Introduction
Osteosarcoma mainly arises from human mesenchymal
cells and is one of the most common primary malignant bone tumors
(1–3). The incidence of osteosarcoma in
adolescents is relatively high (4,5). The
main treatment option for low-grade osteosarcoma is surgery alone,
and the prognosis for patients with low-grade osteosarcoma is
relatively good. High-grade osteosarcoma or osteosarcoma with
metastasis can be treated with surgery and chemotherapy; however,
the mean 5-year survival rate is <25% (1,6,7). Although the treatment for osteosarcoma
has improved, few effective methods are available (8,9).
Research has focused on the mechanisms involved in the initiation
and development of osteosarcoma, yet the intrinsic mechanisms
involved in the malignant behaviors of osteosarcoma remain unclear.
Further studies are needed to improve the understanding of the
molecular mechanisms involved in osteosarcoma initiation, growth,
migration and invasion, which could help the development of novel
treatments for osteosarcoma.
Solute carrier 25 family member 10 (SLC25A10), also
known as dicarboxylate carrier, is an important regulator of human
energy metabolism and redox homeostasis (10,11).
SLC25A10 contains six transmembrane regions and three cognate
repeats, and is localized in the cytoplasm and the inner
mitochondrial membrane (12,13). SLC25A10 transports dicarboxylate
substrates and supplies substrates for several pathways, including
sulfur metabolism and gluconeogenesis (10,14).
SLC25A10 is involved in fatty acid synthesis, glucose-stimulated
insulin secretion and additional physiological processes (12,15,16).
Moreover, Zhou et al (10)
suggested that knockdown of SLC25A10 in human cancer cells markedly
decreased cell growth and increased sensitivity to anticancer
drugs, demonstrating its role as an oncogene. However, the role of
SLC25A10 in different types of human cancer, including
osteosarcoma, remains unclear. Therefore, further studies focusing
on osteosarcoma are required.
The present study demonstrated that the expression
levels of SLC25A10 were higher in human osteosarcoma tissues,
compared with normal bone tissues. In addition, in patients with
osteosarcoma, the expression levels of SLC25A10 were positively
associated with tumor metastasis, clinical Enneking stage, poor
relapse-free survival (RFS) and overall survival (OS) rates.
Knockdown of SLC25A10 with short hairpin RNA (shRNA) significantly
decreased cell proliferation, increased cell apoptosis and
suppressed cell mitosis in osteosarcoma cells. Moreover, cyclin E1
(CCNE1) was positively regulated by SLC25A10, while P21/P27 were
negatively regulated by SLC25A10. CCNE1 was previously described as
an important tumor promoter in many types of human cancer, and
P21/P27 were found to be tumor suppressors in many human cancer
types (17–21). Collectively, CCNE1, P21 and P27 may
mediate the oncogenic role of SLC25A10 in human osteosarcoma
cells.
Materials and methods
Clinical osteosarcoma and normal bone
samples
In total, 60 osteosarcoma tissues and 60 normal bone
tissues were collected in The Department of Orthopedics and The
Department of Pathology in The First Affiliated Hospital of Anhui
Medical University. These tissues were collected from patients with
osteosarcoma or bone diseases who underwent resection in The First
Affiliated Hospital of Anhui Medical University between January
2011 and December 2013. These osteosarcoma tissues and normal bone
tissues were not from the same patients. The clinicopathological
features of the enrolled patients with osteosarcoma were collected
from The Department of Pathology, The First Affiliated Hospital of
Anhui Medical University. The 60 patients with osteosarcoma were
followed-up for >5 years, and the RFS and OS rates were
determined. Ethical approval from The Institutional Review Boards
of Anhui Medical University was obtained prior to the study. All
experiments involving human patients were performed according to
The Code of Ethics of The World Medical Association (Declaration of
Helsinki). Informed consent was obtained from all patients involved
in the present study.
Immunohistochemistry
The protein levels of SLC25A10 in 4-µm thick
paraffin sections of osteosarcoma tissues and normal bone tissues
(10% formalin fixed at room temperature for 24 h) were detected by
immunohistochemistry, as previously described (22,23).
Sections were deparaffinized in xylene, rehydrated in a series of
ethanol solutions (100, 100, 95, 85 and 75%) and heated in 0.01 M
sodium citrate buffer at 100°C for 10 min for antigen retrieval.
Sections were incubated with 3% hydrogen peroxide incubation at
room temperature for 10 min, and then incubated with primary
antibody [SLC25A10 rabbit polyclonal antibody (1:200; 12086-1-AP;
ProteinTech Group, Inc.)] for 3 h at room temperature, followed by
incubation for 15 min at room temperature with horseradish
peroxidase (HRP)-conjugated secondary antibody (1:1; MaxVision-HRP,
KIT-5030; Fuzhou Maixin Biotech Co., Ltd.) 3,3′-diaminobenzidine
tetrahydrochloride (Fuzhou Maixin Biotech Co., Ltd.) was used for
visualization. Sections with <10% positive stained cells were
considered as SLC25A10-negative, and sections with ≥10% positive
stained cells were considered to be SLC25A10-positive using a light
microscope (Olympus Corporation) at ×20 magnification.
Cells and cell culture
The human MG-63 and U2OS osteosarcoma cell lines
(both from American Type Culture Collection) were used in the
present study. MG-63 and U2OS cells were cultured using DMEM medium
(Gibco; Thermo Fisher Scientific, Inc.) containing 10% fetal bovine
serum (Gibco; Thermo Fisher Scientific, Inc.) in humidified
conditions at 37°C and 5% CO2.
shRNA transfection
shRNAs targeting SLC25A10 (shSLC25A10-1 and
shSLC25A10-2) and control shRNA (shCtrl) were used for cell
functional experiments. All of the shRNAs were obtained from
Shanghai GenePharma Co., Ltd. Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) was used for shRNA
transfection (75 pmol/transfection), which was performed as
previously described (22,23). The shRNA sequences were as follows:
shSLC25A10-1, 5′-GTTTAGCTGGAGGCTTCGTGG-3′; shSLC25A10-2,
5′-CAAGCAGCTGGTCCTTAGCAC-3′; and shCtrl,
5′-TCAAGCTGCTAGGCCTATCCG-3′.
Western blotting
Western blotting experiments were performed to
detect the protein expression levels of SLC25A10, CCNE1, P21 and
P27 in human osteosarcoma cells. Western blot analysis was carried
out as previously described (22,23) and
β-actin was used as a control. Polyvinylidene difluoridemembranes
(EMD Millipore) were used for electrotransfer. Membranes were
blocked using incubation with 5% (w/v) non-fat milk powder at room
temperature for 2 h. Membranes were incubated with respective
antibodies (rabbit polyclonal antibodies anti-SLC25A10 (1:1,000;
12086-1-AP; ProteinTech Group, Inc.), anti-CCNE1 (1:1,000;
11554-1-AP; ProteinTech Group, Inc.), anti-P21 (1:1,000;
10355-1-AP; ProteinTech Group, Inc.), anti-P27 (1:1,000;
25614-1-AP; ProteinTech Group, Inc.), rabbit polyclonal antibody
anti-caspase-3 (1:500; #9662; Cell Signaling Technology, Inc.),
rabbit polyclonal anti-poly ADP-ribose polymerase (PARP; 1:500;
#9542; Cell Signaling Technology, Inc.) and rabbit polyclonal
antibody anti-β-actin (1:1,000; 20536-1-AP; ProteinTech Group,
Inc.) overnight at 4°C. Then, then membranes were incubated with
goat anti-rabbit IgG (H+L) HRP-conjugated secondary antibody
(1:50,000; 31460; Invitrogen; Thermo Fisher Scientific, Inc.) for 2
h at room temperature. Femto (34095; Thermo Fisher Scientific,
Inc.) and Pico (34077; Thermo Fisher Scientific, Inc.) were used
for visualization. Protein bands were identified using Image Quant
LAS 4000 (Cytiva) and analyzed using ImageJ version 1.8.0 software
(National Institutes of Health).
Cell proliferation assay
Cell counting, MTT and cell colony formation assays
were performed to evaluate the role of SLC25A10 in the
proliferation of human osteosarcoma cells, as previously described
(22,23). MG-63 and U2OS cells were transfected
with shRNAs, and collected after 24 h. In the cell counting assay,
10,000 cells/well were seeded into six-well plates, and the total
cell number was calculated every day during a period of 5 days. In
the MTT assay, 2,000 cells/well were seeded into 96-well plates,
and the MTT detection was performed for 5 days (the purple formazan
was dissolved in DMSO, and the absorbance was measured at 570 nm).
In the cell colony formation assay, 1,000 cells/well were seeded
into six-well plates, and cell colony formation was examined after
10 days.
Flow cytometry
After transfection, flow cytometry was carried out
to examine apoptosis and mitosis in MG-63 and U2OS cells. After
transfection with shRNA for 72 h, MG-63 or U2OS cells were
collected, fixed in 70% ethanol solution at 4°C for 30 min and
incubated with Annexin V-FITC (Beyotime Institute of Biotechnology)
and propidium iodide (Sigma-Aldrich; Merck KGaA) for 30 min at room
temperature. Flow cytometric analysis was performed to examine cell
apoptosis. Cells were collected, fixed in 70% ethanol solution at
4°C for 30 min and incubated with propidium iodide and RNAse A for
30 min at room temperature. Subsequently, a flow cytometer (LSR II;
BD Biosciences) was used to examine cell mitosis. Data were
analyzed using FlowJo version 7.6 software (Tree Star, Inc.).
Statistical analysis
The data are presented as the mean, calculated from
three independent experiments. SPSS 25.0 (IBM Corp) was used for
statistical analysis. For cell functional experiments, the
respective results were standardized by percentage. For cell
functional experiments and flow cytometry, one-way ANOVA followed
by Bonferroni or Tamhane post hoc tests were used.
Immunohistochemical results were analyzed using Pearson's
χ2 test. For RFS and OS analysis, Kaplan-Meier curves
were calculated and log-rank test was used to analyze the
statistical significance. P<0.05 was considered to indicate a
statistically significant difference.
Results
Clinical analysis of SLC25A10
expression in patients with osteosarcoma
In total, 60 paraffin-embedded osteosarcoma tissue
samples and 60 paraffin-embedded normal bone tissue samples were
collected. The protein levels of SLC25A10 in these tissues were
detected by immunohistochemistry, and a positive signal of SLC25A10
was mainly detected in the cytoplasm. Among these 60 osteosarcoma
tissues, 18 (30.0%) were SLC25A10-negative and 42 (70.0%) were
SLC25A10-positive (P<0.05; Table
I). Among the 60 normal bone tissues, 32 (53.3%) were
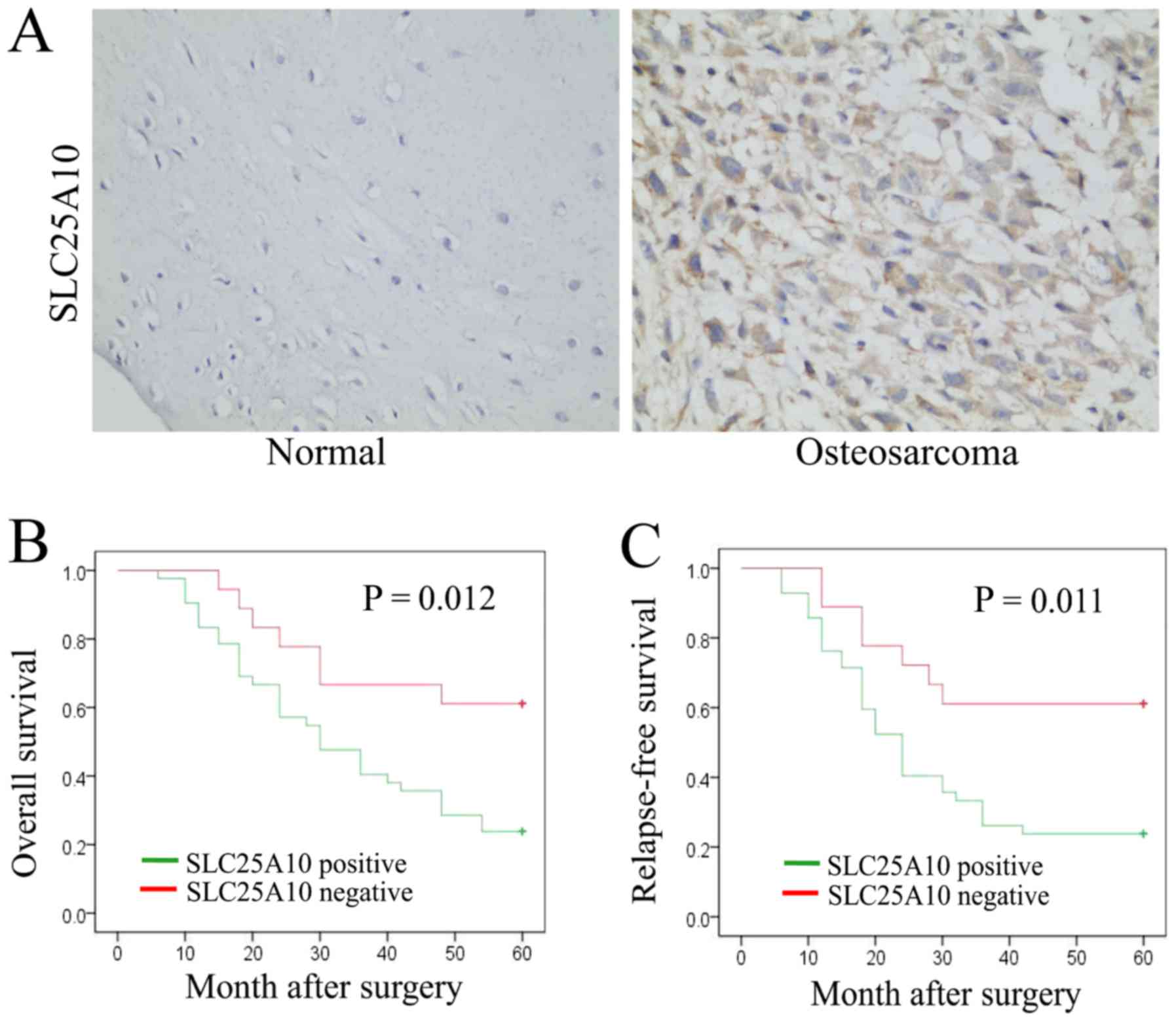
SLC25A10-negative and 28 (46.7%) were SLC25A10-positive (Table I). The representative SLC25A10
expression levels in osteosarcoma and normal bone tissues are
presented in Fig. 1A. Therefore, the
protein levels of SLC25A10 were higher in human osteosarcoma
tissues, compared with normal bone tissues.
 | Table I.Expression of SLC25A10 in
osteosarcoma and normal tissues. |
Table I.
Expression of SLC25A10 in
osteosarcoma and normal tissues.
|
|
| SLC25A10
expression |
|---|
|
|
|
|
|---|
| Group | n | Negative, n
(%) | Positive, n
(%) |
|---|
| Osteosarcoma | 60 | 18 (30.0) | 42
(70.0)a |
| Normal | 60 | 32 (53.3) | 28 (46.7) |
The association between SLC25A10 expression and
various clinicopathological features and survival rates in the 60
patients with osteosarcoma was analyzed. Positive expression of
SLC25A10 was found to be associated with a higher risk of tumor
metastasis (P=0.037) and worse clinical Enneking stage (P=0.042) in
the 60 patients with osteosarcoma (Table II). However, there was no
significant statistical significance in SLC25A10 expression between
patient groups of different age, sex or tumor size (all P>0.05;
Table II).
| Table II.Association of SLC25A10 expression
with clinicopathological parameters from patients with
osteosarcoma. |
Table II.
Association of SLC25A10 expression
with clinicopathological parameters from patients with
osteosarcoma.
|
|
| SLC25A10
expression, n (%) |
|
|
|---|
|
|
|
|
|
|
|---|
| Parameter | n | Negative | Positive | P-value | χ2 |
|---|
| Age, years |
|
|
|
|
|
|
≤20 | 27 | 7 (25.9) | 20 (74.1) | 0.533 | 0.388 |
|
>20 | 33 | 11 (33.3) | 22 (66.7) |
|
|
| Sex |
|
|
|
|
|
|
Male | 37 | 11 (29.7) | 26 (70.3) | 0.954 | 0.003 |
|
Female | 23 | 7 (30.4) | 16 (69.6) |
|
|
| Tumor size, cm |
|
|
|
|
|
| ≤5 | 36 | 9 (25.0) | 27 (75.0) | 0.301 | 1.071 |
|
>5 | 24 | 9 (37.5) | 15 (62.5) |
|
|
| Metastasis |
|
|
|
|
|
| No | 42 | 16 (38.1) | 26 (61.9) | 0.037 | 4.369 |
|
Yes | 18 | 2 (11.1) | 16 (88.9) |
|
|
| Enneking stage |
|
|
|
|
|
|
I–II | 28 | 12 (42.9) | 16 (57.1) | 0.042 | 4.133 |
|
III | 32 | 6 (18.8) | 26 (71.2) |
|
|
Moreover, these 60 patients with osteosarcoma were
all followed-up for >5 years, and potential associations between
SLC25A10 expression and RFS and OS rates were examined. Patients
with osteosarcoma who displayed positive expression of SLC25A10
showed decreased OS and RFS rates, compared with patients with
negative SLC25A10 expression (both P<0.05; Fig. 1B and C). Therefore, high expression
of SLC25A10 was associated with poor prognosis in patients with
osteosarcoma.
Role of SLC25A10 in osteosarcoma cell
proliferation
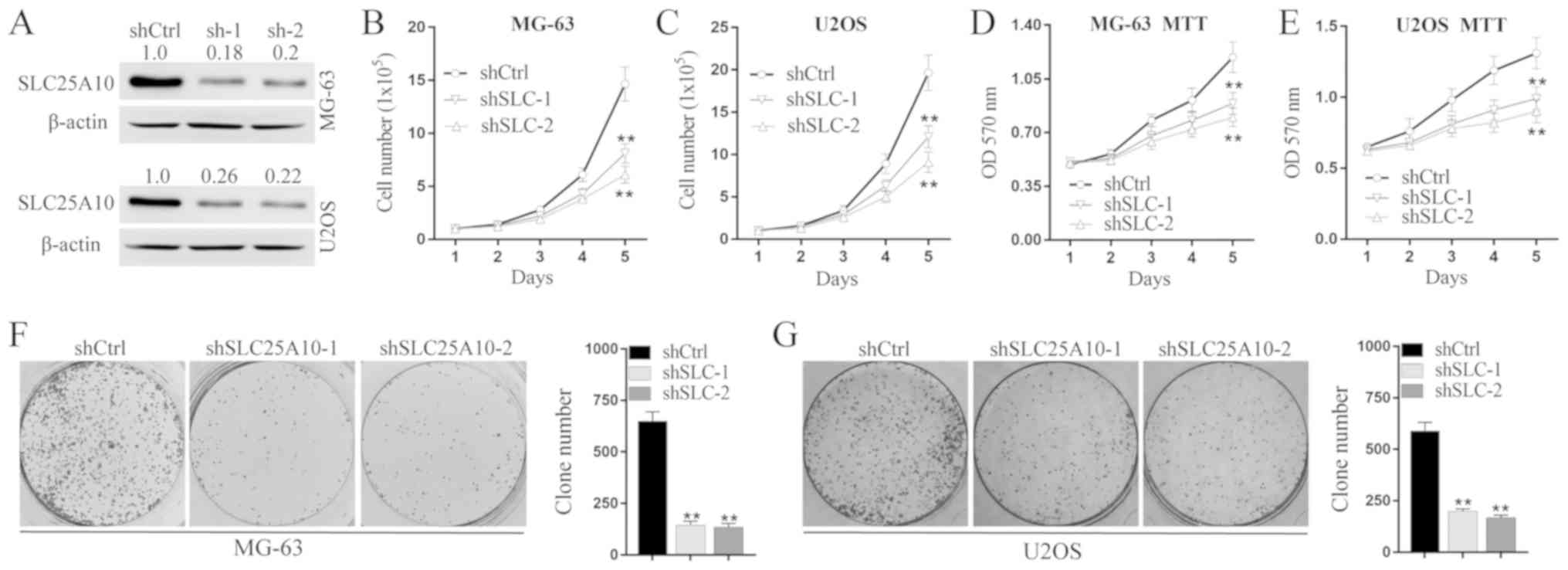
The human osteosarcoma cell lines MG-63 and U2OS
were selected for cell functional experiments. Protein levels of
SLC25A10 decreased after transfection with shSLC25A10-1 or
shSLC25A10-2 compared with shCtrl in both MG-63 and U2OS cells
(Fig. 2A). As determined by the cell
counting assay (Fig. 2B and C), both
shSLC25A10-1 and shSLC25A10-2 significantly reduced the total cell
number, compared with shCtrl in both MG-63 and U2OS cells over 5
days (standardized by percentage: MG-63 shCtrl, 100%; MG-63
shSLC25A10-1, 55.25%; MG-63 shSLC25A10-2, 41.61%; P<0.01; U2OS
shCtrl, 100%; U2OS shSLC25A10-1, 61.55%; U2OS shSLC25A10-2, 46.29%;
P<0.01). Moreover, as detected by the MTT assay (Fig. 2D and E), cell proliferation decreased
significantly during a period of 5 days in both MG-63 and U2OS
cells after transfection with shSLC25A10-1 or shSLC25A10-2,
compared with shCtrl (MTT assay at day 5, standardized by
percentage: MG-63 shCtrl, 100%; MG-63 shSLC25A10-1, 74.79%; MG-63
shSLC25A10-2, 67.23%; P<0.01; U2OS shCtrl, 100%; U2OS
shSLC25A10-1, 75.57%; U2OS shSLC25A10-2, 68.70%; P<0.01). In
addition, MG-63 shSLC25A10-1 and MG-63 shSLC25A10-2 cells showed
decreased cell colony formation compared with MG-63 shCtrl cells
(standardized by percentage: MG-63 shCtrl 100%, MG-63 shSLC25A10-1
22.31%, MG-63 shSLC25A10-2 20.77%; P<0.01). Moreover, U2OS
shSLC25A10-1 and U2OS shSLC25A10-2 cells also showed decreased
colony formation compared with U2OS shCtrl cells (Standardized by
percentage: U2OS shCtrl 100%, U2OS shSLC25A10-1 33.56%, U2OS
shSLC25A10-2 28.64%; P<0.01; Fig. 2F
and G). Collectively, SLC25A10 knockdown suppressed cell
proliferation in human osteosarcoma cells.
Role of SLC25A10 in osteosarcoma cell
apoptosis and mitosis
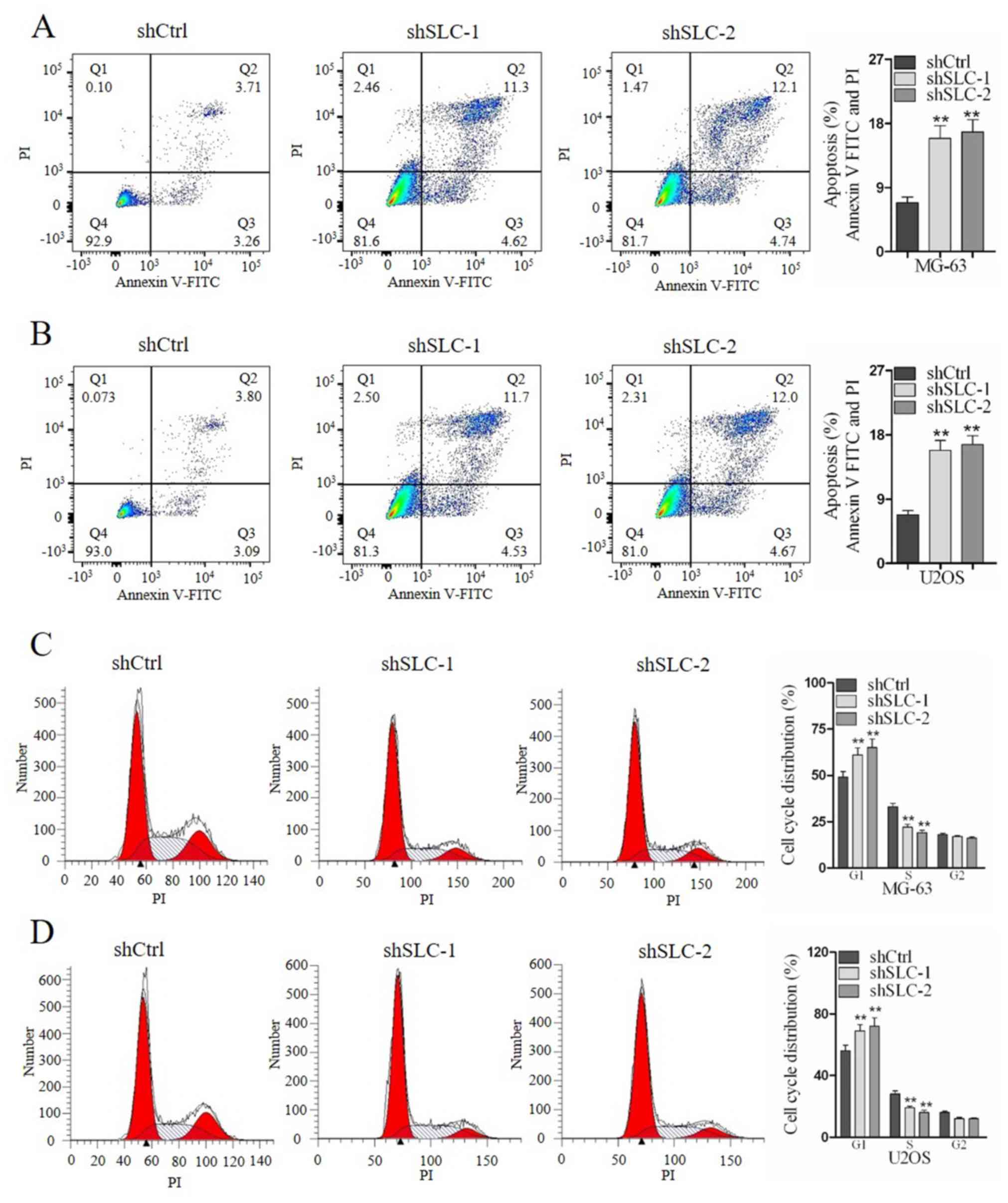
The role of SLC25A10 in apoptosis and mitosis of
osteosarcoma cells was evaluated by flow cytometry. SLC25A10 shRNA
knockdown significantly increased the percentage of apoptotic cells
in both MG-63 and U2OS cells (MG-63 shCtrl, 6.97%; MG-63
shSLC25A10-1, 15.82%; MG-63 shSLC25A10-2, 16.84%; P<0.01; U2OS
shCtrl, 6.89%; U2OS shSLC25A10-1, 16.23%; U2OS shSLC25A10-2,
16.67%; P<0.01; Fig. 3A and B).
Moreover, transfection of shSLC25A10-1 or shSLC25A10-2
significantly increased the percentage of cells in G1
phase and decreased the percentage of cells in S phase in both
MG-63 and U2OS cells (Fig. 3C and
D). Therefore, the present data suggested that SLC25A10
promoted cell mitosis and suppressed cell apoptosis in human
osteosarcoma cells.
Regulation analysis of SLC25A10 in
osteosarcoma cells
To investigate the downstream mechanisms of
SLC25A10, several candidate genes, including MYC, tumor protein 53
(TP53), cyclin D1 (CCND1), CCNE1, PTEN, STAT3, epidermal growth
factor (EGFR), P21 and P27 were examined in MG-63 cells after
transfection with shSLC25A10-1, shSLC25A10-2 or shCtrl. These genes
were selected according to previous studies which demonstrated
their oncogenic role (24–31). Thus, it was hypothesized that these
gene candidates were regulated by SLC25A10, and that they might
mediate the role of SLC25A10 in osteosarcoma cells.
As determined by the western blotting assay, the
protein level of CCNE1 decreased markedly after transfection with
shSLC25A10-1 or shSLC25A10-2, whereas the protein levels of P21 and
P27 increased after transfection with shSLC25A10-1 or shSLC25A10-2
in both MG-63 and U2OS cells, compared with shCtrl (Fig. 4A). However, there were no marked
changes in MYC, TP53, CCND1, PTEN, STAT3 and EGFR protein levels
after shSLC25A10 transfection compared with shCtrl (data not
shown). Therefore, only CCNE1 was positively regulated by SLC25A10.
Moreover, P21 and P27 were negatively regulated by SLC25A10 in
human osteosarcoma cells.
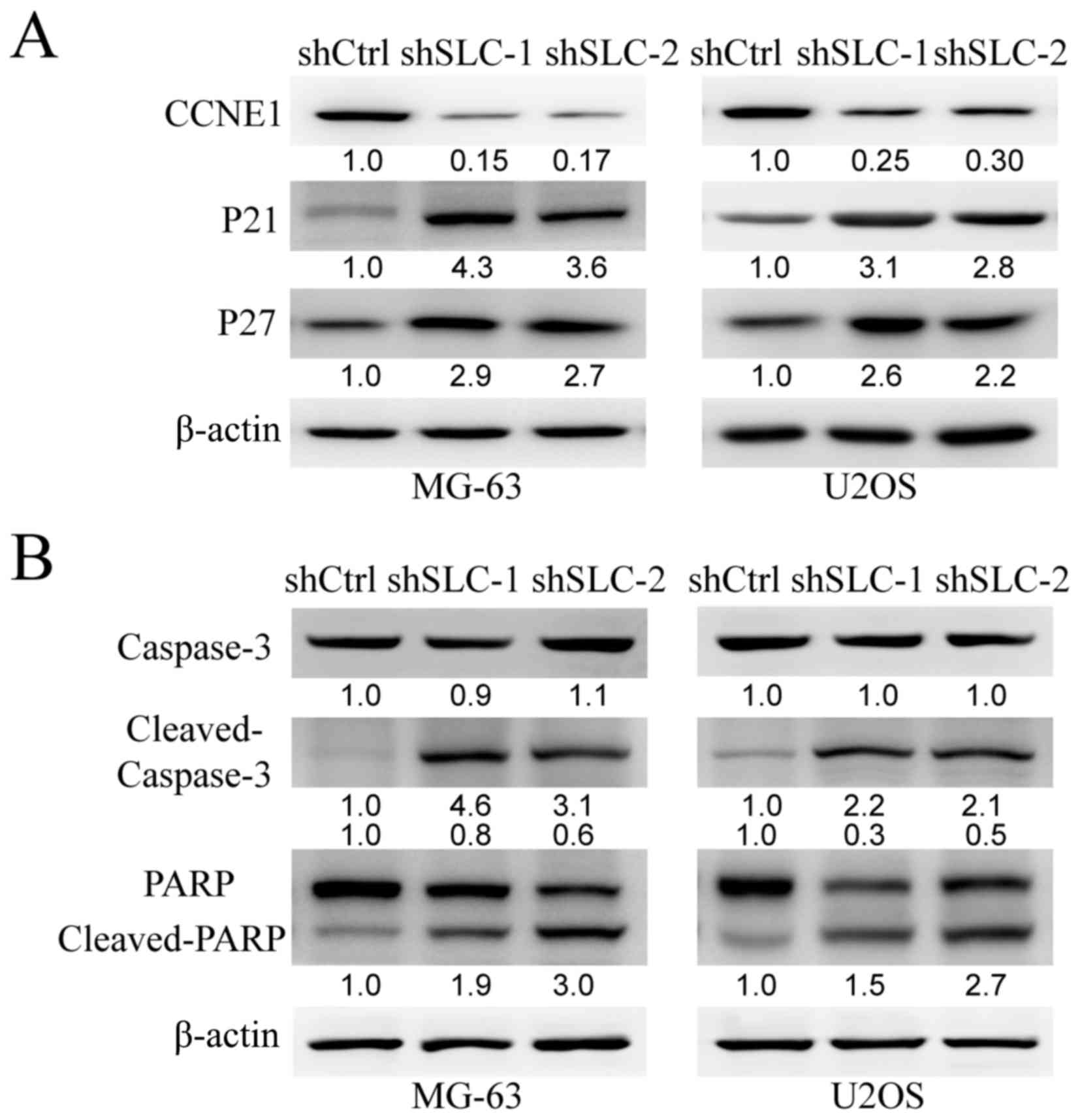 | Figure 4.CCNE1, P21 and P27 are regulated by
SLC25A10 in human osteosarcoma cells. (A) Protein levels of CCNE1,
P21 and P27 in MG-63 and U2OS cells after transfection with
shSLC25A10-1, shSLC25A10-2 or shCtrl were examined by western blot
analysis. β-actin was used as loading control. The numbers below
were fold change compared with reference protein. (B) Protein
levels of caspase-3, cleaved caspase-3, PARP and cleaved PARP in
MG-63 and U2OS cells following transfection with shSLC25A10-1,
shSLC25A10-2 or shCtrl were examined by western blotting. The
numbers below were fold change compared with reference protein
respectively. sh, short hairpin; SLC25A10/SLC, solute carrier 25
family member 10; CCNE1, cyclin E1; PARP, poly ADP-ribose
polymerase; Ctrl, control. |
In addition, cleaved caspase-3 and its substrate
cleaved PARP markedly increased after transfection with
shSLC25A10-1 or shSLC25A10-2 in both MG-63 and U2OS cells.
Therefore, knockdown of SLC25A10 promoted caspase-3-mediated
apoptosis in human osteosarcoma cells.
Discussion
In the present study, several systematical
experiments were performed to examine the role of SLC25A10 in human
osteosarcoma. As examined by immunohistochemistry, the protein
levels of SLC25A10 were much higher in osteosarcoma tissues
compared with normal bone tissues. A high expression level of
SLC25A10 was associated with worse clinicopathological parameters,
including tumor metastasis and clinical Enneking stage in patients
with osteosarcoma. Patients with osteosarcoma with high expression
levels of SLC25A10 showed lower RFS and OS rates, compared with
patients with low expression levels of SLC25A10. In MG-63 and U2OS
human osteosarcoma cells, shRNA-mediated SLC25A10 knockdown
significantly suppressed cell proliferation as determined by cell
counting, MTT and colony formation assays. In addition, knockdown
of SLC25A10 promoted apoptosis and suppressed mitosis in human
osteosarcoma cells, as determined by flow cytometry. In addition,
knockdown of SLC25A10 promoted caspase-3-mediated apoptosis. Zhou
et al (10) demonstrated that
SLC25A10-knockdown in A549 cells decreased cell proliferation,
leading to a less malignant phenotype, as well as increased
glutamine dependency and sensitivity to oxidative stress. The data
from the present study are in line with these previous findings.
However, few previous studies have documented the role of SLC25A10
in other human cancer types. Therefore, to the best of the authors'
knowledge, the present study is the first to show that SLC25A10 may
play a tumor-promoting role in human osteosarcoma.
In addition, the downstream pathway underlying the
oncogenic role of SLC25A10 was examined, and CCNE1 was found to be
positively regulated by SLC25A10. CCNE1 is a member of the cyclin
family, which contributes to the activity of the cell cycle. As
reported in previous study, CCNE1 is upregulated, and plays an
oncogenic role in human osteosarcoma (17). MicroRNA (miR)-874 and miR-132
directly targeted CCNE1, and both acted as tumor suppressors,
inhibiting the malignant features of human osteosarcoma cells. As a
tumor promoter, CCNE1 mediated the tumor-suppressing effect of
miR-874 and miR-132 (26,32). Moreover, CCNE1 was found to act as an
oncogene in various other types of human cancer, including breast
cancer (18,33), lung cancer (34), gastric cancer (35) and hepatocellular carcinoma (36). Therefore, the tumor-promoting role of
SLC25A10 in human osteosarcoma might be partly mediated by
CCNE1.
Furthermore, in the present study, P21 and P27 were
found to be negatively regulated by SLC25A10 in osteosarcoma cells.
P21 is an important cyclin-dependent kinase inhibitor, which
suppresses both cell proliferation and metastasis in human
osteosarcoma cells. Moreover, overexpression of P21 increases
sensitivity of osteosarcoma cells to anti-cancer drugs (37–39).
Furthermore, P21 acts as tumor suppressor in human nasopharyngeal
carcinoma (19), cervical cancer
(20), breast cancer (40), non-small cell lung cancer (41). P27 is a tumor suppressor gene in many
types of human cancer. In osteosarcoma, Liao et al (42) identified that P27 is negatively
correlated with S-phase kinase associated protein 2 and serves a
tumor-suppressing role. Hu et al (43) demonstrated that inhibition of P27
mediated the tumor-promoting role of miR-227 in both cell
proliferation and metastasis of osteosarcoma cells. Accumulating
evidence has demonstrated the tumor-suppressing role of P27 in
human cervical cancer (20),
pancreatic cancer (21), papillary
thyroid cancer (44) and other types
of cancer. Therefore, the downregulation of P21 and P27 increased
by high expression levels of SLC25A10 in osteosarcoma cells may
also mediate the tumor-promoting role of SLC25A10. The
SLC25A10/CCNE1/P2/P27 signaling pathway may play an important role
in human osteosarcoma.
Previous studies suggested that SLC25A10 was
involved in sustaining metabolic shift and redox balance (10,13).
Abnormal redox condition was found to be associated with abnormal
expression of P21 and P27, thus influencing cell proliferation and
mitosis (45,46). CCNE1 is an important gene involved in
cell cycle (47). Therefore,
SLC25A10 may regulate the expression of CCNE1, P21 and P27 by
influencing the redox state in osteosarcoma cells. Moreover, P21
may be regulated by SLC25A10. However, a previous study
demonstrated that the upstream factor TP53 in the TP53/P21 pathway
showed no significant changes after knockdown of SLC25A10 (48). Therefore, SLC25A10 may regulate P21
through TP53-independent mechanisms in osteosarcoma cells. In
addition, knockdown of SLC25A10 may induce G1 cell cycle
arrest by undetermined mechanisms, thus increasing the expression
levels of P21, P27 and CCNE1. The changes in the levels of CCNE1,
P21 and P27 could be an indirect consequence of cell cycle arrest.
The exact molecular mechanisms underlying the regulation of CCNE1,
P21 and P27 by SLC25A10 in osteosarcoma cells require further
examination. The present study did not clarify the energy
metabolism, redox homeostasis and the location of SLC25A10 in
osteosarcoma cells, which was a limitation to the present study and
should be investigated in future experiments.
In summary, the present study identified an
oncogenic role for SLC25A10 in human osteosarcoma. In vitro
functional experiments and analysis of tumor tissues collected from
patients with osteosarcoma were performed. Decreased expression of
SLC25A10 was associated with less malignant features in
osteosarcoma cells and improved clinicopathological parameters and
prognosis in patients with osteosarcoma. In addition, CCNE1 was
positively regulated and P21/P27 were negatively regulated by
SLC25A10. The present results suggested that the
SLC25A10/CCNE1/P21/P27 pathway may be fundamental in human
osteosarcoma.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from Anhui
Province Natural Science Youth Funding Project (grant no.
1808085QH241) and Anhui Province Natural Science Funding Project
(grant no. 1808085MH243).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
GW, XC and BX designed experiments. GW, JX, CC and
JQ performed experiments. GW, JX, PS and ZP analyzed data. XC and
BX wrote the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by Institutional
Review Boards of Anhui Medical University. All participants signed
informed consent before admission.
Patient consent for publication
All patients signed informed consent for the
possible publication of the present study.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
SLC25A10
|
solute carrier 25 family member 10
|
|
RFS
|
relapse-free survival
|
|
OS
|
overall survival
|
|
shRNA
|
short hairpin RNA
|
|
PARP
|
poly ADP-ribose polymerase
|
References
|
1
|
Fuja DG, Rainusso NC, Shuck RL,
Kurenbekova L, Donehower LA and Yustein JT: Transglutaminase-2
promotes metastatic and stem-like phenotypes in osteosarcoma. Am J
Cancer Res. 8:1752–1763. 2018.PubMed/NCBI
|
|
2
|
Abarrategi A, Tornin J, Martinez-Cruzado
L, Hamilton A, Martinez-Campos E, Rodrigo JP, González MV, Baldini
N, Garcia-Castro J and Rodriguez R: Osteosarcoma: Cells-of-Origin,
cancer stem cells, and targeted therapies. Stem Cells Int.
2016:36317642016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhao X, Sun S, Xu J, Luo Y, Xin Y and Wang
Y: MicroRNA-152 inhibits cell proliferation of osteosarcoma by
directly targeting Wnt/β-catenin signaling pathway in a
DKK1-dependent manner. Oncol Rep. 40:767–774. 2018.PubMed/NCBI
|
|
4
|
He R, Wu JX, Zhang Y, Chen H and Yang L:
LncRNA LINC00628 overexpression inhibits the growth and invasion
through regulating PI3K/Akt signaling pathway in osteosarcoma. Eur
Rev Med Pharmaco Sci. 22:5857–5866. 2018.
|
|
5
|
Nie WB, Zhao LM, Guo R, Wang MX and Ye FG:
Circular RNA circ-NT5C2 acts as a potential novel biomarker for
prognosis of osteosarcoma. Eur Rev Med Pharmaco Sci. 22:6239–6244.
2018.
|
|
6
|
Zuo DQ, Shogren KL, Zang J, Jewison DE,
Waletzki BE, Miller AL II, Okuno SH, Cai Z, Yaszemski MJ and Maran
A: Inhibition of STAT3 blocks protein synthesis and tumor
metastasis in osteosarcoma cells. J Exp Clin Canc Res. 37:2442018.
View Article : Google Scholar
|
|
7
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tabak SA, Khalifa SE and Fathy Y: HER-2
Immunohistochemical Expression in Bone Sarcomas: A New hope for
patients with osteosarcoma. Open Access Maced J Med Sci.
6:1555–1560. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Meazza C and Scanagatta P: Metastatic
osteosarcoma: A challenging multidisciplinary treatment. Expert Rev
Anticancer Ther. 16:543–556. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhou XS, Paredes JA, Krishnan S, Curbo S
and Karlsson A: The mitochondrial carrier SLC25A10 regulates cancer
cell growth. Oncotarget. 6:9271–9283. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhao Q, Zhou XS, Curbo S and Karlsson A:
Metformin downregulates the mitochondrial carrier SLC25A10 in a
glucose dependent manner. Biochem Pharmacol. 156:444–450. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mizuarai S, Miki S, Araki H, Takahashi K
and Kotani H: Identification of dicarboxylate carrier Slc25a10 as
malate transporter in de novo fatty acid synthesis. J Biol Chem.
280:32434–32441. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hlouschek J, Ritter V, Wirsdorfer F, Klein
D, Jendrossek V and Matschke J: Targeting SLC25A10 alleviates
improved antioxidant capacity and associated radioresistance of
cancer cells induced by chronic-cycling hypoxia. Cancer Lett.
439:24–38. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kulyte A, Ehrlund A, Arner P and Dahlman
I: Global transcriptome profiling identifies KLF15 and SLC25A10 as
modifiers of adipocytes insulin sensitivity in obese women. PLoS
One. 12:e01784852017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huypens P, Pillai R, Sheinin T, Schaefer
S, Huang M, Odegaard ML, Ronnebaum SM, Wettig SD and Joseph JW: The
dicarboxylate carrier plays a role in mitochondrial malate
transport and in the regulation of glucose-stimulated insulin
secretion from rat pancreatic beta cells. Diabetologia. 54:135–145.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lin Y, Berg AH, Iyengar P, Lam TK, Giacca
A, Combs TP, Rajala MW, Du X, Rollman B, Li W, et al: The
hyperglycemia-induced inflammatory response in adipocytes: The role
of reactive oxygen species. J Biol Chem. 280:4617–4626. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lockwood WW, Stack D, Morris T, Grehan D,
O'Keane C, Stewart GL, Cumiskey J, Lam WL, Squire JA, Thomas DM and
O'Sullivan MJ: Cyclin E1 is amplified and overexpressed in
osteosarcoma. J Mol Diagn. 13:289–296. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao ZM, Yost SE, Hutchinson KE, Li SM,
Yuan YC, Noorbakhsh J, Liu Z, Warden C, Johnson RM, Wu X, et al:
CCNE1 amplification is associated with poor prognosis in patients
with triple negative breast cancer. BMC Cancer. 19:962019.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sang Y, Zhang R, Sun L, Chen KK, Li SW,
Xiong L, Peng Y, Zeng L and Huang G: MORF4L1 suppresses cell
proliferation, migration and invasion by increasing p21 and
E-cadherin expression in nasopharyngeal carcinoma. Oncol Lett.
17:294–302. 2019.PubMed/NCBI
|
|
20
|
Guo Q, Xiong Y, Song Y, Hua K and Gao S:
ARHGAP17 suppresses tumor progression and up-regulates P21 and P27
expression via inhibiting PI3K/AKT signaling pathway in cervical
cancer. Gene. 692:9–16. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li Z, Tao Y, Wang X, Jiang P, Li J, Peng
M, Zhang X, Chen K, Liu H, Zhen P, et al: Tumor-secreted exosomal
miR-222 promotes tumor progression via regulating P27 Expression
and Re-localization in pancreatic cancer. Cell Physiol Biochem.
51:610–629. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen CY, Lei J, Zheng Q, Tan S, Ding KS
and Yu CJ: Poly(rC) binding protein 2 (PCBP2) promotes the
viability of human gastric cancer cells by regulating CDK2. FEBS
Open Bio. 8:764–773. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen C, Zheng Q, Kang W and Yu C: Long
non-coding RNA LINC00472 suppresses hepatocellular carcinoma cell
proliferation, migration and invasion through miR-93-5p/PDCD4
pathway. Clin Res Hepatol Gastroenterol. 43:436–445. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wu X, Cai ZD, Lou LM and Zhu YB:
Expressions of p53, c-MYC, BCL-2 and apoptotic index in human
osteosarcoma and their correlations with prognosis of patients.
Cancer Epidemiol. 36:212–216. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cao W, Fang L, Teng S, Chen H and Liu T:
MicroRNA-466 inhibits osteosarcoma cell proliferation and induces
apoptosis by targeting CCND1. Exp Ther Med. 16:5117–5122.
2018.PubMed/NCBI
|
|
26
|
Wang J, Xu G, Shen F and Kang Y: miR-132
targeting cyclin E1 suppresses cell proliferation in osteosarcoma
cells. Tumour Biol. 35:4859–4865. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu Q, Geng P, Shi L, Wang Q and Wang P:
miR-29 promotes osteosarcoma cell proliferation and migration by
targeting PTEN. Oncol Lett. 17:883–890. 2019.PubMed/NCBI
|
|
28
|
Zhao R, He H, Zhu Y, Wan J, Li Y, Gao S
and Zhang C: MiR-204/14-3-3ζ axis regulates osteosarcoma cell
proliferation through SATA3 pathway. Pharmazie. 72:593–598.
2017.PubMed/NCBI
|
|
29
|
Wang J, Wang G, Li B, Qiu C and He M:
miR-141-3p is a key negative regulator of the EGFR pathway in
osteosarcoma. Onco Targets Ther. 11:4461–4478. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lv H, Gao G, Zhang L and Sun Y: Polo-like
kinase 3 inhibits osteosarcoma cell proliferation and tumorigenesis
via cooperative interaction with p21. Mol Med Rep. 12:6789–6796.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang F and Peng H: LncRNA-ANCR regulates
the cell growth of osteosarcoma by interacting with EZH2 and
affecting the expression of p21 and p27. J Orthop Surg Res.
12:1032017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ghosh T, Varshney A, Kumar P, Kaur M,
Kumar V, Shekhar R, Devi R, Priyanka P, Khan MM and Saxena S:
MicroRNA-874-mediated inhibition of the major G1/S phase
cyclin, CCNE1, is lost in osteosarcomas. J Biol Chem.
292:21264–21281. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shukla K, Sharma AK, Ward A, Will R,
Hielscher T, Balwierz A, Breunig C, Münstermann E, König R,
Keklikoglou I and Wiemann S: MicroRNA-30c-2-3p negatively regulates
NF-κB signaling and cell cycle progression through downregulation
of TRADD and CCNE1 in breast cancer. Mol Oncol. 9:1106–1119. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yao Y, Luo J, Sun Q, Xu T, Sun S, Chen M,
Lin X, Qian Q, Zhang Y, Cao L, et al: HOXC13 promotes proliferation
of lung adenocarcinoma via modulation of CCND1 and CCNE1. Am J
Cancer Res. 7:1820–1834. 2017.PubMed/NCBI
|
|
35
|
Zhang Y, Peng Z, Zhao YS and Chen L:
microRNA-25 inhibits cell apoptosis of human gastric adenocarcinoma
cell line AGS via regulating CCNE1 and MYC. Med Sci Monitor.
22:1415–1420. 2016. View Article : Google Scholar
|
|
36
|
Zhang X, Hu SJ, Zhang X, Wang L, Zhang X,
Yan B, Zhao J, Yang A and Zhang R: MicroRNA-7 arrests cell cycle in
G1 phase by directly targeting CCNE1 in human hepatocellular
carcinoma cells. Biochem Biophys Res Commun. 443:1078–1084. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fujii R, Osaka E, Sato K and Tokuhashi Y:
MiR-1 suppresses proliferation of osteosarcoma cells by
Up-regulating p21 via PAX3. Cancer Genom Proteom. 16:71–79. 2019.
View Article : Google Scholar
|
|
38
|
Huang J, Deng GR, Liu TM, Chen WZ and Zhou
Y: Long noncoding RNA PCAT-1 acts as an oncogene in osteosarcoma by
reducing p21 levels. Biochem Biophys Res Commun. 495:2622–2629.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ding Y, Wang YC, Chen J, Hu Y, Cao Z, Ren
P and Zhang Y: p21 overexpression sensitizes osteosarcoma U2OS
cells to cisplatin via evoking caspase-3 and Bax/Bcl-2 cascade.
Tumor Biol. 35:3119–3123. 2014. View Article : Google Scholar
|
|
40
|
Li Z, Yu D, Li H, Lv Y and Li S: Long
noncoding RNA UCA1 confers tamoxifen resistance in breast cancer
endocrinotherapy through regulation of the EZH2/p21 axis and the
PI3K/AKT signaling pathway. Int J Oncol. 54:1033–1042.
2019.PubMed/NCBI
|
|
41
|
Guo L, Gu J, Hou S, Liu D, Zhou M, Hua T,
Zhang J, Ge Z and Xu J: Long non-coding RNA DANCR promotes the
progression of non-small-cell lung cancer by inhibiting p21
expression. Onco Targets Ther. 12:135–146. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liao QD, Zhong D and Chen Q: Protein
expression of Skp2 in osteosarcoma and its relation with prognosis.
Zhong Nan Da Xue Xue Bao Yi Xue Ban. 33:606–611. 2008.(In Chinese).
PubMed/NCBI
|
|
43
|
Hu XH, Zhao ZX, Dai J, Geng DC and Xu YZ:
MicroRNA-221 regulates osteosarcoma cell proliferation, apoptosis,
migration, and invasion by targeting CDKN1B/p27. J Cell Biochem.
120:4665–4674. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li H, Guan H, Guo Y, Liang W, Liu L, He X,
Ke W, Cao X, Xiao H and Li Y: CITED1 promotes proliferation of
papillary thyroid cancer cells via the regulation of p21 and p27.
Cell Biosci. 8:572018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Vijay K, Sowmya PR, Arathi BP, Shilpa S,
Shwetha HJ, Raju M, Baskaran V and Lakshminarayana R: Low-dose
doxorubicin with carotenoids selectively alters redox status and
upregulates oxidative stress-mediated apoptosis in breast cancer
cells. Food Chem Toxicol. 118:675–690. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhang D, Wang Y, Liang Y, Zhang M, Wei J,
Zheng X, Li F, Meng Y, Zhu NW, Li J, et al: Loss of p27 upregulates
MnSOD in a STAT3-dependent manner, disrupts intracellular redox
activity and enhances cell migration. J Cell Sci. 127:2920–2933.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Tang Z, Fang Y and Du R: MicroRNA-107
induces cell cycle arrests by directly targeting cyclin E1 in
ovarian cancer. Biochem Biophys Res Commun. 512:331–337. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yang M, Peng Y, Liu W, Zhou M, Meng Q and
Yuan C: Sirtuin 2 expression suppresses oxidative stress and
senescence of nucleus pulposus cells through inhibition of the
p53/p21 pathway. Biochem Biophys Res Commun. 513:616–622. 2019.
View Article : Google Scholar : PubMed/NCBI
|