Introduction
According to Globocan, a total of 18.1 million new
cancer cases and 9.6 million cancer-related deaths occurred in 2018
(1). Both in men and women, lung
cancer is the most commonly diagnosed cancer (11.6% of total cases)
and the leading cause of cancer-related death (18.4% of all
cancer-related deaths) (1). It is
estimated that ~80% of lung cancer cases are non-small cell lung
cancer (NSCLC), and ~50% of NSCLC are adenocarcinomas (2). In recent years, radiotherapy has been a
critical therapeutic approach for lung cancer. However, both
recurrence and metastasis after radiotherapy remain
life-threatening complications of cancer and are primarily
responsible for cancer patient mortality (3). Therefore, elucidating the mechanism of
radiation damage is crucial for radiotherapy of lung cancer.
Circular RNA (circRNA) is a broad category of
non-coding RNA characterized by a covalently closed ring framework
without 5′-3′ polarity (4–7). Unlike other forms of RNA, circRNA not
only exists in organisms stably but also has resistance to RNase R
treatment (8–10). Although Sanger et al (11) first identified circRNA in viruses in
1976, its functions in various diseases have only been recently
reported. With the development of new biochemical and computational
methods, circRNA has attracted increasing research attention
(5,12). Previous studies demonstrated that
circRNAs regulate mRNA transcription via two main mechanisms:
Competition with mRNAs leading to inhibition of host gene
expression (13), and elimination of
the inhibitory effects of microRNA (miRNA/miR) on target genes by
acting as miRNA sponges (5,14).
Several previous studies suggested that circRNAs
were involved in various human diseases, such as lupus nephritis
(15), cardiovascular diseases
(16) and systemic neural diseases
(17). The role of circRNAs in human
cancer was also previously examined. For instance, hsa_circ_002059
could be a potential novel and stable biomarker for the diagnosis
of gastric carcinoma (18).
circRNA-000911 might regulate the carcinogenesis of breast cancer
(19). Furthermore, circRNAs could
promote cell proliferation, migration, invasion and metastasis in
NSCLC (20–22). Moreover, previous studies have
suggested that circRNA may play an importantly regulatory role in
radiation-induced esophageal injury (23). circRNA was significantly
differentially expressed in radioresistant esophageal cancer cells
(24) and radioresistant HeLa cells
(25). circRNA_014511 could affect
the radiosensitivity of bone marrow mesenchymal stem cells by
binding to miR-29b-2-5p (26).
However, the circRNA expression profiles of NSCLC cells in response
to radiation treatment remain unclear.
The present study, investigated for the first time
to the best of the authors' knowledge, the regulation of circRNAs
in irradiated and non-irradiated A549 cells using high-throughput
RNA sequencing (RNAseq). The expression levels of dysregulated
circRNAs were also verified by reverse transcription-quantitative
PCR (RT-qPCR), and the application and function of dysregulated
circRNAs in disease diagnosis were assessed. Overall, the present
study suggested that circRNAs could play an important role in lung
cancer radiotherapy.
Materials and methods
Cell culture
The human NSCLC cell lines A549 and NCI-H1299 were
purchased from The American Type Culture Collection and were
authenticated by STR DNA profiling analysis. NSCLC cells were
cultured in RPMI-1640 medium (GE Healthcare) containing 10% FBS
(Thermo Fisher Scientific, Inc.) and 5 mg/ml
penicillin/streptomycin (Thermo Fisher Scientific, Inc.) at 37°C
with 5% CO2. NSCLC cells were digested and passaged
every 3 days by using EDTA-containing trypsin (Thermo Fisher
Scientific, Inc.).
Irradiation
A549 cells were plated at 1×104
cells/well in 96-well plates. After incubation for 24 h, A549 cells
were exposed to various doses (0, 2, 4 and 8 Gy) of irradiation
using the X-RAD 160–225 instrument (42 cm; 225 kV/s; 12.4 mA; 2.0
Gy/min; filter, 2 mm aluminum; Precision X-Ray, Inc.) (27,28).
Radiosensitivity was assessed after treatment with the irradiation
for 24 h or 48 h.
Cell Counting Kit-8 (CCK-8) assay
Following irradiation, cell proliferation was
measured by CCK-8 according to the manufacturer's instructions. The
CCK-8 kit was purchased from Beyotime Biotechnology Co., Ltd. The
450 nm absorbance (630 nm reference) was determined for each well,
using a Multiskan™ FC Microplate Photometer (Thermo Fisher
Scientific, Inc.). Each experiment was repeated three times.
Total RNA extraction
Total RNA was extracted using the Total RNA
extraction kit (Beijing Solarbio Science & Technology Co.,
Ltd.), according to the manufacturer's instructions. RNA
concentration and purity were determined using a NanoDrop™ 2000
spectrophotometer (Thermo Fisher Scientific, Inc.). RNA integrity
was assessed by 1% formaldehyde denaturing gel electrophoresis. The
total RNA of the control group cells (0 Gy/48 h) and the
irradiation group cells (2 Gy/48 h) were extracted separately for
RNAseq, three samples per group.
circRNA sequencing analysis
RNA libraries were constructed by CloudSeq Pte Ltd.
circRNA sequencing analysis was also performed by CloudSeq Pte Ltd
according to the manufacturer's instructions (14). Cluster and TreeView were used to
conduct hierarchical clustering analysis using R (v3.6.3) (29) in order to assess differential
expression of circRNAs in irradiated and non-irradiated A549 cells.
These circRNAs that can be found in the circbase (http://www.circbase.org/) are known circRNAs, and
those that cannot be found are novel circRNAs. DCC (detect circRNAs
from chimeric reads, v0.4.4) software was used to detect circRNAs
from chimeric reads. The functional roles of target genes in
biological process (BP), cellular component (CC) and molecular
function (MF) were examined through Gene Ontology (GO) (current
release 2020-03-24, http://geneontology.org) analysis. The involvement of
genes encoding circRNAs in biological pathways was assessed by the
Kyoto Encyclopedia of Genes and Genomes (KEGG) using the Database
for Annotation, Visualization and Integrated Discovery v6.8
(https://david.ncifcrf.gov/).
RT-qPCR
circRNA expression profiles obtained from
high-throughput sequencing were verified by RT-qPCR. Total RNA was
extracted from cells using the Total RNA extraction kit (Beijing
Solarbio Science & Technology Co., Ltd.). cDNA was synthesized
using the iScript cDNA synthesis kit (Bio-Rad Laboratories, Inc.)
by RT of 2 µl total RNA. The CFX96 Real-time System (Bio-Rad
Laboratories, Inc.) was used to perform RT-qPCR with SYBR Green
Supermix (Bio-Rad Laboratories, Inc.). The extraction of total RNA,
the synthesis of cDNA, and the steps of RT-qPCR were all performed
according to the manufacturers' instructions (27). The thermocycling conditions were:
Denaturation at 95°C for 30 sec, annealing at 60°C for 30 sec,
extension at 72°C for 60 sec, repeated for 30 cycles. Finally, the
product was kept intact at 72°C for 5 min to complete the extension
and stored at 4°C. The primer sequences used in the present study
are presented in Table SI. The
control gene was β-actin and the 2−ΔΔCq method was used
to calculate the mRNA expression relative to the control group
(30).
miRNA prediction and circRNA-miRNA
interaction network analysis
TargetScan (v7.0) and miRanda (v3.3a) were used to
identify potential miRNA targets of circRNAs (14). The Cloudseq software was used to
analyze interactions between circRNA and miRNA; each circRNA has
multiple miRNA binding sites. The construction of circRNA-miRNA
networks was performed using Cytoscape (v3.1.0) software.
Statistical analysis
Data analysis was performed using GraphPad Prism 7.0
(GraphPad Software, Inc.). Results are presented as the mean ±
standard deviation from at least three independent experiments. A
two-tailed Student's t-test was used to analyze the differences
between the two groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
Overview of circRNA expression
profiles
Based on the results of the CCK-8 assay (Fig. S1), after a 2 Gy irradiation
treatment for 48 hours, the cell viability of A549 was
significantly reduced (P<0.05). The total RNA of the control
group A549 cells (0 Gy/48 h) and the irradiation group A549 cells
(2 Gy/48 h) were extracted separately for RNAseq, three samples per
group. The expression profiles of circRNA were first analyzed in
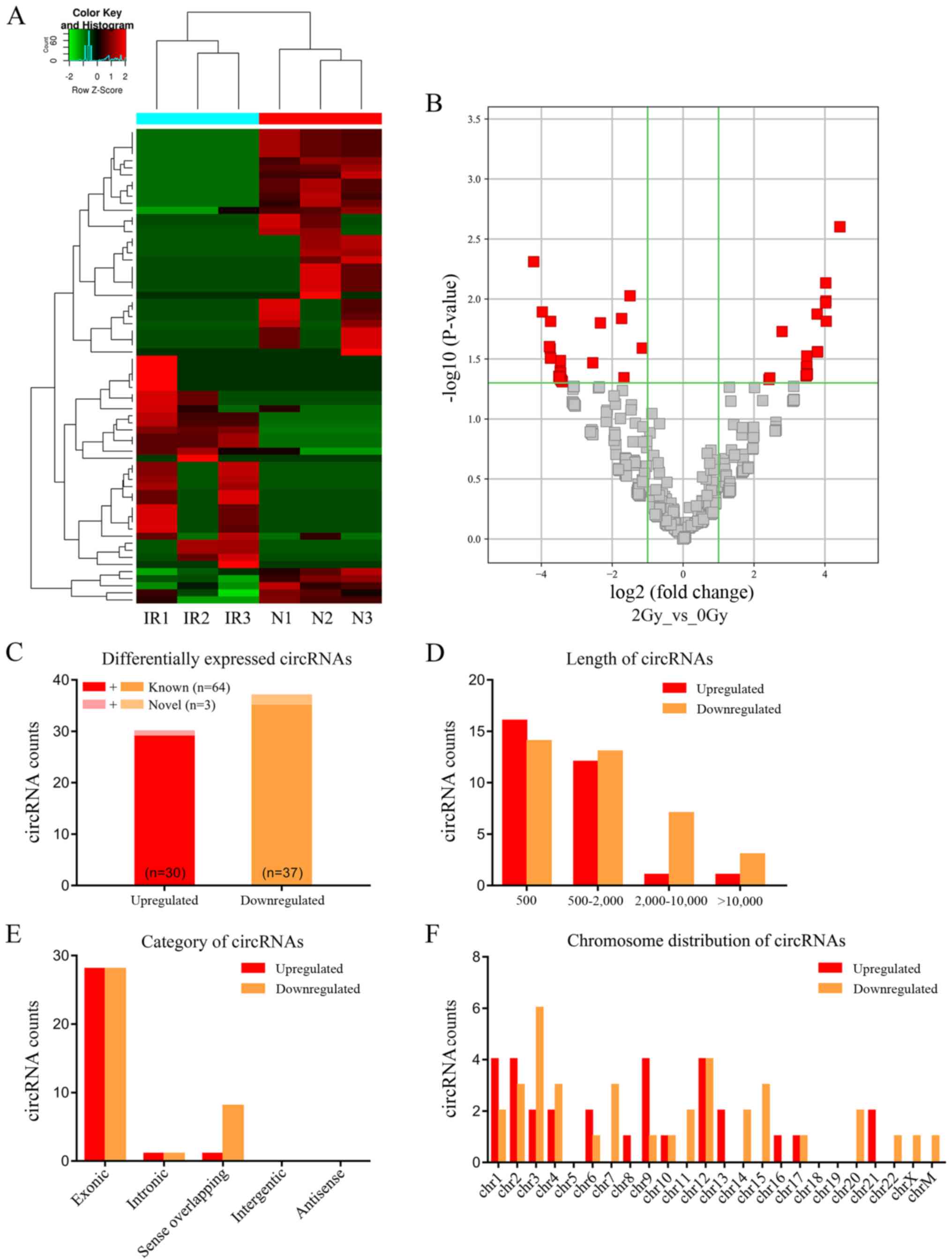
the irradiated and non-irradiated A549 cells. A total of 1,875
circRNA targets was detected by RNAseq. A hierarchical clustering
heatmap (Fig. 1A) and volcano plots
(Fig. 1B) suggested that the
expression levels of circRNA were clustered and distinguishable. A
total of 67 significantly differentially expressed circRNAs in
irradiated and non-irradiated A549 cells (fold change ≥2.0;
P<0.05) were identified. Of these, 30 circRNAs were
significantly upregulated (Table I)
and 37 were downregulated (Table
II) >2-fold in irradiated A549 cells, compared with
non-irradiated A549 cells.
 | Table I.Upregulated circRNAs in irradiated
A549 cells, compared with non-irradiated A549 cells. |
Table I.
Upregulated circRNAs in irradiated
A549 cells, compared with non-irradiated A549 cells.
| circRNA | Log FC | P-value | Chromosomal
location | Gene name |
|---|
|
hsa_circ_0008278 | 4.423 | 0.003 | Chr2:
120885264−120932580+ | EPB41L5 |
|
hsa_circ_0003187 | 4.035 | 0.015 | Chr4:
146767108−146770713− | ZNF827 |
|
hsa_circ_0002174 | 4.029 | 0.01 | Chr3:
195686054−195686957− |
GSE61474_XLOC_045249 |
|
hsa_circ_0070039 | 4.023 | 0.007 | Chr4:
77055328−77065626− | NUP54 |
|
hsa_circ_0004270 | 4.020 | 0.011 | Chr1:
108690901−108703915− | SLC25A24 |
|
hsa_circ_0005591 | 3.794 | 0.028 | Chr12:
110566755−110570441+ | IFT81 |
|
hsa_circ_0029605 | 3.789 | 0.028 | Chr13:
20235838−20244503+ | MPHOSPH8 |
|
hsa_circ_0001865 | 3.787 | 0.028 | Chr9:
86292642−86293514− | UBQLN1 |
|
hsa_circ_0002874 | 3.780 | 0.013 | Chr9:
4286038−4286523− | GLIS3 |
|
hsa_circ_0011218 | 3.506 | 0.042 | Chr1:
31413998−31414970− | PUM1 |
|
hsa_circ_0030632 | 3.499 | 0.042 | Chr13:
96636058−96651561− | UGGT2 |
|
hsa_circ_0005579 | 3.499 | 0.042 | Chr2:
36623757−36691798+ | CRIM1 |
|
hsa_circ_0088239 | 3.497 | 0.043 | Chr9:
119093523−119097353+ | PAPPA |
|
hsa_circ_0105573 | 3.497 | 0.043 | Chr16:
53472928−53504834+ | RBL2 |
|
hsa_circ_0099569 | 3.497 | 0.043 |
Chr12:95663815-95681633+ | VEZT |
| Novel | 3.497 | 0.043 |
Chr6:154749262-154763424− | CNKSR3 |
|
hsa_circ_0044158 | 3.490 | 0.036 |
Chr17:43197692-43198739− | PLCD3 |
|
hsa_circ_0005238 | 3.490 | 0.043 |
Chr21:17205667-17214859+ | USP25 |
|
hsa_circ_0136108 | 3.490 | 0.043 |
Chr8:19442677-19459376− | CSGALNACT1 |
|
hsa_circ_0003677 | 3.488 | 0.03 |
Chr12:117365828-117402659+ | FBXW8 |
|
hsa_circ_0066776 | 3.485 | 0.043 |
Chr3:110830877-110845182+ | PVRL3 |
|
hsa_circ_0009061 | 3.485 | 0.043 |
Chr1:23356962-23377013+ | KDM1A |
|
hsa_circ_0001603 | 3.482 | 0.043 |
Chr6:42559889-42562042+ | UBR2 |
|
hsa_circ_0000199 | 3.479 | 0.037 |
Chr1:243708812-243736350− | AKT3 |
|
hsa_circ_0004820 | 3.475 | 0.044 |
Chr2:183993015-183995273+ | NUP35 |
|
hsa_circ_0002224 | 3.475 | 0.044 |
Chr10:51374370-51387763+ | TIMM23B |
|
hsa_circ_0053317 | 3.468 | 0.044 |
Chr2:28460069-28464260+ | BRE |
|
hsa_circ_0000378 | 2.792 | 0.019 |
Chr12:12397196-12397589− | LRP6 |
|
hsa_circ_0001190 | 2.446 | 0.046 |
Chr21:38792601-38794168+ | DYRK1A |
|
hsa_circ_0005456 | 2.422 | 0.047 |
Chr9:118949433-118950495+ | PAPPA |
| Table II.Downregulated circRNAs in irradiated
A549 cells, compared with non-irradiated A549 cells. |
Table II.
Downregulated circRNAs in irradiated
A549 cells, compared with non-irradiated A549 cells.
| circRNA | Log FC | P-value | Chromosomal
location | Gene name |
|---|
|
hsa_circ_0001009 | −4.213 | 0.005 | Chr2:
58449077−58459247− | FANCL |
|
hsa_circ_0004815 | −3.972 | 0.013 | Chr12:
42745687−42792796+ | PPHLN1 |
|
hsa_circ_0096360 | −3.769 | 0.025 | Chr11:
70505933−70507783− | SHANK2 |
|
hsa_circ_0036627 | −3.767 | 0.025 | Chr15:
85656608−85669605+ | PDE8A |
|
hsa_circ_0004524 | −3.750 | 0.026 | Chr3:
138289160−138291774− | CEP70 |
|
hsa_circ_0006834 | −3.740 | 0.031 | Chr2:
120885264−120932576+ | EPB41L5 |
|
hsa_circ_0006284 | −3.740 | 0.031 | Chr11:
12883797−12886447+ | TEAD1 |
|
hsa_circ_0001345 | −3.736 | 0.015 | Chr3:
142455221−142467302+ | TRPC1 |
|
hsa_circ_0127359 | −3.736 | 0.031 | Chr4:
95796982−95820201+ | BMPR1B |
|
hsa_circ_0098537 | −3.736 | 0.031 | Chr12:
41408030−41423021+ | CNTN1 |
|
hsa_circ_0005925 | −3.503 | 0.044 | Chr7:
99952766−99953427+ | PILRB |
|
hsa_circ_0002658 | −3.479 | 0.045 | Chr3:
15726733−15731727− | ANKRD28 |
|
hsa_circ_0116480 | −3.479 | 0.045 | Chr22:
32664275−32669494+ | G053490 |
|
hsa_circ_0124903 | −3.479 | 0.045 | Chr4:
103644028−103651893− | MANBA |
|
hsa_circ_0000606 | −3.479 | 0.045 | Chr15:
60734615−60737990− | ICE2 |
|
hsa_circ_0001977 | −3.466 | 0.046 | Chr20:
17928130−17937681− | SNX5 |
| Novel | −3.464 | 0.040 | ChrM:
13298−13449+ | MTND5 |
| Novel | −3.464 | 0.046 | Chr20:
45801355−45809584+ | EYA2 |
|
hsa_circ_0004844 | −3.460 | 0.033 | ChrX:
2871184−2876476− | ARSE |
|
hsa_circ_0005062 | −3.455 | 0.047 | Chr9:
710804−713464+ | KANK1 |
|
hsa_circ_0002714 | −3.453 | 0.047 | Chr7:
80418622−80435074− | SEMA3C |
|
hsa_circ_0058495 | −3.453 | 0.046 | Chr2:
227729320−227779067+ | RHBDD1 |
|
hsa_circ_0003692 | −3.453 | 0.046 | Chr3:
171969050−172028671+ | FNDC3B |
|
hsa_circ_0078299 | −3.453 | 0.041 | Chr6:
151669846−151674887+ | AKAP12 |
|
hsa_circ_0101802 | −3.442 | 0.047 | Chr14:
39648295−39648666+ | PNN |
|
hsa_circ_0000532 | −3.440 | 0.047 | Chr14:
45564424−45566208+ | PRPF39 |
|
hsa_circ_0006501 | −3.431 | 0.048 | Chr7:
101870647−101870949+ | CUX1 |
|
hsa_circ_0006021 | −3.418 | 0.048 | Chr1:
235582788−235597595+ | TBCE |
|
hsa_circ_0123217 | −3.418 | 0.048 | Chr3:
196214270−196215554− | RNF168 |
|
hsa_circ_0004532 | −3.418 | 0.048 | Chr17:
12011107−12016677+ | MAP2K4 |
|
hsa_circ_0006668 | −3.407 | 0.049 | Chr12:
51442817−51451911+ | LETMD1 |
|
hsa_circ_0008153 | −2.549 | 0.034 | Chr15:
63845914−63855207+ | USP3 |
|
hsa_circ_0093688 | −2.333 | 0.016 | Chr10:
4872867−4950612+ | AKR1E2 |
|
hsa_circ_0000443 | −1.733 | 0.015 | Chr12:
116668338−116675510− | MED13L |
|
hsa_circ_0001460 | −1.669 | 0.045 |
Chr4:178274462-178281831+ | NEIL3 |
|
hsa_circ_0112879 | −1.497 | 0.009 |
Chr1:247319708-247323115− | ZNF124 |
|
hsa_circ_0005332 | −1.152 | 0.026 |
Chr3:114069121-114070725− | ZBTB20 |
Of the 67 differentially expressed circRNAs, 64 were
known circRNAs listed in circbase, and 3 were novel (Fig. 1C). Most of the 67 identified circRNAs
were <2,000 nucleotides in length (Fig. 1D). Based on structure, five
categories of circRNAs were identified: i) Exonic circRNAs; ii)
intronic circRNAs; iii) sense overlapping circRNAs; iv) intergenic
circRNAs; and v) antisense circRNAs. Exonic circRNAs represented
83.6% (56/67) of all identified circRNAs, sense overlapping
circRNAs were at 13.4% (9/67), and other circRNAs at 3% (2/67;
Fig. 1E). Sixty-six circRNAs were
located on chromosomes, while one downregulated circRNA was located
in the mitochondrial genome (Fig.
1F).
Predicted functions and pathways of
differentially expressed circRNAs
The special RNA analysis software (DCC) was used to
obtain the genomic location of circRNA target genes. In combination
with the gene annotation in the circbase, the target genes of
circRNAs were obtained. The functions and pathways of
differentially expressed circRNAs were predicted by GO and KEGG
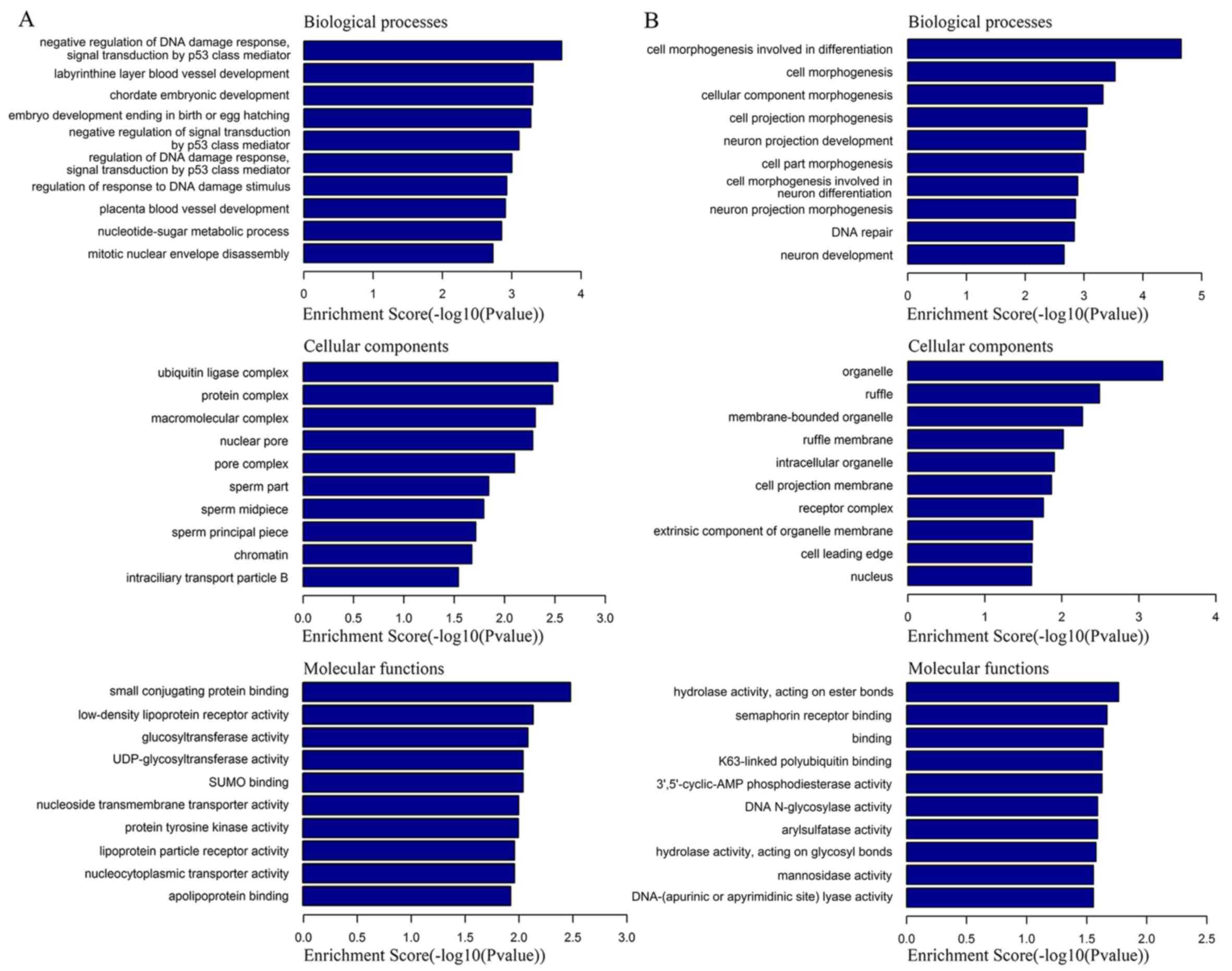
analysis based on their corresponding target genes. GO enrichment
analysis predicted the top 10 functional roles of target host genes
according to BP, CC and MF (Fig. 2).
GO analysis suggested the target genes participated in multiple BPs
related to human tumorigenesis, including ‘cell morphogenesis
involved in differentiation’, ‘neuron development’ and ‘chordate
embryonic development’.
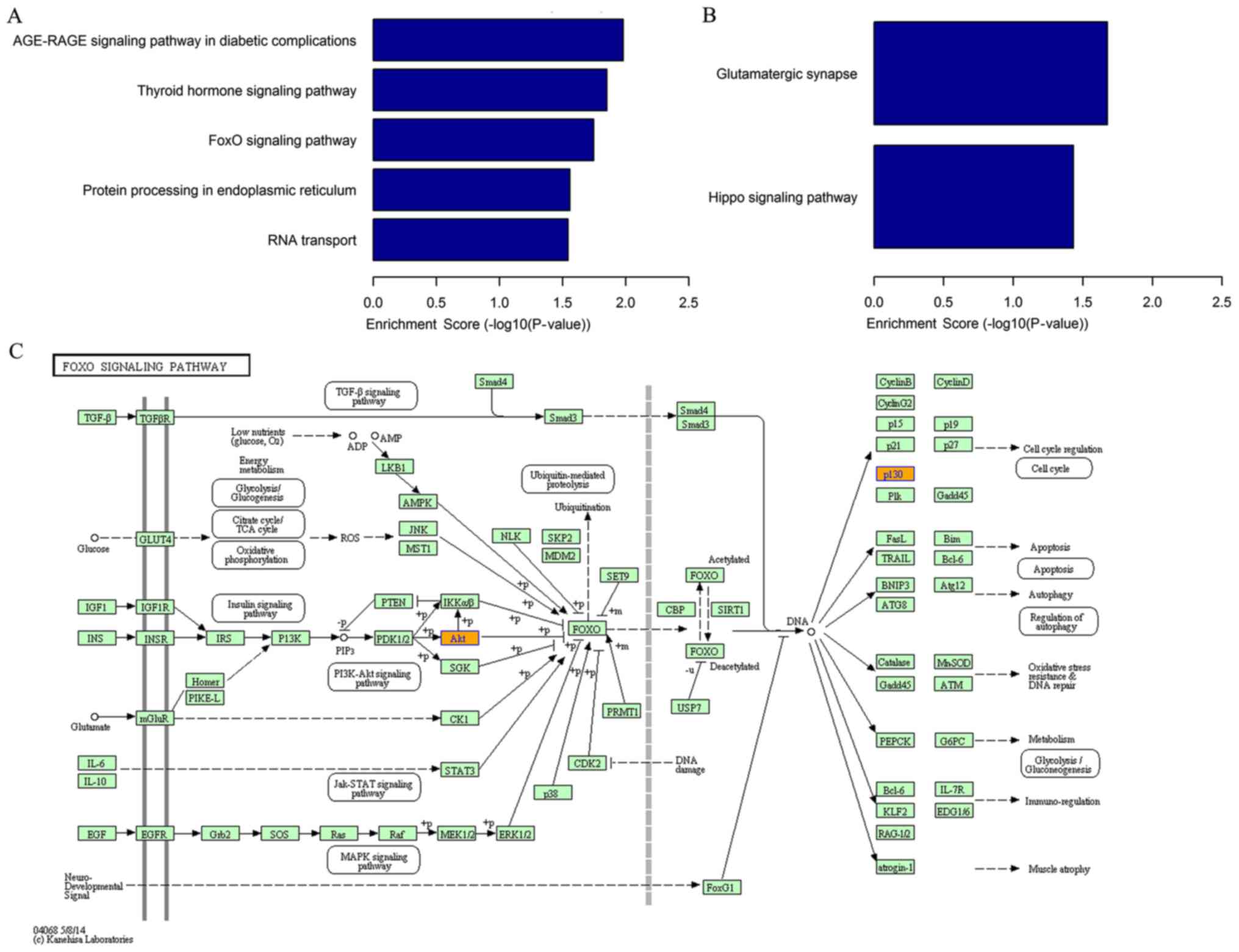
The KEGG enrichment analysis identified five
pathways related to the upregulated circRNAs, the ‘AGE-RAGE
signaling pathway in diabetic complications’, ‘thyroid hormone
signaling pathway’, ‘FoxO signaling pathway’, ‘protein processing
in the endoplasmic reticulum’ and ‘RNA transport’ (Fig. 3A). The pathway analysis also
indicated two pathways associated with downregulated circRNAs,
namely the ‘glutamatergic synapse’ and the ‘Hippo signaling
pathway’ (Fig. 3B). The FOXO
signaling pathway is presented as an example (Fig. 3C). The FOXO signaling pathway was
associated with several BPs, including ‘cell cycle’, ‘apoptosis’
and ‘regulation of autophagy’.
Validation of the accuracy of circRNA
sequencing data by RT-qPCR
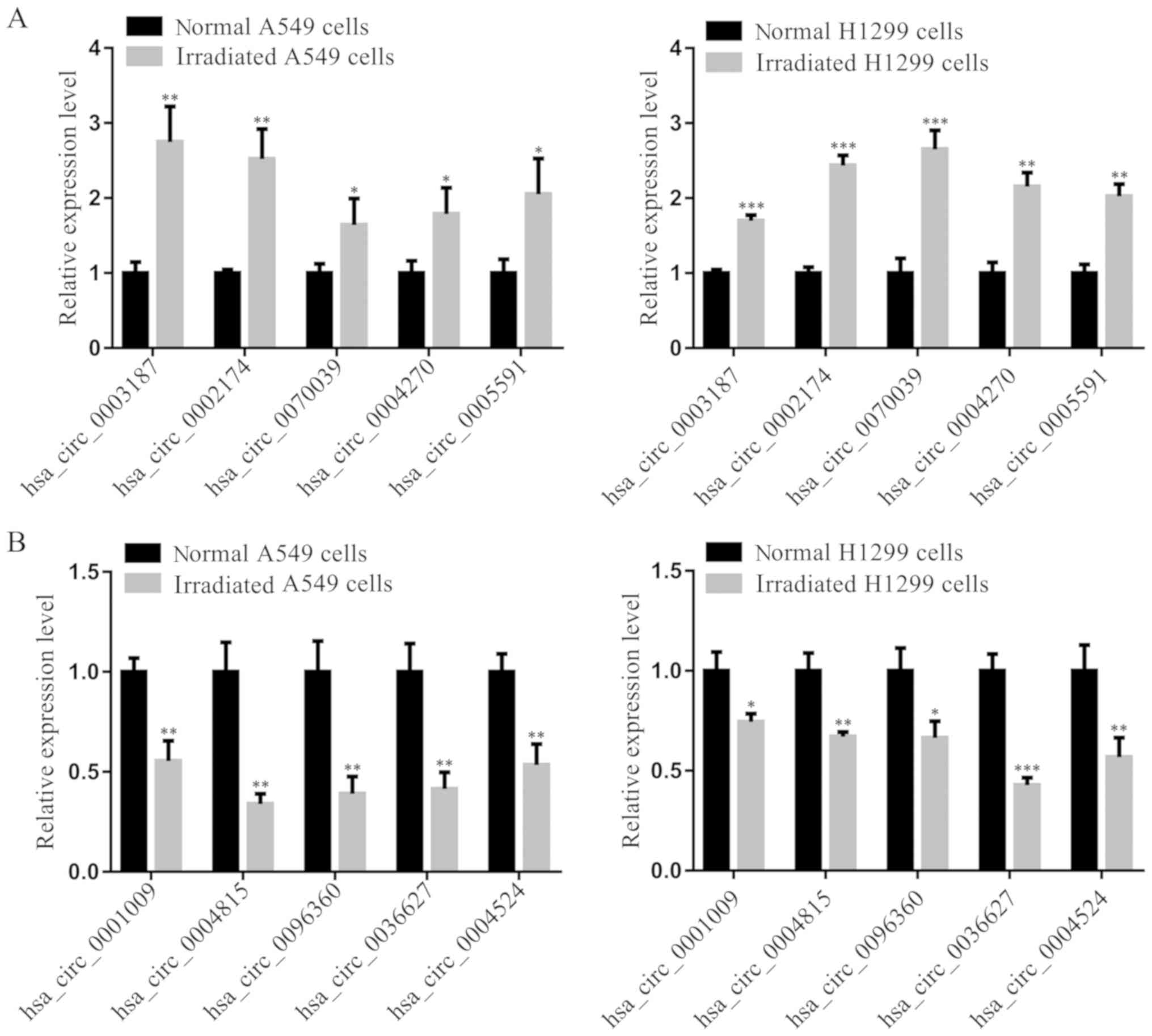
To validate the RNAseq data, the top 5 upregulated
(excluding hsa_circ_0008278 due to its sequence length >40,000
nucleotides) and downregulated circRNAs were selected based on
P-value and fold change, and subjected to RT-qPCR. RT-qPCR
demonstrated that hsa_circ_0003187, hsa_circ_0002174,
hsa_circ_0070039, hsa_circ_0004270 and hsa_circ_0005591 expressions
were significantly upregulated in irradiated cells compared with
corresponding non-irradiated cells (Fig.
4A). Moreover, the expression of hsa_circ_0001009,
hsa_circ_0004815, hsa_circ_0096360, hsa_circ_0036627 and
hsa_circ_0004524 were significantly downregulated in irradiated
cells (Fig. 4B). These 10 circRNA
expression levels detected by RT-qPCR were consistent with RNAseq
data, demonstrating the accuracy of high-throughput RNAseq
results.
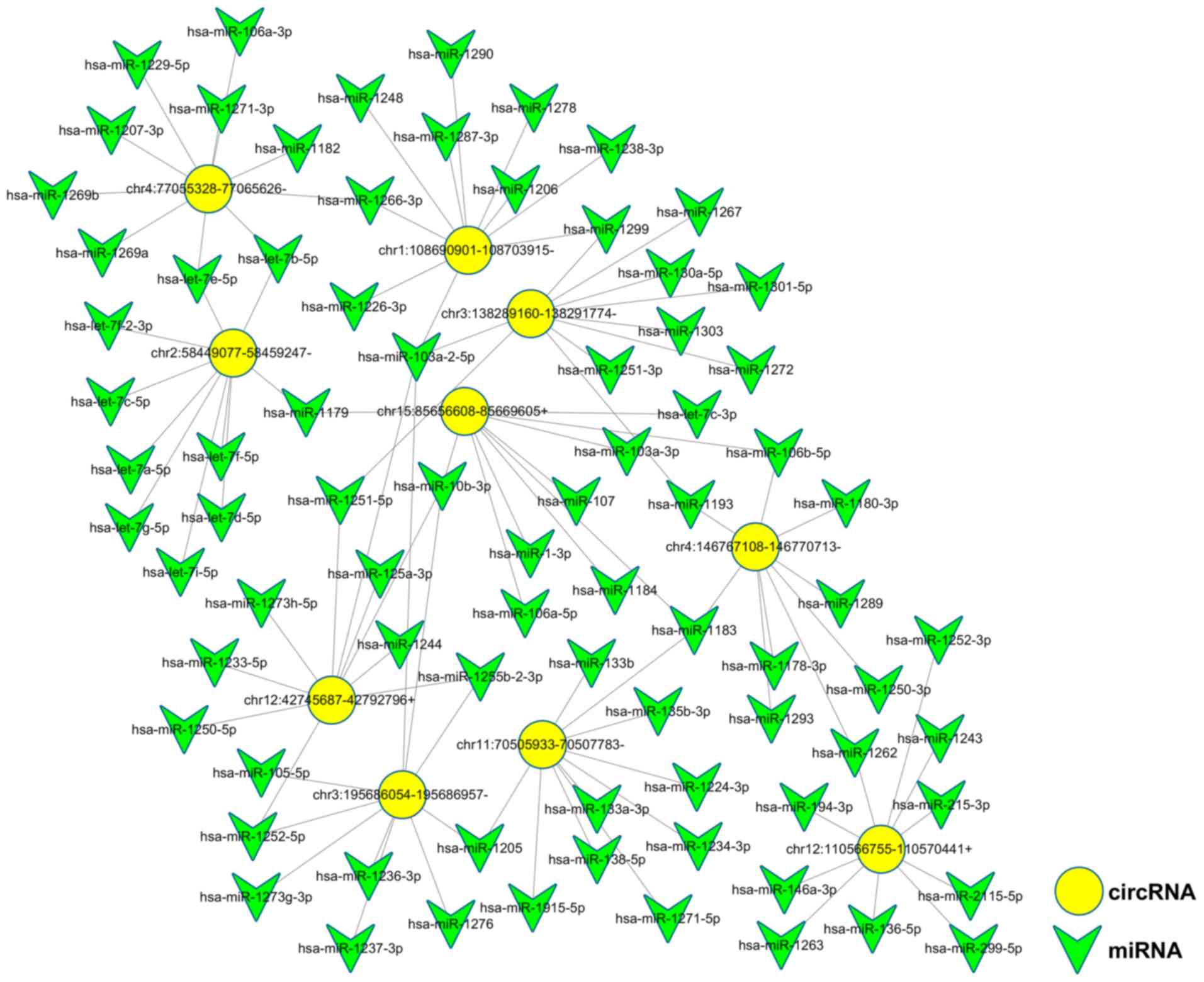
circRNA-miRNA co-expression
network
Potential interactions between the 10 validated
circRNAs and microRNAs were theoretically predicted through
conserved binding sites using miRanda and TargetScan analyses. All
10 circRNAs had complementary miRNA response elements.
circRNA_0002174 and circRNA_0036627 were the two circRNAs with the
largest number of predicted miRNA-binding sites among upregulated
circRNAs and downregulated circRNAs, respectively (Fig. 5; Table
III). These two circRNAs could be investigated to help resolve
the molecular mechanisms of radiotherapy.
| Table III.A total of 10 validated circRNA and
their miRNA targets. |
Table III.
A total of 10 validated circRNA and
their miRNA targets.
| circRNA | Chromosomal
location | No. of miRNA
targets |
|---|
|
hsa_circ_0036627 | Chr15:
85656608−85669605+ | 378 |
|
hsa_circ_0002174 | Chr3:
195686054−195686957− | 276 |
|
hsa_circ_0004815 | Chr12:
42745687−42792796+ | 274 |
|
hsa_circ_0070039 | Chr4:
77055328−77065626− | 229 |
|
hsa_circ_0004270 | Chr1:
108690901−108703915− | 164 |
|
hsa_circ_0004524 | Chr3:
138289160−138291774− | 121 |
|
hsa_circ_0003187 | Chr4:
146767108−146770713− | 114 |
|
hsa_circ_0001009 | Chr2:
58449077−58459247− | 103 |
|
hsa_circ_0005591 | Chr12:
110566755−110570441+ | 66 |
|
hsa_circ_0096360 | Chr11:
70505933−70507783− | 60 |
Discussion
With the development of new computational methods
and high-throughput sequencing, circRNA has been recognized as a
stable and abundant form of RNA, and is presently regarded as an
essential part of the non-coding RNA family (31). Previously, several studies
demonstrated that circRNAs played a vital role in the
pathophysiology of various diseases, with particular relevance in
different types of cancer (18,32,33).
Furthermore, circRNAs could promote cell proliferation, migration,
invasion and metastasis in NSCLC (20–22).
Moreover, previous studies have found that circRNA may play a major
role in response to radiation (23).
circRNA was significantly differently expressed in radioresistant
esophageal cancer cells (24) and
radioresistant HeLa cells (25).
circRNA_014511 could affect the radiosensitivity of bone marrow
mesenchymal stem cells by binding to miR-29b-2-5p (26). However, the functions and biological
features of circRNAs in lung cancer radiotherapy remain poorly
understood. In the present study, the expression levels of circRNAs
in irradiated and non-irradiated A549 cells were determined by
circRNA sequencing, to examine the possible underlying involvement
of differentially expressed circRNAs in the mechanisms of lung
cancer radiotherapy.
To the best of the authors' knowledge, the present
study was the first to identify differentially expressed circRNAs
in irradiated A549 cells by examining junction reads. A549 cells
received a 2 Gy irradiation dose and total RNA was collected after
48 h. This time interval was chosen because most of the
radiation-related genes with high expression were strongly
inhibited after 48 h, based on previous studies (25,34).
Among the total 1,875 identified circRNAs, 30 significantly
upregulated circRNAs and 37 significantly downregulated circRNAs
were detected in irradiated A549 cells. The expression levels of
the top 5 upregulated and downregulated circRNAs were further
validated by RT-qPCR, which were consistent with high-throughput
sequencing data.
Differentially expressed circRNAs may regulate the
transcription of several target genes in irradiated lung cancer
cells. In order to further investigate the potential functions of
circRNAs in irradiated A549 cells, GO and KEGG analyses were
conducted. GO analysis was used to identify the BP, CC and MF of
target genes. Previous studies demonstrated that irradiation can
cause DNA strand breaks via different mechanisms (35,36).
Notably, irradiation induces DNA damage (37). The results of the present study
suggested that the target genes associated with dysregulated
circRNAs might be related to the regulation of DNA damage response
(‘signal transduction by p53 class mediator’). In addition, the
present study also indicated that ‘labyrinthine layer blood vessel
development’ was a notable BP term in response to irradiation.
Similarly, Lee et al (38)
had previously identified that vascular endothelial growth factor
was highly expressed following irradiation, which might lead to
rapid blood vessel development. ‘Small conjugating protein binding’
was also an important GO term. KEGG analysis identified several
pathways associated with the upregulated and downregulated circRNAs
identified in the present study. Among these pathways, the FOXO
signaling pathway was identified. The FOXO signaling pathway was
associated with several cellular functions in previous studies,
including proliferation, apoptosis, inflammation migration,
antitumor activities and metabolism through regulation of numerous
transcriptional targets (39,40). A
further understanding of the function of these differentially
expressed circular RNAs will help to understand the mechanisms
associated with radiation damage.
Previous studies demonstrated that circRNAs could
negatively regulate the effects of miRNAs, and promote the effects
of endogenous mRNAs (15,41). A single circRNA may regulate multiple
miRNAs, thereby affecting several pathways. Zhao et al
(42) suggested that ciRS-7 had
>60 well-known miR-7 binding sites, which is far more than any
known linear RNA sponge. In the present study, a circRNA-miRNA
interaction network was constructed to examine the biological role
of circRNAs following irradiation. The network analysis suggested
that two potential circRNAs (circRNA_0002174 and circRNA_0036627)
might play an important role in the regulation of target miRNAs.
Based on the characteristics of circRNAs, including structural
stability, stable expression and tissue specificity, circRNAs could
serve as a novel biomarker and therapeutic target for cancer
treatment. Chen et al (43)
suggested that circRNA epithelial stromal interaction 1 played a
vital role as a prognostic marker in patients with triple-negative
breast cancer. Nair et al (44) suggested that circRNAs might function
as markers of cell proliferation in breast cancer. circPRKCI may
serve as a potential therapeutic target for patients with lung
adenocarcinoma (45). The present
study demonstrated that differential expression of circRNAs in
irradiated and non-irradiated A549 cells could be a critical marker
of radiation-induced damage.
In conclusion, the preset study identified
differentially expressed circRNAs and their target genes in
irradiated A549 cells. The results of the present study suggested
that circRNAs might play an important role in response to
irradiation. The top 5 upregulated and downregulated circRNAs were
validated; however, further investigation is required in order to
examine the molecular functions of circRNAs in radiotherapy.
Several potential circRNA-miRNA networks were predicted through
bioinformatics analysis. These results warrant further
investigation of the potential biological functions of circRNAs in
lung cancer radiotherapy.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Cloud-Seq Biotech
Ltd. Co. (Shanghai, China) for the circRNA sequencing service and
the subsequent bioinformatics analysis.
Funding
The present study was funded by The National Natural
Science Foundation of China (grant nos. 81802955 and 81972977), The
Foundation of Sichuan Science and Technology Agency (grant nos.
2018JY0648 and 2019YJ0589), The Foundation of The First Affiliated
Hospital of Chengdu Medical College (grant nos. CYFY2017ZD03 and
CYFY2018ZD02) and The Foundation of Chengdu Medical College (grant
no. CYCG16-04).
Availability of data and materials
The datasets supporting the conclusions of this
article are available in the Gene Expression Omnibus repository
(GSE124396; https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE1243
96).
Authors' contributions
TZ, DW and YX conceived and designed the
experiments. TZ and DW performed the experiments and wrote drafts
of the paper; TZ, SD and RH analyzed the data; DW, TL and JL
acquired the data; YX reviewed the drafts of the paper. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhu W, Li Z, Xiong L, Yu X, Chen X and Lin
Q: FKBP3 Promotes proliferation of non-small cell lung cancer cells
through regulating Sp1/HDAC2/p27. Theranostics. 7:3078–3089. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chiu LY, Hsin IL, Yang TY, Sung WW, Chi
JY, Chang JT, Ko JL and Sheu GT: The ERK-ZEB1 pathway mediates
epithelial-mesenchymal transition in pemetrexed resistant lung
cancer cells with suppression by vinca alkaloids. Oncogene.
36:242–253. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Haimovich G, Medina DA, Causse SZ, Garber
M, Millán-Zambrano G, Barkai O, Chávez S, Pérez-Ortín JE, Darzacq X
and Choder M: Gene expression is circular: Factors for mRNA
degradation also foster mRNA synthesis. Cell. 153:1000–10011. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hansen TB, Jensen TI, Clausen BH, Bramsen
JB, Finsen B, Damgaard CK and Kjems J: Natural RNA circles function
as efficient microRNA sponges. Nature. 495:384–388. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wilusz JE and Sharp PA: Molecular biology.
A circuitous route to noncoding RNA. Science. 340:440–441. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Qu S, Yang X, Li X, Wang J, Gao Y, Shang
R, Sun W, Dou K and Li H: Circular RNA: A new star of noncoding
RNAs. Cancer Lett. 365:141–148. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rybak-Wolf A, Stottmeister C, Glažar P,
Jens M, Pino N, Giusti S, Hanan M, Behm M, Bartok O, Ashwal-Fluss
R, et al: Circular RNAs in the mammalian brain are highly abundant,
conserved, and dynamically expressed. Mol Cell. 58:870–885. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Szabo L, Morey R, Palpant NJ, Wang PL,
Afari N, Jiang C, Parast MM, Murry CE, Laurent LC and Salzman J:
Statistically based splicing detection reveals neural enrichment
and tissue-specific induction of circular RNA during human fetal
development. Genome Biol. 16:1262015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Qiu X, Ke X, Ma H, Han L, Chen Q, Zhang S,
Da P and Wu H: Profiling and bioinformatics analyses reveal
differential expression of circular RNA in tongue cancer revealed
by high-throughput sequencing. J Cell Biochem. 120:4102–4112. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sanger HL, Klotz G, Riesner D, Gross HJ
and Kleinschmidt AK: Viroids are single-stranded covalently closed
circular RNA molecules existing as highly base-paired rod-like
structures. Proc Natl Acad Sci USA. 73:3852–3856. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen LL and Yang L: Regulation of circRNA
biogenesis. RNA Biol. 12:381–388. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Salzman J: Circular RNA expression: Its
potential regulation and function. Trends Genet. 32:309–316. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zheng Q, Bao C, Guo W, Li S, Chen J, Chen
B, Luo Y, Lyu D, Li Y, Shi G, et al: Circular RNA profiling reveals
an abundant circHIPK3 that regulates cell growth by sponging
multiple miRNAs. Nat Commun. 7:112152016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Luan J, Jiao C, Kong W, Fu J, Qu W, Chen
Y, Zhu X, Zeng Y, Guo G, Qi H, et al: circHLA-C plays an important
role in lupus nephritis by sponging miR-150. Mol Ther Nucl Acids.
10:245–253. 2018. View Article : Google Scholar
|
|
16
|
Tan WL, Lim BT, Anene-Nzelu CG,
Ackers-Johnson M, Dashi A, See K, Tiang Z, Lee DP, Chua WW and Luu
TD: A landscape of circular RNA expression in the human heart.
Cardiovasc Res. 113:298–309. 2017.PubMed/NCBI
|
|
17
|
Shao Y and Chen Y: Roles of circular RNAs
in neurologic disease. Front Mol Neurosci. 9:252016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li P, Chen S, Chen H, Mo X, Li T, Shao Y,
Xiao B and Guo J: Using circular RNA as a novel type of biomarker
in the screening of gastric cancer. Clin Chim Acta. 444:132–136.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang H, Xiao Y, Wu L and Ma D:
Comprehensive circular RNA profiling reveals the regulatory role of
the circRNA-000911/miR-449a pathway in breast carcinogenesis. Int J
Oncol. 52:743–754. 2018.PubMed/NCBI
|
|
20
|
Wu K, Liao X, Gong Y, He J, Zhou JK, Tan
S, Pu W, Huang C, Wei YQ and Peng Y: Circular RNA F-circSR derived
from SLC34A2-ROS1 fusion gene promotes cell migration in non-small
cell lung cancer. Mol Cancer. 18:982019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li XY, Liu YR, Zhou JH, Li W, Guo HH and
Ma HP: Enhanced expression of circular RNA hsa_circ_000984 promotes
cells proliferation and metastasis in non-small cell lung cancer by
modulating Wnt/β-catenin pathway. Eur Rev Med Pharmacol Sci.
23:3366–3374. 2019.PubMed/NCBI
|
|
22
|
Chang H, Qu J, Wang J, Liang X and Sun W:
Circular RNA circ_0026134 regulates non-small cell lung cancer cell
proliferation and invasion via sponging miR-1256 and miR-1287.
Biomed Pharmacother. 112:1087432019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Luo J, Zhang C, Zhan Q, An F, Zhu W, Jiang
H and Ma C: Profiling circRNA and miRNA of radiation-induced
esophageal injury in a rat model. Sci Rep. 8:146052018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Su H, Lin F, Deng X, Shen L, Fang Y, Fei
Z, Zhao L, Zhang X, Pan H, Xie D, et al: Profiling and
bioinformatics analyses reveal differential circular RNA expression
in radioresistant esophageal cancer cells. J Transl Med.
14:2252016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yu D, Li Y, Ming Z, Wang H, Dong Z, Qiu L
and Wang T: Comprehensive circular RNA expression profile in
radiation-treated HeLa cells and analysis of
radioresistance-related circRNAs. Peer J. 6:e50112018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang Y, Zhang J, Li J, Gui R, Nie X and
Huang R: CircRNA_014511 affects the radiosensitivity of bone marrow
mesenchymal stem cells by binding to miR-29b-2-5p. Bosn J Basic Med
Sci. 19:155–163. 2019.PubMed/NCBI
|
|
27
|
Wu D, Han R, Deng S, Liu T, Zhang T, Xie H
and Xu Y: Protective effects of flagellin A N/C against
radiation-induced NLR pyrin domain containing 3
inflammasome-dependent pyroptosis in intestinal cells. Int J Radiat
Oncol. 101:107–117. 2018. View Article : Google Scholar
|
|
28
|
Wang M, Meng B and Liu Y, Yu J, Chen Q and
Liu Y: MiR-124 inhibits growth and enhances radiation-induced
apoptosis in non-small cell lung cancer by inhibiting STAT3. Cell
Physiol Biochem. 44:2017–2028. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
R Core Team, . 2012.R: A language and
environment for statistical computing. R Foundation for Statistical
Computing. (Vienna, Austria). ISBN 3-900051-07-0, URL
http://www.R-project.org/.
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ivanov A, Memczak S, Wyler E, Torti F,
Porath HT, Orejuela MR, Piechotta M, Levanon EY, Landthaler M,
Dieterich C and Rajewsky N: Analysis of intron sequences reveals
hallmarks of circular RNA biogenesis in animals. Cell Rep.
10:170–177. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Qin M, Liu G, Huo X, Tao X, Sun X, Ge Z,
Yang J, Fan J, Liu L and Qin W: Hsa_circ_0001649: A circular RNA
and potential novel biomarker for hepatocellular carcinoma. Cancer
Biomark. 16:161–169. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Su H, Jin X, Zhang X, Zhao L, Lin B, Li L,
Fei Z, Shen L, Fang Y, Pan H and Xie C: FH535 increases the
radiosensitivity and reverses epithelial-to-mesenchymal transition
of radioresistant esophageal cancer cell line KYSE-150R. J Transl
Med. 13:1042015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang H, Yue J, Jiang Z, Zhou R, Xie R, Xu
Y and Wu S: CAF-secreted CXCL1 conferred radioresistance by
regulating DNA damage response in a ROS-dependent manner in
esophageal squamous cell carcinoma. Cell Death Dis. 8:e27902017.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Samstein RM and Riaz N: The DNA damage
response in immunotherapy and radiation. Adv Radiat Oncol.
3:527–533. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Moreno-Villanueva M, Feiveson AH, Krieger
S, Kay Brinda A, von Scheven G, Bürkle A, Crucian B and Wu H:
Synergistic effects of weightlessness, isoproterenol, and radiation
on DNA damage response and cytokine production in immune cells. Int
J Mol Sci. 19:E36892018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Goldstein M and Kastan MB: The DNA damage
response: Implications for tumor responses to radiation and
chemotherapy. Annu Rev Med. 66:129–143. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lee C, Shim S, Jang H, Myung H, Lee J, Bae
CH, Myung JK, Kim MJ, Lee SB, Jang WS, et al: Human umbilical cord
blood-derived mesenchymal stromal cells and small intestinal
submucosa hydrogel composite promotes combined radiation-wound
healing of mice. Cytotherapy. 19:1048–1059. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Alikhani M, Alikhani Z and Graves DT:
FOXO1 functions as a master switch that regulates gene expression
necessary for tumor necrosis factor-induced fibroblast apoptosis. J
Biol Chem. 280:12096–12102. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang Y, Zhou Y and Graves DT: FOXO
Transcription factors: Their clinical significance and regulation.
Biomed Res Int. 2014:9253502014.PubMed/NCBI
|
|
41
|
Wang K, Long B, Liu F, Wang JX, Liu CY,
Zhao B, Zhou LY, Sun T, Wang M, Yu T, et al: A circular RNA
protects the heart from pathological hypertrophy and heart failure
by targeting miR-223. Eur Heart J. 37:2602–2611. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhao ZJ and Shen J: Circular RNA
participates in the carcinogenesis and the malignant behavior of
cancer. RNA Biol. 14:514–521. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chen B, Wei W, Huang X and Xie X, Kong Y,
Dai D, Yang L, Wang J, Tang H and Xie X: circEPSTI1 as a prognostic
marker and mediator of triple-negative breast cancer progression.
Theranostics. 8:4003–4015. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Nair AA, Niu N, Tang X, Thompson KJ, Wang
L, Kocher JP, Subramanian S and Kalari KR: Circular RNAs and their
associations with breast cancer subtypes. Oncotarget.
7:80967–80979. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Qiu M, Xia W, Chen R, Wang S, Xu Y, Ma Z,
Xu W, Zhang E, Wang J, Fang T, et al: The circular RNA circPRKCI
promotes tumor growth in lung adenocarcinoma. Cancer Res.
78:2839–2851. 2018. View Article : Google Scholar : PubMed/NCBI
|