Introduction
Despite recent advances in cancer prevention, lung
cancer remains the most common malignant cancer type and the
leading cause of cancer-associated mortality worldwide (1). Overall, the 5-year survival rate of
patients with lung cancer at all stages is ~15%, and this survival
rate has not been significantly improved over previous years
(2). The high mortality rate is
primarily a result of the lack of tools available to facilitate
early diagnosis and the lack of curative therapies (3). Non-small cell lung cancer (NSCLC) is
the major subtype of lung cancer and accounts for >85% of all
cases (4). Smoking is a major risk
factor of NSCLC; however, this disease also affects non-smokers
(5). In addition, smoking itself is
insufficient to induce the occurrence of NSCLC (5).
Besides smoking, the tumorigenesis and progression
of NSCLC are closely associated with certain genetic factors
(6,7). Phosphatase and tensin homolog (PTEN) is
a tumor-suppressive gene with pivotal roles in cell cycle
regulation (8). In cancer biology,
PTEN inhibits cancer cells from proliferating rapidly,
predominantly via inhibition of the PI3K/AKT signaling pathway
(9). Certain oncogenic microRNAs
(miR/miRNA), such as miR-221, target PTEN to promote cancer
progression (10). In a recent
study, Li et al (11)
identified a novel long non-coding RNA (lncRNA), zinc finger
protein (ZNF)281, which serves a tumor-suppressive role in glioma
via the inhibition of the NF-κB signaling pathway, and it has been
revealed that the NF-κB signaling pathway interacts directly with
miR-221 (12). Therefore, ZNF281 may
interact with miR-221. The present study aimed to investigate the
interactions between ZNF281 and miR-221 and the consequent effects
on PTEN.
Materials and methods
Patients
A total of 66 patients were selected from the 182
patients with NSCLC admitted to Hiser Medical Center of Qingdao
(Shandong, China) between January 2012 and April 2014. The present
study was approved by the Review Board of Hiser Medical Center and
the Qingdao Ethics Committee. The 66 patients with NSCLC comprised
30 cases of squamous cell carcinoma and 36 cases of adenocarcinoma.
The inclusion criteria were as follows: i) Diagnosed for the first
time; and ii) no treatment had been received before admission. The
exclusion criteria were as follows: i) Recurrent NSCLC; ii)
clinical disorders other than NSCLC were diagnosed; iii) therapy
had already been initiated; and iv) patients who failed to complete
the follow-up or who died from other diseases or accidents.
According to the clinical findings, the 66 patients included 6, 19,
20 and 21 cases at clinical stage I–IV, respectively (13). According to cancer histologic grade,
there were 14, 19, 20 and 13 cases at grade 1–4, respectively
(14). All patients were informed of
the principle of the present study. Written informed consent was
provided by all 66 patients.
Follow-up
Starting from the day of admission, all 66 patients
were followed-up for 5 years. Their survival conditions were
monitored and recorded through monthly telephone calls.
NSCLC cells and tissues
The H1993 human NSCLC cell line (American Type
Culture Collection) was used in the present study. Cells were
cultured in a mixture of 90% RPMI-1640 medium (Sigma-Aldrich; Merck
KGaA) and 10% FBS (Sigma-Aldrich; Merck KGaA) supplemented with 1%
penicillin-streptomycin (Sigma-Aldrich; Merck KGaA). Cell were
cultured at 37°C with 5% CO2 and 95% humidity.
All 66 patients with NSCLC received lung biopsy.
During biopsy, adjacent (≤2 cm from tumor) non-tumorous lung
tissues and NSCLC tissues were obtained from each patient. Based on
histopathological examination results, all non-tumor tissues
contained <1% cancerous cells, and all NSCLC tissues contained
>98% cancerous cells. Fresh tissues were stored in liquid
nitrogen.
Cell transfections
Expression vectors of ZNF281 and PTEN were
constructed using pcDNA3.1 (Sangon Biotech Co., Ltd.). Negative
control (NC) miRNA (5′-UGUGGUUACGAUCGUGGGAACUG-3′) and miR-221
(5′-ACCUGGCAUACAAUGUAGAUUU-3′) were purchased from Guangzhou
RiboBio Co., Ltd. Prior to transfections, H1993 cells were
harvested at a confluency of 70–80%. Lipofectamine 2000®
(Sangon Biotech Co., Ltd.) was used to transfect 40 nM miRNA (NC
miRNA as NC group) or 10 nM vector (empty vector as NC group) into
1×106 cells. Untransfected cells were used as the
control (C) group. Cells were harvested at 24 h post-transfection
to perform all subsequent experiments.
RNA extraction and reverse
transcription-quantitative (RT-q)PCR
H1993 cells were collected at 24 h
post-transfection. Total RNA in 1×105 cells and 0.02 g
tissue sample (ground in liquid nitrogen) was extracted using
Ribozol reagent (Sigma-Aldrich; Merck KGaA). In order to harvest
miRNAs, 80% ethanol was used to precipitate and wash RNA
samples.
All RNA samples were digested with DNase I
(Sigma-Aldrich; Merck KGaA) at 37°C for 2 h to remove genomic DNAs.
All reverse transcriptions were performed using the PrimeScript RT
Reagent kit (Takara Bio, Inc.) to synthesize cDNA, followed by
preparation of qPCR mixtures using the QuantiTect SYBR-Green PCR
kit (Qiagen) according to the manufacturer's instructions with
GAPDH as an endogenous control to measure the expression levels of
ZNF281 and PTEN mRNA.
To measure the expression levels of miR-221, both
reverse transcriptions and qPCR mixture preparations were prepared
using the All-in-One™ miRNA RT-qPCR Detection kit (GeneCopoeia,
Inc.) with U6 as an endogenous control. The sequences of primers
were: ZNF281 forward, 5′-GGACACATAGTGGAGAAAAG-3′ and reverse,
5′-GAGACAACACAGCCAGATTA-3′; PTEN forward, 5′-TGAGTTCCCTCAGCCGT-3′
and reverse, 5′-GAGGTTTCCTCTGGTCC-3′; and GADPH forward,
5′-GGATTTGGTCGTATTGG-3′ and reverse, 5′-GGAAGATGGTGATGGGAT-3′. The
forward primer for miR-221 was: 5′-ACCUGGCAUACAAUGUAG-3′. Reverse
primers for miR-221 and U6 primers were from the All-in-One™ miRNA
RT-qPCR Detection kit (cat. no. QP015, GeneCopoeia, Inc.). The PCR
conditions were as follows: 95°C for 1 min, and then 40 cycles of
95°C for 10 sec and 55°C for 60 sec. All experiments were performed
in three technical replicates and the 2−ΔΔCq method was
used to analyze data (15).
Cell apoptosis analysis
H1993 cells were collected at 24 h post-transfection
and counted. Subsequently, 4×104 cells were mixed with 1
ml serum-free RPMI-1640 medium to prepare single-cell suspensions.
Cells were cultivated in a 6-well cell culture plate (2 ml/well)
for 48 h at 37°C. Subsequently, pre-cooled PBS was used to wash
cells, followed by propidium iodide and Annexin V-FITC staining for
20 min at 4°C. Finally, flow cytometry was performed using CytoFLEX
LX Flow Cytometer (Beckman Coulter) to separate apoptotic cells.
CellQuest Pro v5.1 software (BD Biosciences) was used to analyze
data.
Cell proliferation analysis
H1993 cells were collected at 24 h post-transfection
and counted. Subsequently, 4×104 cells were mixed with 1
ml RPMI-1640 medium (10% FBS) to prepare single-cell suspensions.
Cells were seeded in a 96-well cell culture plate (0.1 ml/well),
followed by the addition of 10 µl Cell Counting Kit-8 (Dojindo
Molecular Technologies, Inc.) for 4 h. Optical density was measured
at 450 nm on a microplate reader.
Western blotting
H1993 cells were collected at 24 h post-transfection
and counted. Total protein of 1×105 cells was extracted
using RIPA solution (Sangon Biotech Co., Ltd.). A bicinchoninic
acid kit (Sangon Biotech Co., Ltd.) was used to measure protein
concentration. After denaturation in boiling water for 5 min,
proteins (30 µg) were separated by 10% SDS-PAGE. Subsequently,
proteins were transferred to PVDF membranes, followed by blocking
for 1 h in 5% non-fat milk at room temperature. The membranes were
first blotted with primary rabbit antibodies of PTEN (dilution,
1:1,500; cat. no. ab31392; Abcam) and GAPDH (dilution, 1:1,300;
cat. no. ab37168; Abcam) for 18 h at 4°C, followed by blotting with
horseradish peroxidase-cinjugated goat goat anti-rabbit
immunoglobulin G secondary antibody (dilution, 1:1,500; cat. no.
ab6721; Abcam) secondary antibodies at 24°C for 2 h. Pierce ECL
Western Blotting Substrate (Thermo Fisher Scientific, Inc.) was
used to develop signals. Signals were processed using ImageJ v1.48
software (National Institutes of Health).
Statistical analysis
All data are presented as the mean values of three
biological replicates. Evaluation of the significance of
differences between non-tumor and NSCLC tissues was performed using
the paired Student's t-test. Differences among cell transfection
groups were analyzed using the Kruskal-Wallis test and post hoc
Dunn's test. Associations were analyzed using linear regression.
The 66 patients with NSCLC were grouped into high-(n=33) and
low-ZNF281 (n=33) expression level groups according to its median
expression level in NSCLC (2.28). GraphPad Prism 6 software
(GraphPad Software, Inc.) was used for statistical analysis
included the plotting of survival curves, and the log-rank test was
used to compare survival curves. Differences in the clinical stage,
cancer grade, subtypes, age and sex between two groups were
analyzed using the χ2 test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Low ZNF281 expression in NSCLC
predicts poor overall survival rate
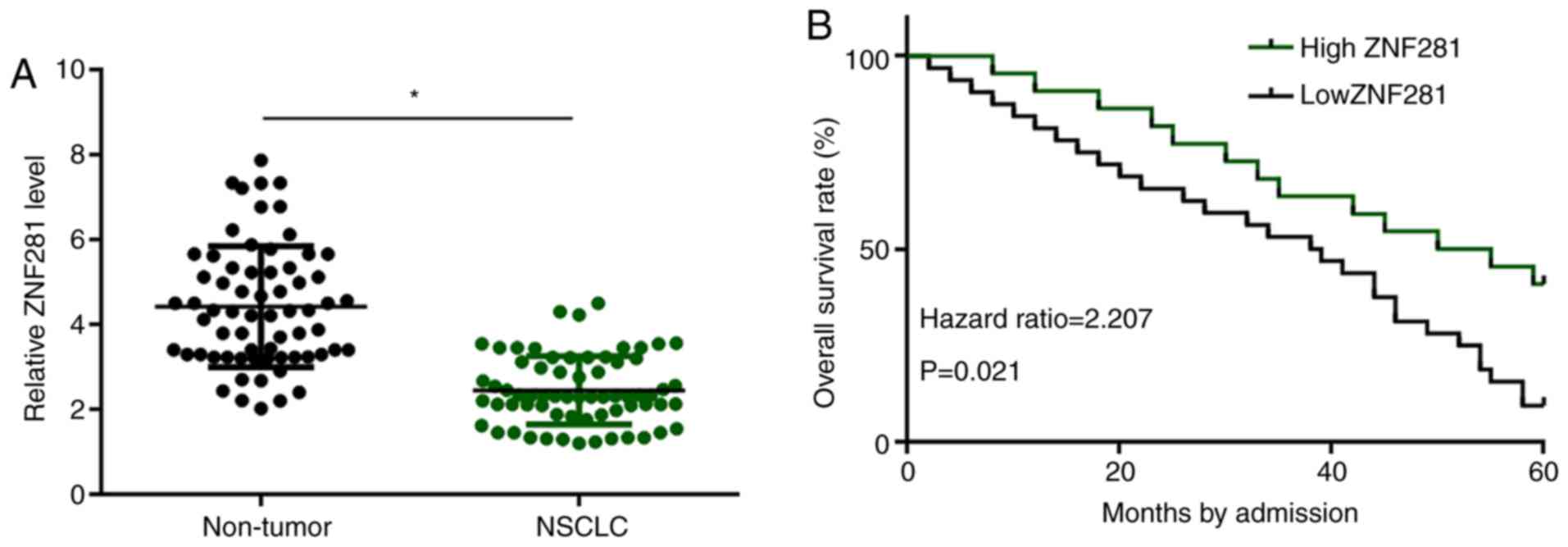
Levels of ZNF281 expression were measured using
RT-qPCR and compared between two types of tissues (non-tumor vs.
NSCLC) using the paired Student's t-test. Compared with non-tumor
samples, expression levels of ZNF281 in NSCLC tissue samples were
significantly lower (P<0.05; Fig.
1A). Patients were grouped into high- and low-expression groups
according to the median expression level, and survival curves were
plotted and compared. No significant differences in the clinical
stage, cancer grade, subtype, age and sex were observed between the
two groups (Table I). It was
observed that the 5-year overall survival rate of patients in the
high-ZNF281 expression group was significantly higher compared with
that of patients in the low-ZNF281 expression group (Fig. 1B).
 | Table I.Comparison of clinical data between
high- and low-ZNF281 expression level groups. |
Table I.
Comparison of clinical data between
high- and low-ZNF281 expression level groups.
| Variable | High-ZNF281
expression, n | Low-ZNF281
expression, n | P-value |
|---|
| Cases | 33 | 33 |
|
| Sex |
|
|
|
| Male | 23 | 21 | 0.60 |
|
Female | 10 | 12 |
|
| Age, years |
|
>50 | 17 | 19 | 0.62 |
| ≤50 | 16 | 14 |
|
| Subtype |
|
|
|
| Squamous
cell carcinoma | 14 | 16 | 0.62 |
|
Adenocarcinoma | 19 | 17 |
|
| Grade |
| I | 6 | 8 | 0.79 |
| II | 9 | 10 |
|
| III | 10 | 10 |
|
| IV | 8 | 5 |
|
| Stage |
| I | 2 | 4 | 0.81 |
| II | 9 | 10 |
|
| III | 11 | 9 |
|
| IV | 11 | 10 |
|
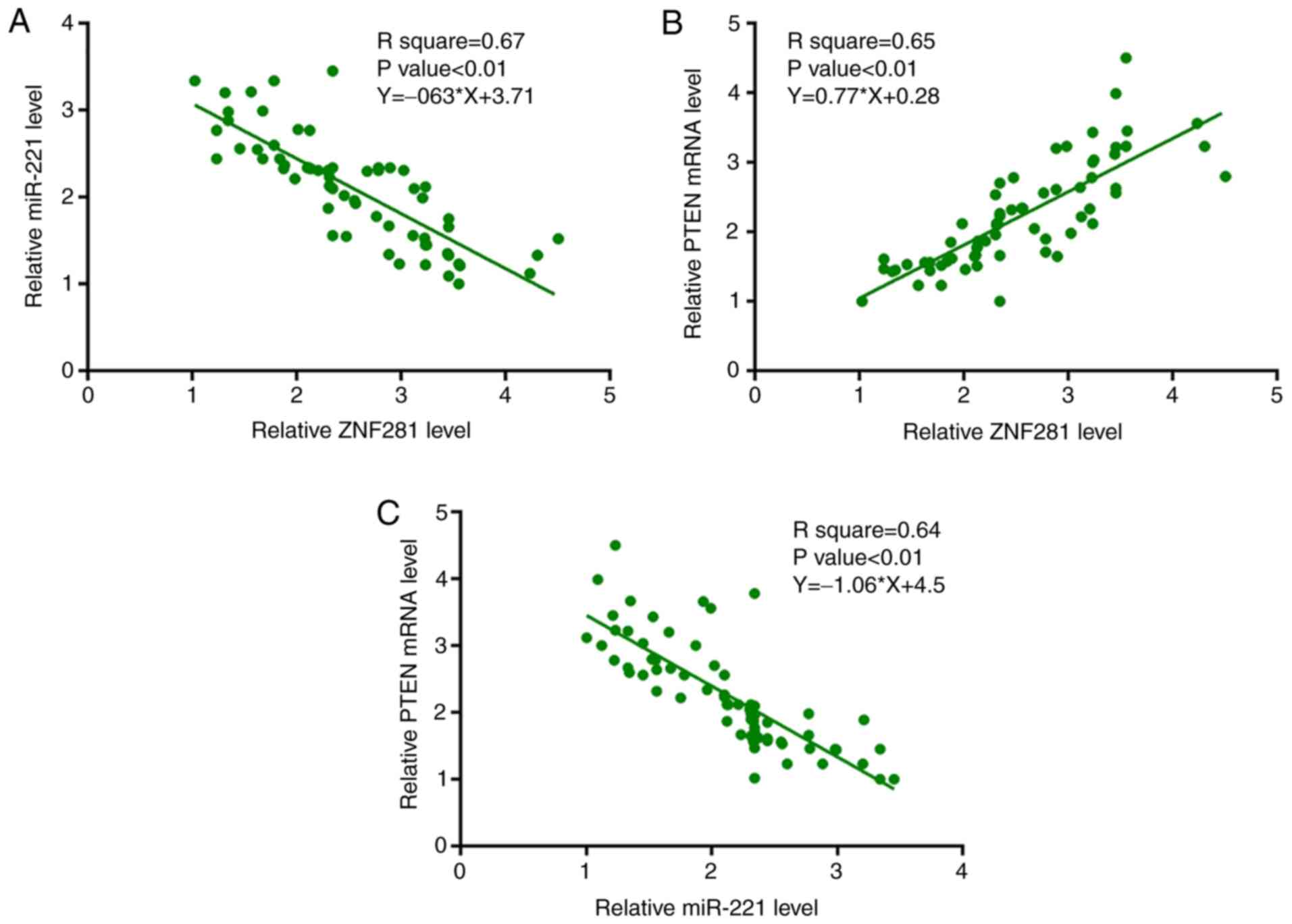
ZNF281 is significantly associated
with miR-221 and PTEN mRNA in NSCLC tissues
Furthermore, expression levels of miR-221 and PTEN
mRNA in NSCLC tissues were measured by performing RT-qPCR.
Associations between ZNF281 and miR-221/PTEN mRNA were analyzed
using linear regression analysis. It was observed that the
expression levels of ZNF281 were significantly negatively
associated with miR-221 expression (Fig.
2A); however, the expression levels of ZNF281 and PTEN mRNA
were significantly positively associated (Fig. 2B). Additionally, the association
between miR-221 and PTEN mRNA expression was analyzed by linear
regression analysis and it was observed that miR-221 and PTEN mRNA
were significantly negatively associated (Fig. 2C).
ZNF281 upregulates PTEN mRNA and
protein expression via the downregulation of miR-221 in H1993
cells
To analyze the interactions between ZNF281, PTEN and
miR-221, expression vectors of ZNF281 and PTEN as well as a miR-221
mimic were transfected into H1993 cells. RT-qPCR results revealed
that, compared with the NC and C groups, expression levels of
ZNF281, PTEN mRNA and miR-221 were significantly increased at 24 h
post-transfection (P<0.05; Fig.
3A). The effects of ZNF281 and miR-221 overexpression on mRNA
and protein levels were analyzed by RT-qPCR and western blotting,
respectively. Cells with ZNF281 overexpression exhibited
downregulated miR-221 expression, whereas cells with miR-221
overexpression exhibited unaffected ZNF281 expression (P<0.05;
Fig. 3B). Compared with the two
control groups, cells with ZNF281 overexpression exhibited
upregulated PTEN expression, whereas cells with miR-221
overexpression exhibited downregulated PTEN expression.
Additionally, miR-221 overexpression reduced the effects of ZNF281
overexpression on PTEN expression at both mRNA and protein levels
(P<0.05; Fig. 3C).
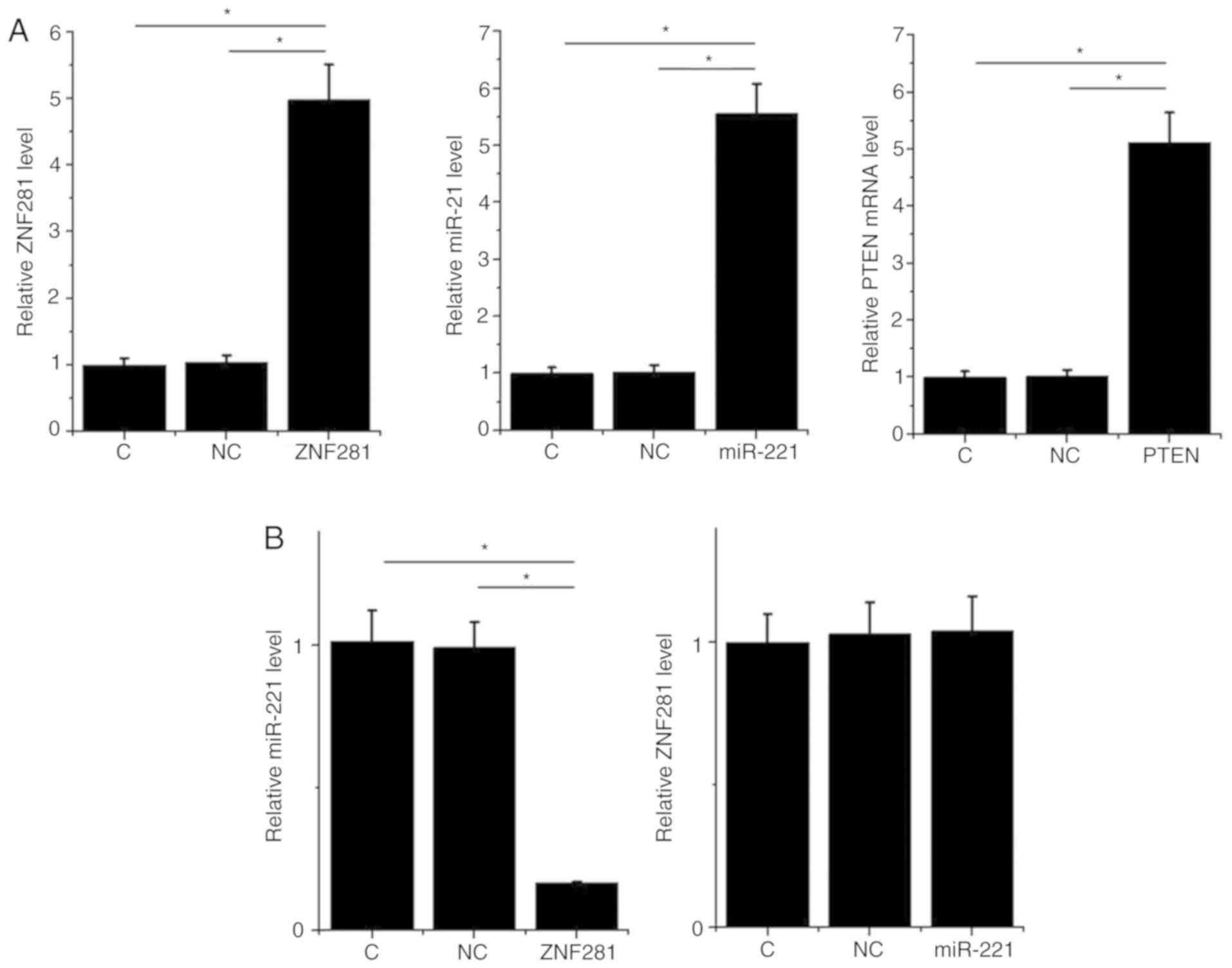 | Figure 3.ZNF281 upregulates PTEN via
downregulation of miR-221 in H1993 cells. In order to analyze the
interactions among ZNF281, PTEN and miR-221, expression vectors of
ZNF281 and PTEN, and a miR-221 mimic were transfected into H1993
cells. (A) Overexpression of ZNF281, PTEN mRNA and miR-221 was
confirmed by RT-qPCR. (B) Interaction between ZNF281 and miR-221
analyzed by RT-qPCR. (C) Effects of ZNF281 and miR-221
overexpression on PTEN expression were analyzed by western blotting
and RT-qPCR. Data are presented as the means ± standard deviation.
*P<0.05. C, control (untransfected cells); NC, negative control
(cells transfected with empty vector or negative control miRNA);
ZNF281, zinc finger protein 281; NSCLC, non-small cell lung cancer;
miR, microRNA; PTEN, phosphatase and tensin homolog; RT-qPCR,
reverse transcription-quantitative PCR. |
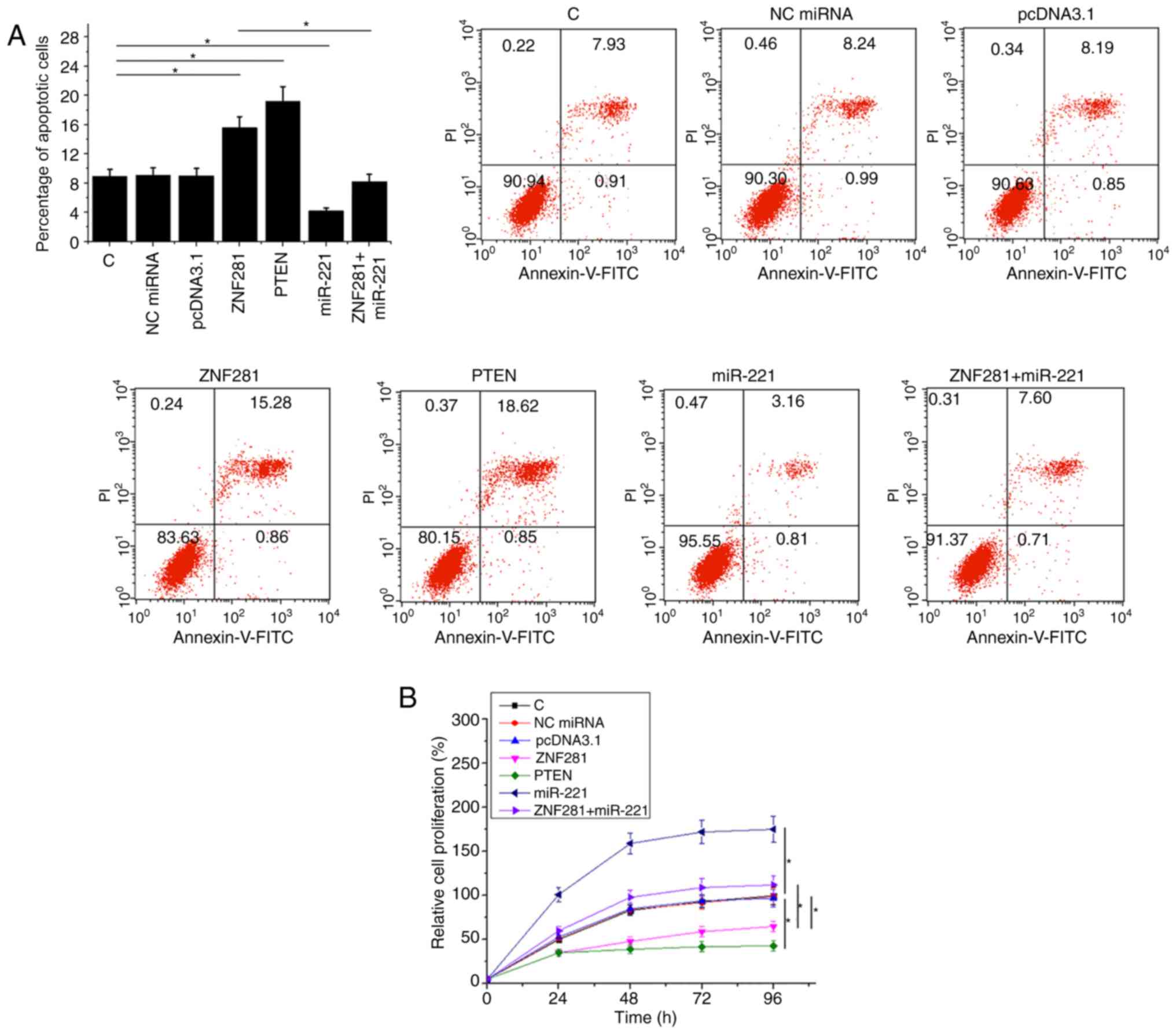
ZNF281 regulates H1993 cell apoptosis
and proliferation via miR-221 and PTEN
Compared with the NC and C groups, ZNF281 and PTEN
overexpression resulted in an increase in cell apoptosis (Fig. 4A) and inhibited cell proliferation
(Fig. 4B) of NSCLC cells
(P<0.05). Additionally, miR-221 overexpression partially
attenuated the functions of ZNF281 (P<0.05).
Discussion
The present study primarily investigated the role of
ZNF281 in NSCLC, a major subtype of lung cancer. In NSCLC, ZNF281
was downregulated and could downregulate oncogenic miR-221 to
upregulate PTEN, thereby promoting cancer cell apoptosis and
inhibiting cancer cell proliferation.
To the best of our knowledge, the expression pattern
and functionality of ZNF281 have only been investigated in glioma
(11). In glioma, ZNF281 is
downregulated and inhibits the stemness, proliferation and
invasiveness of glioma cells via inactivation of the NF-κB
signaling pathway (11), which is a
well-characterized oncogenic signaling pathway that serves critical
roles in numerous aspects of cancer biology (16,17). The
present study was the first to report the downregulation of ZNF281
in NSCLC. In addition, ZNF281 overexpression resulted in the
promotion of cell apoptosis and inhibition of cell proliferation.
Therefore, the present data indicated that ZNF281 serves a
tumor-suppressive role in NSCLC.
miR-221 is an oncogenic miRNA in numerous human
cancer types, including liver cancer (18). However, in a previous study, miR-221
has been reported to have inhibitory effects on the proliferation
of NSCLC cells (19), which was
consistent with the results observed in the present study; for
example, it was revealed that miR-221 overexpression resulted in
the promotion of cancer cell proliferation and inhibited cancer
cell apoptosis. This may be attributable to the different cell line
used in the present study. In another study, miR-221 has been
reported to directly target PTEN to inhibit gastric carcinoma cell
proliferation (10). In the present
study, PTEN was downregulated in NSCLC cells following miR-221
overexpression. Therefore, miR-221 may also target PTEN in
NSCLC.
The present study demonstrated that ZNF281
downregulated miR-221 which resulted in the upregulation of PTEN.
Additionally, it is known that both ZNF281 and miR-221 have direct
interactions with NF-κB (11,12);
therefore, NF-κB may mediate the interaction between ZNF281 and
miR-221. However, further studies are required to investigate the
mechanisms that mediate the interaction between ZNF281 and miR-221.
Future study will include a higher sample number and in vivo
experiments.
In conclusion, ZNF281 was downregulated in NSCLC and
may downregulate miR-221, resulting in the upregulation of PTEN,
thereby promoting cancer cell apoptosis and proliferation.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XL and WJ designed the study. XL, BY and XW
performed the experiments. FW, YL, NW and XY collected and analyzed
the data. WJ drafted the manuscript. All authors read and approved
the final version of the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Review Board
of Hiser Medical Center and the Qingdao Ethics Committee (approval
no. 2011HC08LQ03). All patients provided written informed
consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Miller KD, Siegel RL, Lin CC, Mariotto AB,
Kramer JL, Rowland JH, Stein KD, Alteri R and Jemal A: Cancer
treatment and survivorship statistics, 2016. CA Cancer J Clin.
66:271–289. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Herbst RS, Morgensztern D and Boshoff C:
The biology and management of non-small cell lung cancer. Nature.
553:446–454. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yamamoto H, Yatabe Y and Toyooka S:
Inherited lung cancer syndromes targeting never smokers. Transl
Lung Cancer Res. 7:498–504. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Subramanian J and Govindan R: Molecular
genetics of lung cancer in people who have never smoked. Lancet
Oncol. 9:676–682. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brennan P, Hainaut P and Boffetta P:
Genetics of lung-cancer susceptibility. Lancet Oncol. 12:399–408.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sun H, Lesche R, Li DM, Liliental J, Zhang
H, Gao J, Gavrilova N, Mueller B, Liu X and Wu H: PTEN modulates
cell cycle progression and cell survival by regulating
phosphatidylinositol 3, 4, 5,-trisphosphate and Akt/protein kinase
B signaling pathway. Proc Natl Acad Sci USA. 96:6199–6204. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Carnero A, Blanco-Aparicio C, Renner O,
Link W and Leal JF: The PTEN/PI3K/AKT signalling pathway in cancer,
therapeutic implications. Curr Cancer Drug Targets. 8:187–198.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chun-Zhi Z, Lei H, An-Ling Z, Yan-Chao F,
Xiao Y, Guang-Xiu W, Zhi-Fan J, Pei-Yu P, Qing-Yu Z and Chun-Sheng
K: MicroRNA-221 and microRNA-222 regulate gastric carcinoma cell
proliferation and radioresistance by targeting PTEN. BMC Cancer.
10:3672010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li XT, Li JC, Feng M, Zhou YX and Du ZW:
Novel lncRNA-ZNF281 regulates cell growth, stemness and invasion of
glioma stem-like U251s cells. Neoplasma. 66:118–127. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhao D, Zhuang N, Ding Y, Kang Y and Shi
L: miR-221 activates the NF-κB pathway by targeting A20. Biochem
Biophys Res Commun. 472:11–18. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Edge SB and Compton CC: The American joint
committee on cancer: The 7th edition of the AJCC cancer staging
manual and the future of TNM. Ann Surg Oncol. 17:1471–1474. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sun Z, Aubry MC, Deschamps C, Marks RS,
Okuno SH, Williams BA, Sugimura H, Pankratz VS and Yang P:
Histologic grade is an independent prognostic factor for survival
in non-small cell lung cancer: An analysis of 5018 hospital-and 712
population-based cases. J Thorac Cardiovasc Surg. 131:1014–1020.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rinkenbaugh AL and Baldwin AS: The NF-κB
pathway and cancer stem cells. Cells. 5:162016. View Article : Google Scholar
|
|
17
|
Li F, Zhang J, Arfuso F, Chinnathambi A,
Zayed ME, Alharbi SA, Kumar AP, Ahn KS and Sethi G: NF-κB in cancer
therapy. Arch Toxicol. 89:711–731. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pineau P, Volinia S, McJunkin K, Marchio
A, Battiston C, Terris B, Mazzaferro V, Lowe SW, Croce CM and
Dejean A: miR-221 overexpression contributes to liver
tumorigenesis. Proc Natl Acad Sci USA. 107:264–269. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yamashita R, Sato M, Kakumu T, Hase T,
Yogo N, Maruyama E, Sekido Y, Kondo M and Hasegawa Y: Growth
inhibitory effects of miR-221 and miR-222 in non-small cell lung
cancer cells. Cancer Med. 4:551–564. 2015. View Article : Google Scholar : PubMed/NCBI
|