Introduction
Mucin 1 (MUC1) is a membrane-associated glycoprotein
involved in the protection of mucous membranes and modulation of
immune system (1). MUC1 is
upregulated in >70% of all types of cancer and has important
roles in tumorigenicity and tumor progression; thus, this antigen
may be a potential target for cancer immunotherapy (2). T-cell epitopes of the MUC1 core domain
bind major histocompatibility complex (MHC) class I molecules in
their truncated hypoglycosylated form, which directs the
MHC-restricted recognition of epitopes (3,4).
MUC1-based clinical trials are currently ongoing, including
retargeting human CD8+ and CD4+ T cells to
tumor-associated MUC1 glycoforms (5,6). In
addition, TG4010, a viral vaccine developed by Transgene SA,
expresses full-length MUC1 and interleukin (IL)-2, and has
demonstrated an association between overall survival and
vaccine-induced T-cell responses (7). Lakshminarayanan et al (8) have also constructed a tumor vaccine by
covalent attachment of a T-helper epitope and an aberrantly
glycosylated MUC1 peptide, which induces immunoglobulin (Ig) G
antibodies and cytotoxic T lymphocytes (CTLs) against MUC1.
However, a lack of immune recognition, resulting from the weak
immunogenicity of tumor antigens, is the primary reason for the
inadequacy of vaccines to induce potent immune responses (8).
Toll-like receptors (TLRs) are a family of integral
membrane proteins that are primarily localized on immune cells,
such as dendritic cells (DCs) and macrophages (9). TLRs recognize molecules that are widely
shared by pathogens, known as pathogen-associated molecular
patterns (9). Following the
recruitment of adapter proteins, TLR activation leads to the
stimulation of myeloid differentiation primary response
88-dependent signaling and the subsequent release of inflammatory
cytokines and stimulatory molecules (10). As a bridge between innate and
adaptive immunity, TLRs have been used for conjugation with ligands
and antigens, and have been demonstrated to exhibit advantages over
non-coupled antigens (11). Among
the TLRs, only TLR7 recognizes small synthetic molecules, including
nucleoside analogues, which are easier to obtain and modify
compared with other biomacromolecules, such as TLR4 and TLR9
ligands (12). TLR7 agonists (T7s)
have attracted attention for their promotion of not only
antigen-presenting cells (APCs), but also T cells and natural
killer (NK) cells (13,14). Our previous study investigated small
molecule TLR7 ligands, and a series of T7s were synthesized with
significant immunostimulatory activity (15). Our previous study also constructed
tumor vaccines by conjugating the T7 with gastric cancer antigens,
which had synergistic antitumor effects with chemotherapeutic
agents via T-cell activation and myeloid-derived suppressor cell
inhibition (16).
The present study conjugated a novel T7 and MUC1
peptide together (T7-MUC1) for use as a vaccine and examined its
immune responses and anti-tumor effects. It was hypothesized that
systemic administration of T7-MUC1 may induce antitumor immune
responses and elicit an antitumor effect in a mouse breast cancer
model by enhancing CTL activity and antibody-dependent
cell-mediated cytotoxicity (ADCC). In addition, it was speculated
that the therapeutic effect of T7-MUC1 may occur due to
non-specific anti-tumor responses elicited by the adjuvant T7, and
specific cellular and humoral immune responses elicited by the MUC1
peptide.
Materials and methods
Mice and cell lines
4T1 mouse breast cancer cells, MCF-7 human breast
cancer cells, MB231 human breast cancer cells and K562 human
leukemia cells (American Type Culture Collection) were cultured in
RPMI-1640 medium (K562 cells) or DMEM (4T1, MCF-7 and MB231 cells)
(both HyClone; Cytiva), supplemented with 10% FBS (HyClone;
Cytiva), 100 µg/ml penicillin and 100 µg/ml streptomycin (Gibco;
Thermo Fisher Scientific, Inc.) at 37°C in a humidified atmosphere
with 5% CO2. All experiments were performed with
mycoplasma-free cells.
Female 4-week-old BALB/c mice (n=150; weight, 15–20
g) were purchased from the Medical Laboratory Animal Centre of
Guangdong Province. All mice were housed in constant specific
pathogen-free laboratory conditions at 18–22°C and 50–60% humidity
with a 12 h light/dark cycle and ad libitum access to water
and food. The protocols of the animal experiments were approved by
the Laboratory Animal Ethics Committee of Shenzhen University
(approval no. AEWC-201712025).
Synthesis of T7 and T7-MUC1
The MUC1 peptide used in the present study is a
well-documented murine MUC1 epitope (8). T7 (SZU101), MUC1 and T7-MUC1 were
synthesized in our laboratory; MUC1 and T7-MUC1 were synthesized by
solid phase using an Fmoc strategy as previously described
(12,17). T7-K was prepared separately as an
amino acid for T7-MUC1, by first mixing the T7 activated ester with
lysine, and subsequently stirring for 4 h at room temperature. The
structure of T7 was determined as previously described (17).
Cytokine assay
Bone marrow dendritic cells (BMDCs) were generated
from the femurs and tibiae of one BALB/c mouse as previously
described (12), and were cultured
in X-vivo 15 medium (Lonza Group AC) with granulocyte-macrophage
colony-stimulating factor (GM-CSF) and IL-4 for 6 days. Spleen
lymphocytes were isolated from the same BALB/c mouse used to
generate BMDCs as previously described (12) using Mouse Lymphocyte Separation
Medium (Dakewe Biotech Co., Ltd.) and cultured in RPMI-1640 medium
with 10% FBS. Cells were seeded in 96-well plates at a density of
5×104 cells per well. T7-MUC1 was added at a final
concentration ranging between 0.1 and 100 µg/ml and incubated at
37°C for 24 h; T7 or MUC1 were used with an equal molar quantity to
T7-MUC1. Then, culture supernatants were collected, and the levels
of tumor necrosis factor (TNF)-α, interferon (IFN)-γ and IL-12 were
assessed using mouse TNF-α (cat. no. 88-7324-22), IFN-γ (cat. no.
88-7314-22) and IL-12 p70 (cat. no. 88-7121-22) ELISA kits
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions.
Immunizations and antibody
detection
All experiments were routinely performed in groups
of eight mice. On days 0, 14 and 28, the mice were
intraperitoneally (i.p.) administered PBS, 20 µg MUC1 peptide, 8.5
µg T7 or 28 µg T7-MUC1. A total of 0.2 ml blood was collected from
the tail of the mice 7 days after the last vaccination, and serum
was harvested from the blood for serologic assays. Anti-MUC1 IgG,
IgG1, IgG2a and IgM antibody titers were determined by ELISA as
previously described (18). ELISA
plates (Thermo Fisher Scientific, Inc.) were coated with the MUC1
peptide, and serial dilutions of the serum were allowed to bind to
immobilized MUC1. Detection was performed by adding alkaline
phosphatase-conjugated anti-mouse antibodies and phosphorylated
nitrophenyl phosphate (Sigma-Aldrich; Merck KGaA), and the optical
density was measured at 405 nm using a microplate reader.
Western blotting
Whole cell protein was extracted from K562, MCF-7
and MB231 cells using RIPA lysis buffer with 1 mM PMSF (Beyotime
Institute of Biotechnology), and protein concentrations were
detected using a bicinchoninic acid assay (Beyotime Institute of
Biotechnology). Proteins were separated by 10% SDS-PAGE and
transferred onto PVDF membranes. The membranes were first blocked
in 5% bovine serum albumin (Beyotime Institute of Biotechnology) at
room temperature for 1 h, and then incubated with rabbit anti-human
MUC1 (cat. no. 14161) or β-actin (cat. no. 4970) monoclonal
antibodies (1:1,000; Cell Signaling Technology, Inc.) at 4°C
overnight, and finally with goat anti-rabbit peroxidase
conjugated-secondary antibodies (cat. no. 7074; 1:2,000; Cell
Signaling Technology, Inc.) at room temperature for 1 h. Antibodies
bound to the blots were detected using the Clarity Western ECL
Substrate (Bio-Rad Laboratories, Inc.).
DC-cytokine-induced killer cell (CIK)
co-culture
Ethics approval was obtained for the use of human
tissues in the present study by the Medical Ethics Committee of the
Third Affiliated Hospital of Shenzhen University (approval no.
2019-SZLH-LW-009), and informed consent was provided by the healthy
donors. PBMCs (peripheral blood mononuclear cells) were isolated
from human blood samples of 3 male healthy donors (age range, 25–35
years; mean age, 30 years), and separated by density gradient
centrifugation at 400 × g for 30 min, according to the
manufacturer's instructions (Tianjin Haoyang Biological Co., Ltd.).
DCs and CIKs were cultured from PBMCs as previously described
(19). For the induction of CIKs,
non-adherent cells were cultured in RPMI-1640 medium containing
1,000 U/ml IFN-γ, 1,000 U/ml IL-2 and 100 ng/ml CD3 antibody
(PeproTech, Inc.) at 37°C for 7 days. Adherent cells were
differentiated to DCs using 1,000 U/ml GM-CSF, 500 U/ml IL-4 and
500 U/ml TNF-α (PeproTech, Inc.) for 7 days; MUC1 and the T7-MUC1
conjugate were added as an antigen on day 3 or 5. DCs and CIKs were
co-cultured between days 7 and 14 with a supplement of 1,000 U/ml
IL-2. The phenotype molecules of DCs
(CD80+/CD83+/CD86+) and CIKs
(CD3+/CD56+) were determined via flow
cytometry using a FACScalibur flow cytometer (BD Biosciences) and
the FlowJo v10 software (BD Biosciences). The mouse anti-human
CD80-FITC (cat. no. 305205), CD83-APC (cat. no. 305311),
CD86-PerCP/Cy5.5 (cat. no. 374215), CD3-FITC (cat. no. 300305) and
CD56-APC (cat. no. 362503) monoclonal antibodies (1:100) for flow
cytometry were purchased from BioLegend, Inc. On day 14, tumor
cells (MCF-7, MB231 and K562) and DC-CIKs were used as target cells
and effector cells, respectively, at an effector/target cell ratio
of 10:1 for 4 h. The levels of lactate dehydrogenase (LDH) released
by target cells in the supernatant were determined using a CytoTox
96 Non-Radioactive Cytotoxicity assay (Promega Corporation)
according to the manufacturer's instructions.
Evaluation of antitumor effects in
vaccinated mice
On day 21, mice were subcutaneously injected in the
mid-back region with 100 µl 4T1 cell suspension (1×106
cells/ml in PBS). The tumor dimensions were measured twice a week,
and tumor volume was calculated according to the equation Volume =
LxW2/2, where L is length (mm) of the long axis of the
tumor. On day 42, the mice were sacrificed by CO2
inhalation to minimize animal suffering, and the tumors were
surgically dissected, weighed and measured. Additionally, another
30 mice were used for the evaluation of the long-term survival
until the mice died naturally or the tumor diameter reached 15
mm.
Hematoxylin and eosin (H&E)
staining
The tumor tissues were cut into thin slices and
stained with a Hematoxylin and Eosin Staining kit, according to the
manufacturer's protocol (Beyotime Institute of Biotechnology) and
observed under a light microscope at ×200 magnification.
CTL assay
At the time of sacrifice, the spleens were removed
and lymphocytes were obtained by filtering the organs through a
sterile nylon mesh (70 µm). Then, 4T1 tumor cells used as target
cells and lymphocytes used as effect cells were seeded in 96-well
culture plates (Corning, Inc.) at an effector/target cell ratio of
50:1 at 37°C for 4 h. The levels of LDH released by the target
cells in the supernatants were determined as aforementioned.
Determination of ADCCs
4T1 tumor cells (5,000 cells/well) were incubated
with serum (1:20 dilution) obtained from the vaccinated mice for 30
min at 37°C. NK cells separated from the spleen were used as
effectors and were seeded with the antibody-labeled tumor cells in
96-well culture plates (Corning, Inc.) at an effector/target cell
ratio of 50:1 at 37°C for 4 h. The levels of LDH in the
supernatants were determined as aforementioned.
Flow cytometry
At the time of sacrifice, the mouse spleen was
collected, and splenocytes were prepared by removing the red blood
cells (RBCs) with RBC lysis buffer (BioLegend, Inc.) after
separating the cells by a 70-µm cell strainer. Subsequently,
~1×106 cells were stained with the corresponding
florescence antibodies and analyzed using a FACScalibur flow
cytometer (BD Biosciences) and the FlowJo v10 software (BD
Biosciences). The rat anti-mouse CD4-FITC (cat. no. 100406), CD8-PE
(cat. no. 100708) and CD3-APC (cat. no. 100236) monoclonal
antibodies (1:100 dilution) for flow cytometry were purchased from
BioLegend, Inc.
Statistical analysis
Data are presented as the mean ± SEM from the
indicated number of independently performed experiments. Two-way
ANOVA with Bonferroni post hoc test was used to compare the tumor
volumes in different groups collected over all time points. One-way
ANOVA with Bonferroni post hoc test was used for the determination
of statistical significance for all other experiments. P<0.05
was considered to indicate a statistically significant
difference.
Results
Synthesis of the T7-MUC1
conjugate
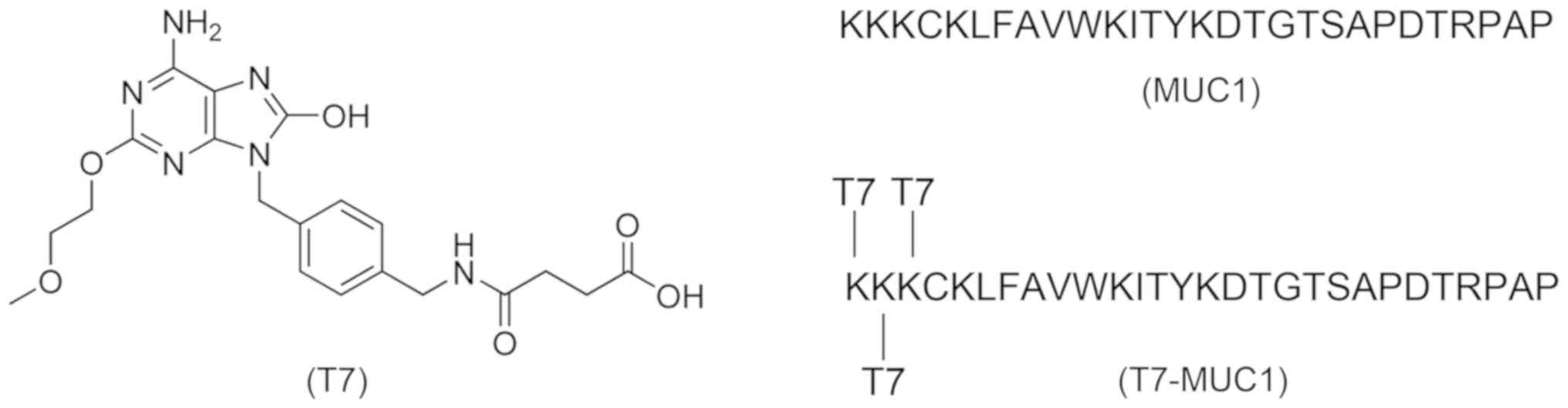
Structures of all of the vaccines used in this study
are presented in Fig. 1. T7
(Fig. S1A) was synthesized in the
laboratory following a previously described procedure (17), with the 1H and
13C NMR being consistent with the assigned structure
(Fig. S2A and B). The certificates
of the T7-MUC1 conjugate are presented in Fig. S3A and B, and the ratio of the T7
conjugate to MUC1 was 3:1 (Fig.
S1B).
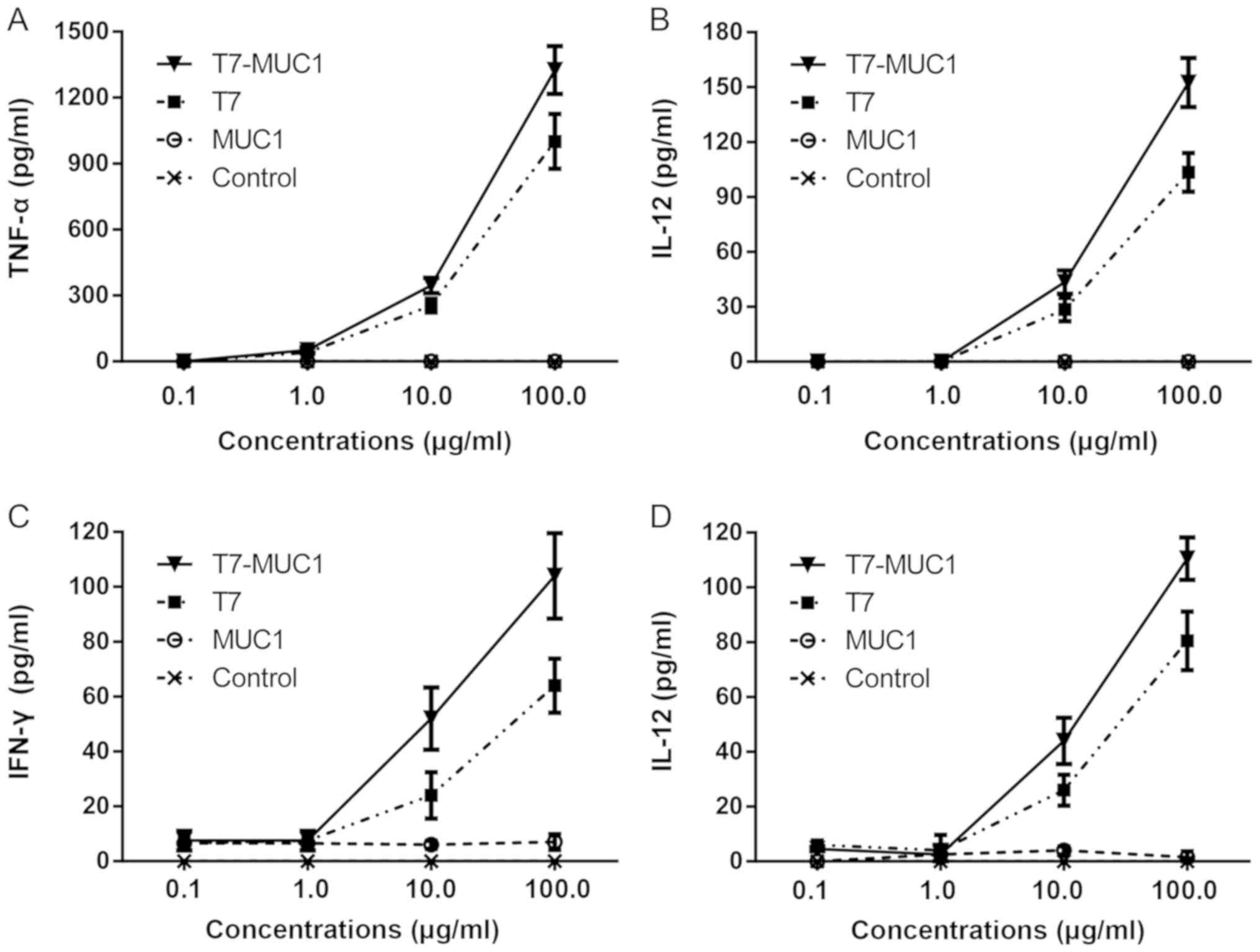
In vitro cytokine release in response
to the T7-MUC1 conjugate
In order to evaluate the immunological activity of
T7-MUC1, BMDCs and spleen-derived lymphocytes were incubated with
different concentrations of T7, MUC1 and T7-MUC1, and ELISA was
used to determine the release of cytokines. The results indicated
that the levels of IL-12 and TNF-α remained unchanged when BMDCs
were incubated with MUC1 alone (Fig. 2A
and B). However, the levels of IL-12 and TNF-α in the T7-MUC1
and T7 groups were significantly higher compared with those in the
control group. The results also demonstrated that the levels of
IL-12 and TNF-α released by BMDCs in the T7-MUC1 group were
significantly increased compared those in with the T7 group. A
similar trend was observed for the release of IL-12 and IFN-γ in
spleen lymphocytes (Fig. 2C and
D).
T7-MUC1 conjugate induces IgG and IgM
antibody responses
The anti-MUC1 IgG, IgG1, IgG2a and IgM antibody
titers were determined by ELISA. The results demonstrated that T7
and MUC1 alone had no effect on IgG and IgM antibody responses, but
the T7-MUC1 conjugate significantly elicited the IgG total, IgG1,
IgG2a and IgM antibody responses (Table
I).
 | Table I.ELISA anti-MUC1 antibody titers after
three immunizations with T7-MUC1. |
Table I.
ELISA anti-MUC1 antibody titers after
three immunizations with T7-MUC1.
| Treatment | IgG total | IgM | IgG1 | IgG2a |
|---|
| PBS | 0 | 0 | 0 | 0 |
| T7 | 55 | 13 | 8 | 15 |
| MUC1 | 1,719 | 303 | 920 | 688 |
| T7-MUC1 | 11,147 | 9,552 | 12,890 | 13,257 |
In vitro cytolytic response in the
high MUC1 protein-expressing cells is induced by the T7-MUC1
conjugate
To observe the antigen effect of the T7-MUC1
conjugate, the present study developed a DC-CIK co-culture
procedure using human samples. DCs and CIKs were cultured
separately, and the T7-MUC1 conjugate was added as an antigen on
day 3 and 5 to the DC culture. Then, DCs and CIKs were co-cultured
on day 7, and the CTL effect was measured on day 14. Western
blotting results for the three types of human tumor cells, MCF-7,
MB231 and K562, demonstrated high protein expression levels of MUC1
in MCF-7 cells, whereas K562 and MB231 cells exhibited relatively
low MUC1 expression; MB231 cells expressed undetectable levels of
MUC1 protein (Fig. 3A). The CTL
results demonstrated that the conjugate-induced CTLs exerted a more
potent effect on MCF-7, but not K562 and MB231 cells (Fig. 3B-D). Therefore, it was speculated
that the conjugate-induced CTLs exerted increased target cell
cytotoxicity, and this effect was partially dependent on the
expression of MUC1 in the target cells.
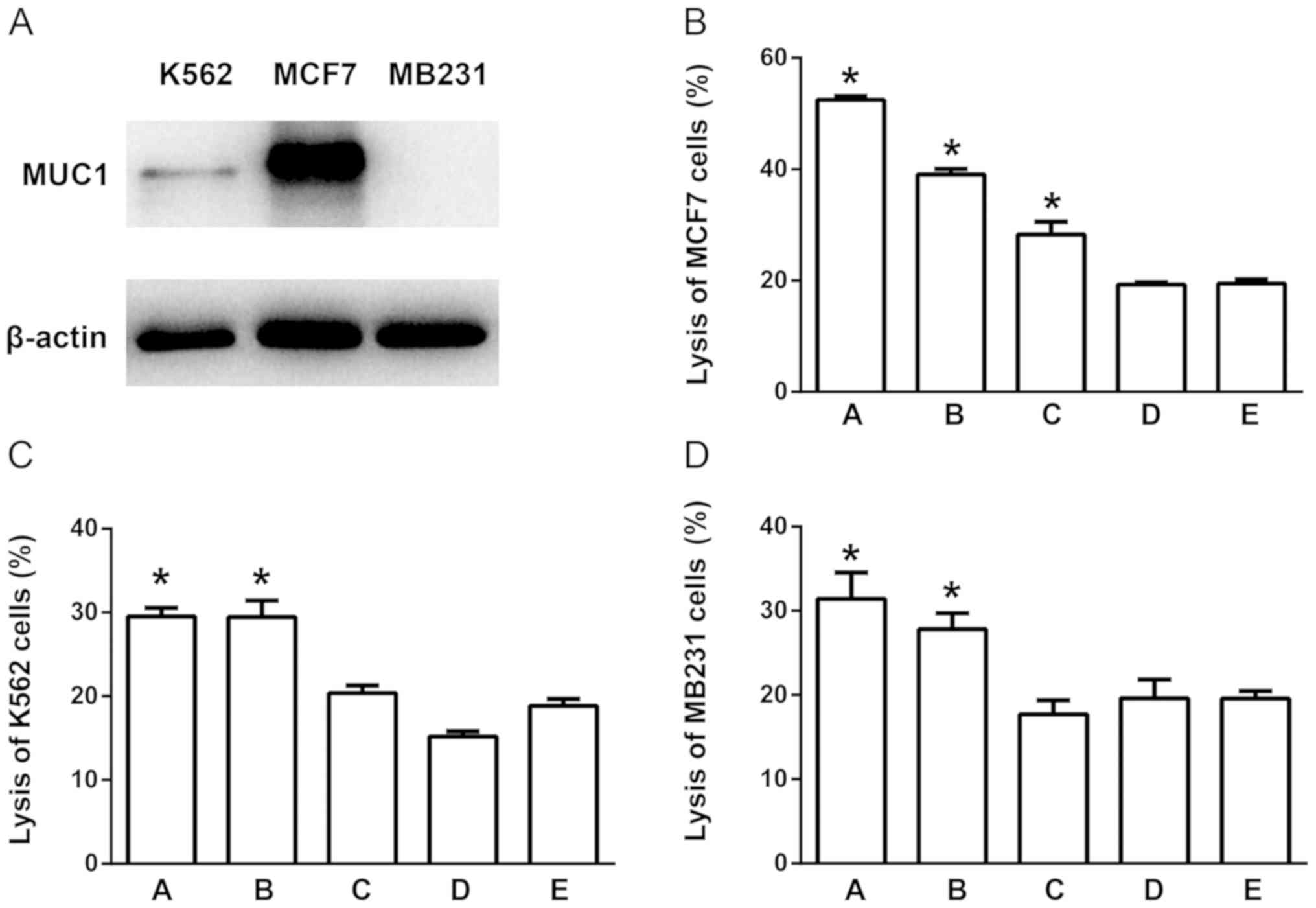 | Figure 3.T7-MUC1 conjugate induces a cytolytic
response in cells expressing high levels of MUC1 protein. (A) MUC1
expression in MCF-7, MB231 and K562 cells. (B-D) Induction of the
CTL effect on (B) MCF-7, (C) K562 and (D) MB231 cells. The groups
are as follows: A, T7-MUC1 conjugate added on day 3; B, MUC1 added
on day 3; C, T7-MUC1-conjugate added on day 5; D, MUC1 added on day
5; and E, control with no added antigen. Data are presented as the
mean ± SD; n=5. *P<0.05 vs. E. MUC1, mucin 1; T7, Toll-like
receptor 7 agonist; PBMCs, peripheral blood mononuclear cells; CTL,
cytotoxic T lymphocytes. |
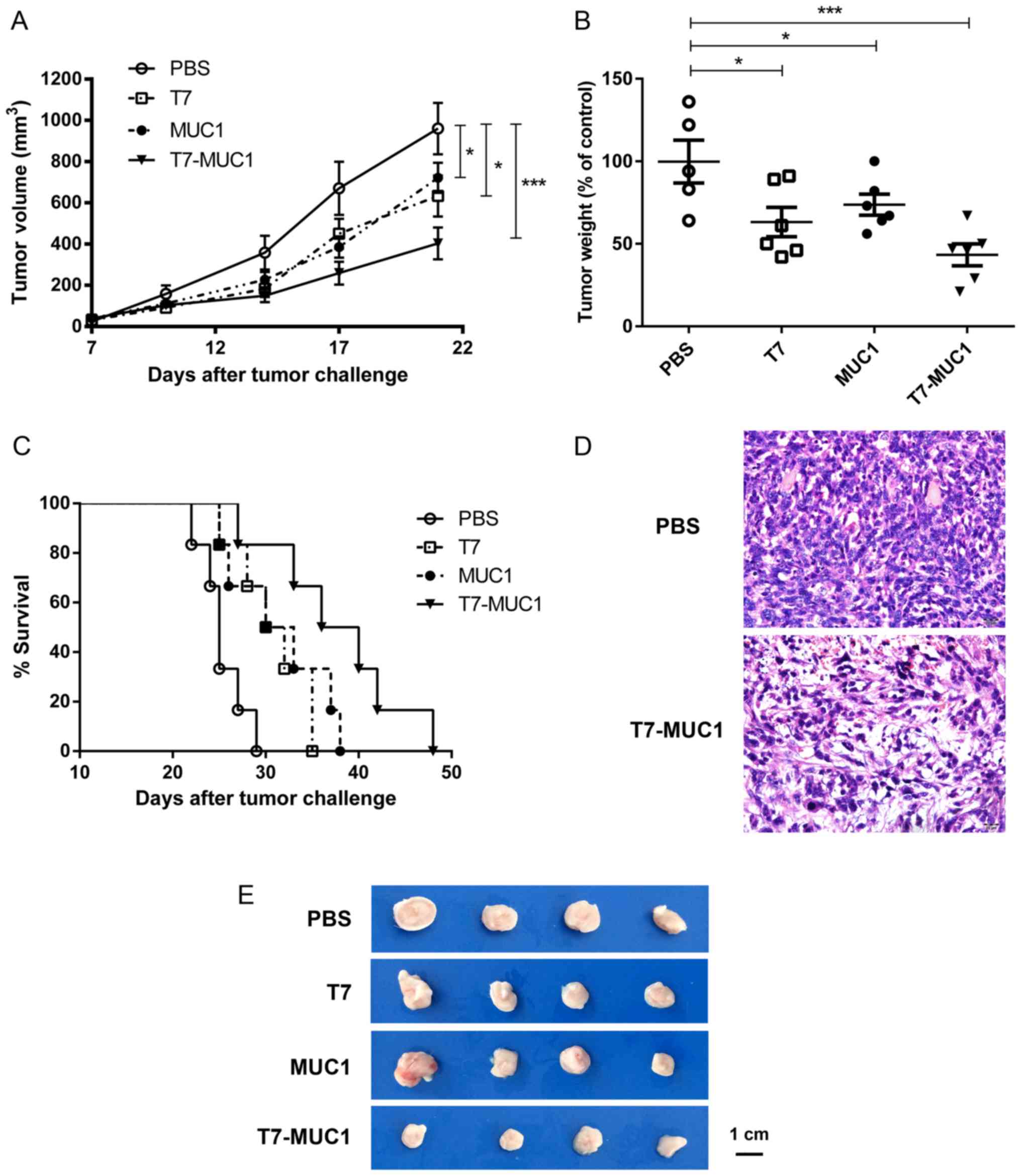
Antitumor effect of the T7-MUC1
conjugate in a mouse breast cancer model
To assess the antitumor effect of the T7-MUC1
conjugate, BALB/c mice were injected subcutaneously with 4T1 mouse
breast cancer cells after two immunizations. Then, 21 days after
the 4T1 cell injection, the mice were sacrificed, and the tumors
were dissected, weighed and measured. The results demonstrated a
significant growth inhibition in the T7, MUC1 and conjugate-treated
groups compared with that in the control group (P<0.05; Fig. 4A, B and E). Of note, compared with
the PBS control, tumor weight was lower (43.3%) after treatment
with T7-MUC1-conjugate than with T7 (63.2%) or MUC1 (73.7%) alone
(Fig. 4B). The results also
demonstrated that the T7-MUC1 conjugate group exhibited a
significantly increased long-term survival rate compared with that
in the other groups during a 50-day observation period (Fig. 4C). Tumor tissues were also examined
by H&E staining, and it was identified that the T7-MUC1
conjugate exerted an antitumor effect in the mouse breast cancer
model (Fig. 4D).
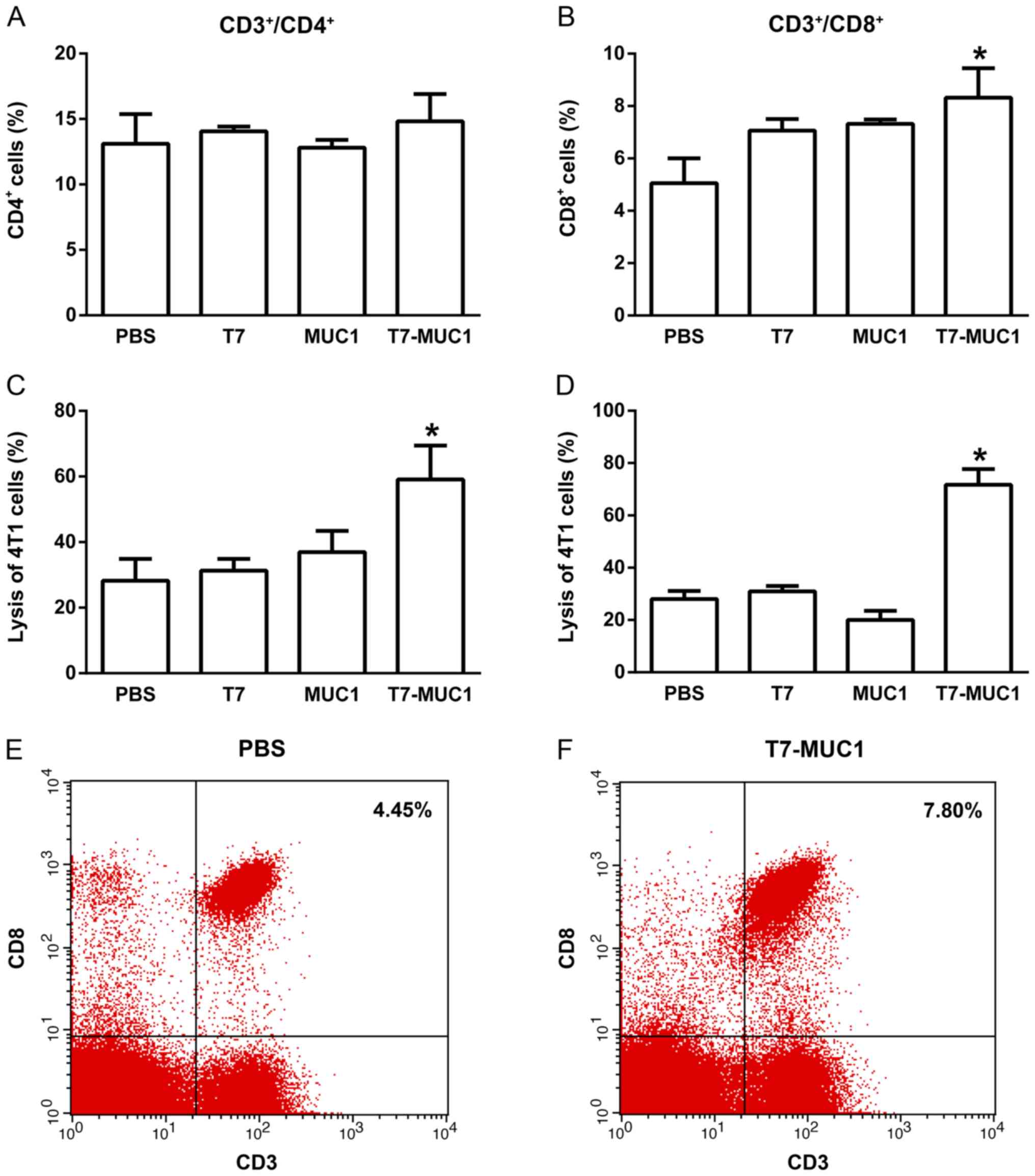
T7-MUC1 conjugate induces
tumor-specific immune responses
To determine the capacity of the T7-MUC1 vaccine to
elicit a tumor-specific immune response, lymphocytes were isolated
from the spleen in the different groups, and the percentages of
CD3+/CD4+ and CD3+/CD8+
T cells were measured by flow cytometry. No changes in the
percentages of CD3+/CD4+ T cells were
observed among the groups (Fig. 5A).
However, the percentages of CD3+/CD8+ T cells
were significantly higher in the T7-MUC1 group compared with in the
PBS control group (Fig. 5B).
Representative flow cytometry results of
CD3+/CD8+ T-cells are demonstrated for the
PBS control (Fig. 5E) and T7-MUC1
conjugate (Fig. 5F) groups. Further
representative flow cytometry results are presented in Fig. S4. Regarding ADCCs and CTLs, the
cytotoxicity effects in 4T1 cells induced by NK cells following the
addition of antisera or by lymphocytes in the T7-MUC1 group were
significantly higher compared with those in the other groups
(Fig. 5C and D).
Discussion
Breast cancer is a major cause of cancer mortality
among women, especially in the least developed countries, and the
risk factors include reproductive and endocrine dysfunction,
obesity and physical inactivity (20). Traditional treatments of breast
cancer include surgery, chemotherapy, radiotherapy and
endocrinotherapy (20). Currently,
immunotherapy is a promising novel therapeutic strategy due to its
advantages such as effective targeting and limited side effects
(21). The immune system can
recognize, process and present tumor antigens, leading to the
cross-priming and activation of T cells and tumor elimination
(21). In addition, multiple
tumor-associated antigens, such as human epidermal growth factor
receptor and MUC1, are targets of novel drugs developed for breast
cancer vaccines, a number of which have been demonstrated to be
clinically beneficial (22).
In recent years, multimodal cancer vaccines have
received increased attention due to their ability to simultaneously
stimulate different aspects of the immune system (23). However, it has been revealed to be
difficult to obtain vaccines with both cellular and humoral immune
responses against MUC1, although either of these responses are
associated with a satisfactory outcome in clinical cancer treatment
(24,25). It has also been demonstrated that
patients with breast cancer who are vaccinated with a 30-amino acid
sequence of MUC1 peptide conjugated with keyhole limpet hemocyanin
(KLH) and the immune adjuvant QS-21 produce high IgG and IgM
antibody titers to MUC1 and high response to KLH, but the T-cell
response against MUC1 peptide is minimal and inconsistent (26,27).
Furthermore, a previous study has reported that lack of
glycosylation in MUC1 peptides used in vaccines leads to
ineffective immune responses (22).
In the present study, following conjugation with T7, the MUC1
peptide without the glycosylated structure generated an overall
therapeutic response, suggesting that the conjugation of T7 and
MUC1 may be a potential effective vaccine against breast
tumors.
TLR ligands are widely used as adjuvants in the
design of vaccines to boost the immunogenicity of antigens,
producing strong and long-lasting immunity. For example, TLR2
peptide-overexpressing Bacille Calmette-Guérin triggers MHC
II-dependent antigen presentation and elicits T helper (Th) 1
cytokine release in vitro, leading to increased expansion of
the effector and central memory T cells in vivo (28). Furthermore, Maynard et al
(29) have reported that CpG, a TLR9
agonist, may be a potent adjuvant to a synthetic long peptide
containing the human papilloma virus 16 E7 antigen, which can
induce multifunctional antigen specific CD8+ T cells
with prophylactic and therapeutic benefits in a murine lung tumor
model. Previous studies have also demonstrated that the conjugation
of a TLR ligand and an antigen induces improved immunogenicity
compared with a simple mixture of them, which is due to the fact
that the distribution and metabolism of TLR ligand and antigen may
be more systemically coincident in the design of a conjugate
(8,18). Our previous study revealed the
necessity of a covalent attachment in the design of tumor vaccines
containing T7 and tumor-associated antigens (12,16).
Therefore, in the present study, a similar strategy was used to
construct the T7-MUC1 vaccine by conjugating T7 to lysine and
linking it to the N-terminal of MUC1 peptide at a ratio of 3:1.
The results of the present study suggested that the
T7-MUC1 conjugate caused a rapid induction of inflammatory
mediators TNF-α, IFN-γ and IL-12 in mouse BMDCs and spleen
lymphocytes in vitro, indicating the activation of innate
immunity. As TNF-α, IFN-γ and IL-12 are Th1 cytokines (30), these results suggested that TLR7
activation may lead to Th1-biased immune responses. Furthermore,
TLR-produced cytokines stimulate the expression of a number of
co-stimulatory molecules, promoting the optimal performance of T
and B cells (8). Additionally, IgG
and IgM are two types of Ig produced by B cells based on a specific
antigen, thus mediating complement-dependent cytotoxicity and ADCC
activity (31). The results of the
present study demonstrated that the T7-MUC1 vaccine elicited
anti-MUC1 IgG and IgM antibodies in the immunized mice and was more
effective compared with T7 or MUC1 alone. Therefore, the present
results indicated that the covalent attachment of T7 facilitated
selective internalization by TLR7-expressing immune cells, such as
B cells and APCs.
Following mouse immunization with the vaccines, the
present study generated a tumor challenge model in BALB/c mice via
subcutaneous implantation of mouse 4T1 breast cancer cells in order
to investigate the in vivo antitumor effects of T7-MUC1. The
results demonstrated that the T7-MUC1 conjugate exhibited the
highest prophylactic performance among the tested vaccines for
tumor treatment, and several aspects of tumor-specific immune
responses were further examined. The results indicated that
significant cytotoxicity to 4T1 tumor cells was caused by the
antisera collected from mice immunized with T7-MUC1 with the
presence of NK cells. In addition, the results of ADCC activity
were similar to those of the IgG and IgM titers, suggesting the
activation on humoral immunity by the T7-MUC1 vaccine. In terms of
cellular immunity, CTL activity was identified to exhibit similar
outcomes of tumor cell lysis as ADCC activity. Among the subsets of
T cells, CD4+ and CD8+ T cells serve
essential roles in the host defense against different types of
cancer, such as melanoma (32).
Naïve CD4+ T cells can differentiate into a variety of
subsets, including Th1, Th2, Th9, Th17 and T regulatory cells,
whereas naïve CD8+ T cells differentiate into memory and
effector CD8+ T cells (33). In line with the CTL results, T7-MUC1
increased the proportions of CD3+/CD8+ T
cells, but not CD3+/CD4+ T-cells, in
splenocytes, which was expected as CTLs belong to the
CD8+ T-cell subset. Therefore, the present results may
facilitate the development of a MUC1-derived vaccine, which may
initiate a response against residual cancer cells, thus improving
long-term survival.
DC-CIK adoptive cellular immunotherapy is
significant due to its strong antitumor activity against a broad
spectrum of solid tumor types, including breast cancer, as DCs are
stimulators of tumor-specific T cell responses, and CIKs are ex
vivo-expanded T lymphocytes with a NK/T-cell phenotype
(34,35). The present study used an ex
vivo model to examine the antitumor mechanisms of T7-MUC1; DCs
were treated with the vaccines, followed by co-culture of
antigen-loaded DCs and CIKs, and their toxicity to tumor cells was
tested using the LDH method. The present results demonstrated that
T7-MUC1 significantly induced the DC-CIK response in human breast
cancer MCF-7 cells with high MUC1 expression, but not in those with
low (K562) or no (MB231) MUC1 expression. The results also
demonstrated that the antigen was more effective when added on day
3 compared with day 5. Collectively, the results of the present
study demonstrated the selectivity and specificity of T7-MUC1 for
the application of cellular immunotherapy in breast cancer.
In conclusion, the novel T7 was conjugated with the
unglycosylated MUC1 peptide to develop a safe and effective
immunotherapeutic vaccine against breast cancer. The results of the
present study demonstrated that T7 elicited non-specific immune
responses and strengthened the specific humoral and cellular immune
responses to the MUC1 antigen. In addition, the vaccine induced
CTLs and ADCC-mediating antibodies recognizing MUC1, and overall
exhibited a potential to inhibit tumor growth. Therefore, this
vaccine candidate may have beneficial effects for the prevention of
tumor recurrence in patients with breast cancer.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the Guangdong
Science and Technology Department (grant nos. 2017A030310400 and
2018A0303130225) and the Shenzhen Science and Technology Innovation
Commission (grant nos. JCYJ20180305163318492 and
JSGG20160331161046511).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XW, GJ, YL and LT designed the study. YL, LT, NG,
YWD and JZ performed the experiments. YL, LT, NG, YQD and ZW
analyzed the data. XW, GJ, YL and LT wrote the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Ethics approval was obtained for the use of human
tissues in the present study by the Medical Ethics Committee of the
Third Affiliated Hospital of Shenzhen University (approval no.
2019-SZLH-LW-009), and informed consent was provided by the healthy
donors. The protocols of animal experiments were approved by the
Laboratory Animal Ethics Committee of Shenzhen University (approval
no. AEWC-201712025).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Curigliano G, Spitaleri G, Pietri E,
Rescigno M, de Braud F, Cardillo A, Munzone E, Rocca A, Bonizzi G,
Brichard V, et al: Breast cancer vaccines: A clinical reality or
fairy tale? Ann Oncol. 17:750–762. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sangha R and Butts C: L-BLP25: A peptide
vaccine strategy in non small cell lung cancer. Clin Cancer Res. 13
(Suppl):S4652–S4654. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ninkovic T and Hanisch FG: O-glycosylated
human MUC1 repeats are processed in vitro by immunoproteasomes. J
Immunol. 179:2380–2388. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Roulois D, Grégoire M and Fonteneau JF:
MUC1-specific cytotoxic T lymphocytes in cancer therapy: Induction
and challenge. Biomed Res Int. 2013:8719362013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Taylor-Papadimitriou J, Burchell JM,
Graham R and Beatson R: Latest developments in MUC1 immunotherapy.
Biochem Soc Trans. 46:659–668. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Scheid E, Major P, Bergeron A, Finn OJ,
Salter RD, Eady R, Yassine-Diab B, Favre D, Peretz Y, Landry C, et
al: Tn-MUC1 DC vaccination of rhesus macaques and a phase I/II
trial in patients with nonmetastatic castrate-resistant prostate
cancer. Cancer Immunol Res. 4:881–892. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tosch C, Bastien B, Barraud L, Grellier B,
Nourtier V, Gantzer M, Limacher JM, Quemeneur E, Bendjama K and
Préville X: Viral based vaccine TG4010 induces broadening of
specific immune response and improves outcome in advanced NSCLC. J
Immunother Cancer. 5:702017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lakshminarayanan V, Thompson P, Wolfert
MA, Buskas T, Bradley JM, Pathangey LB, Madsen CS, Cohen PA,
Gendler SJ and Boons GJ: Immune recognition of tumor-associated
mucin MUC1 is achieved by a fully synthetic aberrantly glycosylated
MUC1 tripartite vaccine. Proc Natl Acad Sci USA. 109:261–266. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Braunstein MJ, Kucharczyk J and Adams S:
Targeting toll-like receptors for cancer therapy. Target Oncol.
13:583–598. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhao S, Gao N, Qi H, Chi H, Liu B, He B,
Wang J, Jin Z, He X, Zheng H, et al: Suppressive effects of
sunitinib on a TLR activation-induced cytokine storm. Eur J
Pharmacol. 854:347–353. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Coffman RL, Sher A and Seder RA: Vaccine
adjuvants: Putting innate immunity to work. Immunity. 33:492–503.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang XD, Gao NN, Diao YW, Liu Y, Gao D, Li
W, Wan YY, Zhong JJ and Jin GY: Conjugation of toll-like receptor-7
agonist to gastric cancer antigen MG7-Ag exerts antitumor effects.
World J Gastroenterol. 21:8052–8060. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Vascotto F, Petschenka J, Walzer KC,
Vormehr M, Brkic M, Strobl S, Rosemann R, Diken M, Kreiter S,
Töreci Ö and Sahin U: Intravenous delivery of the toll-like
receptor 7 agonist SC1 confers tumor control by inducing a
CD8+ T cell response. Oncoimmunology. 8:16014802019.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim H, Sehgal D, Kucaba TA, Ferguson DM,
Griffith TS and Panyam J: Acidic pH-responsive polymer
nanoparticles as a TLR7/8 agonist delivery platform for cancer
immunotherapy. Nanoscale. 10:20851–20862. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gao D, Liu Y, Diao Y, Gao N, Wang Z, Jiang
W and Jin G: Synthesis and evaluation of conjugates of novel TLR7
inert ligands as self-adjuvanting immunopotentiators. ACS Med Chem
Lett. 6:249–253. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang X, Liu Y, Diao Y, Gao N, Wan Y, Zhong
J, Zheng H, Wang Z and Jin G: Gastric cancer vaccines synthesized
using a TLR7 agonist and their synergistic antitumor effects with
5-fluorouracil. J Transl Med. 16:1202018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhu J, He S, Du J, Wang Z, Li W, Chen X,
Jiang W, Zheng D and Jin G: Local administration of a novel
Toll-like receptor 7 agonist in combination with doxorubicin
induces durable tumouricidal effects in a murine model of T cell
lymphoma. J Hematol Oncol. 8:212015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lin G, Wang X, Yi W, Zhang C, Xu G, Zhu X,
Cai Z, Liu Y, Diao Y, Lin MC and Jin G: A conjugate of
octamer-binding transcription factor 4 and toll-like receptor 7
agonist prevents the growth and metastasis of testis embryonic
carcinoma. J Transl Med. 13:1662015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Qiao G, Wang X, Zhou L, Zhou X, Song Y,
Wang S, Zhao L, Morse MA, Hobeika A, Song J, et al: Autologous
dendritic cell-cytokine induced killer cell immunotherapy combined
with S-1 plus cisplatin in patients with advanced gastric cancer: A
prospective study. Clin Cancer Res. 25:1494–1504. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Akram M, Iqbal M, Daniyal M and Khan AU:
Awareness and current knowledge of breast cancer. Biol Res.
50:332017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang Y: Cancer immunotherapy: Harnessing
the immune system to battle cancer. J Clin Invest. 125:3335–3337.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mohit E, Hashemi A and Allahyari M: Breast
cancer immunotherapy: Monoclonal antibodies and peptide-based
vaccines. Expert Rev Clin Immunol. 10:927–961. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Morse MA and Whelan M: A year of
successful cancer vaccines points to a path forward. Curr Opin Mol
Ther. 12:11–13. 2010.PubMed/NCBI
|
|
24
|
Cazet A, Julien S, Bobowski M, Burchell J
and Delannoy P: Tumour-associated carbohydrate antigens in breast
cancer. Breast Cancer Res. 12:2042010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Beatson RE, Taylor-Papadimitriou J and
Burchell JM: MUC1 immunotherapy. Immunotherapy. 2:305–327. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Adluri S, Gilewski T, Zhang S, Ramnath V,
Ragupathi G and Livingston P: Specificity analysis of sera from
breast cancer patients vaccinated with MUC1-KLH plus QS-21. Br J
Cancer. 79:1806–1812. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Musselli C, Ragupathi G, Gilewski T,
Panageas KS, Spinat Y and Livingston PO: Reevaluation of the
cellular immune response in breast cancer patients vaccinated with
MUC1. Int J Cancer. 97:660–667. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Khan A, Bakhru P, Saikolappan S, Das K,
Soudani E, Singh CR, Estrella JL, Zhang D, Pasare C, Ma Y, et al:
An autophagy-inducing and TLR-2 activating BCG vaccine induces a
robust protection against tuberculosis in mice. NPJ Vaccines.
4:342019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Maynard SK, Marshall JD, MacGill RS, Yu L,
Cann JA, Cheng LI, McCarthy MP, Cayatte C and Robbins SH:
Vaccination with synthetic long peptide formulated with CpG in an
oil-in-water emulsion induces robust E7-specific CD8 T cell
responses and TC-1 tumor eradication. BMC Cancer. 19:5402019.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Habijanic J, Berovic M, Boh B, Plankl M
and Wraber B: Submerged cultivation of Ganoderma lucidum and the
effects of its polysaccharides on the production of human cytokines
TNF-α, IL-12, IFN-γ, IL-2, IL-4, IL-10 and IL-17. N Biotechnol.
32:85–95. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sánchez Ramírez J, Morera Díaz Y,
Bequet-Romero M, Hernandez-Bernal F, Selman-Housein Bernal KH, de
la Torre Santos A, Santiesteban Álvarez ER, Martín Bauta Y,
Bermudez Badell CH, de la Torre Pupo J, et al: Characteristics of
the specific humoral response in patients with advanced solid
tumors after active immunotherapy with a VEGF vaccine, at different
antigen doses and using two distinct adjuvants. BMC Immunol.
18:392017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mei Y, Zhao L, Liu Y, Gong H, Song Y, Lei
L, Zhu Y, Jin Z, Ma S, Hu B, et al: Combining DNA vaccine and
AIDA-1 in attenuated salmonella activates tumor-specific
CD4+ and CD8+ T-cell responses. Cancer
Immunol Res. 5:503–514. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jiang S and Yan W: T-cell immunometabolism
against cancer. Cancer Lett. 382:255–258. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lin M, Liang S, Jiang F, Xu J, Zhu W, Qian
W, Hu Y, Zhou Z, Chen J, Niu L, et al: 2003-2013, a valuable study:
Autologous tumor lysate-pulsed dendritic cell immunotherapy with
cytokine-induced killer cells improves survival in stage IV breast
cancer. Immunol Lett. 183:37–43. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhao Y, Qiao G, Wang X, Song Y, Zhou X,
Jiang N, Zhou L, Huang H, Zhao J, Morse MA, et al: Combination of
DC/CIK adoptive T cell immunotherapy with chemotherapy in advanced
non-small-cell lung cancer (NSCLC) patients: A prospective
patients' preference-based study (PPPS). Clin Transl Oncol.
21:721–728. 2019. View Article : Google Scholar : PubMed/NCBIPubMed/NCBIPubMed/NCBIPubMed/NCBI
|