Introduction
Neuroblastoma (NB) is the most common extracranial
solid tumor that occurs in children, with >90% of patients being
diagnosed before the age of 10 years (1). In fact, it is estimated that NB
affected 10.72/1,000,000 children <15 years each year in the
United States between 2001–2009 (2).
At the time of diagnosis, 50% of all NB cases present with
metastasis, which are classified as a high-risk clinical phenotype
(3). Although the treatment of
high-risk NB has significantly improved in the past few decades,
the long-term survival rate remains <50%, which is markedly
lower compared with other pediatric oncology diseases (4).
Apatinib is a novel tyrosine kinase inhibitor of the
vascular endothelial growth factor receptor-2 (VEGFR2) (5). At the cellular level, apatinib has been
reported to selectively block the downstream signal transduction of
VEGFR2 (6). Apatinib has previously
been demonstrated to exhibit promising antitumor effects in various
types of tumor in vitro, such as osteosarcoma, anaplastic
thyroid cancer, lung cancer and hepatocellular carcinoma (7–10). In
addition, in a phase 3 clinical trial, apatinib was identified to
significantly improve the median overall and progression-free
survival time of advanced gastric cancer, which was refractory to
fewer than two lines of prior chemotherapy (11). However, to the best of our knowledge,
the antitumor effects of apatinib in NB remain unclear.
The antitumor effects of apatinib reportedly act
downstream of the VEGFR2 signaling pathway, such as via
PI3K/AKT/mTOR, STAT3/Bcl-2 or mitogen-activated protein kinase
(MAPK)/ERK (7,12,13). In
particular, it has been reported that the upregulation of
phosphorylated (p)-AKT, p-mTOR and MAPK are recurrent events
occurring in NB cell lines, and predict a poor prognosis for
patients with NB (14–16).
The present study evaluated the antitumor effects of
apatinib and revealed the main signaling pathway affecting the
induction of apoptosis and autophagy in NB cells in
vitro.
Materials and methods
Chemicals
Apatinib was obtained from Jiangsu Hengrui Medicine
Co., Ltd.. Apatinib (0.5 mg) was dissolved in 1 ml DMSO to a
concentration of 1 mM and then diluted with DMEM (cat. no.
10566016; Thermo Fisher Scientific, Inc.) for cell culture.
Cell culture
NB cell lines [BE(2)-M17, SH-SY5Y and IMR-32] were
purchased from the American Type Culture Collection. The cells were
cultured in DMEM supplemented with 10% FBS (cat. no. 10100147C;
Thermo Fisher Scientific, Inc.) and maintained at 37°C with 5%
CO2.
Cell viability, proliferation, cell
cycle, apoptosis assays and autophagy observation
Cell viability assays were performed using a Cell
Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.) assay.
Briefly, the NB cells were plated at a density of 5×103
cells/well into 96-well plates in triplicate and incubated for 24
h. The effects on harvested cells treated with 20 µM near the
IC50 concentration of apatinib were too poor to perform
the cell cycle assay and other experiments; therefore, lower
concentrations of apatinib (5 or 10 µM) were used in the present
study and the results demonstrated an obvious effect of apatinib on
NB cells. Subsequently, the cells were cultured with fresh medium
containing 0, 5 or 10 µM apatinib for 24 or 48 h and the absorbance
was subsequently measured at 450 nm to assess the effect on cell
proliferation. The cell cycle and cell apoptosis assays were
performed as previously described (17). For autophagy observation, the NB
cells were cultured with 10 µM apatinib at 37°C with 5%
CO2 for 48 h. Then the cellular vacuolization of
neuroblastoma cells was visualized via light microscopy (×400
magnification; CKX41; Olympus Corporation, Inc.)
Colony formation assay
BE(2)-M17 and SH-SY5Y cells were treated with 0, 5
or 10 µM apatinib for 48 h, and then 1×103 cells were
seeded and cultured in 6-well plates without apatinib. After 14
days of incubation, the cells were fixed with 100% anhydrous
methanol at room temperature for 10 min and stained with 0.5%
crystal violet at room temperature for 20 min. The colony numbers
were observed using a G:BOX Chemi XRQ (Synoptics Ltd., http://www.syngene.com). All experiments were
performed in triplicate.
Wound healing and invasion assays
BE(2)-M17 and SH-SY5Y cells were seeded at a density
of 5×104 cells/well into 6-well plates and cultured
until they reached 100% confluency. Then, pipette tips were used to
make a scratch in the monolayer of the cells. Due to the poor
adhesion ability of neuroblastoma cells in medium without FBS, the
cells were cultured in DMEM supplemented with 2% FBS and 0, 5 or 10
µM apatinib. Although the medium with 2% FBS had little effect on
the proliferation of neuroblastoma cells in 24 h, there was a
limitation of our wound healing experiment. The wounded area was
monitored every 6 h under a light microscope (×100 magnification;
CKX41; Olympus Corporation) and the degree of wound healing induced
by cell migration was quantified using Image-Pro Plus software 6.0
(Media Cybernetics, Inc.). The invasion assay was performed as
previously described (18). For cell
invasion analysis, 1×105 cells in serum-free DMEM were
plated in the upper membrane of Transwell inserts (8 mm) coated
with 60 µl Matrigel (BD Biosciences) in the 24-well plates.
High-glucose DMEM containing 20% FBS was added to the lower wells.
Migration was allowed to proceed for 24 h at 37°C. Cells that did
not migrate through the filters were removed using cotton swabs,
and cells that migrated through the inserts were fixed in anhydrous
methanol for 10 min and stained with 0.5% crystal violet at room
temperature for 20 min. The number of migratory cells were observed
under a light microscope (×200 magnification; CKX41; Olympus
Corporation). All experiments were performed in triplicate.
Bioinformatics analysis
The molecular interactions between the VEGFR2 and
PI3K/AKT signaling pathways were analyzed using Search Tool for the
Retrieval of Interacting Genes/Proteins (STRING) database
(http://string-db.org). In the STRING database,
kinase insert domain-containing receptor (KDR) is the official
symbol name also known as VEGFR2, and phosphatidylinositol
4,5-bisphosphate 3-kinase catalytic subunit alpha isoform (PIK3CA)
is the official symbol name also known as PI3K (19).
Western blotting
Following treatment with apatinib (0, 5, 10 or 20
µM) at 37°C for 48 h in 5% CO2, total protein was
extracted from BE(2)-M17 and SH-SY5Y cells as previously described
(17). In the autophagy assay, the
starvation group was cultured without FBS at 37°C for 48 h. Total
protein was quantified using the BCA Protein Assay kit (cat. no.
23225; Thermo Fisher Scientific, Inc.). Then, 30 µg protein/lane
was separated on precast 10% SDS-PAGE gels and subsequently
transferred onto polyvinylidene fluoride membranes. The membranes
were blocked with 5% bovine serum albumin dissolved in
Tris-buffered saline containing 0.1% Tween-20 for 30 min at room
temperature followed by incubation with primary antibodies against:
Bcl-2 (cat. no. sc-7382; 1:200), Bax (cat. no. sc-7480, 1:200),
cyclin D1 (cat. no. sc-8396; 1:200), p70S6 kinase α (cat. no.
sc-230, 1:200), phosphorylated (p)-p70S6 kinase αSer411
(cat. no. sc-7983-R, 1:200), mTOR (cat. no. sc-517464, 1:200) and
p-mTORSer2448 (cat. no. sc-293133, 1:200) were purchased
from Santa Cruz Biotechnology, Inc., overnight at 4°C. The primary
antibody against β-actin (cat. no. A1978, 1:10,000) was purchased
from Sigma-Aldrich; Merck KGaA. The primary antibodies against AKT
(cat. no. 4691, 1:1,000), p-AKTSer473 (cat. no. 4058,
1:1,000), ERK (cat. no. 4696, 1:2,000),
p-ERKThr202/Tyr204 (cat. no. 4376, 1:1,000) and LC3B
(cat. no. 3868, 1:1,000) were purchased from Cell Signaling
Technology, Inc. The primary antibodies against autophagy related 5
(ATG5; cat. no. 10181-2-AP, 1:1,000) and P62 (cat. no. 18420-1-AP,
1:1,000) were purchased from Proteintech Group, Inc.. Following the
primary incubation, membranes were incubated with goat against
rabbit IgG-horseradish peroxidase (cat. no. ZB-2301, 1:10,000) and
goat against mouse IgG-horseradish peroxidase (cat. no. ZB-2305,
1:10,000; both from Beijing Zhongshan Golden Bridge Biotechnology
Co., Ltd.) secondary antibodies for 2 h at 37°C. Protein bands were
visualized using the Super ECL plus super sensitive luminous liquid
(cat. no. P1050; Applygen Technologies, Inc.). The images were
captured using the ImageQuant™ LAS 4000 system (GE Healthcare Life
Sciences). The densitometry of the scanned immunoblot bands was
analyzed using ImageJ software (version 1.52q; Bharti Airtel Ltd,
http://imagej.nih.gov/ij). Each experiment was
performed at least three times.
Immunofluorescence
BE(2)-M17 and SH-SY5Y cells were seeded at a density
of 1×105 cells/well onto glass coverslips in 12-well
plates and treated with 0 or 10 µM apatinib for 48 h at 37°C with
5% CO2. The NB cells were subsequently fixed for
immunofluorescence analysis as previously described (20). Following fixation, the cells were
penetrated and blocked with PBS containing 10% FBS and 0.4%
TritonX-100 at room temperature for 30 min. Subsequently, cells
were incubated with primary antibodies against LC3B (cat. no. 3868;
1:200; Cell Signaling Technology, Inc.) and Ki-67 (cat. no. 9129;
1:400; Cell Signaling Technology, Inc.) overnight at 4°C. Following
the primary incubation, cells were incubated with rabbit IgG
secondary antibody (Alexa Fluor 488, cat. no. ZF-0511; 1:2,000;
Beijing Zhongshan Golden Bridge Biotechnology Co., Ltd.) at room
temperature for 1 h and washed three times with PBS containing DAPI
(cat. no. D9524; 1:1,000; Sigma-Aldrich; Merck KGaA) at room
temperature for 3 min. Cells were observed under a fluorescence
microscope (magnification ×200; DM2500, Leica Microsystems, Ltd.)
or a laser confocal microscope (magnification ×400; SP8, Leica
Microsystems, Ltd.). Staining intensity was calculated using ImageJ
software (version 1.52q; National Institutes of Health).
Statistical analysis
Statistical analysis was performed using SPSS 19.0
software (IBM Corp.). All experiments were performed in triplicate
and data are presented as the mean ± standard deviation.
Statistical differences between two groups were determined using
unpaired Student's t-test, whereas one-way ANOVA followed by
Bonferroni's post hoc test were used for ≥2 groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
Apatinib inhibits cell proliferation
and the colony formation of NB cells
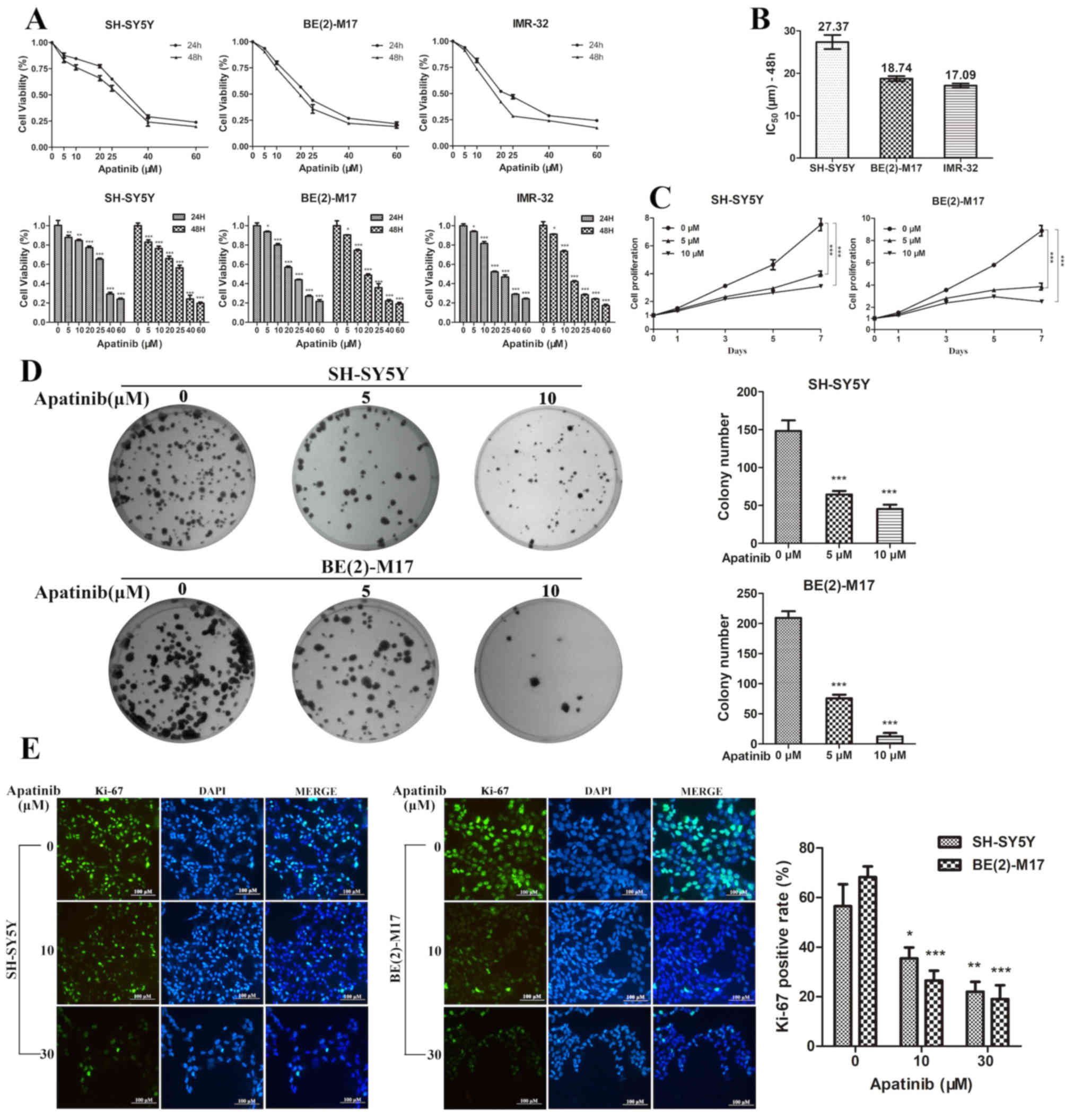
To determine the antitumor effects of apatinib on NB
cells, the NB cell lines BE(2)-M17, SH-SY5Y and IMR-32 were
cultured with different concentrations of apatinib. The CCK-8 assay
revealed that the viability and proliferation of NB cells were
significantly inhibited by apatinib in both a dose- and
time-dependent manner (Fig. 1A and
C). At 48 h, the IC50 of apatinib was 18.74 µM for
BE(2)-M17 cells, 27.37 µM for SH-SY5Y cells and 17.09 µM for IMR-32
cells (Fig. 1B). Neuroblastoma is
frequently associated with the presence of MYCN amplification, a
genetic biomarker for poor prognosis (4). The SH-SY5Y cell line without MYCN
amplification and BE(2)-M17 cell line with MYCN amplification were
selected for further experimentation. Subsequently, BE(2)-M17 and
SH-SY5Y cells were treated with apatinib for 48 h and then seeded
into 6-well plates for 14 days. Apatinib significantly decreased
the colony formation efficiency of BE(2)-M17 and SH-SY5Y cells
(Fig. 1D). In addition, the
immunofluorescence assay demonstrated that the expression level of
Ki-67 was significantly decreased following apatinib treatment
(Fig. 1E). These results indicated
that apatinib may inhibit the growth of NB cells in
vitro.
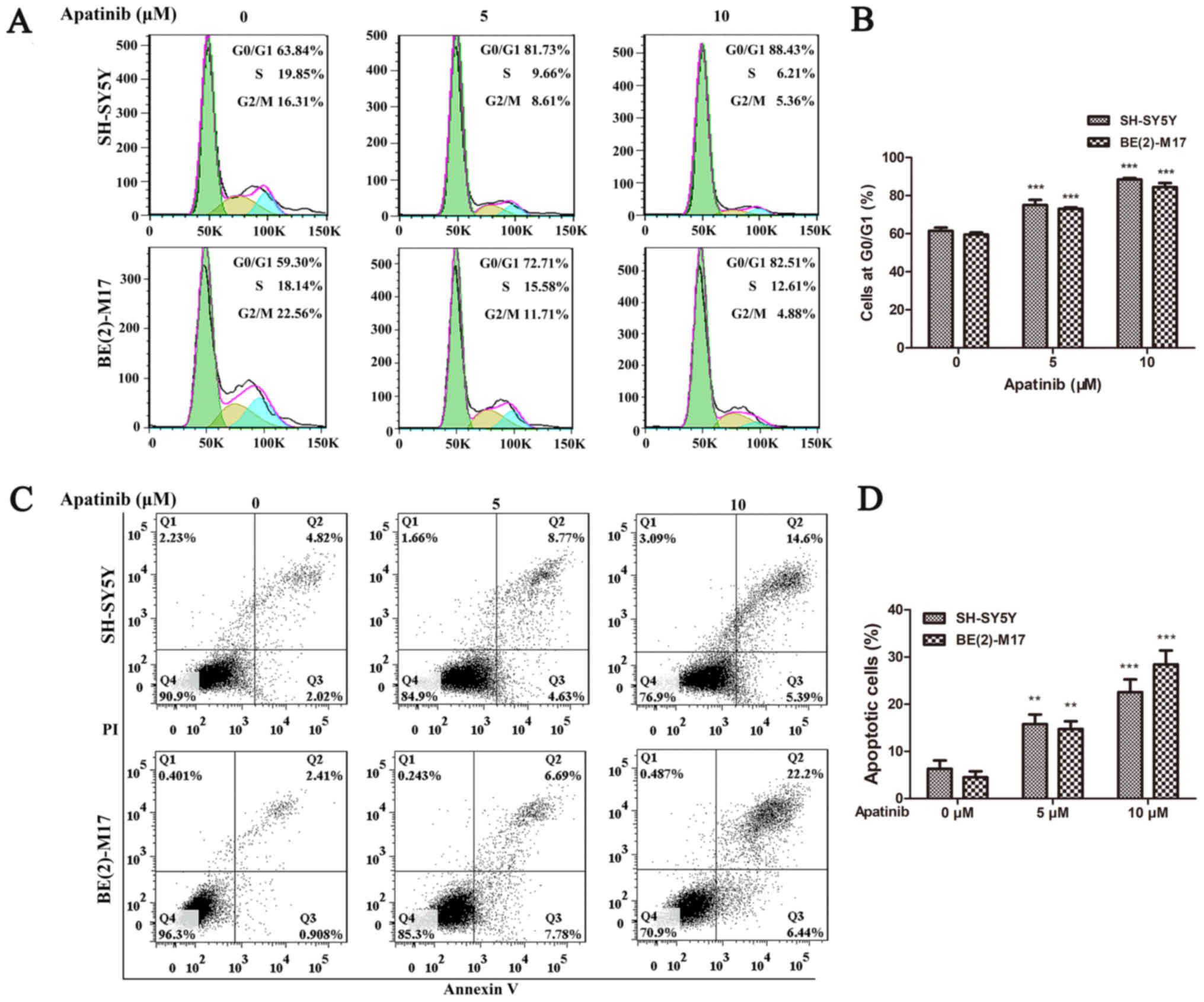
Apatinib induces cell cycle arrest and
apoptosis of NB cells
To determine the potential mechanism of cell
proliferation inhibition, the effects of apatinib on the cell cycle
of BE(2)-M17 and SH-SY5Y cells were analyzed using flow cytometry.
The cell cycle was significantly arrested at the
G0/G1 phase in BE(2)-M17 and SH-SY5Y cells
following treatment with 5 or 10 µM apatinib for 48 h (Fig. 2A and B). Furthermore, flow cytometric
analysis also revealed that apatinib significantly increased the
percentage of apoptotic cells in both the BE(2)-M17 and SH-SY5Y
cell lines in a concentration-dependent manner (Fig. 2C and D). These findings indicated
that apatinib may induce cell cycle arrest and the apoptosis of NB
cells.
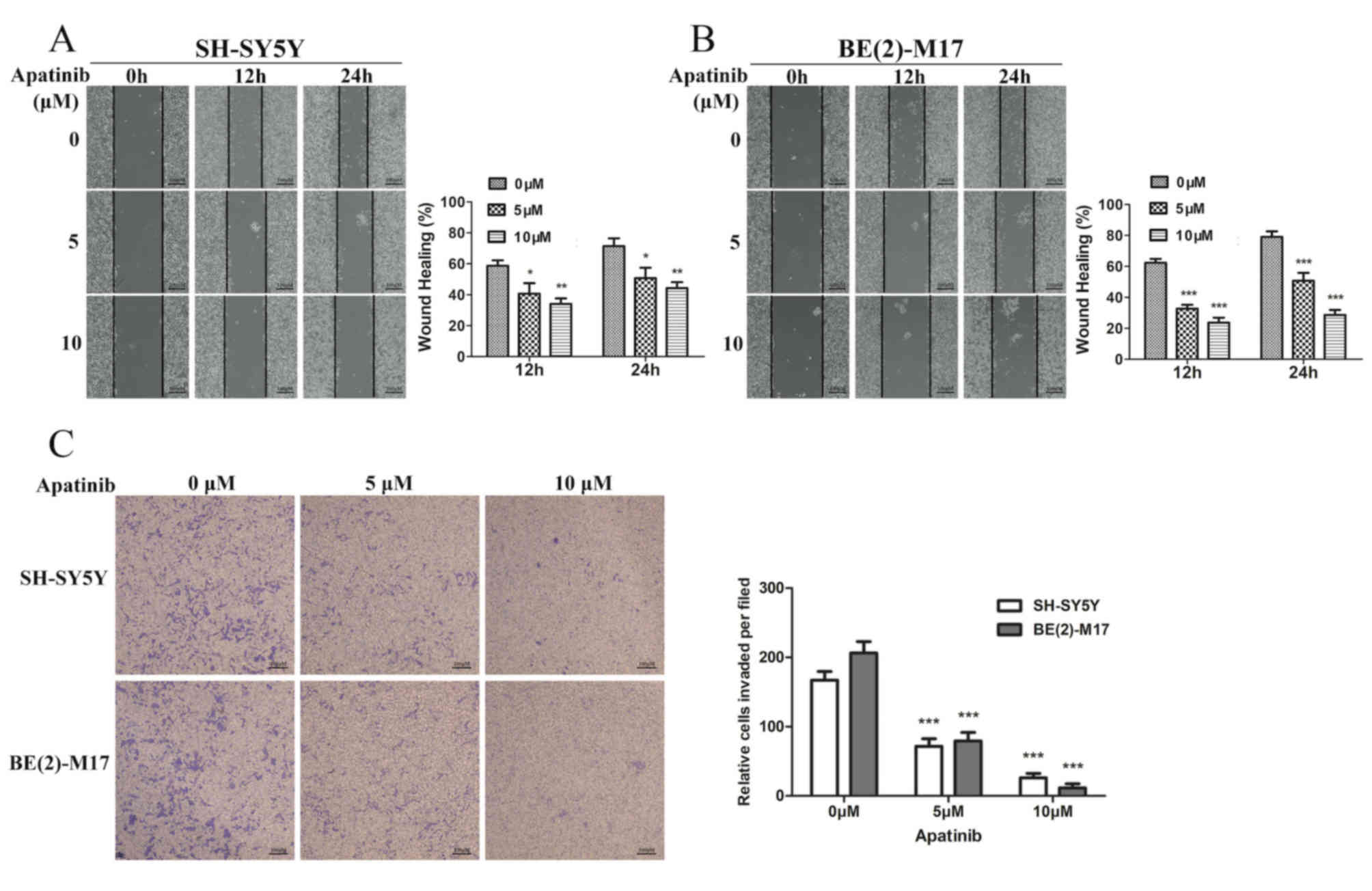
Apatinib inhibits the migration and
invasion of NB cells
To investigate whether apatinib affects the
processes of invasion and migration in NB cells, wound healing and
Transwell assays were performed. The wound healing assay revealed
that apatinib significantly suppressed the migration of BE(2)-M17
and SH-SY5Y cells (Fig. 3A and B).
In addition, the invasive abilities of NB cells were significantly
reduced following treatment with apatinib for 24 h (Fig. 3C). These results suggested that
apatinib may be a potential anti-metastatic agent for NB.
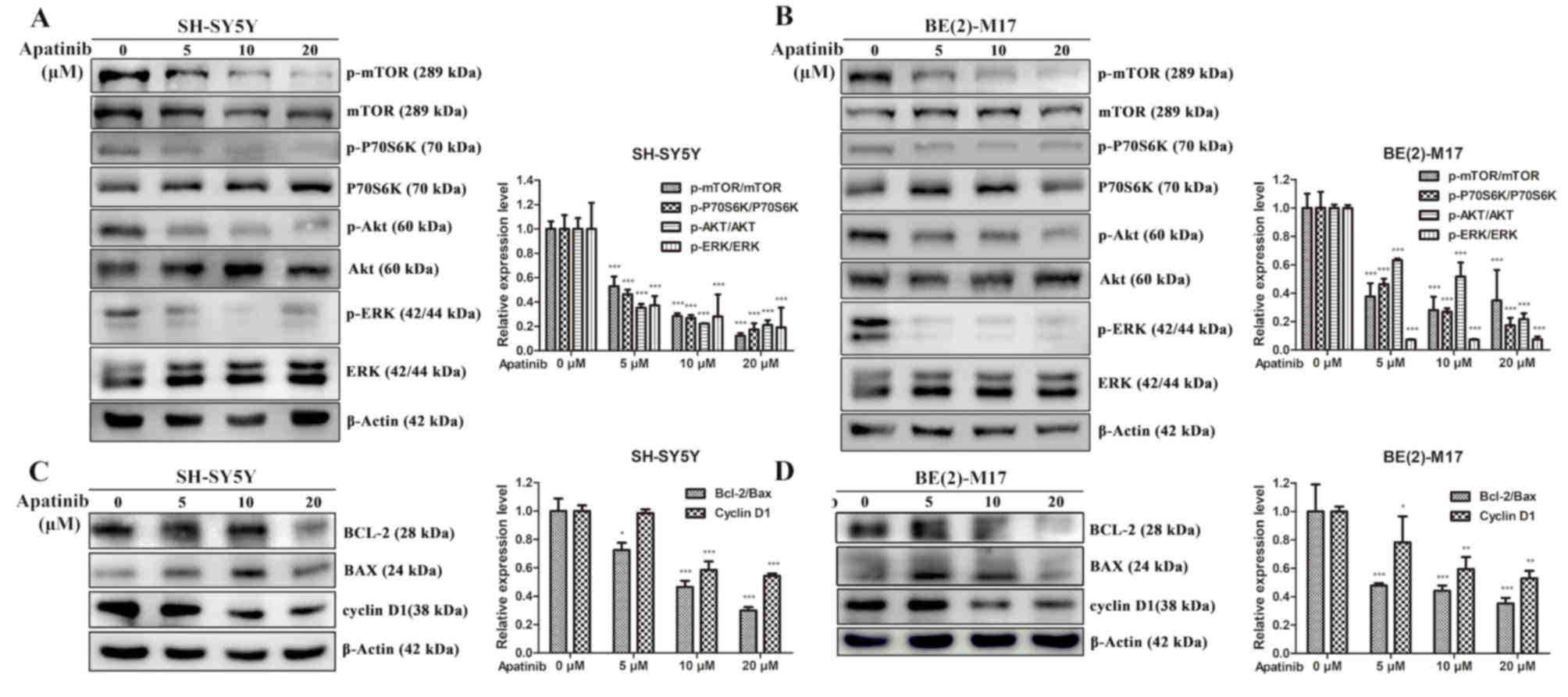
Apatinib inhibits the PI3K/AKT/mTOR
and MAPK/ERK signaling pathways
Bioinformatics analysis revealed molecular
interactions between the VEGFR2 and PI3K/AKT signaling pathway
(Fig. S1). Western blotting
subsequently demonstrated that the expression ratios of p-AKT/AKT,
p-mTOR/mTOR and p-p70S6K/p70S6K were significantly decreased by
apatinib treatment in NB cells (Fig. 4A
and B). Since MAPK/ERK is the downstream signaling pathway of
VEGFR2 (21), the present study also
demonstrated that apatinib inhibited the phosphorylation of ERK in
NB cells (Fig. 4C and D). As
apatinib induced cell cycle arrest and the apoptosis of NB cells,
the expression levels of Bcl-2, Bax and cyclin D1 following the
treatment with apatinib were also investigated (Fig. 4C and D). The results demonstrated
that as the concentration of apatinib increased, the expression of
Bcl-2 decreased, while the expression of Bax increased first and
then decreased in the SH-SY5Y and BE(2)-M17 cell lines,
respectively. It was speculated that the accumulation of apoptotic
cells was induced by a high concentration of apatinib, which
decreased Bax protein expression. However, after the gray value
statistics, the Bcl-2/Bax ratio, which determines the direction of
apoptosis, was decreased. It was also identified that apatinib
significantly reduced the expression levels of cyclin D1 in NB
cells, which is important for the progression of the cell cycle.
Altogether, these findings suggested that apatinib may induce
apoptosis through inhibition of the PI3K/AKT/mTOR and MAPK/ERK
signalling pathways.
Apatinib induces autophagy in NB
cells
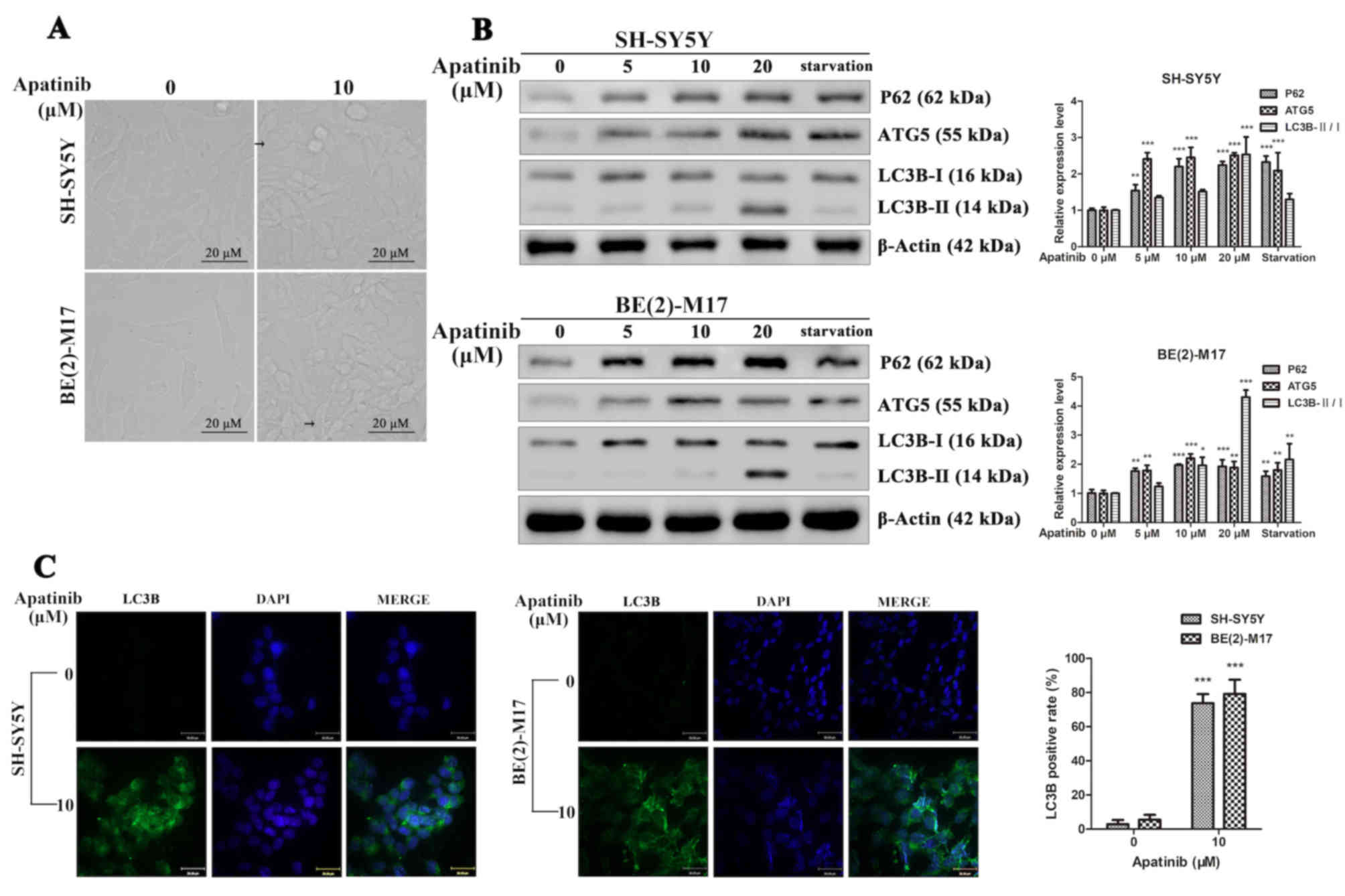
Furthermore, under the microscope, it was observed
that apatinib induced the vacuolization of NB cells (Fig. 5A). Thus, the expression levels of
p62, ATG5, LC3-I and LC3-II, which are important proteins involved
in the initial stages of autophagy (22), were analyzed. LC3B is the main
isoforms of LC3 and was used as an autophagy indicator. The
starvation group was used as a positive control of autophagy. The
western blotting analysis revealed that following apatinib
treatment, the expression levels of p62, ATG5 and LC3B-II/I were
significantly increased in NB cells, which was consistent with the
starvation group (Fig. 5B).
Furthermore, NB cells exhibited a dot fluorescence pattern of LC3B
following treatment with apatinib (Fig.
5C). In addition, following apatinib treatment, the ratios of
LC3B positive cells in the SH-SY5Y and BE(2)-M17 cell lines
significantly increased compared with the control groups. These
results suggested that apatinib may induce autophagy in NB
cells.
Discussion
Apatinib, a highly selective tyrosine kinase
inhibitor to VEGFR2, has been reported to exert antitumor effects
in various types of tumors, such as osteosarcoma, thyroid cancer,
cervical cancer and gastric cancer (7,8,23,24). The
present study aimed to demonstrate the potential antitumor effects
of apatinib in NB cells in vitro. It was identified that
apatinib induced apoptosis and autophagy in NB cells, which mainly
occurred via the PI3K/AKT/mTOR and MAPK/ERK signaling pathways.
The induction of angiogenesis is a hallmark of
cancer (25) and an important step
for cancer metastasis (26). It is
reported that ~50% of newly diagnosed patients with NB present with
metastasis, usually at the site of the bone or bone marrow
(3). Unfortunately, patients with NB
that present with bone or bone marrow metastasis have a poor
prognosis (27). VEGFR serves an
important role in the process of angiogenesis in cancer (28), and the use of VEGFR tyrosine kinase
inhibitors, such as axitinib, sorafenib and sunitinib, has been
revealed to effectively inhibit the angiogenesis of NB both in
vitro and in vivo (29–31).
It has been reported that apatinib induces apoptosis
and autophagy via the VEGFR2/STAT3/Bcl-2 signalling pathway in
osteosarcoma and via ER stress in colorectal cancer (7,32). In
the present study, apatinib was observed to inhibit cell
proliferation, colony formation, and the migration and invasion of
NB cells in vitro. The inhibitory effects of apatinib were
associated with apoptosis and cell cycle arrest in both a time- and
dose-dependent manner. Subsequently, western blotting analysis
revealed that the Bax/Bcl-2 ratio was increased, which is
associated with cell apoptosis (33), whilst the expression level of cyclin
D1 was downregulated, which is associated with arrest of the cell
cycle (34). Furthermore, apatinib
inhibited the colony formation, migration and invasion of NB cells.
The PI3K/AKT/mTOR and MAPK/ERK signaling pathways regulate numerous
cellular processes and physiological functions, including cell
proliferation, cell migration, cell metabolism and cell survival
(35,36). PI3K/AKT/mTOR and MAPK/ERK act
downstream of the VEGFR2 signaling pathway (37). The findings of the present study also
demonstrated that apatinib downregulated the phosphorylation of
AKT, mTOR, p70S6K and ERK.
Autophagy serves an important role in the
progression and treatment of cancer (38). A crucial regulator of autophagy is
the nutrient sensor mTOR, which negatively regulates autophagy
(38–40). A previous study has revealed that
Bcl-2 suppresses autophagy and that inhibiting Bcl-2 promotes the
autophagy of cells (41). The
presence of LC3 in autophagosomes and the conversion of LC3 to the
lower migrating form, LC3-II, are considered to be indicators of
autophagy (42). ATG5, which
associates with the phagophore, and p62, which targets specific
substrates to autophagosomes, may also be used as measurements of
the autophagic flux (38,43). In the present study, the expression
levels of LC3-II, p62 and ATG5 were significantly upregulated,
while the expression levels of Bcl-2 and p-mTOR were significantly
inhibited following the treatment with apatinib. These results may
further explain the autophagic mechanism of apatinib. However,
further in vivo analysis is required to determine whether
apatinib can exert antitumor effects and even suppress the
metastasis of neuroblastoma by inhibiting angiogenesis via
VEGFR2.
In conclusion, to the best of our knowledge, the
present study was the first to reveal the antitumor effects of
apatinib in NB cells in vitro. The findings suggested that
apatinib may inhibit the VEGFR2-dependent PI3K/AKT/mTOR and
MAPK/ERK signaling pathways, further to inducing the apoptosis and
autophagy of NB cells. These findings may provide a basis for
future clinical trials to investigate whether apatinib could be
used for the treatment of NB.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81972572 and
81602138), CAMS Innovation Fund for Medical Sciences (grant no.
2016-I2M-1-001) and the Capital's Funds for Health Improvement and
Research (grant no. 2018-2-2095).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding authors upon reasonable
request.
Authors' contributions
WJ, MJ and XM were involved in the conception and
design of the study. XY, HF and WZ performed the cellular and
molecular experiments. XY, HF, XJ and YY assisted with the
statistical analysis. XY and HF were major contributors in drafting
the initial manuscript. All authors read and approved the final
version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Matthay KK, Maris JM, Schleiermacher G,
Nakagawara A, Mackall CL, Diller L and Weiss WA: Neuroblastoma. Nat
Rev Dis Primers. 2:160782016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel DA, King J, Tai E, Buchanan N,
Ajani UA and Li J: Cancer incidence rates and trends among children
and adolescents in the United States, 2001–2009. Pediatrics.
134:e945–e955. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Morandi F, Corrias MV and Pistoia V:
Evaluation of bone marrow as a metastatic site of human
neuroblastoma. Ann N Y Acad Sci. 1335:23–31. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pinto NR, Applebaum MA, Volchenboum SL,
Matthay KK, London WB, Ambros PF, Nakagawara A, Berthold F,
Schleiermacher G, Park JR, et al: Advances in risk classification
and treatment strategies for neuroblastoma. J Clin Oncol.
33:3008–3017. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hu L, Sun F, Sun Z, Ni X, Wang J, Wang J,
Zhou M, Feng Y, Kong Z, Hua Q and Yu J: Apatinib enhances the
radiosensitivity of the esophageal cancer cell line KYSE-150 by
inducing apoptosis and cell cycle redistribution. Oncol Lett.
17:1609–1616. 2019.PubMed/NCBI
|
|
6
|
Tian S, Quan H, Xie C, Guo H, Lü F, Xu Y,
Li J and Lou L: YN968D1 is a novel and selective inhibitor of
vascular endothelial growth factor receptor-2 tyrosine kinase with
potent activity in vitro and in vivo. Cancer Sci. 102:1374–1380.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu K, Ren T, Huang Y, Sun K, Bao X, Wang
S, Zheng B and Guo W: Apatinib promotes autophagy and apoptosis
through VEGFR2/STAT3/BCL-2 signaling in osteosarcoma. Cell Death
Dis. 8:e30152017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jin Z, Cheng X, Feng H, Kuang J, Yang W,
Peng C, Shen B and Qiu W: Apatinib inhibits angiogenesis via
suppressing Akt/GSK3β/ANG signaling pathway in anaplastic thyroid
cancer. Cell Physiol Biochem. 44:1471–1484. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang C and Qin S: Apatinib targets both
tumor and endothelial cells in hepatocellular carcinoma. Cancer
Med. 7:4570–4583. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lin C, Wang S, Xie W, Zheng R, Gan Y and
Chang J: Apatinib inhibits cellular invasion and migration by
fusion kinase KIF5B-RET via suppressing RET/Src signaling pathway.
Oncotarget. 7:59236–59244. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li J, Qin S, Xu J, Xiong J, Wu C, Bai Y,
Liu W, Tong J, Liu Y, Xu R, et al: Randomized, double-blind,
placebo-controlled phase III trial of apatinib in patients with
chemotherapy-refractory advanced or metastatic adenocarcinoma of
the stomach or gastroesophageal junction. J Clin Oncol.
34:1448–1454. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Feng H, Cheng X, Kuang J, Chen L, Yuen S,
Shi M, Liang J, Shen B, Jin Z, Yan J and Qiu W: Apatinib-induced
protective autophagy and apoptosis through the AKT-mTOR pathway in
anaplastic thyroid cancer. Cell Death Dis. 9:10302018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang M, Huang B, Li G and Zeng S:
Apatinib affect VEGF-mediated cell proliferation, migration,
invasion via blocking VEGFR2/RAF/MEK/ERK and PI3K/AKT pathways in
cholangiocarcinoma cell. BMC Gastroenterol. 18:1692018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Santo EE, Stroeken P, Sluis PV, Koster J,
Versteeg R and Westerhout EM: FOXO3a is a major target of
inactivation by PI3K/AKT signaling in aggressive neuroblastoma.
Cancer Res. 73:2189–2198. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mei H, Wang Y, Lin Z and Tong Q: The mTOR
signaling pathway in pediatric neuroblastoma. Pediatr Hematol
Oncol. 30:605–615. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lambertz I, Kumps C, Claeys S, Lindner S,
Beckers A, Janssens E, Carter DR, Cazes A, Cheung BB, De Mariano M,
et al: Upregulation of MAPK negative feedback regulators and RET in
mutant ALK neuroblastoma: Implications for targeted treatment. Clin
Cancer Res. 21:3327–3339. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jiang XR, Yu XY, Fan JH, Guo L, Zhu C,
Jiang W and Lu SH: RFT2 is overexpressed in esophageal squamous
cell carcinoma and promotes tumorigenesis by sustaining cell
proliferation and protecting against cell death. Cancer Lett.
353:78–86. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xue L, Yu X, Jiang X, Deng X, Mao L, Guo
L, Fan J, Fan Q, Wang L and Lu SH: TM4SF1 promotes the self-renewal
of esophageal cancer stem-like cells and is regulated by miR-141.
Oncotarget. 8:19274–19284. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wu HB, Yang S, Weng HY, Chen Q, Zhao XL,
Fu WJ, Niu Q, Ping YF, Wang JM, Zhang X, et al: Autophagy-induced
KDR/VEGFR-2 activation promotes the formation of vasculogenic
mimicry by glioma stem cells. Autophagy. 13:1528–1542. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dong Z, Zhu C, Zhan Q and Jiang W: Cdk
phosphorylation licenses Kif4A chromosome localization required for
early mitotic progression. J Mol Cell Biol. 10:358–370. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sun J, Huang W, Yang SF, Zhang XP, Yu Q,
Zhang ZQ, Yao J, Li KR, Jiang Q and Cao C: Gαi1 and Gαi3mediate
VEGF-induced VEGFR2 endocytosis, signaling and angiogenesis.
Theranostics. 8:4695–4709. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Larrue C, Saland E, Boutzen H, Vergez F,
David M, Joffre C, Hospital MA, Tamburini J, Delabesse E, Manenti
S, et al: Proteasome inhibitors induce FLT3-ITD degradation through
autophagy in AML cells. Blood. 127:882–892. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang Y, Deng M, Chen Q, Li Y, Guo X, Shi
P, He L, Xie S, Yu L, Zhang H and Xu B: Apatinib exerts anti-tumor
activity to non-Hodgkin lymphoma by inhibition of the Ras pathway.
Eur J Pharmacol. 843:145–153. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Qiu H, Li J, Liu Q, Tang M and Wang Y:
Apatinib, a novel tyrosine kinase inhibitor, suppresses tumor
growth in cervical cancer and synergizes with paclitaxel. Cell
Cycle. 17:1235–1244. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Weilbaecher KN, Guise TA and McCauley LK:
Cancer to bone: A fatal attraction. Nat Rev Cancer. 11:411–425.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Viprey VF, Gregory WM, Corrias MV,
Tchirkov A, Swerts K, Vicha A, Dallorso S, Brock P, Luksch R,
Valteau-Couanet D, et al: Neuroblastoma mRNAs predict outcome in
children with stage 4 neuroblastoma: A European HR-NBL1/SIOPEN
study. J Clin Oncol. 32:1074–1083. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
McTigue M, Murray BW, Chen JH, Deng YL,
Solowiej J and Kania RS: Molecular conformations, interactions, and
properties associated with drug efficiency and clinical performance
among VEGFR TK inhibitors. Proc Natl Acad Sci USA. 109:18281–18289.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rössler J, Monnet Y, Farace F, Opolon P,
Daudigeos-Dubus E, Bourredjem A, Vassal G and Geoerger B: The
selective VEGFR1-3 inhibitor axitinib (AG-013736) shows antitumor
activity in human neuroblastoma xenografts. Int J Cancer.
128:2748–2758. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kakodkar NC, Peddinti RR, Tian Y, Guerrero
LJ, Chlenski A, Yang Q, Salwen HR, Maitland ML and Cohn SL:
Sorafenib inhibits neuroblastoma cell proliferation and signaling,
blocks angiogenesis, and impairs tumor growth. Pediatr Blood
Cancer. 59:642–647. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nilsson MB, Zage PE, Zeng L, Xu L, Cascone
T, Wu HK, Saigal B, Zweidler-McKay PA and Heymach JV: Multiple
receptor tyrosine kinases regulate HIF-1alpha and HIF-2alpha in
normoxia and hypoxia in neuroblastoma: Implications for
antiangiogenic mechanisms of multikinase inhibitors. Oncogene.
29:2938–2949. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cheng X, Feng H, Wu H, Jin Z, Shen X,
Kuang J, Huo Z, Chen X, Gao H, Ye F, et al: Targeting autophagy
enhances apatinib-induced apoptosis via endoplasmic reticulum
stress for human colorectal cancer. Cancer Lett. 431:105–114. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mukhopadhyay S, Panda PK, Sinha N, Das DN
and Bhutia SK: Autophagy and apoptosis: Where do they meet?
Apoptosis. 19:555–566. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hydbring P, Malumbres M and Sicinski P:
Non-canonical functions of cell cycle cyclins and cyclin-dependent
kinases. Nat Rev Mol Cell Biol. 17:280–292. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Thorpe LM, Yuzugullu H and Zhao JJ: PI3K
in cancer: Divergent roles of isoforms, modes of activation and
therapeutic targeting. Nat Rev Cancer. 15:7–24. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wagner EF and Nebreda AR: Signal
integration by JNK and p38 MAPK pathways in cancer development. Nat
Rev Cancer. 9:537–549. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Simons M, Gordon E and Claesson-Welsh L:
Mechanisms and regulation of endothelial VEGF receptor signalling.
Nat Rev Mol Cell Biol. 17:611–625. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Levy JMM, Towers CG and Thorburn A:
Targeting autophagy in cancer. Nat Rev Cancer. 17:528–542. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hansen M, Rubinsztein DC and Walker DW:
Autophagy as a promoter of longevity: Insights from model
organisms. Nat Rev Mol Cell Biol. 19:579–593. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang G, Wang Z, Du Z and Zhang H: mTOR
regulates phase separation of PGL granules to modulate their
autophagic degradation. Cell. 174:1492–1506 e22. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
You L, Wang Z, Li H, Shou J, Jing Z, Xie
J, Sui X, Pan H and Han W: The role of STAT3 in autophagy.
Autophagy. 11:729–739. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ohsumi Y: Historical landmarks of
autophagy research. Cell Res. 24:9–23. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Klionsky DJ, Abdelmohsen K, Abe A, Abedin
MJ, Abeliovich H, Acevedo Arozena A, Adachi H, Adams CM, Adams PD,
Adeli K, et al: Guidelines for the use and interpretation of assays
for monitoring autophagy (3rd edition). Autophagy. 12:1–222. 2016.
View Article : Google Scholar : PubMed/NCBI
|