cultured in DMEM supplemented with 10% heat inactivated FBS at 37°C
in a 5% CO2 incubator. Cells were plated into 6-well
plates at a density of 1×105 cells/ml/plate and
transfected with 6 µl of Fugene HD (Promega Corporation), followed
by 1.5 µg of TMOD1 shRNAs (1 µg/µl) with or without 1.5 µg of TMOD1
overexpression vector (1 µg/µl), thus the final volume of
transfection mixture was 100 µl. After 24 h, cells were treated
with puromycin (Thermo Fisher Scientific, Inc.) at 37°C, for 2
consecutive days.
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted from the cultured cells
using ISOGEN lysis buffer (Nippon Gene Co., Ltd.), according to the
manufacturer's protocol. Total RNA was reverse transcribed into
cDNA using the ReverTra Ace® qPCR RT kit (Toyobo Life
Science), according to the manufacturer's protocol. qPCR was
subsequently performed using the SYBR® Green Real-time
PCR master mix (Toyobo Life Science), as previously described
(17). The method of quantification
was performed according to a previous report (18). The primer sequences of TMOD1, Twist,
Snail and GAPDH are presented in Table
II.
| Table II.Primer sequences for PCR
experiments. |
Table II.
Primer sequences for PCR
experiments.
| Primer name | Forward primer
sequence (5′-3′) | Reverse primer
sequence (5′-3′) |
|---|
| Human TMOD1 |
GTGGAAATGGAGATTGTGAGC |
TCATGCTGCCAGCATTTTGC |
| Human GAPDH |
ATCATCCCTGCCTCTACTGG |
CCCTCCGACGCCTGCTTCAC |
| Human Snail |
TCGGAAGCCTAACTACAGCGA |
AGATGAGCATTGGCAGCGAG |
| Human Twist |
GTCCGCAGTCTTACGAGGAG |
GCTTGAGGGTCTGAATCTTGCT |
Western blot analysis
Total protein was extracted from HeLa or CaSki cells
using RIPA lysis buffer (50 mM Tris-HCl pH 7.4, 150 mM NaCl, 1%
NP-40, 1% sodium dexycholate, 0.1% SDS), and protein concentration
was measured using a Pierce bicinchoninic acid protein assay kit
(Thermo Fisher Scientific, Inc). A total of 15 µg protein/lane was
separated via SDS-PAGE on a 12% gel. The separated proteins were
subsequently transferred onto a nitrocellulose membrane (GE
Healthcare) and blocked with 5% skimmed milk, supplemented in PBS
with 0.1% Tween-20 for 1 h at room temperature. The membranes were
incubated with primary antibodies against TMOD1 (1:1,000; cat. no.
H00007111-B01P; Novus Biologicals, LCC) and β-actin (1:1,000; cat.
no. A5441; Sigma-Aldrich; Merck KGaA) overnight at 4°C. Membranes
were washed three times with 0.1% Tween-20 in PBS. Following the
primary antibody incubation, membranes were incubated with
horseradish peroxidase-conjugated secondary antibodies (1:5,000;
cat. no. 115-035-003; Jackson ImmunoResearch Laboratories,
Inc.) for 1 h at room temperature. Membranes were washed three
times again with 0.1% Tween-20 in PBS, and protein bands were
visualized using the ECL western blotting substrate (GE
Healthcare), according to the manufacturer's protocol.
In vitro wound-healing assay
The wound-healing assay was performed as described
by Zhang et al (19), with a
modification on the culture time following wound generation. A
total of 8 ×105 cells/ml HeLa cells or 1.2×
106 cells/ml CaSki cells were seeded into a 6-well plate
and incubated at 37°C with DMEM supplied with 10% heat-inactivated
FBS, until ~100% confluence was achieved. The straight cell-free
wound was created using a 100 µl pipette tip in the center of the
plate. Cells were subsequently washed twice with PBS to remove any
debris and incubated with serum-free media at 37°C for 24 h. Image
was captured using a light microscope (magnification, ×100). The
distance migrated by the cell monolayer was measured during this
period, in order to close the wounded area. The results were
presented as a relative migration ratio as follows: Distance
migrated by TMOD1 shRNAs or shRNA-resistant TMOD1, with TMOD1
shRNA-1 treated cells are relative to the distance migrated by
control shRNAs treated cells. The distance was determined using
ImageJ software [version 1.8.0; (20)].
Cell proliferation assay
The transfected cells were re-seeded into 96 well
plates at a density of 1×104 cells/ml of HeLa cells or
3×104 cells/ml of CaSki cells and allowed to adhere
overnight. At day 0, the culture medium was changed from 10% FBS to
DMEM supplemented with 2% heat inactivated FBS. At day 4, cell
number was measured using the Cell Counting Kit-8 (Dojindo
Molecular Technologies, Inc.) at a wavelength of 450 nm, according
to the manufacturer's protocol.
Flow cytometry
Cell cycle analysis was performed as described by Lu
et al (17). A total of
1×106 cells/ml HeLa cells or CaSki cells were collected
following treatment with trypsin at 37°C for 5 min, and washed once
with 5 ml of cold PBS prior to fixation with absolute ethanol
overnight at −20°C. Fixed cells were centrifuged at 167.7 × g/min
at 4°C for 5 min and re-suspended in 1 ml of PBS. Subsequently,
cells were treated with 10 µl of RNase A (10 mg/ml) for 30 min at
37°C, and supplemented with 50 µl of propidium iodide (1 mg/ml) for
30 min at 4°C. Cell cycle analysis was performed using a FACS Canto
flow cytometer (BD Biosciences) and FlowJo software (version 7.6;
(TOMY Digital Biology Co., Ltd.).
Terminal
deoxynucleotidyl-transferase-mediated dUTP nick end labeling
(TUNEL) assay
Following transfection with TMOD1 shRNA or control
shRNA, HeLa or CaSki cells were re-seeded into 8-well plates (BD
Biosciences) at a density of 3×104 cells/chamber, after
3 days. After 24 h, the TUNEL assay was performed, according to the
manufacturer's protocol (21).
Briefly, cells were washed three times with PBS and subsequently
fixed with 4% formaldehyde for 15 min at 4°C. Cells were incubated
with 0.5% Tween 20 in PBS supplemented with 0.2% BSA at room
temperature for 15 min and re-washed three times with PBS, prior to
incubation in 100 µl of TdT mixture (90 µl of TdT buffer, 5 µl of
FITC-dUTP and 5 µl of TdT; Medical & Biological Laboratories,
Co., Ltd.) at 37°C for 1 h. Cell nuclei were counterstained with
DAPI (final concentration: 2 µg/ml) for 10 min at room temperature
and mounted with mounting medium (Abcam). TUNEL-positive cells were
observed in three randomly-selected fields under a fluorescent
microscope (magnification, ×100).
Matrigel invasion assay
Transwell membranes (pore size, 8 mm; BD
Biosciences) were precoated with 100 µl of 5 mg/ml Matrigel (BD
Biosciences) at 37°C for 6 h. A total of 5×104 HeLa or
CaSki cells were harvested 48 h post-transfection and plated in the
upper chambers of Transwell plates in % BSA. Simultaneously, 10%
FBS was plated in the lower chambers. Following incubation at 37°C
for 24 h, the non-migratory cells in the upper chambers were
removed and the migratory cells were stained with DAPI for 10 min
at room temperature. Stained cells were counted in four
randomly-selected fields under a fluorescent microscope
(magnification, ×100).
Clinical patient data
The raw TMOD1 gene expression data and corresponding
clinical parameters were downloaded from the publicly available R2
database (http://hgserver1.amc.nl/cgi-bin/r2/main.cgi). The
GSE2109 cervical cancer dataset (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE2109)
employed in R2 was downloaded from the Gene Expression Omnibus
(GEO) database (http://www.ncbi.nlm.nih.gov/geo). The GEO database
includes 36 clinical samples, of which 25 cervical cancer specimens
where selected in the present study, which exhibited detectable
TMOD1 expression. All samples were hybridized to Affymetrix human
genome U133 plus 2.0 microarrays. TMOD1 expression levels of 25
individuals are presented in Table
III.
| Table III.TMOD1 expression levels in the
clinical database. |
Table III.
TMOD1 expression levels in the
clinical database.
| Stage | Sample name | Relative TMOD1
expression level | Histology | Patient age,
years |
|---|
| I (n=12) | gsm117626 | 4.27 | Endometrioid
carcinoma | >40 |
|
| gsm152580 | 4.68 | Squamous cell
carcinoma, spindle cell type | <40 |
|
| gsm152635 | 6.12 | Squamous cell
carcinoma | <40 |
|
| gsm152667 | 5.09 | Adenocarcinoma | <40 |
|
| gsm152719 | 4.47 | Adenocarcinoma | <40 |
|
| gsm179853 | 7.02 | Adenosquamous
carcinoma | <40 |
|
| gsm179907 | 4.49 | Squamous cell
carcinoma, non-keratinizing | >40 |
|
| gsm179956 | 4.49 | Squamous cell
carcinoma, non-keratinizing | <40 |
|
| gsm203742 | 4.69 | Squamous cell
carcinoma, non-keratinizing | >40 |
|
| gsm325835 | 8.46 | Mucinous
Adenocarcinoma, endocervical type | <40 |
|
| gsm46919 | 5.46 | Squamous cell
carcinoma, verrucous | >40 |
|
| gsm46942 | 6.01 | Squamous cell
carcinoma, non-keratinizing | <40 |
| II (n=1) | gsm203799 | 4.79 | Squamous cell
carcinoma, non-keratinizing | >40 |
| III (n=9) | gsm117576 | 2.00 | Squamous cell
carcinoma, keratinizing | <40 |
|
| gsm152587 | 2.68 | Squamous cell
carcinoma | >40 |
|
| gsm152723 | 3.66 | Squamous cell
carcinoma, keratinizing | >40 |
|
| gsm152751 | 0.58 | Squamous cell
carcinoma, keratinizing | >40 |
|
| gsm152788 | 2.94 | Squamous cell
carcinoma | >40 |
|
| gsm102527 | 1.49 | Squamous cell
carcinoma | >40 |
|
| gsm203622 | 5.35 | Adenocarcinoma | >40 |
|
| gsm76614 | 1.32 | Squamous cell
carcinoma, non-keratinizing | >40 |
|
| gsm89050 | 5.30 | Squamous cell
carcinoma, non-keratinizing | <40 |
| IV (n=3) | gsm102481 | 4.83 | Squamous cell
carcinoma, non-keratinizing | <40 |
|
| gsm152787 | 2.83 | Squamous cell
carcinoma, non-keratinizing | <40 |
|
| gsm152800 | 1.77 | Squamous cell
carcinoma | >40 |
Statistical analysis
Statistical analysis was performed using GraphPad
(version 5; GraphPad Software, Inc.). Data are presented as the
mean ± standard deviation. Statistical analysis was determined
using one-way analysis of variance followed by Tukey's post hoc
test. All experiments were performed in triplicate. P<0.05 was
considered to indicate a statistically significant difference.
Results
Downregulation of TMOD1 promotes cell
motility in HeLa and CaSki cells
Given the close association of TMOD1 with
filamentary actin dynamics, the present study determined to
investigate the involvement of TMOD1 in cell motility.
First, HeLa and CaSki cells were transfected with
control non-targeting shRNA, TMOD1 shRNA-1, TMOD1 shRNA-2, or
shRNA-resistant TMOD1 plus TMOD1 shRNA-1, respectively. After 24 h,
puromycin (final concentration: 2 µg/ml) was added to the cell
culture medium, in order to obtain the shRNA positive cells.
Following transfection after 3 days, total RNA and protein were
extracted, and RT-qPCR and western blot analysis were performed,
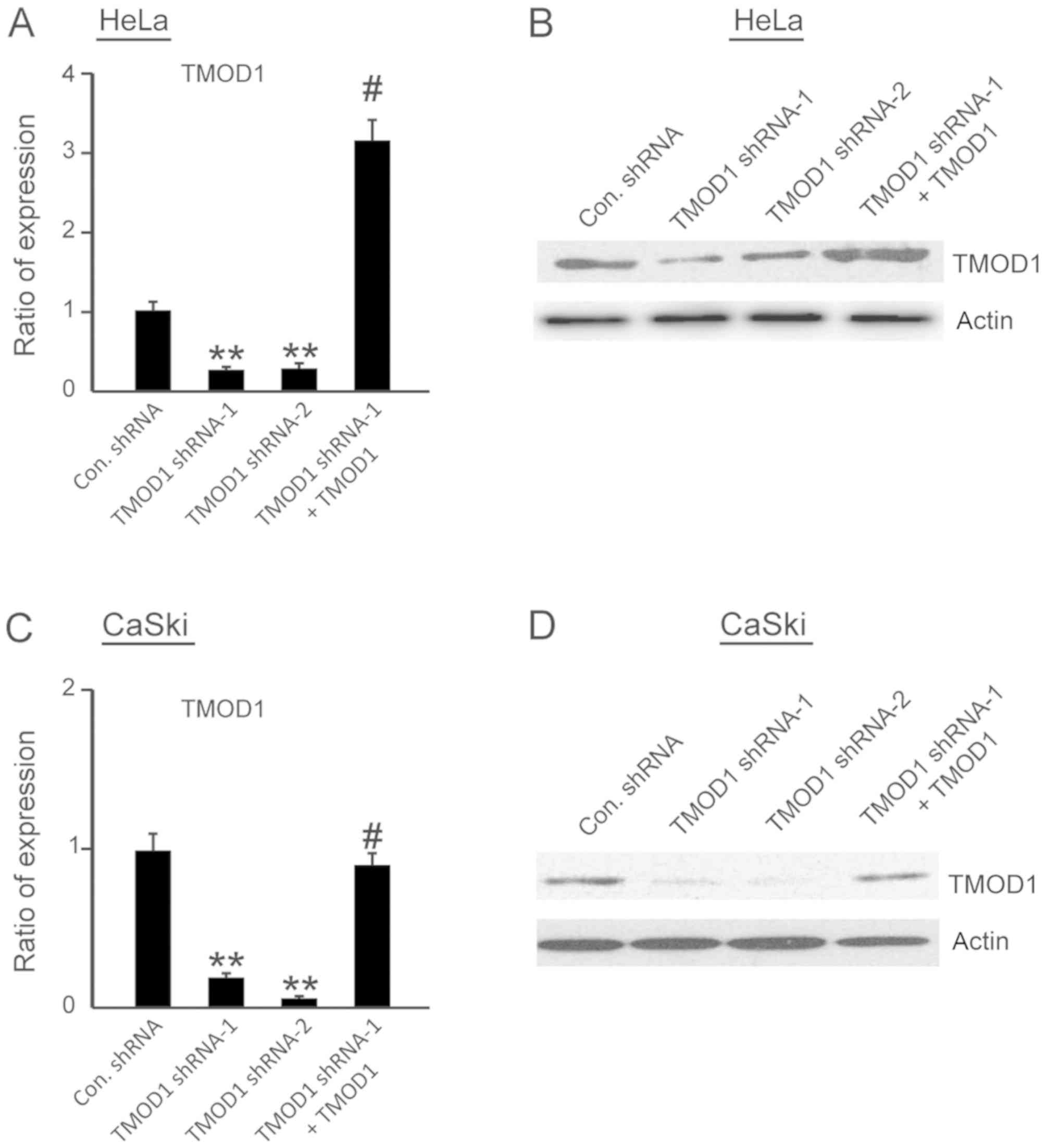
respectively, to determine knockdown efficiency. The results
demonstrated that both TMOD1 shRNA-1 and TMOD1 shRNA-2
significantly suppressed TMOD1 expression in HeLa and CaSki cells
(all P<0.001; Fig. 1A-D).
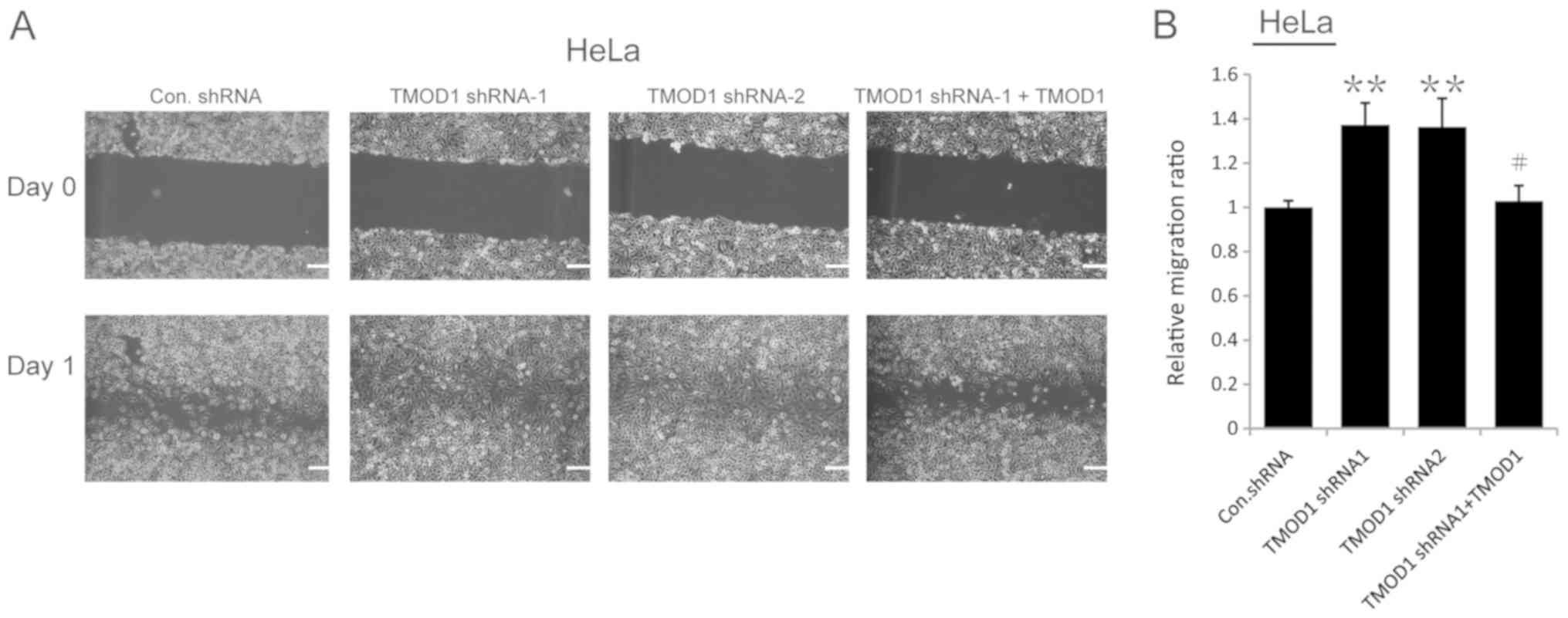
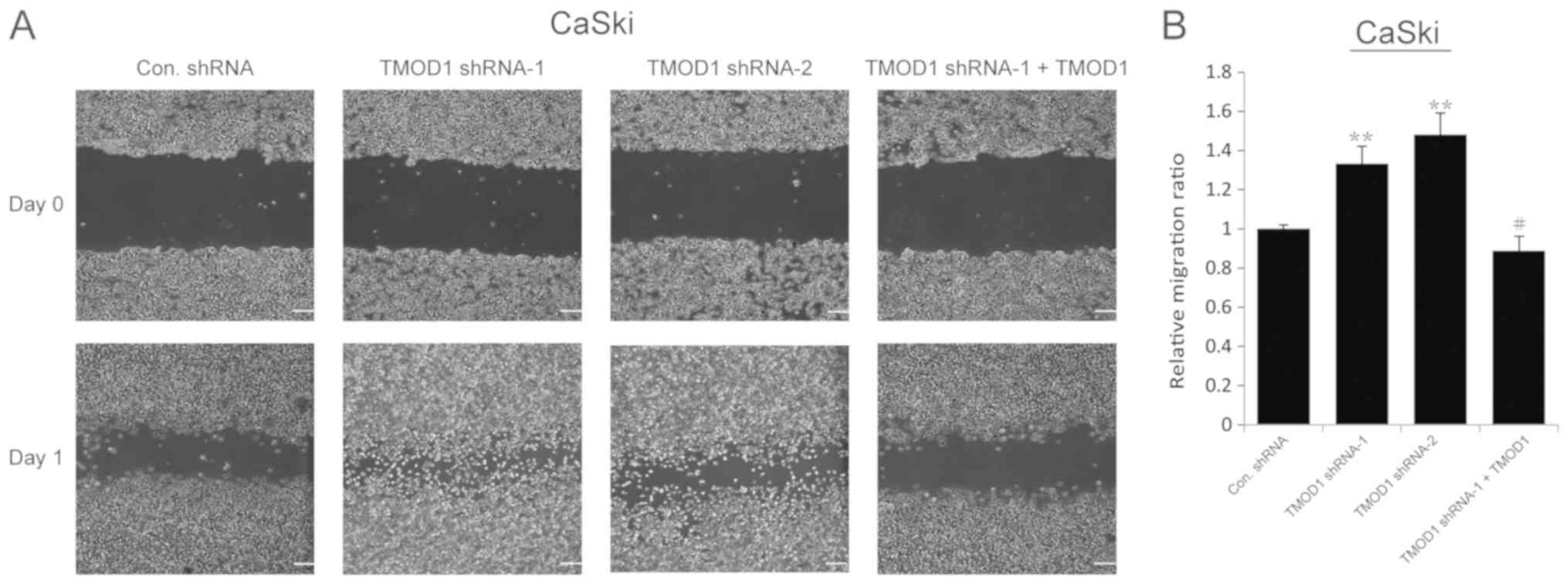
The results of the wound-healing assay demonstrated
that the motility of TMOD1 shRNAs-treated cells was more notable
compared with the control cells (all P<0.001; Figs. 2 and 3). In order to confirm this phenomenon,
rescue experiments were performed in which a shRNA-1-resistant
TMOD1 expressing vector was used to transfect TMOD1 shRNA-1 treated
cells. The results indicated that TMOD1 expression significantly
rescued the phenotype caused by TMOD1 shRNA-1 (all P<0.01;
Figs. 2 and 3). A Matrigel assay was performed to
investigate the role of TOMOD1 in cervical cancer cell invasion.
The results demonstrated that the tendency of cell invasion was
promoted in both Hela and CaSki cells following TMOD1 knockdown,
and differences between the control and TMOD1 shRNAs treated groups
were not significant (data not shown). The expression of two
epithelial-mesenchymal transition (EMT) markers, Twist and Snail,
was also analyzed via RT-qPCR to determine whether or not TMOD1
modulates cell migration by EMT, and no significant difference was
demonstrated (data not shown).
Downregulation of TMOD1 promotes cell
proliferation and G1/S transition in HeLa and CaSki
cells
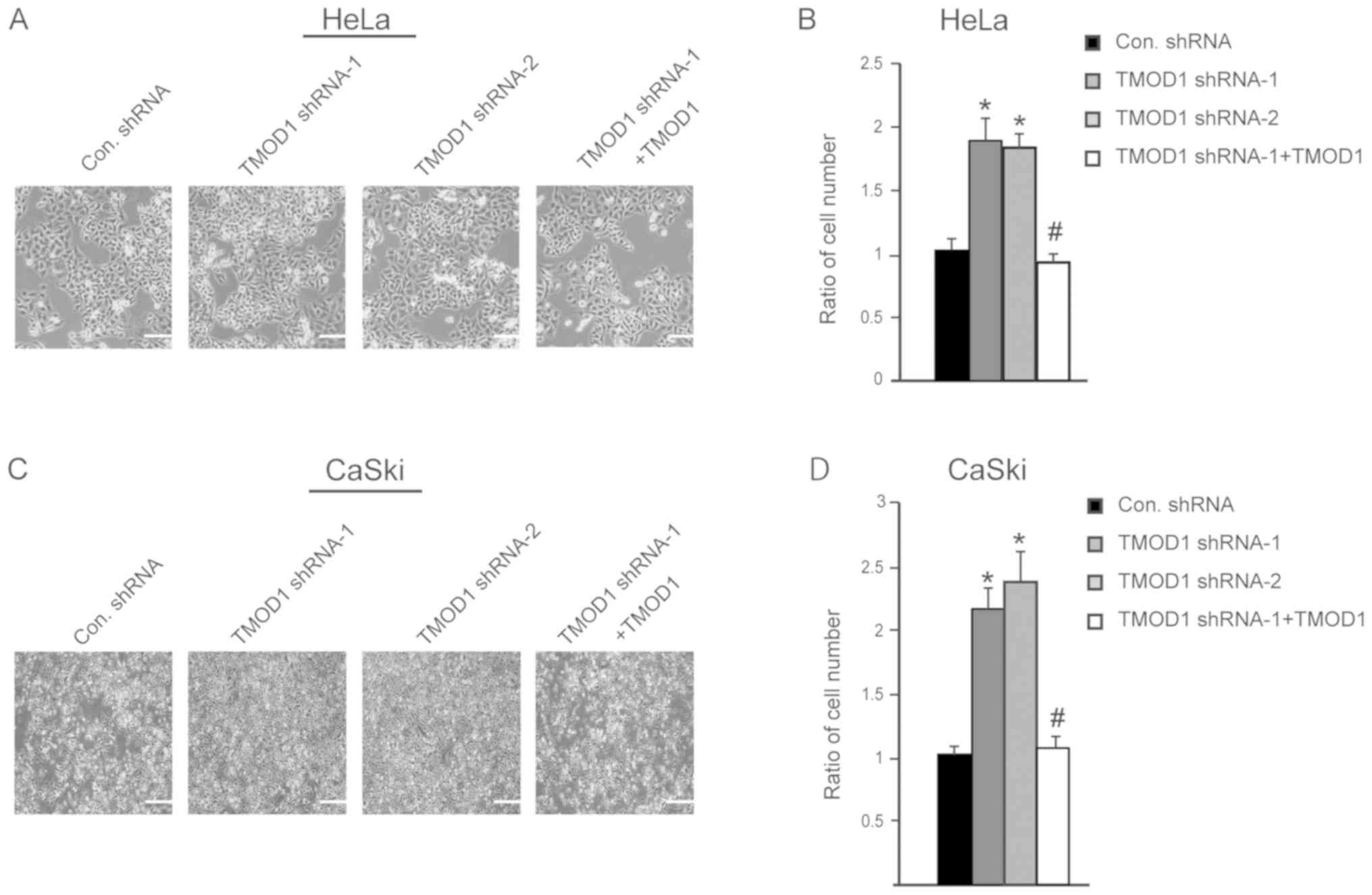
In order to improve the understanding of the role of
TMOD1 in cervical cancer, its expression on HeLa and CaSki cell
proliferation was subsequently assessed. The stable shRNA
transfected cells were seeded and cultured in DMEM supplied with 2%
heat inactivated FBS, and cell number was counted at day 4. The
results indicated that the cells transfected with TMOD1 shRNAs
proliferated at a faster rate compared with control cells (all
P<0.01; Fig. 4A-D).
Simultaneously, the level of apoptotic cells was detected via the
TUNEL assay at day 4. No significant differences in cell death were
demonstrated between the four groups (data not shown). Cell cycle
analysis was performed via flow cytometry, in order to determine
whether TMOD1 regulates cell proliferation by altering the cell
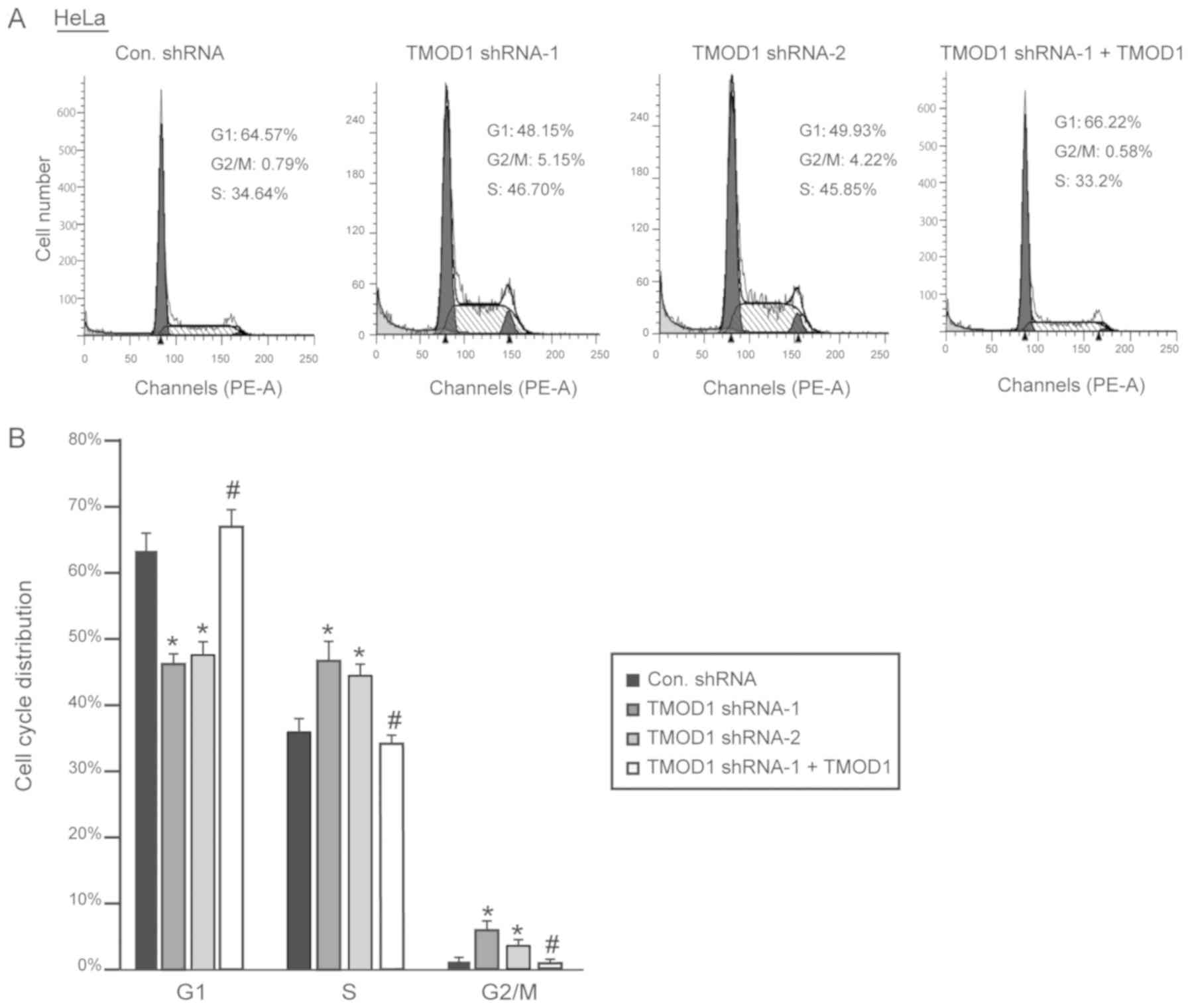
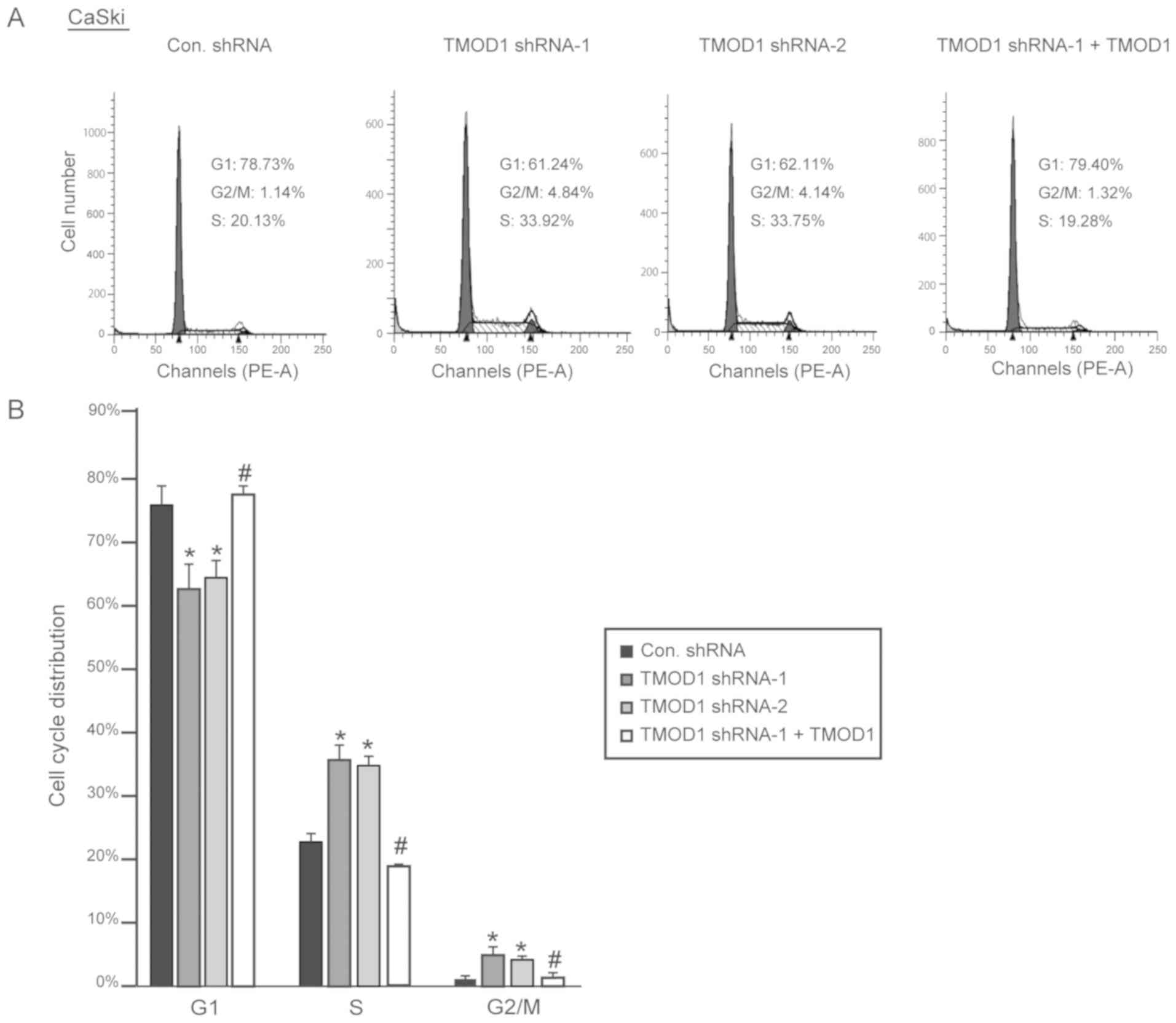
cycle. The results demonstrated that downregulation of TMOD1
promoted G1/S phase transition in HeLa and CaSki cells,
and the percentage of cells in the G2/M phase was
significantly increased, while shRNA-resistant TMOD1 expression in
TMOD1 shRNA treated cells was demonstrated to rescue the phenomenon
induced by TMOD1 shRNA-1 (all P<0.05; Figs. 5 and 6).
Low TMOD1 expression is associated
with worse pathological status
Whether or not the in vitro data were
relevant to the clinical features of human cervical cancer was
subsequently determined. TMOD1 expression was analyzed in different
pathological stages using the R2 database (http://r2.amc.nl), which is a free, and publicly
accessible web-based genomics analysis and visualization platform
that allows researchers to integrate, analyze and visualize
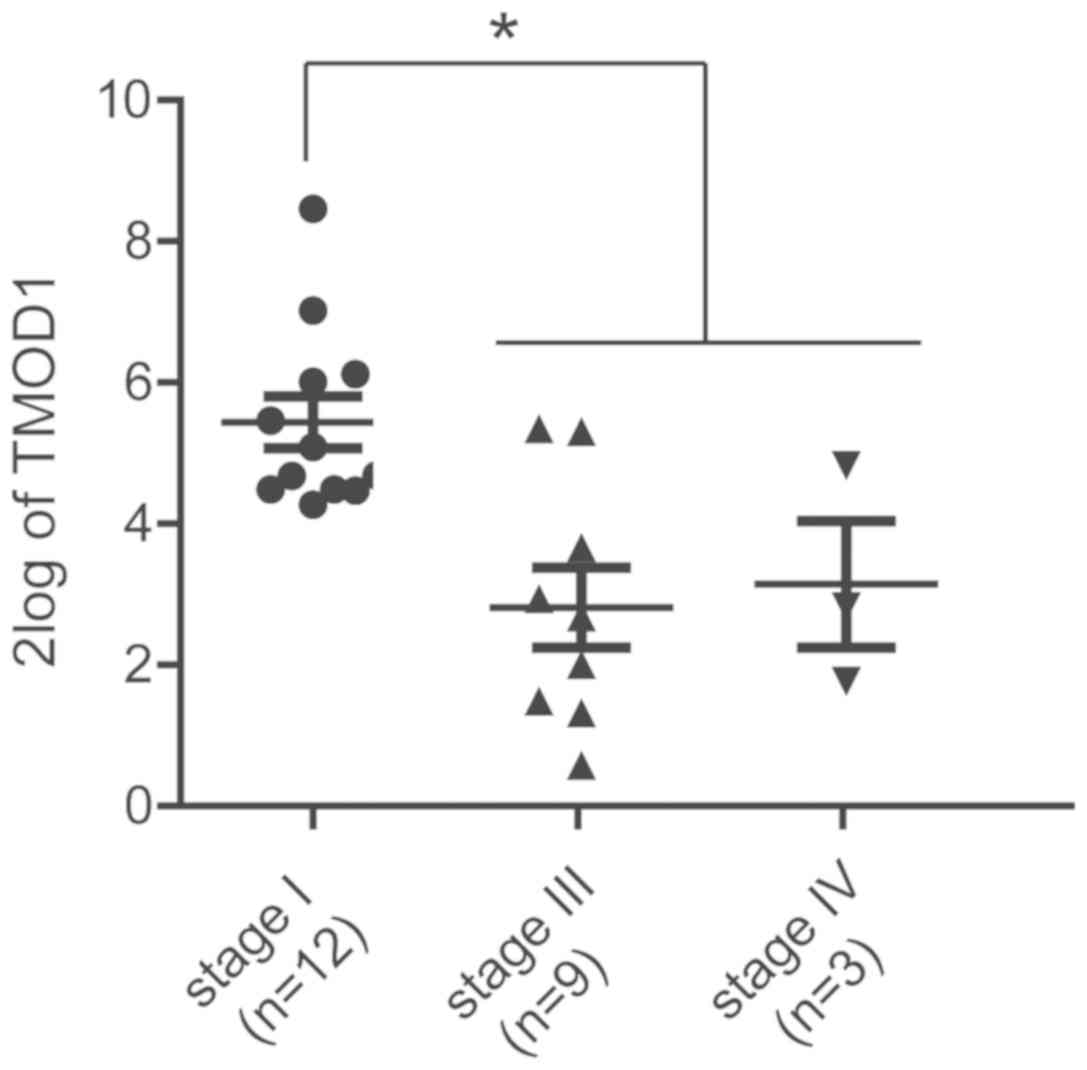
clinical and genomics data. High TMOD1 expression was observed in
patients in the early pathological stages, while low TMOD1
expression was presented in patients in the late stages (P<0.05;
Fig. 7). Patients within the Stage
II group was excluded due to lack of an adequate sample size
required for statistical analysis. In all, these results were
consistent with the in vitro data, further suggesting that
TMOD1 may act as a tumor suppressor gene in human cervical
cancer.
Discussion
Actin is one of the multi-functional proteins in
eukaryotic cells, which can polymerize into filamentary actin and
also depolymerize into monomer actin (22). Depending on the dynamics of
filamentary actin, actin regulates several physiological
activities, such as regulating the subcellular localization of p53
(22) and regulating gene
transcription (23). Filamentary
actin has two ends, the barded end and the pointed end. Thus far,
TMODs are the only known molecules that have the ability to cap the
pointed end of filamentary actin. Capping by TMOD1 inhibits the
release of free monomer actin from the pointed end, therefore
prevents filamentary actin renewal and elongation (24). The tight association between actin
and TMOD1 led researchers to focus on the function of TMOD1 on cell
morphology (25,26), motility (10) and metastasis (15) or neuronal growth (9). It has been reported that TMOD1
knockdown in N2a cells increases the number of neuritis/cell and
decreases the mean primary neurite length (9).
Despite its extensive roles within the nervous
system, a number of previous reports have demonstrated that TMOD1
promotes cancer cell proliferation and metastasis by regulating
filamentary actin, which indicates that TMOD1 may act as an
oncogene in certain types of cancer (13–15,27).
However, the results of the present study indicated that
downregulation of TMOD1 promoted cell migration and proliferation
in both HeLa and CaSki cell lines. Furthermore, the results from
the clinical database suggested that low TMOD1 expression is
associated with relative higher pathological grading and staging,
while high expression is associated with relative lower
pathological grading and staging in patients with human cervical
cancer. Taken together, these results indicated that TMOD1 may act
as a tumor suppressor in human cervical cancer, which seems to vary
in its role across different types of cancer.
Cell cycle analysis demonstrated that downregulation
of TMOD1 increased the percentage of cells in the S phase; however,
it decreased the number of cells in the G1 phase.
Despite the lack of current data on the association between TMOD1
and cell cycle progression, the notable roles of actin dynamics in
cell cycle progress have been broadly researched (28,29).
Disruption of filamentary actin dynamics with drugs, including
cytochalasin D (inhibits actin monomer assembly by binding to the
barbed ends of microfilaments), latrunculin B (prevents actin
polymerization by binding to actin monomers near their ATP-binding
site) and Jasplakinolide (stabilizes and promotes actin
polymerization by binding to F-actin), has been reported to result
in cell cycle arrest (30–32). Actin and several actin binding
proteins, including Myosin II and septin have been demonstrated to
interact with cyclins and cyclin-dependent kinases in different
phases of the cell cycle (28).
Furthermore, Sui et al (33)
demonstrated that TMOD3 depletion increases the number of cells in
the S phase, while fewer cells were observed in the
G0/G1 phase in TMOD3−/− fetal
liver tissue. Among the four TMODs, the amino acid sequences of
their two major function domains (the TM-Cap and the LRR-Cap
domains) have a 70–80% and 80–90% similarity, respectively
(34). Considering the similar
molecular structure and biological functions between TMOD1 and
TMOD3, TMOD1 may also be involved in cell cycle regulation;
however, its underlying molecular mechanism remains unclear.
The wound-healing assay results demonstrated that
cell migration was notably enhanced following TMOD1 depletion.
Conversely, the role of TMOD1 in cell invasion and EMT was assessed
via a Matrigel invasion assay, and analysis of Twist and Snail
expression levels, respectively. However, no significant
differences were demonstrated between control cells and TMOD1
shRNA-treated cells in these experiments. Overall, TMOD1 expression
was demonstrated to inhibit cell migration, but not invasion.
Increasing evidence suggests that the cyclins which
modulate G1/S transition also promote cell migration.
For instance, cyclin D1 has been reported to promote cell migration
by stimulating the activity of Rac1 and Ral GTPases (35), and cyclin E2 has been demonstrated to
enhance cell proliferation and migration of non-small cell lung
cancer (36). The results of the
present study demonstrated that downregulation of TMOD1 promoted
G1/S transition, which suggests that TMOD1 may regulate
the cell cycle by affecting the expression levels of G1
cyclins. Future studies will focus on investigating the
associations between TMOD1, G1 cyclins and cell migration.
Acknowledgements
The authors of the present study would like to thank
Dr. Peng Zou from Shenyang Medical College (Shenyang, China) for
his general support.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FL and PM designed the present study. FL, DC and LZ
acquired the data, while FL and BM analyzed and interpreted the
data. FL and PM drafted the initial manuscript and made substantial
revisions. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Pimple S, Mishra G and Shastri S: Global
strategies for cervical cancer prevention. Curr Opin Obstet
Gynecol. 28:4–10. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
GLOBOCAN 2012, . Estimated cancer
incidence, mortality and prevalence worldwide in 2012.
International Agency for Research on Cancer, World Health
Organization.
|
|
3
|
Small W Jr, Bacon MA, Bajaj A, Chuang LT,
Fisher BJ, Harkenrider MM, Jhingran A, Kitchener HC, Mileshkin LR,
Viswanathan AN and Gaffney DK: Cervical cancer: A global health
crisis. Cancer. 123:2404–2412. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lippman SM and Hawk ET: Cancer prevention:
From 1727 to milestones of the past 100 years. Cancer Res.
69:5269–5284. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu Y, Yu J, Qian L, Zhang H and Ma J:
Extended field intensity-modulated radiotherapy plus concurrent
nedaplatin treatment in cervical cancer. Oncol Lett. 11:3421–3427.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Duenas-Gonzaleza A, Cetina L, Coronel J
and Gonzalez-Fierro A: The safety of drug treatments for cervical
cancer. Expert Opin Drug Saf. 15:169–180. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fischer RS and Fowler VM: Tropomodulins:
Llife at the slow end. Trends Cell Biol. 13:593–601. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gokhin DS and Fowler VM: Cytoplasmic
gamma-actin and tropomodulin isoforms link to the sarcoplasmic
reticulum in skeletal muscle fibers. J Cell Biol. 194:105–120.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fath T, Fischer RS, Dehmelt L, Halpain S
and Fowler VM: Tropomodulins are negative regulators of neurite
outgrowth. Eur J Cell Biol. 90:291–300. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fischer RS, Fritz-Six KL and Fowler VM:
Pointed-end capping by tropomodulin3 negatively regulates
endothelial cell motility. J Cell Biol. 161:371–380. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nowak RB, Papoin J, Gokhin DS, Casu C,
Rivella S, Lipton JM, Blanc L and Fowler VM: Tropomodulin 1
controls erythroblast enucleation via regulation of F-actin in the
enucleosome. Blood. 130:1144–1155. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rho SB, Chun T, Lee SH, Park K and Lee JH:
The interaction between E-tropomodulin and thymosin beta-10 rescues
tumor cells from thymosin beta-10 mediated apoptosis by restoring
actin architecture. FEBS Lett. 557:57–63. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ting SB, Deneault E, Hope K, Cellot S,
Chagraoui J, Mayotte N, Dorn JF, Laverdure JP, Harvey M, Hawkins
ED, et al: Asymmetric segregation and self-renewal of hematopoietic
stem and progenitor cells with endocytic Ap2a2. Blood.
119:2510–2522. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ito-Kureha T, Koshikawa N, Yamamoto M,
Semba K, Yamaguchi N, Yamamoto T, Seiki M and Inoue J: Tropomodulin
1 expression driven by NF-κB enhances breast cancer growth. Cancer
Res. 75:62–72. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Suzuki T, Kasamatsu A, Miyamoto I, Saito
T, Higo M, Endo-Sakamoto Y, Shiiba M, Tanzawa H and Uzawa K:
Overexpression of TMOD1 is associated with enhanced regional lymph
node metastasis in human oral cancer. Int J Oncol. 48:607–612.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gharahkhani P, Fitzgerald RC, Vaughan TL,
Palles C, Gockel I, Tomlinson I, Buas MF, May A, Gerges C, Anders
M, et al: Genome-wide association studies in oesophageal
adenocarcinoma and Barrett's oesophagus: A large-scale
meta-analysis. Lancet Oncol. 17:1363–1373. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lu F, Zheng Y, Donkor PO, Zou P and Mu P:
Downregulation of CREB promotes cell proliferation by mediating
G1/S phase transition in hodgkin lymphoma. Oncol Res. 24:171–179.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang CG, Huang JC, Liu T and Li XY:
Anticancer effects of bishydroxycoumarin are mediated through
apoptosis induction, cell migration inhibition and cell cycle
arrest in human glioma cells. J BUON. 20:1592–1600. 2015.PubMed/NCBI
|
|
20
|
Schneider CA, Rasband WS and Eliceiri KW:
NIH Image to ImageJ: 25 years of image analysis. Nature Methods.
9:671–675. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mu P, Nagahara S, Makita N, Tarumi Y,
Kadomatsu K and Takei Y: Systemic delivery of siRNA specific to
tumor mediated by atelocollagen: Combined therapy using siRNA
targeting Bcl-xL and cisplatin against prostate cancer. Int J
Cancer. 125:2978–2990. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Saha T, Guha D, Manna A, Panda AK, Bhat J,
Chatterjee S and Sa G: G-actin guides p53 nuclear transport:
Potential contribution of monomeric actin in altered localization
of mutant p53. Sci Rep. 6:326262016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sadhukhan S, Sarkar K, Taylor M, Candotti
F and Vyas YM: Nuclear role of WASp in gene transcription is
uncoupled from its ARP2/3-dependent cytoplasmic role in actin
polymerization. J Immunol. 193:150–160. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Weber A, Pennise CR, Babcock GG and Fowler
VM: Tropomodulin caps the pointed ends of actin filaments. J Cell
Biol. 127:1627–1635. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Moyer JD, Nowak RB, Kim NE, Larkin SK,
Peters LL, Hartwig J, Kuypers FA and Fowle VM: Tropomodulin 1-null
mice have a mild spherocytic elliptocytosis with appearance of
tropomodulin 3 in red blood cells and disruption of the membrane
skeleton. Blood. 116:2590–2599. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nowak RB, Fischer RS, Zoltoski RK, Kuszak
JR and Fowler VM: Tropomodulin1 is required for membrane skeleton
organization and hexagonal geometry of fiber cells in the mouse
lens. J Cell Biol. 186:915–928. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Agnelli L, Mereu E, Pellegrino E, Limongi
T, Kwee I, Bergaggio E, Ponzoni M, Zamò A, Iqbal J, Piccaluga PP,
et al: Identification of a 3-gene model as a powerful diagnostic
tool for the recognition of ALK-negative anaplastic large-cell
lymphoma. Blood. 120:1274–1281. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Heng YW and Koh CG: Actin cytoskeleton
dynamics and the cell division cycle. Int J Biochem Cell Biol.
42:1622–1633. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lee YJ and Keng PC: Studying the effects
of actin cytoskeletal destabilization on cell cycle by cofilin
overexpression. Mol Biotechnol. 31:1–10. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Moulding DA, Blundell MP, Spiller DG,
White MR, Cory GO, Calle Y, Kempski H, Sinclair J, Ancliff PJ,
Kinnon C, et al: Unregulated actin polymerization by WASp causes
defects of mitosis and cytokinesis in X-linked neutropenia. J Exp
Med. 204:2213–2224. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lee K and Song K: Actin dysfunction
activates ERK1/2 and delays entry into mitosis in mammalian cells.
Cell Cycle. 6:1487–1495. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gachet Y, Tournier S, Millar JB and Hyams
JS: A MAP kinase-dependent actin checkpoint ensures proper spindle
orientation in fission yeast. Nature. 412:352–355. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sui Z, Nowak RB, Bacconi A, Kim NE, Liu H,
Li J, Wickrema A, An XL and Fowler VM: Tropomodulin3-null mice are
embryonic lethal with anemia due to impaired erythroid terminal
differentiation in the fetal liver. Blood. 123:758–767. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yamashiro S, Gokhin DS, Kimura S, Nowak RB
and Fowler VM: Tropomodulins: Pointed-end capping proteins that
regulate actin filament architecture in diverse cell types.
Cytoskeleton (Hoboken). 69:337–370. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pedraza N, Cemeli T, Monserrat MV, Garí E
and Ferrezuelo F: Regulation of small GTPase activity by G1
cyclins. Small GTPases. 10:47–53. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Vannini I, Wise PM, Challagundla KB,
Plousiou M, Raffini M, Bandini E, Fanini F, Paliaga G, Crawford M,
Ferracin M, et al: Transcribed ultraconserved region 339 promotes
carcinogenesis by modulating tumor suppressor microRNAs. Nat
Commun. 8:18012017. View Article : Google Scholar : PubMed/NCBI
|