Introduction
Liver cancer is the sixth most common cancer and is
the second leading cause of cancer-associated mortality worldwide
in 2018 (1,2). Chemotherapy is one of the most common
treatment methods for liver cancer (3,4);
however, it is not very efficacious in certain patients (5). Currently, transarterial
chemoembolization (TACE) is the preferred therapy for patients with
advanced hepatocellular carcinoma (HCC) (6). Adriamycin (ADR), also known as
doxorubicin, is the first-line chemotherapeutic drug used in TACE
(7). However, the prognosis of
patients with HCC remains poor due to inherent or acquired
chemoresistance to Adriamycin (8,9).
Understanding the molecular mechanisms underlying ADR resistance in
liver cancer may result in improved liver cancer prognosis and the
development of suitable therapeutic targets to overcome
chemoresistance.
Prolyl 4-hydroxylase beta polypeptide (P4HB)
is the core member of the protein disulfide isomerase gene family,
and can serve as an endoplasmic reticulum chaperone to inhibit the
aggregation of misfolded proteins (10). Previous reports have demonstrated
that overexpression of P4HB promotes liver cancer progression via
the upregulation of epithelial-mesenchymal transition (EMT)
(11), which is closely associated
with drug-resistance in malignant gliomas (12), glioblastoma multiforme (13) and non-small cell lung cancer
(14). However, whether P4HB
regulates drug-resistance in liver cancer is unknown.
EMT was initially considered to be an important
physiological process in tissue differentiation and organogenesis
during embryonic development (15).
Previous studies have revealed that EMT is closely associated with
drug-resistance and tumor metastasis (16,17). EMT
occurs in both gemcitabine-resistant pancreatic adenocarcinoma cell
lines (such as MiaPaCa-2, Panc-1 and Aspc-1) (18) and adriamycin-induced drug-resistant
breast cancer cells (such as MCF7) (19). Snail and β-catenin are two of the
several known regulators of EMT-associated. Snail is a zinc finger
transcription factor and an important regulator in tumor
progression, which can promote tumor invasion and metastasis
(20). Overexpression of Snail can
promote epithelial mesenchymal transformation and the invasion and
migration of breast cancer cells (21). Snail-induced EMT is partly due to the
direct repression of E-cadherin transcription both during
development and tumour progression (22). β-catenin serves a key role in
regulating cell proliferation and differentiation (23,24).
Epithelial integrity requires the stability of E-cadherin/β-catenin
complexes (25). Previous studies
have reported that Snail, β-catenin and EMT contribute to tumor
chemotherapeutic resistance to sorafenib (26) and cisplatin (27–29).
In the present study, the expression of P4HB was
measured in drug-resistant liver cancer cells and its effects on
invasion, migration and chemoresistance were investigated. The aim
was to investigate the therapeutic value of targeting P4HB
for liver cancer therapy.
Materials and methods
Cell culture
The liver cancer cell line HepG2 was purchased from
The Cell Bank of Type Culture Collection of the Chinese Academy of
Sciences. Dulbecco's modified Eagle's medium (DMEM; Invitrogen;
Thermo Fisher Scientific, Inc.), supplemented with 10% fetal bovine
serum (FBS; Invitrogen; Thermo Fisher Scientific, Inc.), 100 U/ml
penicillin (Invitrogen; Thermo Fisher Scientific, Inc.) and 100
µg/ml streptomycin (Invitrogen; Thermo Fisher Scientific, Inc.),
was used as the cell culture medium. Cells were cultured in a
humidified chamber containing 5% CO2 at 37°C. To
establish the HepG2/ADR cell lines, ADR (Selleckchem Chemicals) was
added to HepG2 cells in a stepwise increasing titration from
0.001–0.5 mg/l for 6 months. Resistant cell colonies were
subsequently obtained. Adriamycin resistance was maintained by
culturing the cells at low concentrations of ADR (0.10 mg/l). These
resistant sub-lines were named HepG2/ADR. At least three
independent experiments were performed.
Cell proliferation studies
Cell Counting Kit-8 (CCK-8; Beyotime Biotechnology,
Inc.) assays were used to assess drug sensitivity. HepG2 and
HepG2/ADR cells were seeded into 96-well plates at a density of
3,000 cells/well. HepG2/ADR cells were cultured in fresh medium
containing increased concentrations of adriamycin (from 0–8 µM)
substituted medium and incubated at 37°C with 5% CO2 for
48 h. CCK-8 assay was performed according to the manufacturer's
protocols. Briefly, each well was treated with 10 ul CCK-8 reagent.
After incubating at 37°C for additional 2 h, the optical density at
a wavelength of 450 nm was determined using a Spectramax M5
microplate reader (Molecular Devices, LLC). Each assay was
performed with 5 replications.
Transwell migration and invasion
assays
The Transwell chambers used for the migration assay
contained polycarbonate filters with a 8-µm pore size (BD
Biosciences). DMEM containing 10% FBS was placed in the lower
chambers. HepG2 and HepG2/ADR cells (1×105 in 500 µl
serum-free DMEM) were seeded onto the upper chamber and incubated
at 37°C for 24 h. Cells that had migrated to the lower chambers
were fixed with 70% methanol and then stained with 0.1% crystal
violet at room temperature, and imaged using an Olympus ix 71 light
microscope at ×100 magnification (Olympus Corporation, Inc.).
In order to measure cell invasion, 1×105
HepG2 and HepG2/ADR cells were serum-starved overnight and seeded
onto the upper chamber precoated with Matrigel (for 30 min at
37°C). The cells that penetrated the Matrigel-coated filter were
fixed, stained, and counted as aforementioned, and subsequently
five cell fields were randomly selected from each membrane to count
using an Olympus ix 71 light microscope at ×100 magnification
(Olympus Corporation, Inc.). Each assay was performed in
triplicate. At least three independent experiments were
performed.
Wound healing assay
HepG2 and HepG2/ADR cells were seeded onto 6-well
plates at a density of 5×105 cells/well and cultured in
a humidified chamber containing 5% CO2 at 37°C for one
day. After the cells reached 100% confluence, a line was scraped at
the center of each well using a 200-ul pipette tip. Cellular debris
was then carefully removed by washing with PBS three times.
Subsequently, cells were cultured in serum-free medium at 5%
CO2 and 37°C for 24 h. The images of the confluent cells
were captured using an Olympus ix 71 light microscope at ×100
magnification (Olympus Corporation, Inc.) to determine the wound
width at time 0 and 24 h, respectively. Wound healing was
visualized by comparing the images taken at 0 and 24 h, and
analyzing the migration distance using the leading edge of the
wound at each time point for all the treatment groups. The relative
wound width was calculated as wound width at 24 h divided by wound
width at the 0 h time point. At least three independent experiments
were performed.
Western blotting
After treatment, total cellular protein from HepG2
and HepG2/ADR cells was extracted using RIPA buffer containing 1 mM
phenylmethanesulfonyl fluoride (Aidlab Biotechnologies Co., Ltd.).
Cell lysates were collected, and protein concentrations were
determined using the bicinchoninic acidprotein assay kit (Beyotime
Biotechnology, Inc.). Cell lysates (40 µg total protein and 40 µg
nuclear protein) were subsequently separated by SDS-PAGE (10% gel)
and transferred onto PVDF membranes (Bio-Rad Laboratories, Inc.).
Next, 5% fat-free milk was used to block the membrane at 4°C for 1
h. Membranes were then incubated overnight with different primary
antibodies including anti-P4HB (1:1,000; cat. no. ab137110; Abcam),
anti-E-cadherin (1:1,000; cat. no. 3195; Cell Signaling Technology,
Inc.), anti-N-cadherin (1:1,000; cat. no. 4061; Cell Signaling
Technology, Inc.), anti-vimentin (1:1,000; cat. no. 5741; Cell
Signaling Technology, Inc.), anti-Snail (1:1,000; cat. no. 3879;
Cell Signaling Technology, Inc.), anti-β-catenin (1:1,000; cat. no.
8480; Cell Signaling Technology, Inc.), anti-β-actin (1:1,000; cat.
no. 4970; Cell Signaling Technology, Inc.) or anti-histone H3
(1:1,000; cat. no. ab1791; Abcam) at 4°C. β-Actin and histone
proteins were used as the internal controls for total protein and
nuclear protein, respectively. After washing, the membranes were
incubated with the corresponding HRP-linked secondary antibody
(1:5,000; cat. no. 7074; Cell Signaling Technology, Inc.) at room
temperature for 1 h. Protein bands were visualized with an enhanced
chemiluminescent reagent (GE Healthcare). Band intensity was
measured using the gel imaging system (ProteinTech Group, Inc.) and
analyzed using the FluorChem FC3 software (Proteinsimple, Inc.). At
least three independent experiments were performed.
Reverse transcription-quantitative
(RT-qPCR)
Total RNA was isolated from HepG2 and HepG2/ADR
liver cancer cells using TRIzol® reagent (Invitrogen;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
instructions. RNA concentration was determined using the NanoDrop
2000 spectrophotometer (Thermo Fisher Scientific, Inc.). According
to the manufacturer's instructions, a total of 1 µg RNA was
reverse-transcribed (at 42°C for 1 h; and at 70°C for 10 min) using
the First-Strand cDNA Synthesis kit (Invitrogen; Thermo Fisher
Scientific, Inc.). qPCR was performed using a SYBR green qPCR
SuperMix-UDG kit (Life Technologies; Thermo Fisher Scientific,
Inc.) on an ABI 7500 system (Applied Biosystems; Thermo Fisher
Scientific, Inc.) to determine the expression levels of the target
mRNAs in accordance with the manufacturer's instructions (95°C for
10 sec; 60°C for 15 sec; 72°C for 15 sec; 45 cycles).
Relative mRNA expression levels were calculated
using GAPDH as the internal control. Each sample was run in
triplicate. The primer pairs used were as follows: P4HB forward,
5′-GGAATGGAGACACGGCTTC-3′ and reverse, 5′-TTCAGCCAGTTCACGATGTC-3′;
and β-actin forward, 5′-AGCGCGGCTACAGCTTCA-3′, and reverse,
5′-GGCCATCTCTTGCTCGAAGT-3′. The gene expression levels for all
samples were normalized to GAPDH expression using the
2−ΔΔCq method (30). At
least three independent experiments were performed.
Cell transfection
Small interfering RNA (siRNA) specific for human
P4HB (5′-AAGATGAACTGTAATACGCAA-3′) and a scrambled siRNA
(5′-UUCUCCGAACGUGUCACGUTT-3′), which was used as the negative
control (NC), were designed and synthesized by Shanghai GenePharma
Co., Ltd. HepG2/ADR cells were seeded at a density of
3×105 cells/ml onto 6-well plates and transfected with 2
ug control siRNA and P4HB siRNA using Lipofectamine®
2000 (Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol for 48 h. At least three independent
experiments were performed.
Statistical analysis
Data are presented as mean ± SD. Data was analyzed
using the SPSS 21.0 software (IBM, Corp.). The Mann-Whitney U test
was performed due to abnormal distribution or heterogeneity of
variance. Additionally, the Student's t-test or one-way analysis of
variance with Dunnett's post-hoc test was used for normally
distributed data. P<0.05 was considered to indicate a
statistically significant difference.
Results
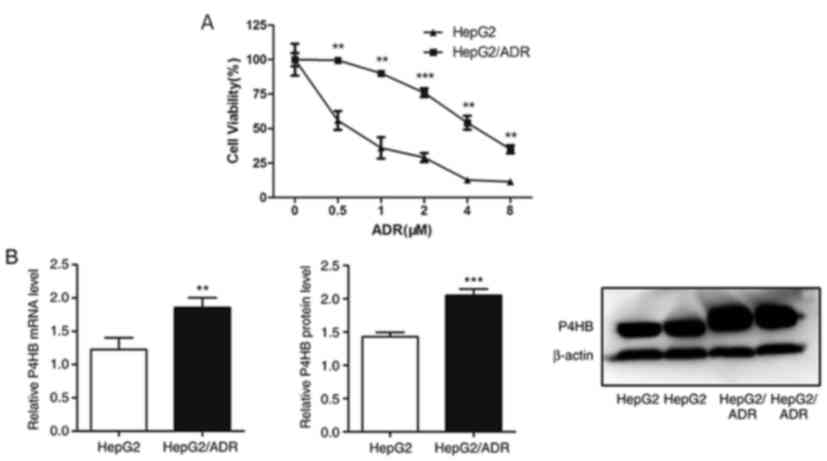
P4HB expression levels are upregulated
in chemoresistant liver cancer sub-line HepG2/ADR
In order to investigate the possible mechanisms
underlying chemoresistance in liver cancer, a liver cancer sub-line
that is resistant to ADR was established. The liver cancer cell
line HepG2/ADR was more resistant to ADR compared with its parental
cell line (Fig. 1A). The half
maximal inhibitory concentration (IC50) of HepG2/ADR and
HepG2 cell lines resistant to ADR were 4.85 and 0.61 µM,
respectively. The data also revealed that P4HB mRNA and protein
levels were higher in HepG2/ADR cells compared with HepG2 cells
(P<0.01; Fig. 1B). This indicates
that P4HB may serve an important role in liver cancer
chemoresistance.
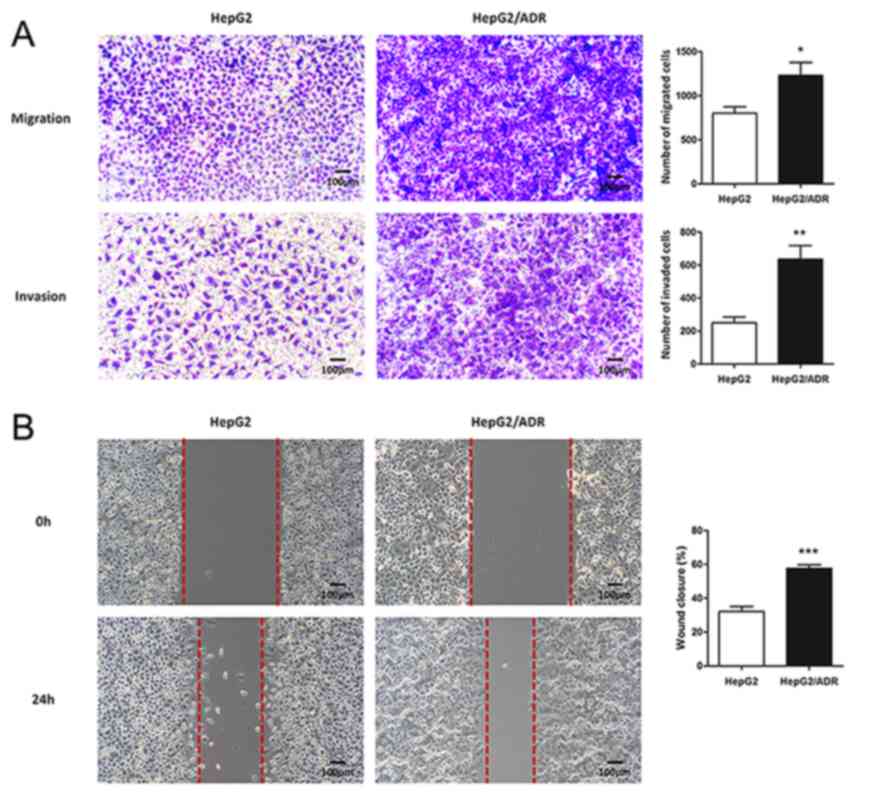
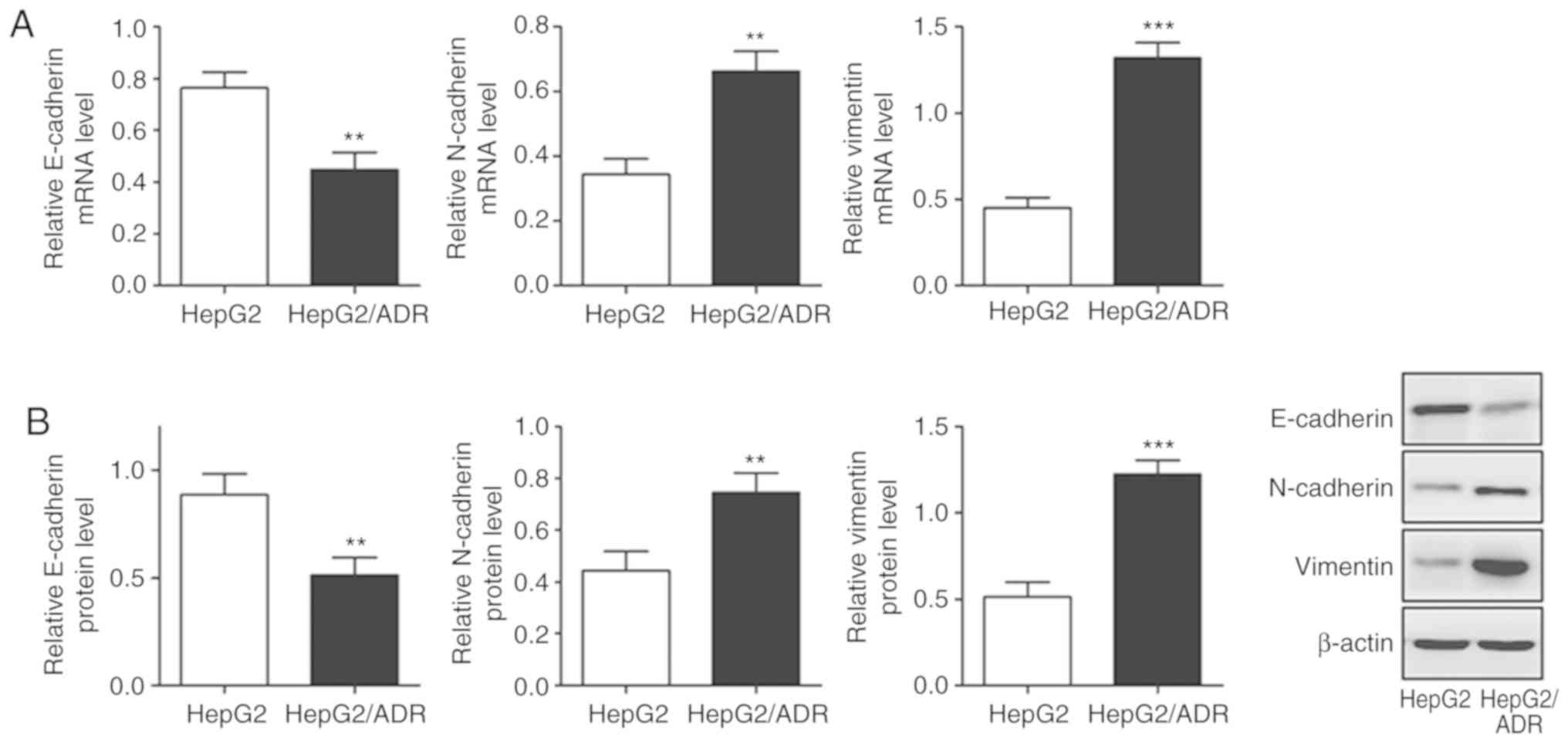
Chemoresistant liver cancer subline
exhibits an EMT phenotype with high migration and invasion
abilities
To determine the migration and invasion ability of
HepG2/ADR cells, Transwell and wound healing assays were performed.
HepG2/ADR cells had significantly increased numbers of cells with
migratory and invasive ability after 24 h, compared with HepG2
cells (P<0.05; Fig. 2A and B),
indicating that HepG2/ADR cells acquired enhanced migration and
invasion ability. To determine whether HepG2/ADR cells acquired
specific molecular changes consistent with EMT, the mRNA and
protein expression levels of epithelial adhesion molecule
E-cadherin and mesenchymal markers, including vimentin and
N-cadherin, were measured. It was observed that the expression of
E-cadherin was significantly downregulated in HepG2/ADR cells at
both mRNA and protein levels (P<0.01), whereas the expression
levels of vimentin and N-cadherin were upregulated (P<0.01;
Fig. 3). These results indicate that
EMT was activated in adriamycin-resistant liver cancer cells.
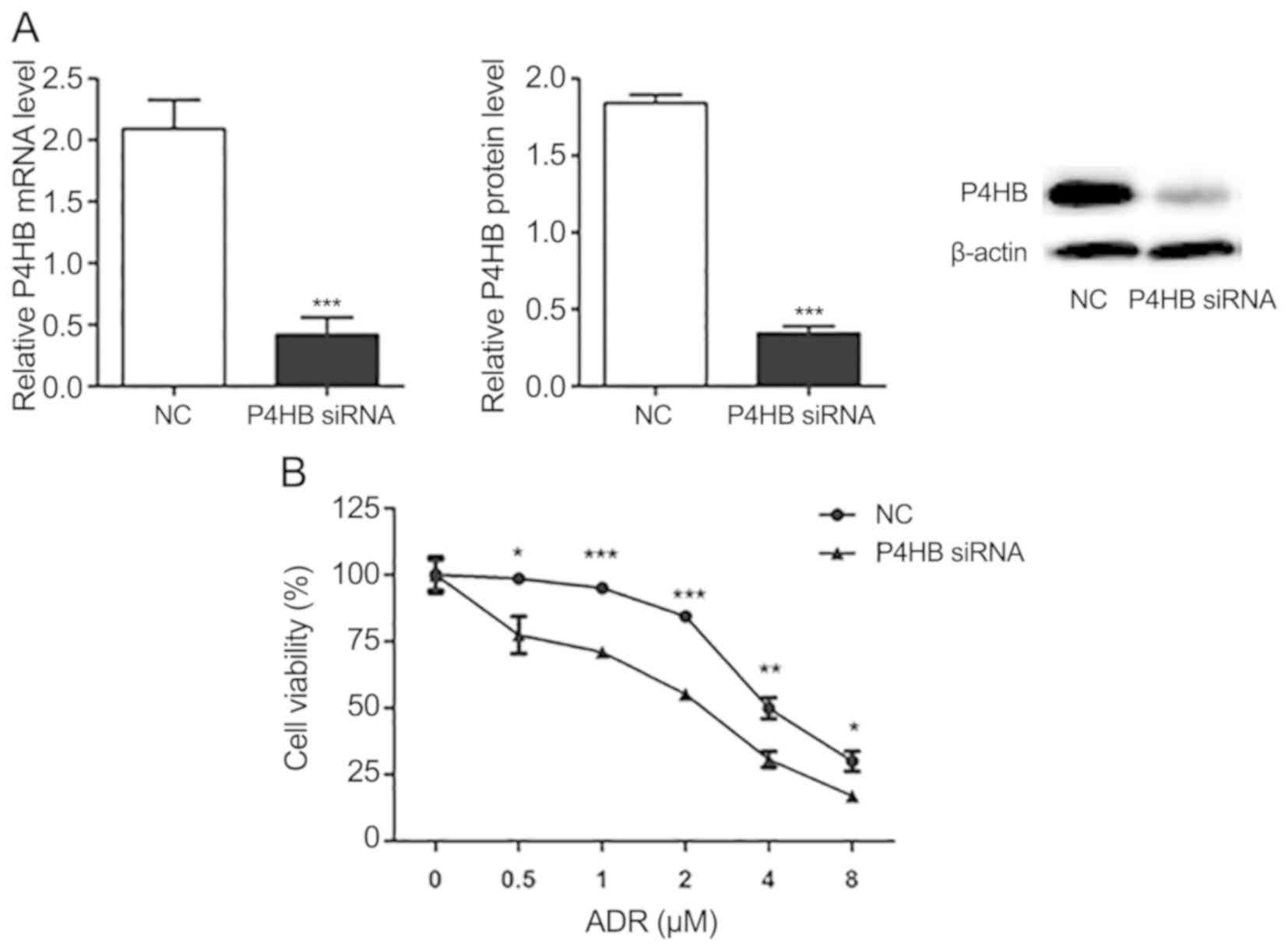
Knockdown of P4HB decreases
drug-resistance in HepG2/ADR cells
Following transfection of HepG2/ADR cells with P4HB
siRNA, the expression levels of P4HB at the mRNA and protein levels
were significantly downregulated (P<0.001; Fig. 4A). Subsequently, the effect of P4HB
inhibition on ADR resistance was investigated in HepG2/ADR cells.
CCK-8 assays demonstrated that ADR was more effective in cells
transfected with P4HB siRNA compared with cells transfected
with NC siRNA. The IC50 of the NC group and the
P4HB siRNA group of ADR resistant cells were 4.64 and 2.05
µM, respectively (Fig. 4B). These
findings suggest that knockdown of P4HB partially reverses
drug-resistance in liver cancer cell lines.
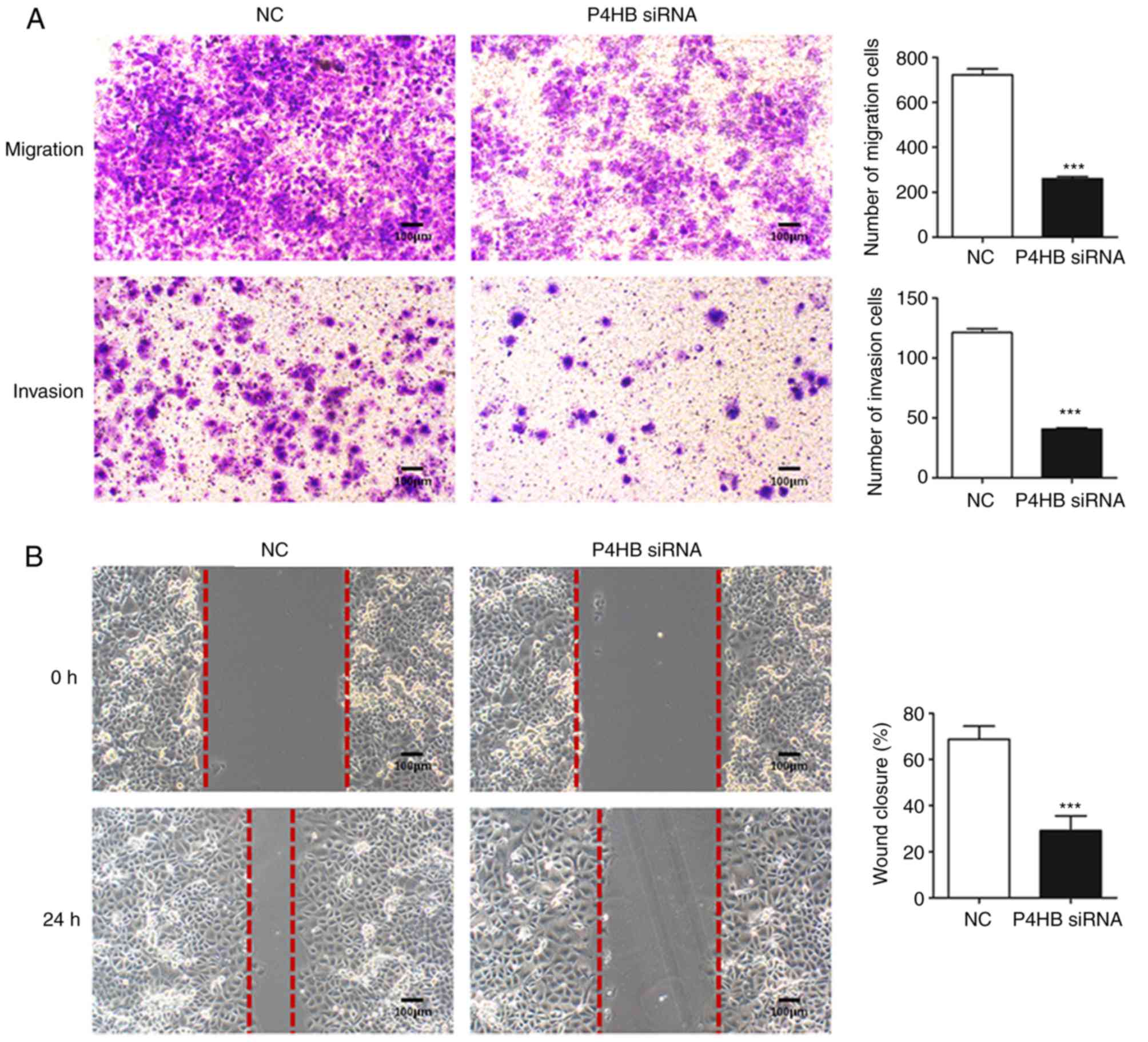
Knockdown of P4HB inhibits the
migration and invasion of HepG2/ADR cells
To further understand the role of P4HB, the
migration and invasive abilities of HepG2/ADR cells transfected
with P4HB siRNA were investigated. Transwell assays
demonstrated that knockdown of P4HB significantly decreased the
migratory and invasive ability of HepG2/ADR cells (P<0.001;
Fig. 5A). Consistent with these
results, P4HB siRNA inhibited cell motility as assessed by
wound healing assays in HepG2/ADR cells (P<0.001; Fig. 5B).
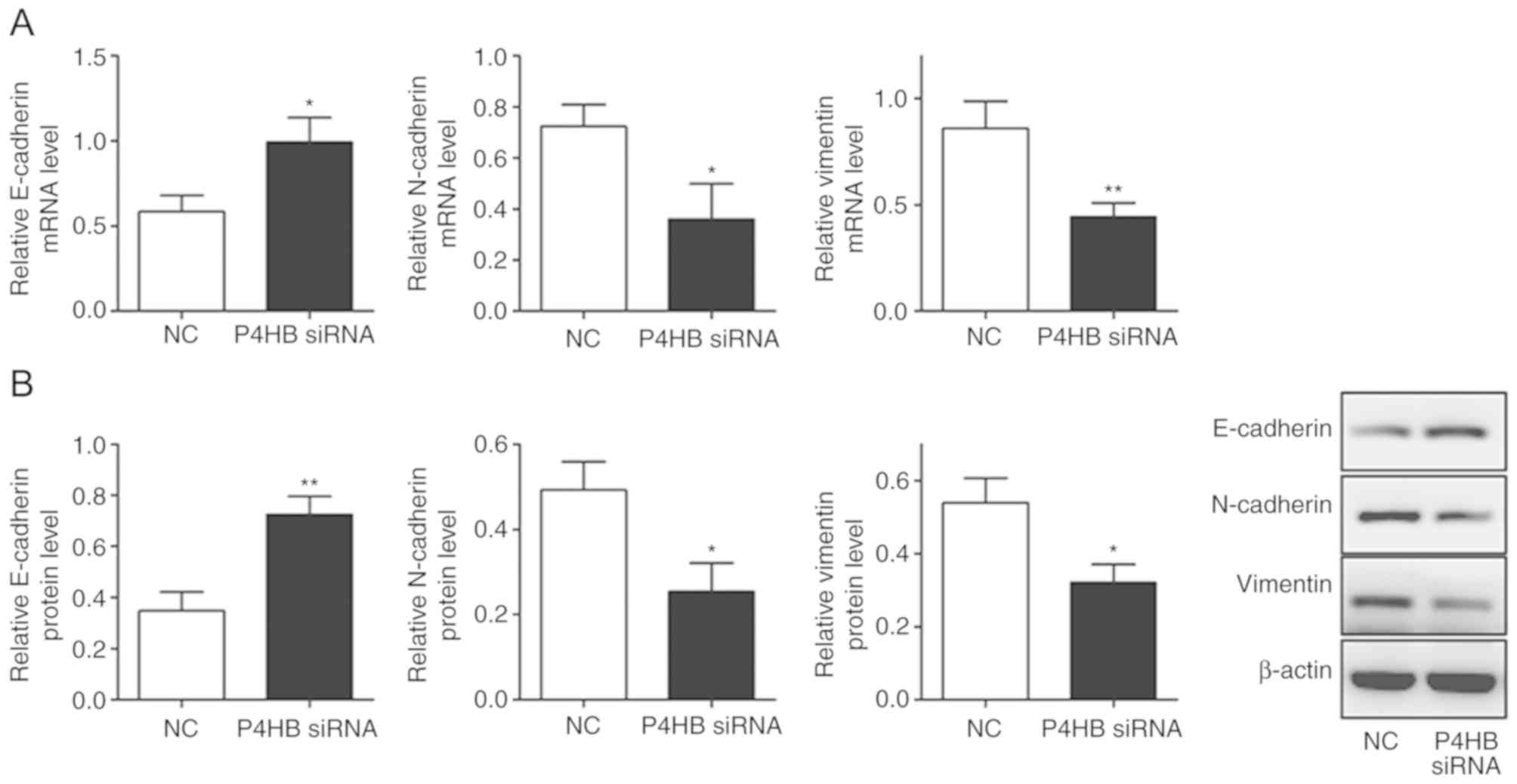
Knockdown of P4HB influences EMT in
HepG2/ADR cells
It was observed that the expression levels of
vimentin and N-cadherin decreased (P<0.05), whereas E-cadherin
levels increased significantly in HepG2/ADR cells transfected with
P4HB siRNA (P<0.05; Fig. 6A,
B). This suggests that the downregulation of P4HB leads to the
inhibition of EMT.
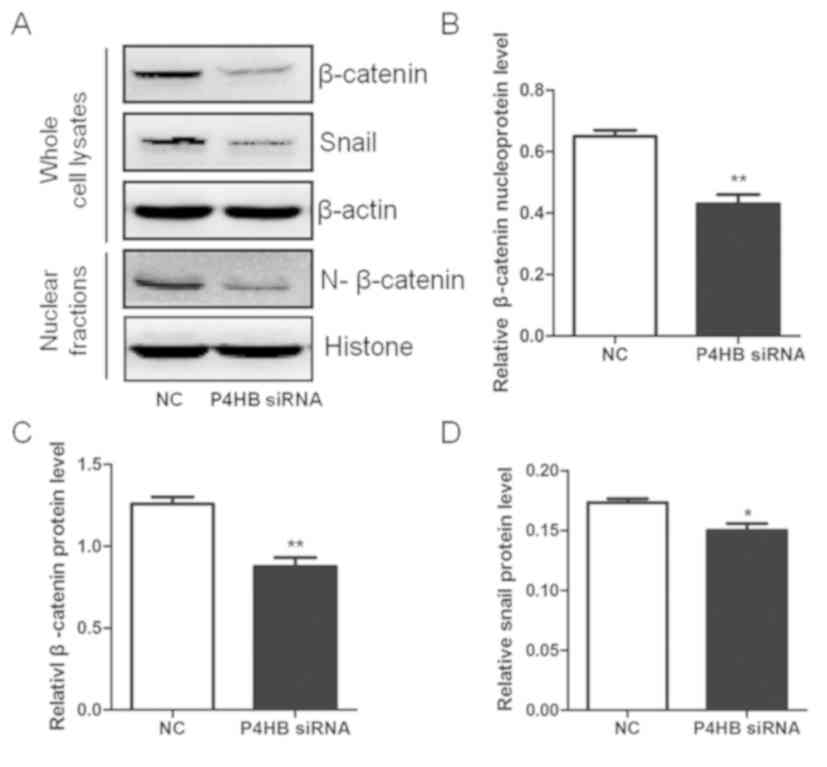
Knockdown of P4HB inhibits the Snail
and β-catenin pathways in HepG2/ADR cells
To investigate the pathway interaction between P4HB,
Snail and β-catenin, the expression level changes in Snail and
β-catenin were measured by western blotting after silencing P4HB
expression in vitro. It was observed that the knockdown of
P4HB significantly decreased the expression of total and nuclear
β-catenin (P<0.01) and downregulated the expression of Snail
(P<0.05) (Fig. 7). This indicates
that P4HB may influence the EMT process via the Snail and β-catenin
pathways (Figs. 7 and 8).
Discussion
Liver cancer is one of the most common malignant
tumors worldwide in 2018 (1,2). Resistance to cytotoxic agents is the
major cause of treatment failure in liver cancer. Several studies
have demonstrated that P4HB is associated with
chemoresistance (14–16). The present study aimed to investigate
whether P4HB influences liver cancer chemotherapy
resistance. It was revealed that P4HB expression was significantly
upregulated in adriamycin-resistant HepG2/ADR cells, compared with
the parental HepG2 cell lines. Silencing P4HB increased the
sensitivity of adriamycin-resistant cells to adriamycin. In
addition, HepG2/ADR cells exhibited increased invasion and
migration abilities, whereas the knockdown of P4HB significantly
decreased cell viability and the number of invasive and migratory
cells. Notably, the knockdown of P4HB inhibited EMT in HepG2/ADR
cells. Overall, the current findings indicate that P4HB knockdown
may enhance the sensitivity of HepG2/ADR cells to ADR, and inhibit
its invasive and migratory ability.
EMT is a complex molecular program that regulates
changes to cell morphology and function during embryogenesis and
tissue development (15). During
EMT, epithelial cells acquire enhanced motility and invasiveness
that are typical of mesenchymal cells. EMT also contributes to
tumor progression and metastasis (31). Emerging evidence suggests that cells
undergoing EMT have increased chemotherapy resistance, and abnormal
activation of genes associated with drug metabolism (19,32).
This indicates that EMT is closely associated with chemotherapy
resistance in tumor cells. Consistent with these findings, the
present study demonstrated the involvement of P4HB in
chemoresistance in adriamycin-resistant HepG2/ADR cells, whereas
P4HB knockdown resulted in reduced EMT and enhanced
chemosensitivity.
Snail is a member of the zinc finger transcription
factor family and is an important regulatory factor in
tumorigenesis which can inhibit gene transcription via competitive
binding to promoter sequences (33–35). The
phenotypic transformation of epithelial cells to mesenchymal cells
results in the occurrence of EMT (36). The data of the present study
demonstrated that the knockdown of P4HB significantly decreased the
expression of Snail in HepG2/ADR cells. β-catenin is the core
component of the Wnt signaling pathway regulates the transcription
of several downstream target genes of Wnt, such as cyclin D1, c-myc
and vimentin, which mediates metastasis and invasion (37). Snail and β-catenin have been reported
to regulate various cellular processes, such as cancer cell
proliferation, apoptosis, invasion, metastases and EMT in
colorectal cancer cells (38). The
present study demonstrated that the knockdown of P4HB significantly
decreased the expression of total and nuclear β-catenin, and
downregulated the expression of Snail. This indicates that
P4HB may influence the EMT process via the β-catenin/Snail
pathway.
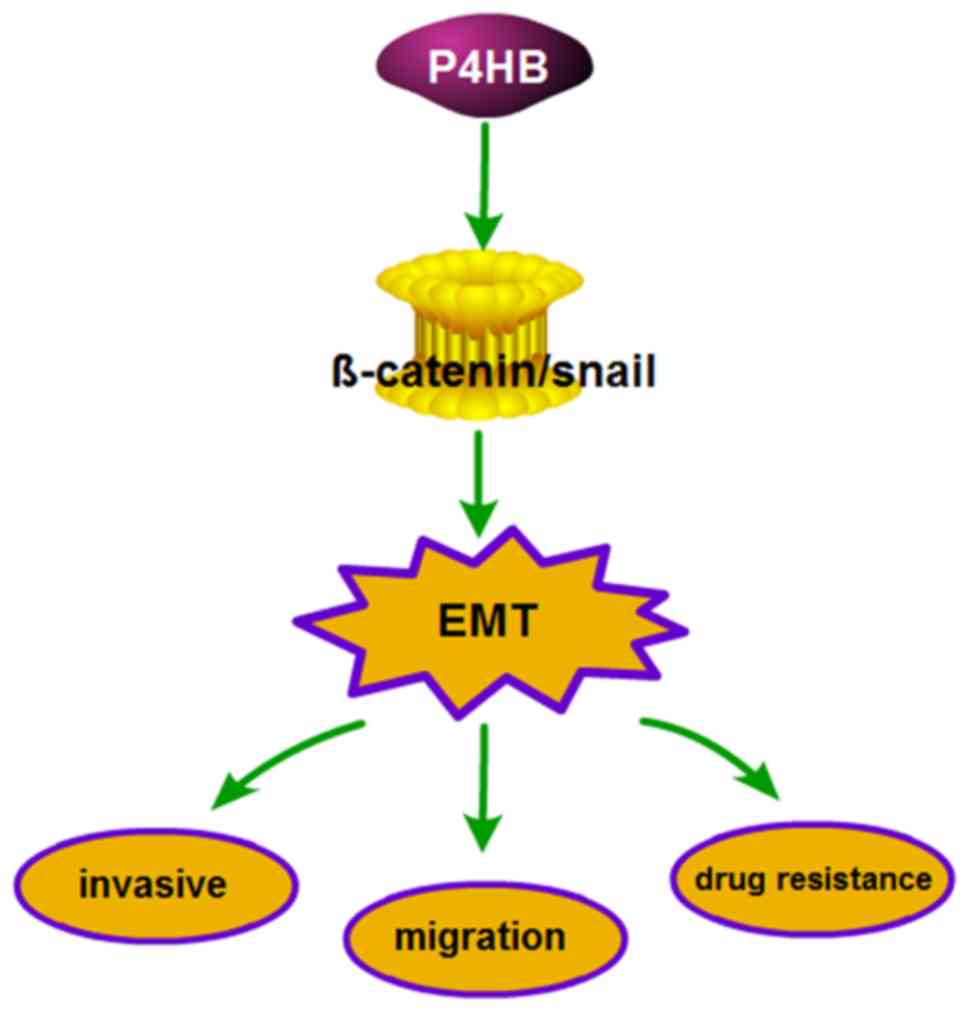
In summary, the present study provides evidence that
P4HB protects HepG2 cells from ADR. Furthermore, the data
demonstrate the role of P4HB in the chemosensitivity, invasion and
migration of HepG2/ADR cells may be mediated via EMT, which is
regulated by the β-catenin/Snail pathway. Thus, P4HB may represent
a novel target to treat liver cancer with acquired ADR resistance.
It is well known that P4HB and its downstream targets may induce
EMT; however, the mechanisms by which P4HB regulates EMT remain to
be deciphered.
Acknowledgements
Not applicable.
Funding
The present study was funded by the National Natural
Science Foundation of China (grant nos. 81703791 and 81873178),
Shanghai Municipal Commission of Health and Family Planning (grant
no. 201740318), Talents Training Program of Seventh People's
Hospital of Pudong Health and Family Planning Commission of
Shanghai (grant no. PWRq2017-02), Science and Technology
Development Fund of Shanghai Pudong New Area (grant no.
PKJ2016-Y50) and Talents Training Program of Seventh People's
Hospital of Shanghai University of Traditional Chinese Medicine
(grant nos. QMX2017-01, XX2017-04 and XX2017-06).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
WX ,YS and JW conceived and designed the study. XM,
JW, JZ, XKM and NZ performed the experiments. WX, YS and XM
analyzed the data and wrote the manuscript. WX, JM, JW, XKM and NZ
reviewed and edited the manuscript. All authors read and approved
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Raoul JL, Kudo M, Finn RS, Edeline J, Reig
M and Galle PR: Systemic therapy for intermediate and advanced
hepatocellular carcinoma: Sorafenib and beyond. Cancer Treat Rev.
68:16–24. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
El Dika I and Abou-Alfa GK: The role (if
any) of chemotherapy in hepatocellular carcinoma. Lancet
Gastroenterol Hepatol. 2:387–389. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhu Q, Li N, Zeng X, Han Q, Li F, Yang C,
Lv Y, Zhou Z and Liu Z: Hepatocellular carcinoma in a large medical
center of China over a 10-year period: Evolving therapeutic option
and improving survival. Oncotarget. 6:4440–4450. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pan LH, Zhao C and Ma YL: Is Y90
radioembolization superior or comparable to transarterial
chemoembolization for treating hepatocellular carcinoma?
Gastroenterology. 152:1627–1628. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Peck-Radosavljevic M: Drug therapy for
advanced-stage liver cancer. Liver Cancer. 3:125–131. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Govaere O, Wouters J, Petz M, Vandewynckel
YP, Van den Eynde K, Van den Broeck A, Verhulst S, Dollé L,
Gremeaux L, Ceulemans A, et al: Laminin-332 sustains
chemoresistance and quiescence as part of the human hepatic cancer
stem cell niche. J Hepatol. 64:609–617. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li Y, Ye Y, Feng B and Qi Y: Long
noncoding RNA lncARSR promotes doxorubicin resistance in
hepatocellular carcinoma via modulating PTEN-PI3K/Akt pathway. J
Cell Biochem. 118:4498–4507. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Noiva R: Protein disulfide isomerase: The
multifunctional redox chaperone of the endoplasmic reticulum. Semin
Cell Dev Biol. 10:481–493. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xia W, Zhuang J, Wang G, Ni J, Wang J and
Ye Y: P4HB promotes HCC tumorigenesis through downregulation of
GRP78 and subsequent upregulation of epithelial-to-mesenchymal
transition. Oncotarget. 8:8512–8521. 2017.PubMed/NCBI
|
|
12
|
Sun S, Wong TS, Zhang XQ, Pu JK, Lee NP,
Day PJ, Ng GK, Lui WM and Leung GK: Protein alterations associated
with temozolomide resistance in subclones of human glioblastoma
cell lines. J Neurooncol. 107:89–100. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lee D, Sun S, Ho AS, Kiang KM, Zhang XQ,
Xu FF and Leung GK: Hyperoxia resensitizes chemoresistant
glioblastoma cells to temozolomide through unfolded protein
response. Anticancer Res. 34:2957–2966. 2014.PubMed/NCBI
|
|
14
|
Wang SM, Lin LZ, Zhou DH, Zhou JX and
Xiong SQ: Expression of prolyl 4-hydroxylase beta-polypeptide in
non-small cell lung cancer treated with Chinese medicines. Chin J
Integr Med. 21:689–696. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Thiery JP and Sleeman JP: Complex networks
orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell
Biol. 7:131–142. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wilson C, Nicholes K, Bustos D, Lin E,
Song Q, Stephan JP, Kirkpatrick DS and Settleman J: Overcoming
EMT-associated resistance to anti-cancer drugs via Src/FAK pathway
inhibition. Oncotarget. 5:7328–7341. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sánchez-Tilló E, Liu Y, de Barrios O,
Siles L, Fanlo L, Cuatrecasas M, Darling DS, Dean DC, Castells A
and Postigo A: EMT-activating transcription factors in cancer:
Beyond EMT and tumor invasiveness. Cell Mol Life Sci. 69:3429–3456.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li Y, VandenBoom TG II, Kong D, Wang Z,
Ali S, Philip PA and Sarkar FH: Up-regulation of miR-200 and let-7
by natural agents leads to the reversal of
epithelial-to-mesenchymal transition in gemcitabine-resistant
pancreatic cancer cells. Cancer Res. 69:6704–6712. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li QQ, Xu JD, Wang WJ, Cao XX, Chen Q,
Tang F, Chen ZQ, Liu XP and Xu ZD: Twist1-mediated
adriamycin-induced epithelial-mesenchymal transition relates to
multidrug resistance and invasive potential in breast cancer cells.
Clin Cancer Res. 15:2657–2665. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Heerboth S, Housman G, Leary M, Longacre
M, Byler S, Lapinska K, Willbanks A and Sarkar S: EMT and tumor
metastasis. Clin Transl Med. 4:62015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Palma Cde S, Grassi ML, Thomé CH, Ferreira
GA, Albuquerque D, Pinto MT, Ferreira Melo FU, Kashima S, Covas DT,
Pitteri SJ and Faça VM: Proteomic analysis of epithelial to
mesenchymal transition (EMT) reveals cross-talk between SNAIL and
HDAC1 proteins in breast cancer cells. Mol Cell Proteomics.
15:906–917. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang Y, Shi J, Chai K, Ying X and Zhou BP:
The Role of Snail in EMT and tumorigenesis. Curr Cancer Drug
Targets. 13:963–972. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Angers S and Moon RT: Proximal events in
Wnt signal transduction. Nat Rev Mol Cell Biol. 10:468–477. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nowicki A, Sporny S and Duda-Szymańska J:
β-catenin as a prognostic factor for prostate cancer (PCa). Cent
European J Urol. 65:119–123. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ghahhari NM and Babashah S: Interplay
between microRNAs and WNT/β-catenin signalling pathway regulates
epithelial-mesenchymal transition in cancer. Eur J Cancer.
51:1638–1649. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen W, Yang J, Zhang Y, Cai H, Chen X and
Sun D: Regorafenib reverses HGF-induced sorafenib resistance by
inhibiting epithelial-mesenchymal transition in hepatocellular
carcinoma. FEBS Open Bio. 9:335–347. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Roy S, Kar M, Roy S, Saha A, Padhi S and
Banerjee B: Role of β-catenin in cisplatin resistance, relapse and
prognosis of head and neck squamous cell carcinoma. Cell Oncol
(Dordr). 41:185–200. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang D, Qian G, Wang J, Wang T, Zhang L,
Yang P and Lin F: Visfatin is involved in the cisplatin resistance
of osteosarcoma cells via upregulation of Snail and Zeb1. Cancer
Biol Ther. 20:999–1006. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Patel N, Garikapati KR, Makani VKK, Nair
AD, Vangara N, Bhadra U and Pal Bhadra M: Regulating BMI1
expression via miRNAs promote mesenchymal to epithelial transition
(MET) and sensitizes breast cancer cell to chemotherapeutic drug.
PLoS One. 13:e01902452018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nieto MA, Huang RY, Jackson RA and Thiery
JP: EMT: 2016. Cell. 166:21–45. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fischer KR, Durrans A, Lee S, Sheng J, Li
F, Wong ST, Choi H, El Rayes T, Ryu S, Troeger J, et al:
Epithelial-to-mesenchymal transition is not required for lung
metastasis but contributes to chemoresistance. Nature. 527:472–476.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Batlle E, Sancho E, Francí C, Domínguez D,
Monfar M, Baulida J and García De Herreros A: The transcription
factor snail is a repressor of E-cadherin gene expression in
epithelial tumour cells. Nat Cell Biol. 2:84–89. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cano A, Pérez-Moreno MA, Rodrigo I,
Locascio A, Blanco MJ, del Barrio MG, Portillo F and Nieto MA: The
transcription factor snail controls epithelial-mesenchymal
transitions by repressing E-cadherin expression. Nat Cell Biol.
2:76–83. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Peinado H, Ballestar E, Esteller M and
Cano A: Snail mediates E-cadherin repression by the recruitment of
the Sin3A/histone deacetylase 1 (HDAC1)/HDAC2 complex. Mol Cell
Biol. 24:306–319. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Pastushenko I, Brisebarre A, Sifrim A,
Fioramonti M, Revenco T, Boumahdi S, Van Keymeulen A, Brown D,
Moers V, Lemaire S, et al: Identification of the tumour transition
states occurring during EMT. Nature. 556:463–468. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
MacDonald BT, Tamai K and He X:
Wnt/beta-catenin signaling: Components, mechanisms, and diseases.
Dev Cell. 17:9–26. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liang G, Fang X, Yang Y and Song Y:
Silencing of CEMIP suppresses Wnt/β-catenin/Snail signaling
transduction and inhibits EMT program of colorectal cancer cells.
Acta Histochem. 120:56–63. 2018. View Article : Google Scholar : PubMed/NCBI
|