Introduction
Laryngeal cancer is a common primary malignant tumor
in the head and neck (1). The
incidence of laryngeal cancer is 5.7–7.6% of total body tumors
(2). It is estimated that there were
177,422 cases of laryngeal cancer and 94,711 mortalities worldwide
in 2012 (3). Currently, there is no
exact cause of laryngeal cancer, which may be caused by several
factors, including smoking, alcohol consumption, air pollution and
viral infection (4,5). Thus, the key to improving the prognosis
of laryngeal cancer is to determine the biological characteristics
of laryngeal cancer cells.
Transmembrane protein 158 (TMEM158), also known as
RIS-1 protein, is recognized for its increased expression when Ras
protein induces senescence of haploid fibroblasts infected with RAS
V12 lentivirus (6). The expression
and biological function of TMEM158 have been reported in different
types of cancer (7–9); however, its role in laryngeal cancer
remains unknown, thus, the present study aimed to investigate the
regulatory mechanism and biological function of TMEM158 in
laryngeal cancer.
MicroRNAs (miRNAs/miRs) are small non-coding RNA
molecules that play key roles in the regulation of gene expression
(10). Translation inhibition or
silencing of target mRNAs is induced following complementary
binding to the 3′-untranslated region (UTR) (11,12).
miR-548 is a large and poorly conserved primate-specific miRNA gene
family that consists of 68 members, and several members of this
family have been reported to be associated with cancer (13). For example, miR-548b-3p functions as
a tumor suppressor in lung cancer (14) and as an antioncogenic regulator in
breast cancer (15).
The purpose of the present study is to explore the
molecular mechanism of the development of laryngeal cancer and to
identify the biomarker for diagnosis and treatment of laryngeal
cancer. Reverse transcription-quantitative PCR (RT-qPCR) and the
analysis of The Cancer Genome Atlas (TCGA) database were used to
demonstrate overexpression of TMEM158 in laryngeal cancer. The
carcinogenic effect of TMEM158 and the anticancer effect of
miR-548ac was investigated through in vitro and in
vivo assays (MTT assay, colony formation assay, flow cytometry
assay, western blotting and tumor xenograft assay). Luciferase
reporter assay, western blotting and RT-qPCR were used to
demonstrate miR-548 directly targeted the 3′-untranslated region of
TMEM158 and inhibited TMEM158 expression. These results suggest
that miR-548ac functions as a crucial cancer suppressor in
laryngeal cancer, which induces apoptosis in laryngeal cancer cells
by suppressing TMEM158. The present findings enrich the
pathogenesis of laryngeal cancer and provide a new target and new
ideas for the treatment on laryngeal cancer.
Materials and methods
Cell lines and culture
Human LSC-1 cells and NP69 nasopharyngeal epithelial
cells were purchased from the Institute of Oncology, Chinese
Academy of Medical Sciences. Cells were maintained in DMEM medium
(Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10% fetal
bovine serum (cat. no. 10100147; Thermo Fisher Scientific, Inc.),
penicillin (100 IU/ml) and streptomycin (100 mg/ml) at 37°C in 5%
CO2. LSC-1 cells were transfected with 10 µM miR-548ac
agomir (LSC-1-miR-548ac) or 10 µM miR-548ac agomir negative control
(NC; LSC-1-NC; Guangzhou RiboBio Co., Ltd.) using
Lipofectamine® RNAiMAX reagent (cat. no. 13778030;
Thermo Fisher Scientific, Inc.). The short hairpin (sh)RNA
targeting positions 1520–1540 (5′-AAATGACCAAATCCTGTGTAT-3′; Sangon
Biotech Co. Ltd.) of human TMEM158 mRNA was cloned into a
lentiviral vector (PLKO.1). A non-specific scrambled shRNA sequence
(5′-CCTAAGGTTAAGTCGCCCTCG-3′; Sangon Biotech Co. Ltd.) was used as
the NC. 1 µg (50 pmol) shRNA was added into Opti-MEM (cat. no.
31985062; Thermo Fisher Scientific, Inc.) to prepare RNA diluent,
and the final volume was 25 µl. LSC-1 cells were subsequently
transfected with lentiviral packaging vectors (LSC-1-NC or
LSC-1-shTMEM158) using Lipofectamine® 3000 reagent (cat.
no. L3000001; Thermo Fisher Scientific, Inc.), according to the
manufacturer's instructions. Subsequent experiments began at 24 h
after transfection of LSC-1 cells with shRNA. The LSC-1 cells
stably transfected with NC, miR-548ac and shTMEM158 were
constructed respectively.
Tumor xenograft model
A total of 30 4-week-old BALB/cA female nude mice
(14–18 g) were purchased from the Beijing Vital River Laboratory
Animal Technologies Co. Ltd. The present study was approved by the
Ethics Committee of Tianjin Union Medical Center (Tianjin, China;
approval no. 2018-269). All nude mice were kept at 28°C and SPF
level with light/dark cycle. Mice received pathogen-free food and
water. All food and water were sterilized and then sent into the
animal isolator through a sterile entrance chamber to ensure that
the nude mice were in a sterile living state. All animal
experiments were performed in accordance with the National
Institutes of Health Guide for the Care and Use of Laboratory
Animals (NIH Publications no. 8023, revised in 1978).
LSC-1 cells (2×106 cells/mouse) were
inoculated into the back of the nude mice following light
anesthesia with 350 mg/kg chloral hydrate. The mice were randomly
divided into four groups (6 mice/group), as follows: i) LSC-1 cells
with a blank vector; ii) LSC-1 cells with shTMEM158; ii) LSC-1
cells with miR-548ac NC and iv) LSC-1 cells with miR-548ac agomir.
The experimental procedures lasted for 3 weeks. The health and
behavior of all animals were monitored every day and the
individuals with poor health were terminated in time. The nude mice
were free from stress, discomfort, pain and injury. All mice were
sacrificed via intraperitoneal injection with 200 mg/kg
pentobarbital sodium. Mortality was confirmed by checking for a
heartbeat and examining the pupil dilation of the nude mice. The
maximum diameter of the tumor was 0.87 cm. The tumors were
subsequently removed using resection, photographed using a camera
(model, D3600; Nikon Corporation) and weighed. Tumor volume was
calculated using the following formula: V=(length ×
width2)/2.
MTT assay
The cell suspensions (LSC-1-NC, LSC-1- miR-548ac;
LSC-1-NC or LSC-1-shTMEM158) were added into 96-well plates at a
density of 6,000 cells/well. After 24 h of cell attachment, the
culture medium was discarded. Cells were subsequently incubated
with serum-free DMEM (100 µl) and 10 µl MTT (5 mg/ml; cat. no.
ST316; Beyotime Institute of Biotechnology, Inc.) for 4 h at 37°C.
Following the MTT incubation, the purple formazan crystals were
dissolved using 100 µl dimethyl sulfoxide and shaking vigorously
for 10 min using a vortex oscillator (model, MixMax; AIBENSEN, Co.
Ltd.). Cell viability was subsequently analyzed at a wavelength of
570 nm. All experiments were performed in triplicate and three
biological repeats were conducted.
Colony formation assay
The cell suspensions (LSC-1-NC, LSC-1-miR-548ac;
LSC-1-NC or LSC-1-shTMEM158) were added into 6-well plates at a
density of 300 cells/well. Cells were maintained in DMEM medium
supplemented with 10% fetal bovine serum (cat. no. 10100147; Thermo
Fisher Scientific, Inc.) for 7–10 days at 37°C. After ~70 cells
appeared in each colony, each pore was fixed with 1 ml methanol for
15 min at room temperature. Cell colonies were subsequently stained
with 0.5% crystal violet for 10 min at room temperature, and excess
stain was discarded using ultrapure water. Cell colonies were
observed and recorded using a camera (model, D3600; Nikon
Corporation).
Annexin V-propidium iodide (PI)
assay
Cells (LSC-1-NC, LSC-1-miR-548ac; LSC-1-NC or
LSC-1-shTMEM158) were digested with trypsin-EDTA solution (0.05%
trypsin and 0.02% EDTA; cat. no. C0202; Beyotime Institute of
Biotechnology, Inc.) and collected by centrifugation at 161 × g and
room temperature. Cells were washed three times with sterile PBS
and subsequently resuspended in 500 µl Annexin V-PI binding
solution. PI (10 µl) and annexin V-fluorescein isothiocyanate (5
µl) (cat. no. C1062S; Beyotime Institute of Biotechnology, Inc.)
were added to the cell suspensions of each group and manually
gently mixed. The whole reaction was 515 µl at room temperature and
dark for 30 min. Apoptotic cells were subsequently detected using a
flow cytometer.
Flow cytometric analysis of cell cycle
distribution
Cells (LSC-1-NC, LSC-1-miR-548ac; LSC-1-NC or
LSC-1-shTMEM158) at 1×106 cells/ml were collected
following digestion with trypsin-EDTA solution (cat. no. C0202;
Beyotime, Inc.), washed twice with PBS, and resuspended and fixed
with 70% ethanol overnight at −20°C. The cell suspension was
re-washed twice with PBS and stained with 400 µl PI at 4°C for 30
min in the dark. Cell cycle distribution was subsequently detected
via flow cytometric analysis.
Western blotting
Total protein from cells (LSC-1-NC, LSC-1-miR-548ac;
LSC-1-NC or LSC-1-shTMEM158) was extracted using RIPA lysis buffer
(cat. no. R0010; Beijing Solarbio Science & Technology Co.,
Ltd.). Detection of protein concentration was used BCA assay. Equal
amounts (15 µg) of protein were separated via SDS-PAGE (12%
separating gel and 5% concentrating gel) and subsequently
transferred onto polyvinylidene difluoride (PVDF) membranes. The
PVDF membrane was blocked in 5% skimmed milk at room temperature
for 2 h. The membranes were incubated with primary antibodies
against: Bax (1:1,000; cat. no. ab32503), B-cell lymphoma-2 (Bcl-2;
1:1,000; cat. no. ab8227), TMEM158 (1:1,000; cat. no. ab98335) and
β-actin (1:2,000; cat. no. ab98335) (all purchased from Abcam) at
room temperature for 2 h. Following the primary incubation,
membranes were incubated with secondary antibodies (HRP goat
anti-Rabbit; 1:5,000; cat. no. A0208; Beyotime Institute of
Biotechnology, Inc.) at room temperature for 1 h. Protein bands
were visualized using enhanced chemiluminescence buffer and
detected using the multifunctional gel imaging system (model, Gel
Doc XR+; Bio-Rad Laboratories, Inc.). Protein expression was
calculated via the gray value.
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted from cells (LSC-1-NC,
LSC-1-miR-548ac; LSC-1-NC or LSC-1-shTMEM158) using
TRIzol® reagent (cat. no. 5301100; SIMGEN) according to
the manufacturer's instructions, and quantified using a NanoDrop
2000 spectrophotometer (Thermo Fisher Scientific, Inc.). Total RNA
was reverse transcribed into cDNA using the TransScript RT Reagent
kit (cat. no. AH411-02; TransGen Biotech Co., Ltd.). The reverse
transcription reaction conditions were 25°C, 10 min (miRNA skip);
42°C, 30 min; 85°C, 5 min. qPCR was subsequently performed using
the 2×SYBR Green qPCR mix (cat. no. 153301; Beijing BLKW
Biotechnology Co. Ltd.) and a ABI 7500 system (Thermo Fisher
Scientific, Inc.). The qPCR reaction conditions were 1 cycle at
94°C for 2 min; 40 cycles (94°C for 15 sec; 55°C for 15 sec; and
72°C for 20 sec). The following primer sequences were used for
qPCR: TMEM158 forward, 5′-TGCCCAACGGCATGGAACA-3′ and reverse,
5′-AGGAGCGGAGCGGGTCACTT-3′; GAPDH forward,
5′-ATGACATCAAGAAGGTGGTGAAGCAGG-3′ and reverse,
GCGTCAAAGGTGGAGGAGTGGGT; miR-548ac forward,
5′-CAAAAACCGGCAATTACTTTTG-3′ and reverse, 5′-CTCAACTGGTGTCGTGGA-3′;
and U6 forward, 5′-CTCGCTTCGGCAGCACA-3′ andreverse,
5′-CTCAACTGGTGTCGTGGA-3′. Relative expression levels were
calculated using the 2−ΔΔCq method (16) and normalized to the internal
reference genes GAPDH and U6, respectively.
Dual-luciferase reporter assay
The wild (WT)/mutant (MUT) 3′-UTR fragments of
TMEME158 were amplified via PCR using 2X PCR Master mix (cat. no.
D7228; Beyotime Institute of Biotechnology, Inc.). The PCR reaction
conditions were: 1 cycle at 94°C for 5 min; 35 cycles (94°C for 30
sec; 60°C for 1 min; 72°C at 1 min); and 1 cycle (72°Cfor 10 min).
Th fragments were cloned into the Dual-Luciferase miRNA targeted
expression vector (Promega Corporation) to synthesize
WT/MUT-TMEM158-3′-UTR. LSC-1 cells with 70% confluence were
transfected with WT-TMEM158-3′-UTR or MUT-TMEM158-3′-UTR and
miR-548ac agomirs or miR-548ac agomirs NC/miR-548ac antagonists or
miR-548ac antagonists NC(Guangzhou RiboBio Co., Ltd.)
usingLipofectamine®3000 reagent (cat. no. L3000008;
Thermo Fisher Scientific, Inc.). Following incubation at 37°Cfor 48
h, Luciferase activity was measured using a multimode microplate
reader (BioTek Instruments, Inc.) and normalized with Renilla
luciferase activity.
Bioinformatics analysis
The mRNA and miRNA sequencing data and the
corresponding clinical information of the patients with laryngeal
cancer were downloaded from the TCGA data portal (http://cancergenome.nih.gov). The data were screened
and laryngeal cancer samples and non-tumor samples, which had
detailed clinical information (laryngeal cancer, 116 cases; normal,
11 cases), were selected. The clinical features of the patients are
shown in Table I. Patients with head and neck cancer were divided
into four stages, according to tumor-node-metastasis (TNM)
(17) and differences in TMEM158
expression at different Tumor-Node-Metastasis stages were analyzed.
RNAInter software (http://www.rna-society.org/raid/; Department of
Bioinformatics, Southern Medical University) was used to predict
the target association between miR-548ac and TMEM158. Univariate
Cox proportional hazards regression analysis was performed to
determine the association between TMEM158 expression and overall
survival rate. Kaplan-Meier survival analysis and log-rank test
were used to assess the association between TMEM158 expression and
overall survival time. A total of 38 cases of TMEM158 with high or
low expression were selected.
Statistical analysis
Statistical analyses were performed using SPSS 18.0
software (SPSS, Inc.). All experiments were performed in triplicate
and data are presented as the mean ± standard deviation. Unpaired
Student's two-tailed t-test was used to compare differences between
two independent samples, whilst one-way analysis of variance and
Bonferroni's correction were used to compare differences between
multiple groups (Figs. 1B, 2F, 3G,
4D and 5B
and C). P<0.05 was considered to indicate a statistically
significant difference.
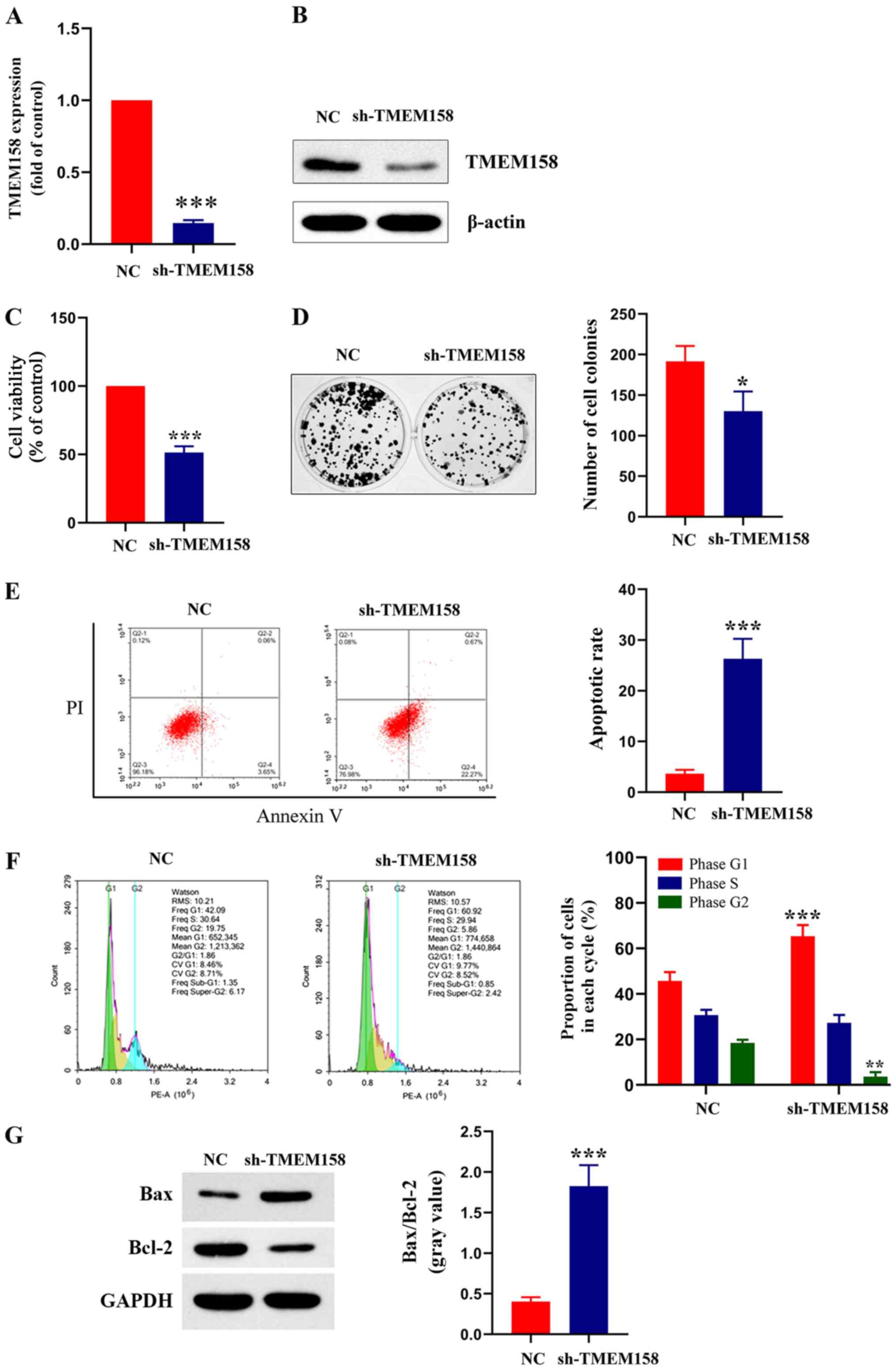 | Figure 2.TMEM158 affects viability, formation
of colonies, cell cycle distribution and apoptosis of laryngeal
cancer cells. (A) Reverse transcription-quantitative PCR and (B)
Western blot analyses were performed to assess mRNA and protein
expression of TMEM158 in LSC-1 cells transfected with sh-TMEM158 or
NC, respectively. ***P<0.001, shTMEM158 vs. NC. (C) The MTT
assay was performed to assess cell viability in LSC-1 cells
transfected with sh-TMEM158 or NC. ***P<0.001, shTMEM158 vs. NC.
(D) Effect of TMEM158 on colony formation of laryngeal cancer
cells. Representative photos (left) and relative quantification
analysis of colonies (right). *P<0.05, shTMEM158 vs. NC. (E)
Flow cytometric analysis of apoptosis. ***P<0.001, shTMEM158 vs.
NC. (F) Flow cytometric analysis of cell cycle distribution.
Histograms depict the proportion of LSC-1 cells in the G1, S and G2
phases. ***P<0.001, shTMEM158 (G1) vs. NC
(G1); **P<0.01, shTMEM158 (G2) vs. NC
(G2). (G) Western blot analysis was performed to assess
protein expression levels of Bax and Bcl-2. Histograms depict the
gray value of Bax/Bcl-2. ***P<0.001, shTMEM158 vs. NC. TMEM158,
transmembrane protein 158; LSC-1, laryngeal squamous cell
carcinoma; sh, short hairpin; NC, negative control; Bcl-2, B-cell
lymphoma 2; PI, propidium iodide. |
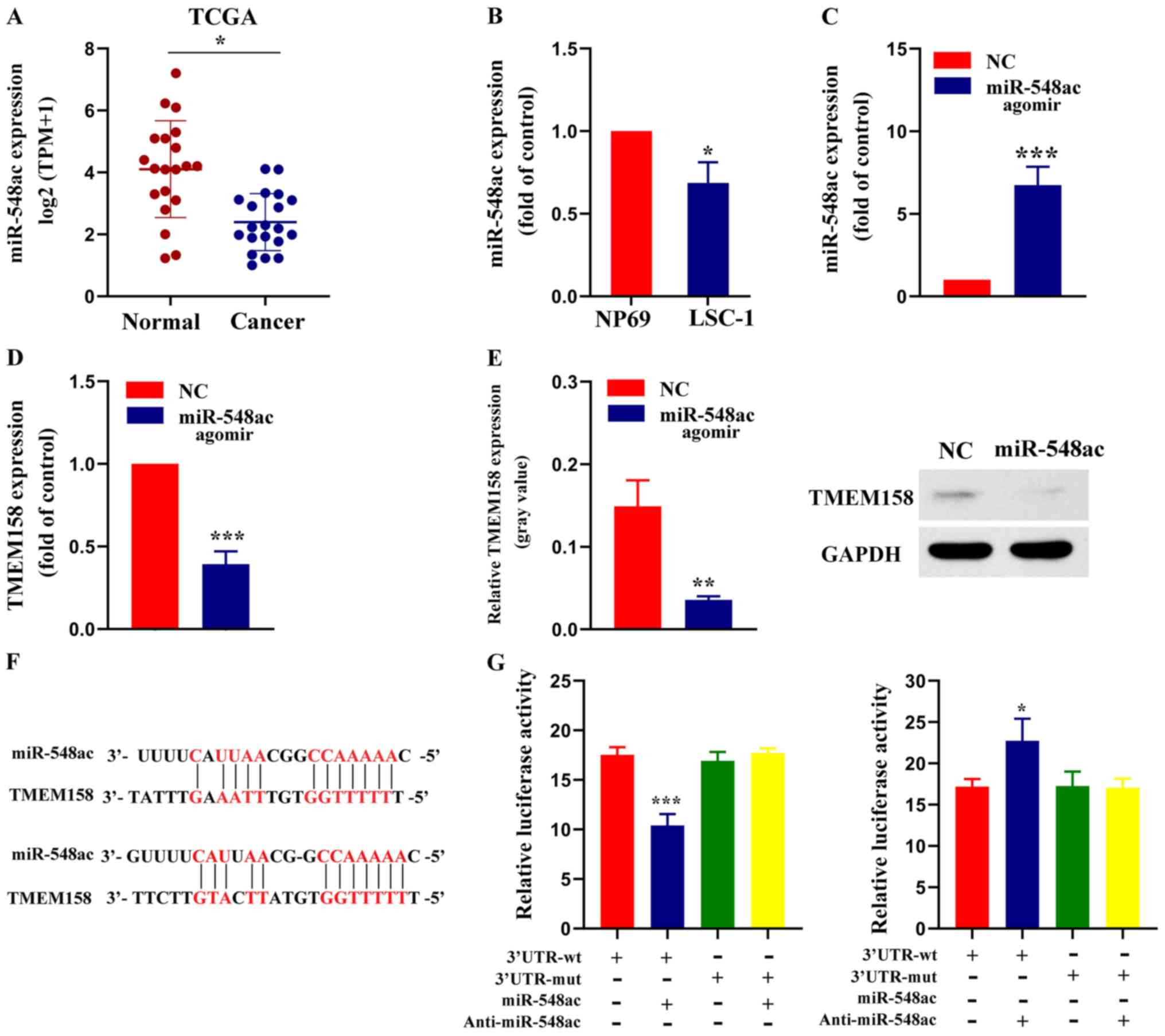 | Figure 3.miR-548ac suppresses TMEM158
expression by directly binding to its 3′-UTR. (A) miR-548ac
expression was assessed in laryngeal carcinoma tissues and adjacent
normal tissues using miRNA sequencing data from TCGA database.
*P<0.05 normal group vs. tumor group. (B) RT-qPCR analysis was
performed to assess miR-548ac expression in NP69 and LSC-1 cells.
*P<0.05, LSC-1 cells vs. NP69 cells. (C) RT-qPCR assay was
performed to assessmiR-548ac expression in LSC-1 cells transfected
with miR-548ac agomir or NC. ***P<0.001, miR-548ac agomirs vs.
NC. (D) RT-qPCR analysis was performed to assess TMEM158 expression
in LSC-1 cells transfected with miR-548ac agomir or NC.
***P<0.001, miR-548ac agomirs vs. NC. (E) Western blot analysis
was performed to assess TMEM158 protein expression in LSC-1 cells
transfected with miR-548ac agomir or NC. Histograms depict the gray
value of TMEM158/GAPDH. **P<0.01, miR-548ac agomirs vs. NC. (F)
Sequence alignment of miR-548ac and its predicted binding sites
(red) in TMEM158 3′-UTR. (G) Luciferase reporter assay.
***P<0.001, 3′UTR-WT (+miR-548ac) vs. 3′UTR-WT; *P<0.05,
3′UTR-WT (+Anti-miR-548ac) vs. 3′UTR-WT. miR, microRNA; TMEM158,
transmembrane protein 158; UTR, untranslated region; TCGA, The
Cancer Genome Atlas; RT-qPCR, reverse transcription-quantitative
PCR; LSC-1, laryngeal squamous cell carcinoma; NC, negative
control. |
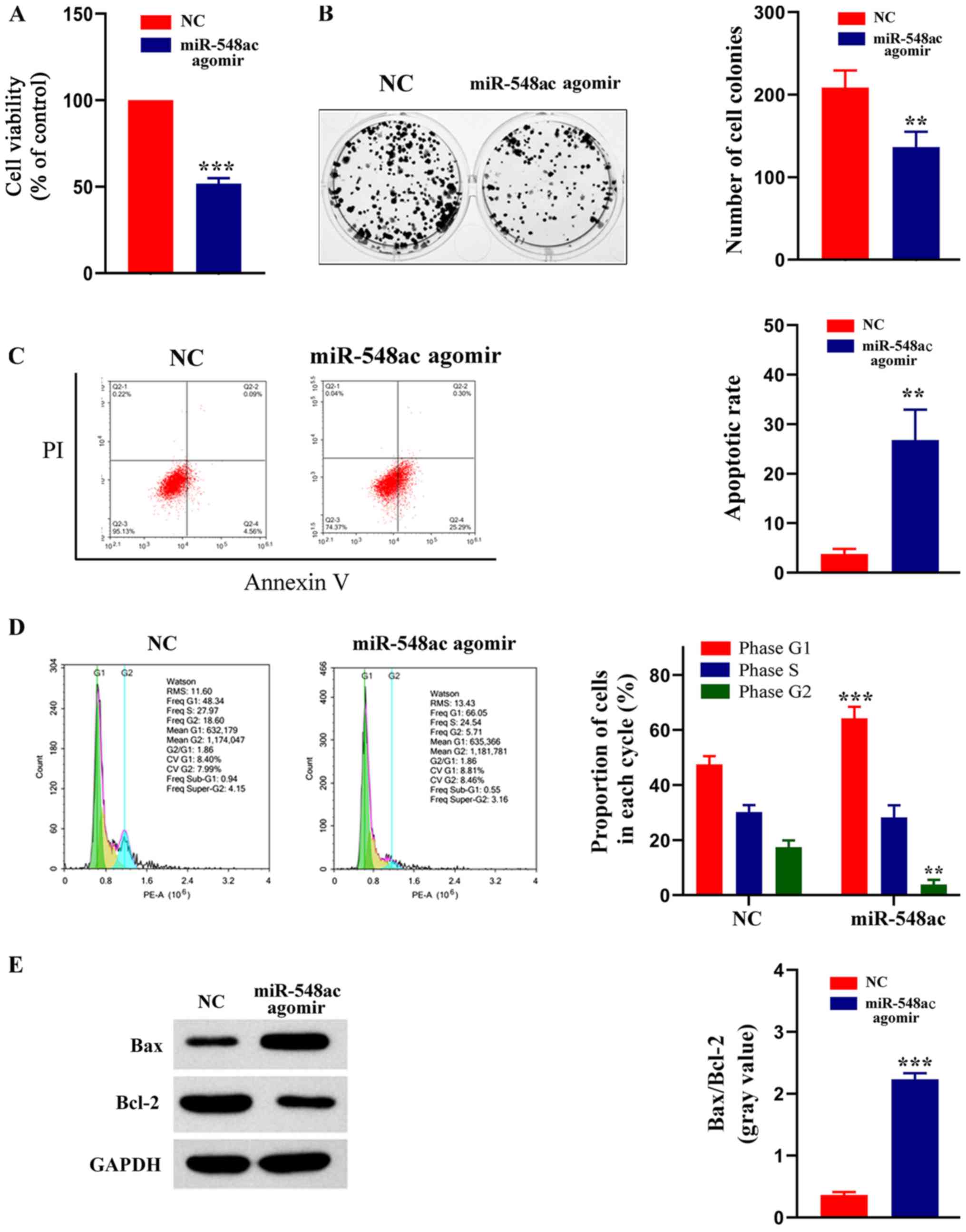 | Figure 4.miR-548ac affects viability, formation
of colonies, cell cycle distribution and apoptosis of laryngeal
cancer cells. (A) MTT assay was performed to assess cell viability
in LSC-1 cells transfected with miR-548ac antagomir or NC.
***P<0.001, miR-548ac agomir vs. NC. (B) Effect of miR-548ac on
colony formation of laryngeal cancer cells. Representative
micrographs (left) and relative quantification analysis of colonies
(right). **P<0.01, miR-548ac agomir vs. NC. (C) Flow cytometric
analysis of apoptosis. **P<0.01, miR-548ac agomir vs. NC. (D)
Flow cytometric analysis of cell cycle distribution. Histograms
depict the proportion of LSC-1 cells in the G1, S and G2 phases.
***P<0.001, miR-548ac agomir (G1) vs. NC
(G1); **P<0.01, miR-548ac agomir (G2) vs.
NC (G2). (E) Western blot analysis was performed to
assess protein expression levels of Bax and Bcl-2. Histograms
depict the gray value of Bax/Bcl-2. ***P<0.001, miR-548ac agomir
vs. NC. miR, microRNA; LSC-1, laryngeal squamous cell carcinoma;
NC, negative control; Bcl-2, B-cell lymphoma 2; PI, propidium
iodide. |
Results
TMEM158 is upregulated in laryngeal
cancer tissues and cell lines
The Cancer Genome Atlas (TCGA) database was used to
compare mRNA sequencing data between laryngeal carcinoma tissues
and adjacent normal tissues. The results demonstrated that TMEM158
expression was significantly higher in laryngeal carcinoma tissues
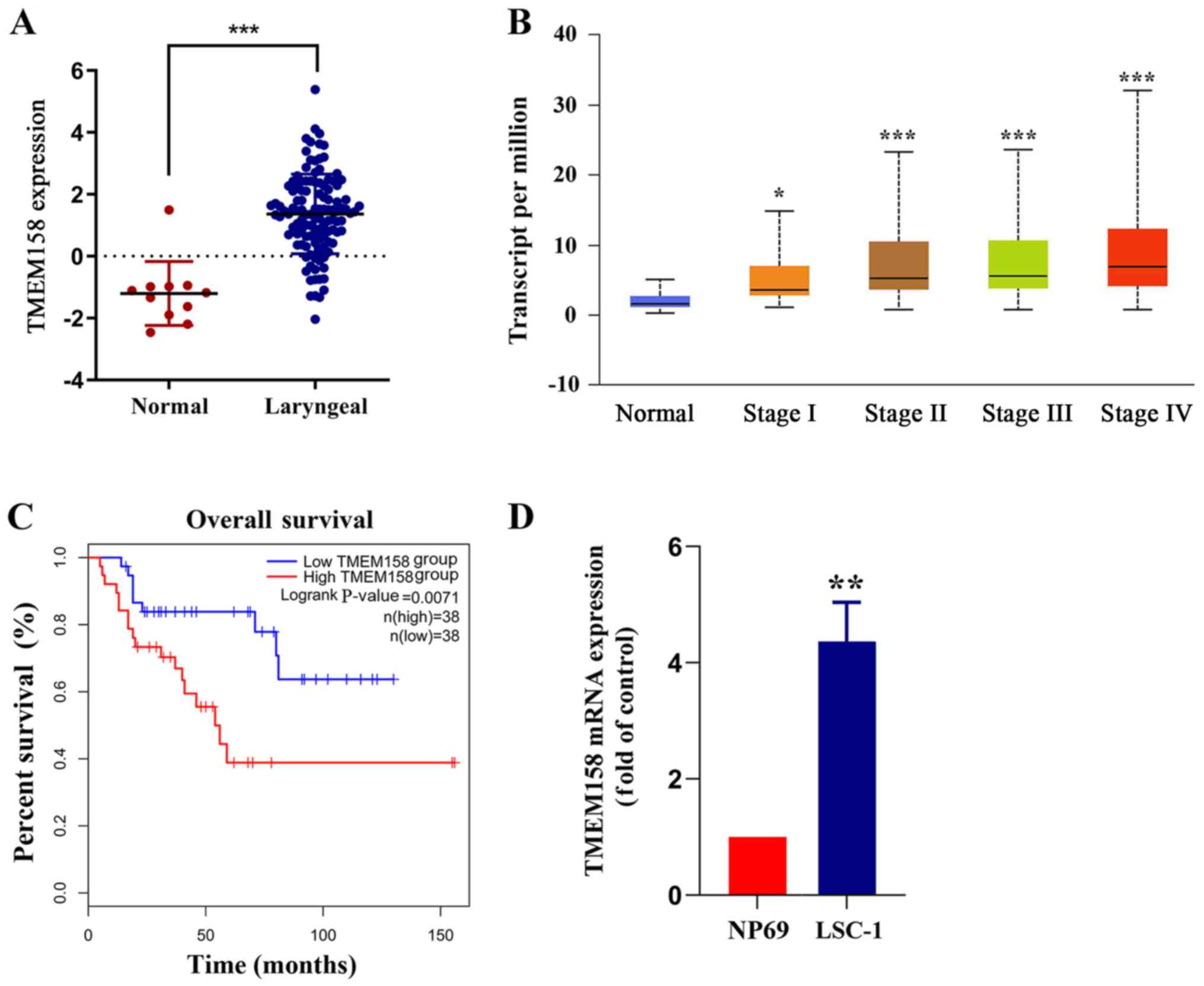
compared with adjacent normal tissues (Fig. 1A). TMEM158 expression according to
different tumor-node-metastasis (TNM) stages was also assessed. The
results demonstrated that TMEM158 expression increased in
accordance with the increase of the TNM stage (Fig. 1B). The prognostic value of TMEM158 in
laryngeal carcinoma was determined via overall survival analysis.
The results demonstrated that patients with low TMEM158 expression
had a significantly longer survival time than that of patients with
high TMEM158 expression (Fig. 1C).
TMEM158 mRNA expression was further assessed in NP69 and LSC-1
cells. The results demonstrated that TMEM158mRNA expression was
4.35±0.56 times greater in LSC-1 cells compared with NP69 cells
(P=0.0052; Fig. 1D). Taken together,
these results suggest that upregulated TMEM158 expression may play
a critical role in the development and progression of laryngeal
cancer.
Effect of TMEM158 on the viability of
laryngeal cancer cells
Normal LSC-1 cells were used as the control group,
while cells transfected with sh-TMEM158 were used as the
experimental group. RT-qPCR analysis demonstrated that TMEM158 mRNA
expression was significantly lower in LSC-1 cells transfected with
sh-TMEM158 compared with LSC-1 cells transfected with NC
(14.67±2.07% of the control; P<0.001; Fig. 2A). The results of western blot
analysis were consistent with these findings (Fig. 2B). Similarly, the results of the MTT
assay demonstrated that cell viability was significantly lower in
LSC-1 cells transfected with sh-TMEM158 compared with the control
group (51.49±4.51% of the control; P<0.0001; Fig. 2C). The results of the colony
formation assay demonstrated that the number of cell colonies was
significantly lower in the experimental group compared with the
control group (67.62±4.72% of the control; P=0.025; Fig. 2D). The results of the Annexin V-PI
assay indicated that the apoptotic rate was significantly higher in
the experimental group compared with the control group (7.3±1.33 of
the control; P=0.00062; Fig. 2E).
The cell cycle distribution assay demonstrated a higher proportion
of cells transfected with sh-TMEM158 in the G1 phase
compared with the control group (Fig.
2F). The protein expression levels of Bax and Bcl-2 were
determined via western blot analysis, and the gray value was used
for relative quantification. The results demonstrated that Bax and
Bcl-2 expression levels were significantly higher in the
experimental group compared with the control group (4.53±0.03 of
the control; P=0.00072; Fig. 2G).
Collectively, these results indicate that silencing TMEM158
significantly inhibits the viability of laryngeal cancer cells.
miR-548ac is downregulated in
laryngeal cancer tissues and cell lines
RNAInter software analysis demonstrated that
miR-548ac has a high probability of binding to TMEM158. TCGA
database was used to compare miRNA sequencing data between
laryngeal carcinoma tissues and adjacent normal tissues. The
results demonstrated that miR-548ac expression was significantly
lower in laryngeal carcinoma tissues compared with adjacent normal
tissues (Fig. 3A). miR-548ac
expression was further assessed inNP69 and LSC-1 cells. The results
demonstrated that miR-548ac expression was significantly lower in
LSC-1 cells compared with NP69 cells (68.67±10.21% of NP69 cells;
P=0.027; Fig. 3B). Taken together,
these results suggest that downregulation of miR-548ac may play a
critical role in the progression of laryngeal cancer.
Association between miR-548ac and
TMEM158
miR-548ac agomir (experimental group) or miR-548ac
agomir NC (control group) were transfected into LSC-1 cells,
respectively. RT-qPCR analysis demonstrated that miR-548ac
expression was significantly higher in the experimental group
compared with the control group (7.76±1.1-fold of the control;
P<0.001; Fig. 3C). Furthermore,
TMEM158 mRNA expression was significantly lower in the experimental
group compared with the control group (39.33±6.34% of the control;
P=0.00071; Fig. 3D). The results of
western blot analysis were consistent with these findings (Fig. 3E). The results of the dual-luciferase
reporter assay demonstrated that luciferase activity decreased in
LSC-1 cells transfected with WT-TMEM158-3′-UTR and miR-548ac agomir
compared with WT-TMEM158-3′-UTR alone (59.58±6.78% of
WT-TMEM158-3′-UTR alone; P<0.001). However, the luciferase
activity remained unchanged whenLSC-1 cells were transfected with
MUT-TMEM158-3′-UTR and miR-548ac agomir. Luciferase activity
increased whenLSC-1 cells were transfected withWT-TMEM158-3′-UTR
and anti-miR-548ac agomir compared with WT-TMEM158-3′-UTR alone
(1.32±0.07 of WT-TMEM158-3′-UTR alone; P=0.03). However, the
luciferase activity remained unchanged whenLSC-1 cells were
transfected with MUT-TMEM158-3′-UTR and anti-miR-548ac agomir
compared with MUT-TMEM158-3′-UTR alone (Fig. 3G). Collectively, these results
indicate that TMEM158 is a direct target gene of miR-548ac.
Effect of miR-548ac on the viability
of laryngeal cancer cells
Normal LSC-1 cells were used as the control group,
while cells transfected with miR-548ac agomir were used as the
experimental group. The results of the MTT assay demonstrated that
cell viability significantly decreased in the experimental group
compared with the control group (51.86±3.11% of the control;
P<0.0001; Fig. 4A). The results
of the colony formation assay demonstrated that the number of cell
colonies was significantly lower in the experimental group compared
with the control group (65.44±4.34% of the control; P=0.0094;
Fig. 4B). The results of the Annexin
V-PI assay demonstrated that the apoptotic rate was significantly
higher in the experimental compared with the control group
(7.27±1.22-fold of the control; P=0.003; Fig. 4C). The cell cycle distribution assay
demonstrated a higher proportion of cells transfected with
miR-548ac agomir in the G1 phase compared with the
control group (Fig. 4D). The protein
expression levels of Bax and Bcl-2 were detected via western blot
analysis, and the gray value was used for relative quantification.
The western blots showed that Bcl-2 protein expression was higher
and Bax protein expression was lower in NC group compared with the
experimental group. Gray value ratio of Bax and Bcl-2 was
significantly higher in the experimental group compared with the
control group (6.19±0.46 fold of the control; P=0.00045; Fig. 4E). Collectively, these results
indicate that overexpressing miR-548ac significantly inhibits the
viability of laryngeal cancer cells. Thus, it is speculated that
miR-548ac plays a cancer suppressive role in laryngeal cancer.
Effect of TMEM158 and miR-548ac on
tumor growth in vivo
A xenograft model was used to determine the effects
of TMEM158 and miR-548ac on tumor growth in vivo. As
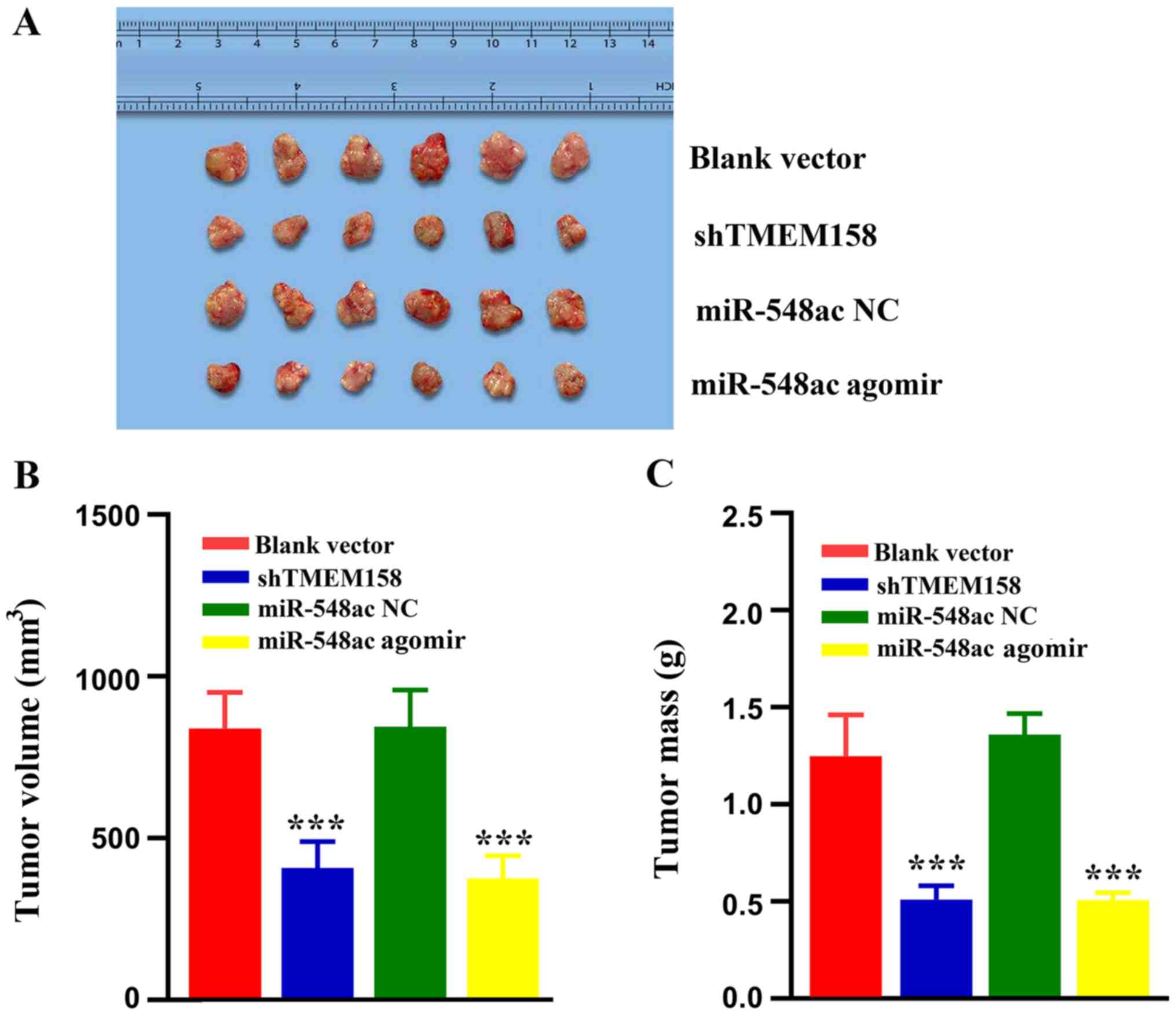
presented in Fig. 5A, the tumors
were smaller in the shTMEM158 and miR-548ac agomir groups compared
with the corresponding control groups. The tumor volume in the
shTMEM158 group was 49.05±9.78% that of the blank vector group
(P<0.001), while the tumor volume in the miR-548ac group was
45.36±10.54% that of the miR-548ac NC group (P<0.001) (Fig. 5B). The tumor mass in the shTMEM158
group was 42.21±11.47% that of the blank vector group
(P<0.0001), while the tumor mass in the miR-548ac group was
37.49±4.43% that of the miR-548ac NC group (P<0.001) (Fig. 5C).
Discussion
miRNAs function in repressing translation of target
mRNAs or degrading it, thus negatively regulating the expression of
target genes following transcription (18). The regulatory mechanism of miRNAs to
their target genes has been extensively studied and demonstrated to
be involved in biological events, such as cell proliferation,
apoptosis, differentiation, exercise and angiogenesis (19,20). It
has been reported that the miR-548 family can inhibit the growth of
several tumors. For example, Shi et al (15) reported that mir-548-3p inhibits the
proliferation of breast cancer cells by regulating ECHS1
expression, suggesting its potential as a therapeutic target for
breast cancer. Furthermore, Wang et al (14) demonstrated that mir-548-3p can
effectively inhibit the proliferation of lung cancer cells by
inhibiting the PI3K/Akt signaling pathway, indicating that
miR-548b-3p has the potential to be developed as an antitumor
target of lung cancer. The results of the present study
demonstrated that LSC-1 cell viability significantly decreased
following overexpression of miR-548ac, whereby some cells exhibited
early apoptosis accompanied by activation of the apoptosis factor,
Bax, and further cell cycle arrest at the G1 phase.
Thus, it was speculated that overexpression of miR-548ac in
laryngeal cancer cells significantly inhibits cell proliferation
and induces apoptosis. Collectively, the results of the present
study suggest that miR-548ac may function as a tumor suppressor in
laryngeal cancer.
The expression and molecular mechanism of TMEM158
have been reported in different types of tumor. For example,
Iglesias et al (21)
suggested that TMEM158 acts as an oncogene in the development of
colon cancer. In addition, Mohamed et al (8) demonstrated that TMEM158 interference
significantly decreases the toxic effect exerted by cisplatin on
NSCLC cells, suggesting that TMEM158 is associated with the
sensitivity of chemotherapy drugs. Zirn et al (22) reported that TMEM158 is highly
expressed in CTNNB1 mutant nephroblastoma, which may be associated
with the Ras and Wnt signaling pathways. Furthermore, Liu et
al (23) demonstrated that
TMEM158 is a key regulatory factor of colorectal carcinogenesis and
drug resistance, and thus may be a promising target for colorectal
cancer treatment. The results of the present study demonstrated
that TMEM158 was highly expressed in laryngeal carcinoma tissues
from the TCGA database, which was verified via cell line
experiments. The results demonstrated that LSC-1 cell viability
significantly decreased following TMEM158 silencing, which induced
early apoptosis, accompanied by activation of the apoptotic factors
(Bax). Simultaneously, TMEM158 silencing resulted in cell arrest in
the G1 phase. Collectively, these results suggest that
TMEM158 may act as an oncogene, affecting the viability of
laryngeal cancer cells.
miRNAs regulate the expression of ~30% of human
protein-coding genes. Each miRNA can regulate hundreds of mRNA
genes, and the same target gene can be regulated by multiple
miRNAs, to regulate the expression of thousands of proteins
directly or indirectly (24). miRNAs
regulate gene expression by specifically pairing with the 3′-UTR
region of their target gene, resulting in the degradation of the
target mRNA or post transcriptional silencing, thus inhibiting
protein synthesis (25,26). The results of the present study
demonstrated that TMEM158 mRNA and protein expression levels
significantly decreased following overexpression of miR-548ac in
LSC-1 cells. The results of the dual-luciferase reporter assay
confirmed that miR-548ac directly binds to the 3′-UTR of TMEM158,
thus inhibiting TMEM158 expression. Consistent with previous
findings, the results of the present study demonstrated that
miR-548ac induces apoptosis in LSC-1 cells by targeting
TMEM158.
The present results suggest that miR-548ac functions
as a crucial cancer suppressor in laryngeal cancer, which induces
apoptosis in laryngeal cancer cells by suppressing TMEM158. The
present findings enrich the pathogenesis of laryngeal cancer and
provide a new target and new insights for the treatment of
laryngeal cancer. The limitation of this study is that there is no
biological detection of TMEM158 and miR-548ac in clinical samples.
At the same time, there is no detection on the influence of the
follow-up signal pathway. These limitations will be addressed in
future studies.
Acknowledgements
The authors would like to thank The Cancer Genome
Atlas (TCGA) project for providing the data.
Funding
Support for the study was provided by the Tianjin
Natural Science Foundation (No 20170842).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
FS designed the experiment and wrote the manuscript;
YY contributed to animal and cell experiments; JL performed the
statistical and bioinformatics analysis. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Tianjin Union Medical Center (Tianjin, China; approval
no. 2018-269,). All animal experiments were performed in accordance
with the National Institutes of Health Guide for the Care and Use
of Laboratory Animals (NIH Publications no. 8023, revised in
1978).
Patients consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Beibei Y, Rong Y, Yunfei Y and Wenchao Z:
Research progress regarding surgical margins, molecular margins,
and prognosis of laryngeal carcinoma. Ear Nose Throat J
145561320903146. 2020.(Epubahead of print). View Article : Google Scholar
|
|
2
|
Olaleye O, Siddiq S, Ekrikpo U and Kazi R:
Regional differences in incidence and mortality trends in cancers
of the larynx, thyroid, oral cavity and pharynx in England and
Scotland: 1975–2002. Open Epidemiol J. 3:70–78. 2013. View Article : Google Scholar
|
|
3
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Steuer CE, El-Deiry M, Parks JR, Higgins
KA and Saba NF: An update on larynx cancer. CA Cancer J Clin.
67:31–50. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Menicagli R, Bolla G, Menicagli L and
Esseridou A: The possible role of diabetes in the etiology of
laryngeal cancer. Gulf J Oncolog. 1:44–51. 2017.PubMed/NCBI
|
|
6
|
Barradas M, Gonos ES, Zebedee Z, Kolettas
E, Petropoulou C, Delgado MD, Leon J, Hara E and Serrano M:
Identification of a candidate tumor-suppressor gene specifically
activated during Ras-induced senescence. Exp Cell Res. 273:127–137.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cheng Z, Guo J, Chen L, Luo N, Yang W and
Qu X: Overexpression of TMEM158 contributes to ovarian
carcinogenesis. J Exp Clin Cancer Res. 34:752015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mohammed Ael S, Eguchi H, Wada S, Koyama
N, Shimizu M, Otani K, Ohtaki M, Tanimoto K, Hiyama K, Gaber MS and
Nishiyama M: TMEM158 and FBLP1 as novel marker genes of cisplatin
sensitivity in non-small cell lung cancer cells. Exp Lung Res.
38:463–474. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fu Y, Yao N, Ding D, Zhang X, Liu H, Ma L,
Shi W, Zhu C and Tang L: TMEM158 promotes pancreatic cancer
aggressiveness by activation of TGFβ1 and PI3K/AKT signaling
pathway. J Cell Physiol. 235:2761–2775. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hummel R, Maurer J and Haier J: MicroRNAs
in brain tumors: A new diagnostic and therapeutic perspective? Mol
Neurobiol. 44:223–234. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rouleau S, Glouzon JS, Brumwell A,
Bisaillon M and Perreault JP: 3′ UTR G-quadruplexes regulate miRNA
binding. RNA. 23:1172–1179. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xiao X, Cao Y and Chen H: Profiling and
characterization of microRNAs responding to sodium butyrate
treatment in A549 cells. J Cell Biochem. 119:3563–3573. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liang T, Guo L and Liu C: Genome-wide
analysis of mir-548 gene family reveals evolutionary and functional
implications. J Biomed Biotechnol. 2012:6795632012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang Z, Wu X, Hou X, Zhao W, Yang C, Wan W
and Chen L: MiR-548b-3p functions as a tumor suppressor in lung
cancer. Lasers Med Sci. 35:833–839. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shi Y, Qiu M, Wu Y and Hai L: MiR-548-3p
functions as an anti-oncogenic regulator in breast cancer. Biomed
Pharmacother. 75:111–116. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tashiro K, Kuroki N, Einama T, Iwasaki T,
Miyata Y, Aosasa S, Inoue Y, Takahashi Y, Ogata S, Ueno H, et al:
Prognostic significance of regional lymph node metastasis according
to station in ampullary carcinoma. J Hepatobiliary Pancreat Sci.
2020.(Epub ahead of print). View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mellis D and Caporali A: MicroRNA-based
therapeutics in cardiovascular disease: Screening and delivery to
the target. Biochem Soc Trans. 46:11–21. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Qadir MI and Faheem A: MiRNA: A diagnostic
and therapeutic tool for pancreatic cancer. Crit Rev Eukaryot Gene
Expr. 27:197–204. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Iglesias D, Fernandez-Peralta AM, Nejda N,
Daimiel L, Azcoita MM, Oliart S and Gonzalez-Aguilera JJ: RIS1, a
gene with trinucleotide repeats, is a target in the mutator pathway
of colorectal carcinogenesis. Cancer Genet Cytogenet. 167:138–144.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zirn B, Samans B, Wittmann S, Pietsch T,
Leuschner I, Graf N and Gessler M: Target genes of the
WNT/beta-catenin pathway in Wilms tumors. Genes Chromosomes Cancer.
45:565–574. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu L, Zhang J, Li S, Yin L and Tai J:
Silencing of TMEM158 inhibits tumorigenesis and multidrug
resistance in colorectal cancer. Nutr Cancer. 72:662–671. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kura B, Kalocayova B, LeBaron TW, Frimmel
K, Buday J, Surovy J and Slezak J: Regulation of microRNAs by
molecular hydrogen contributes to the prevention of
radiation-induced damage in the rat myocardium. Mol Cell Biochem.
457:61–72. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fabian MR, Sonenberg N and Filipowicz W:
Regulation of mRNA translation and stability by microRNAs. Annu Rev
Biochem. 79:351–379. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Correia de Sousa M, Gjorgjieva M, Dolicka
D, Sobolewski C and Foti M: Deciphering miRNAs' Action through
miRNA Editing. Int J Mol Sci. 20:62492019. View Article : Google Scholar
|