Introduction
Non-small-cell lung cancer (NSCLC) is one of the
most lethal types of cancer worldwide, accounting for 85% of lung
cancer cases (1). The majority of
patients with NSCLC die within one year of diagnosis and the 5-year
survival rate is as low as 17.8% (2). Surgical resection combined with
postoperative adjuvant therapies, such as chemotherapy and
radiotherapy, is a common treatment for NSCLC (3). The inability to diagnose and treat the
condition in the early stages is an important factor underlying the
low survival rate of patients with NSCLC (4). Thus, understanding the molecular
mechanisms of NSCLC onset and progression would provide invaluable
insight into early diagnosis and treatment, thereby enabling
effective targeted therapy.
In recent years, dysregulation of long non-coding
RNAs (lncRNAs) has been demonstrated to be an important causative
factor behind the development of NSCLC. Several lncRNAs, such as
actin filament associated protein 1 antisense RNA, X-inactive
specific transcript and maternally expressed 3, have been
implicated in NSCLC progression (5–7) and may
represent potential effective targets for diagnosis and targeted
therapy for NSCLC. Ribonuclease P RNA component H1 (RPPH1), a novel
lncRNA, serves a role in the pathogenesis of several human
diseases. For instance, a previous study suggested that RPPH1 could
promote hippocampal neuron dendritic spine formation by regulating
microRNA (miR)-330-5p and cell division cycle 42 in Alzheimer's
disease (8). RPPH1 is also thought
to be involved in the inflammatory pathogenesis of diabetic
nephropathy. Indeed, in a mouse model of diabetic nephropathy,
RPPH1 was abnormally overexpressed in renal tissue and enhanced the
release of inflammatory cytokines, including tumor necrosis
factor-α and monocyte chemotactic protein-1 (9). Moreover, Zhang et al (10) demonstrated that RPPH1 upregulation
enhanced the proliferative and colony formation abilities of breast
cancer cells. It has also been suggested that RPPH1 may aggravate
the development of breast cancer by suppressing the expression of
miR-122 (10). Liang et al
(10) demonstrated that RPPH1 could
promote colorectal cancer metastasis by interacting with tubulin β3
class III and promoting exosome-mediated macrophage M2
polarization. Furthermore, Lei et al (11) suggested that RPPH1 could enhance
human acute myeloid leukemia cell proliferation, migration and
invasion by reducing the expression of miR-330-5p (11).
Although evidence suggested that RPPH1 may be
associated with the onset of several diseases, the exact function
and mechanism of action underlying the role of RPPH1 in human
disease remain unclear. In the present study, the role of RPPH1 in
NSCLC progression was evaluated by studying its interaction with
miR-326 and Wnt family member 2B (WNT2B). miR-326 has been
previously implicated in the development of several types of human
tumors, including cervical cancer, lung cancer and breast cancer
(12–14). WNT2B is a crucial protein in the Wnt
signaling pathway that can act as an oncogene (15,16).
These findings may provide a novel molecular target and a
theoretical basis for the targeted therapy of NSCLC.
Materials and methods
The Cancer Genome Atlas (TCGA)
analysis
RPPH1 expression data in tumor (n=180) and adjacent
normal tissues (n=171) from patients with NSCLC were downloaded
from TCGA (https://tcga-data.nci.nih.gov/tcga/). The effect of
RPPH1 expression levels on patient 80-month overall survival of
patients was also evaluated using Kaplan-Meier survival analysis.
Lastly, the effect of RRPH1 expression on clinical stages was also
assessed (17).
Cell lines and culture
The human BEAS-2B normal lung epithelial cell line,
as well as H358, A549, H1299 and H1650 NSCLC cell lines were
purchased from The Cell Bank of Type Culture Collection of the
Chinese Academy of Sciences. All cell lines were separately
maintained in complete DMEM (Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Thermo Fisher Scientific, Inc.) at 37°C
with 5% CO2.
Reverse transcription-quantitative PCR
(RT-qPCR)
BEAS-2B, H358, A549, H1299 and H1650 cells were
trypsinized, then collected for total RNA extraction using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). Reverse transcription was performed in 20-µl reactions using
the Prime Script™ RT reagent kit (Takara Bio, Inc.) at 37°C for 15
min. The resulting cDNA templates were used for RT-qPCR, which was
performed using the SYBR-Green PCR Master Mix kit (Takara Bio,
Inc.) on the ABI Prism 7300 Sequence Detection System (Applied
Biosystems; Thermo Fisher Scientific, Inc.) The thermocycling
conditions were as follows: Initial denaturation at 95°C for 5 min;
36 cycles at 95°C for 30 sec, 58°C for 30 sec and 72°C for 40 sec;
and a final extension at 72°C for 10 min. The following primer
pairs were used for the qPCR: RPPH1 forward,
3′-CGAGCTGAGTGCGTCCTGTC-5′ and reverse, 3′-TCGCTGGCCGTGAGTCTGT-5′;
miR-326 forward, 3′-GGCGCCCAGAUAAUGCG-5′ and reverse,
3′-CGTGCAGGGTCCGAGGTC-5′; WNT2B forward, 3′-CTGAAGAGCCAAGCAAT-5′
reverse, 3′-CTCCAAGGCCATCCAAC-5′ and U6 forward,
3′-CTCGCTTCGGCAGCACA-5′ and reverse, 3′-AACGCTTCACGAATTTGCGT-5′. U6
served as the internal control, and the relative expression of
RPPH1, miR-326 and WNT2B was calculated using the 2−ΔΔCq
method (18).
Cell transfection
A549 and H1299 cells were plated at a density of
2×105 cells/well in six-well plates with serum-free DMEM
for transfections. RPPH1 short hairpin (sh) RNA (shRPPH1; 10 nM;
5′-UUGCCUUCAGUCGUGUGUAUCAG-3′; Shanghai Jima Biotechnology Co.,
Ltd.) and its negative control (sh-NC; 10 nM;
5′-AGUUCUGCGAACGUCGCACGU-3′; Shanghai Jima Biotechnology Co.,
Ltd.), were transfected into the cells. pcDNA3.1-RPPH1
overexpression vectors (GenePharma) and empty vectors (GenePharma)
were also constructed to transfect cells. Further transfections
were performed using a miR-326 mimic (10 nM;
5′-GCAGGGCACGACUGAUCUUGG-3′), miR-326 inhibitor (10 nM;
5′-UCGCUCGGUCCYGAUCGGGAG-3′) and their respective negative controls
(miR-NC, 5′-GGAAGUCAUCCAAUGUGCAUU-3′ and NC-inhibitor,
5′-UCCCUGGUUGCAGAUCGCGAA-3′). Co-transfection experiments were also
carried out using combinations of RPPH1 vectors, WNT2B
overexpression vector, miR-326 mimic and miR-326 inhibitor
combinations. The miR-326 mimic, miR-326 inhibitor, miR-NC and
WNT2B overexpression vectors were obtained from Shanghai Jima
Biotechnology Co., Ltd. All transfections were carried out using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.), following the manufacturer's instructions. Cells
were incubated at 37°C with 5% CO2 for 8 h, and the
medium was replaced with fresh DMEM containing 10% FBS. After 48-h
incubation, cells were harvested for subsequent
experimentation.
Transwell Matrigel assays
A549 and H1299 cells were harvested and re-suspended
in serum-free DMEM medium at a density of 1×105
cells/ml. A volume of 200 µl sample was added into Transwell
chambers with 8-µm pores pre-coated with a layer of Matrigel. All
Transwell chambers were then inserted into 24-well plates with 500
µl DMEM (10% FBS) added to the bottom of each well. The cells were
incubated at 37°C with 5% CO2 for 48 h. The Transwell
chambers were then removed and the membranes with adhered cells
were collected. Cells on the upper surface of the membrane were
gently scraped off using a cotton swab, whereas cells attached
under the surface of the membrane were gently rinsed with PBS.
Cells on the lower surface were fixed using 4% paraformaldehyde
solution, then stained 0.1% crystal violet for 20 min at room
temperature. The number of invading cells was counted under a light
microscope (magnification, ×200; Olympus Corporation).
Western blot analysis
Cells were collected and lysed on ice for 5 min
using RIPA buffer (Beyotime Institute of Biotechnology). The cell
lysate was then centrifuged for 10 min at 10,000 × g, 4°C and the
concentration of total protein in the supernatant was determined
using a bicinchoninic acid kit (Thermo Fisher Scientific, Inc.). A
total of 30 µg protein was resolved by SDS-PAGE on 10% gels (80 V
for 30 min and 120 V for 60 min). Protein samples were transferred
to a PVDF membrane at 110 V, which was then blocked with 5% skimmed
milk for 2 h at room temperature. The membrane was subsequently
incubated with the following primary antibodies (all 1:1,000;
Abcam): Mouse anti-E-cadherin (1:1,000; cat. no. ab76055; Abcam),
mouse anti-vimentin (1:1,000; cat. no. ab22651; Abcam), mouse
anti-β-catenin (1:1,000; cat. no. ab231305; Abcam) and mouse
anti-GAPDH (1:1,000; cat. no. ab8245; Abcam) for 12 h at 4°C. After
three washing steps with TBS-Tween-20 (TBS-T; 0.1% Tween-20), the
membranes were incubated with secondary antibodies (Beijing
Solarbio Science and Technology Co., Ltd.) for 1 h at room
temperature. The membranes were washed three times in TBST, and the
proteins bands were visualized using the Pierce ECL Western
Blotting kit (Pierce; Thermo Fisher Scientific, Inc.). Protein
expression was quantified using Image-Pro® Plus software
(version 6.0; Media Cybernetics, Inc.). GAPDH was used as an
internal control.
Cell Counting Kit-8 (CCK-8) assay
A549 and H1299 cells were seeded in 96-well plates
at 1×104 cells per well, in the presence or absence of 5
µg/ml cisplatin/cis-diamminedichloridoplatinum (CDDP; 100 µl;
Sigma-Aldrich: Merck KGaA). Cell were incubated 37°C, 5%
CO2 for 24, 48 and 72 h. A volume of 10 µl CCK-8
solution (Dojindo Molecular Technologies, Inc.) was added into each
well, and the cells were incubated for 4 h at room temperature.
After incubation, the optical density was measured in each well
using a microplate reader at a wavelength of 450 nm. In this assay,
optical density recording was proportional to cell viability and
therefore indicative of cell resistance to CDDP.
Colony formation assay
A549 and H1299 cells were plated at a density of
1×103 cells/dish in 90-mm cell culture dishes containing
5 ml complete DMEM complete + 10% FBS (with or without 5 µg/ml of
CDDP). All dishes were placed in a sterile incubator at 37°C with
5% CO2 for 2 weeks. The medium was changed every two
days. Cells were washed three times with PBS to remove residual
cells then fixed in 4% formaldehyde for 10 min at room temperature.
Crystal violet (0.1%) was then used to stain cells for 10 min at
room temperature. Colonies were counted under a light microscope.
Groups of >50 cells were considered a clone.
Luciferase reporter gene assay
StarBase 2.0 (http://starbase.sysu.edu.cn) was used to predict the
potential miRNAs that can bind to RPPH1, and TargetScan 7.2
(http://www.targetscan.org) was used to
predict the potential downstream target of miR-326. Wild-type
RPPH1, mutant RPPH1, wild-type WNT2B and mutant WNT2B constructs
were designed and purchased from Shanghai GenePharma Co., Ltd. All
four constructs were cloned into the pmirGLO reporter vectors. A549
cells were seeded into six-well plates at a density of
2×105 cells/well and co-transfected with the miR-NC,
miR-326 mimic, NC-inhibitor and miR-326 inhibitor group, as well as
the pmirGLO reporter vectors using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.). After 48-h incubation
at 37°C with 5% CO2, cells were collected and lysed. To
obtain the supernatant, solutions were centrifuged for 10 min at
10,000 × g and 4°C. The luciferase activity of cells was detected
using the Dual-Luciferase Reporter Assay System (Promega
Corporation). Renilla luciferase activity was used as
internal reference.
Statistical analysis
All experiments were repeated at least three times.
Statistical analysis was conducted using SPSS 19.0 (IBM Corp.).
Kaplan-Meier curves and the log-rank test were used to compare
differences in patient survival times. Cut-off values were
determined using median of expression. Differential gene expression
among tumor stages was evaluated by one-way ANOVA, and violin plots
were generated using the ‘vioplot’ R package (http://CRAN.R-project.org/package=vioplot).
Comparisons between two groups were carried out using Student's
t-test. Multigroup comparisons were performed using one-way ANOVA,
followed by Tukey's post hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
High RPPH1 expression levels is
associated with poor prognosis in patients with NSCLC
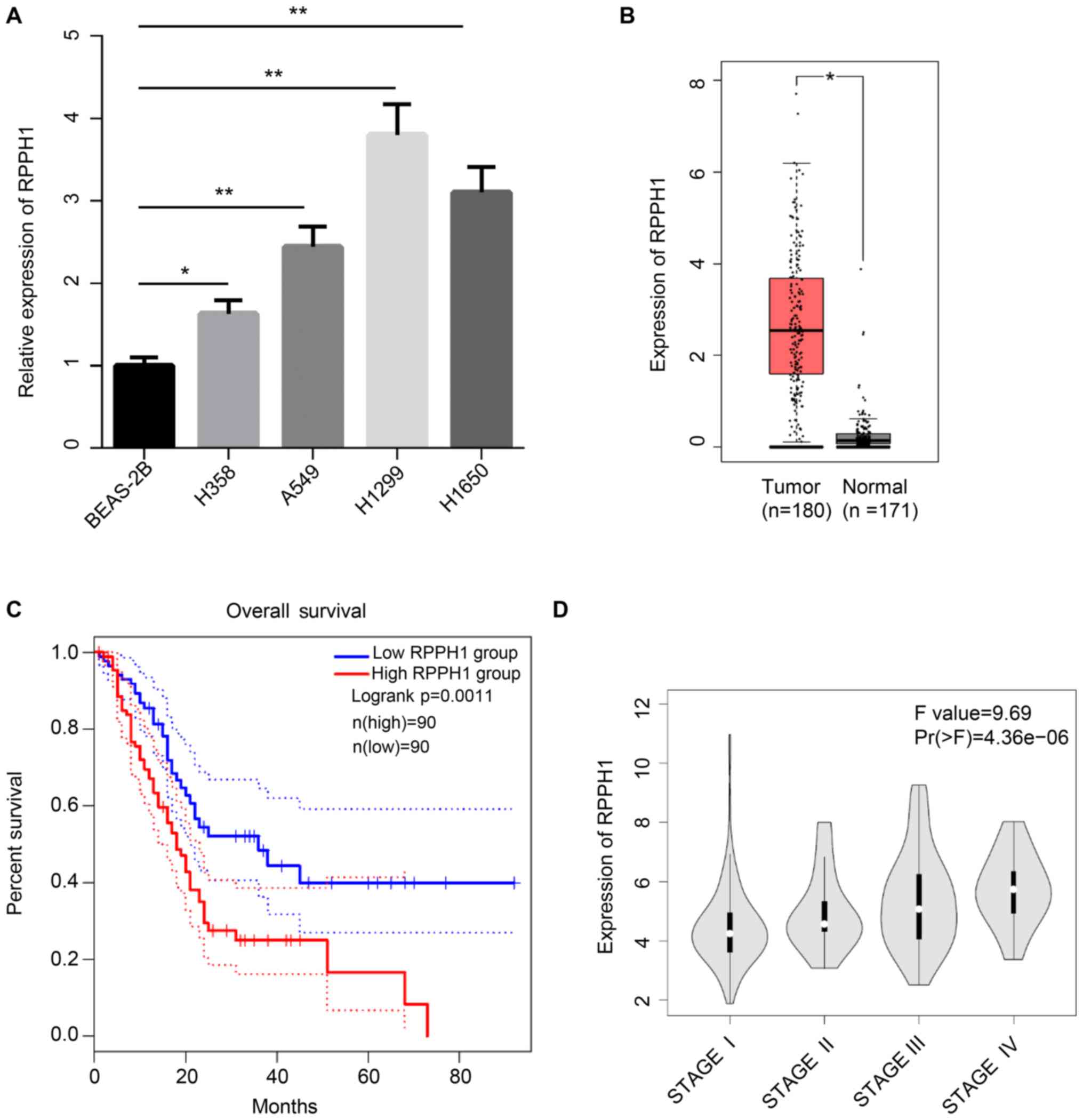
RPPH1 expression levels were significantly higher in
four NSCLC cell lines (H358, A549, H1299 and H1650), compared with
the BEAS-2B normal lung epithelial cell line (Fig. 1A). The association between RPPH1
expression levels and the prognosis of patients with NSCLC was also
analyzed in data obtained from TCGA. RPPH1 expression was
significantly increased in NSCLC tumor tissues compared with normal
tissues (Fig. 1B). Furthermore,
patients with low RPPH1 expression levels exhibited significantly
higher 80-month overall survival compared with patients with high
RPPH1 levels (Fig. 1C). The RPPH1
expression levels in patients with stage-IV NSCLC was also markedly
higher compared with patients with stage I, II and III (Fig. 1D). Therefore, high RPPH1 expression
levels in patients with NSCLC were associated with poor
prognosis.
RPPH1 knockdown inhibits NSCLC cell
invasion, epithelial- mesenchymal transition (EMT) and drug
resistance
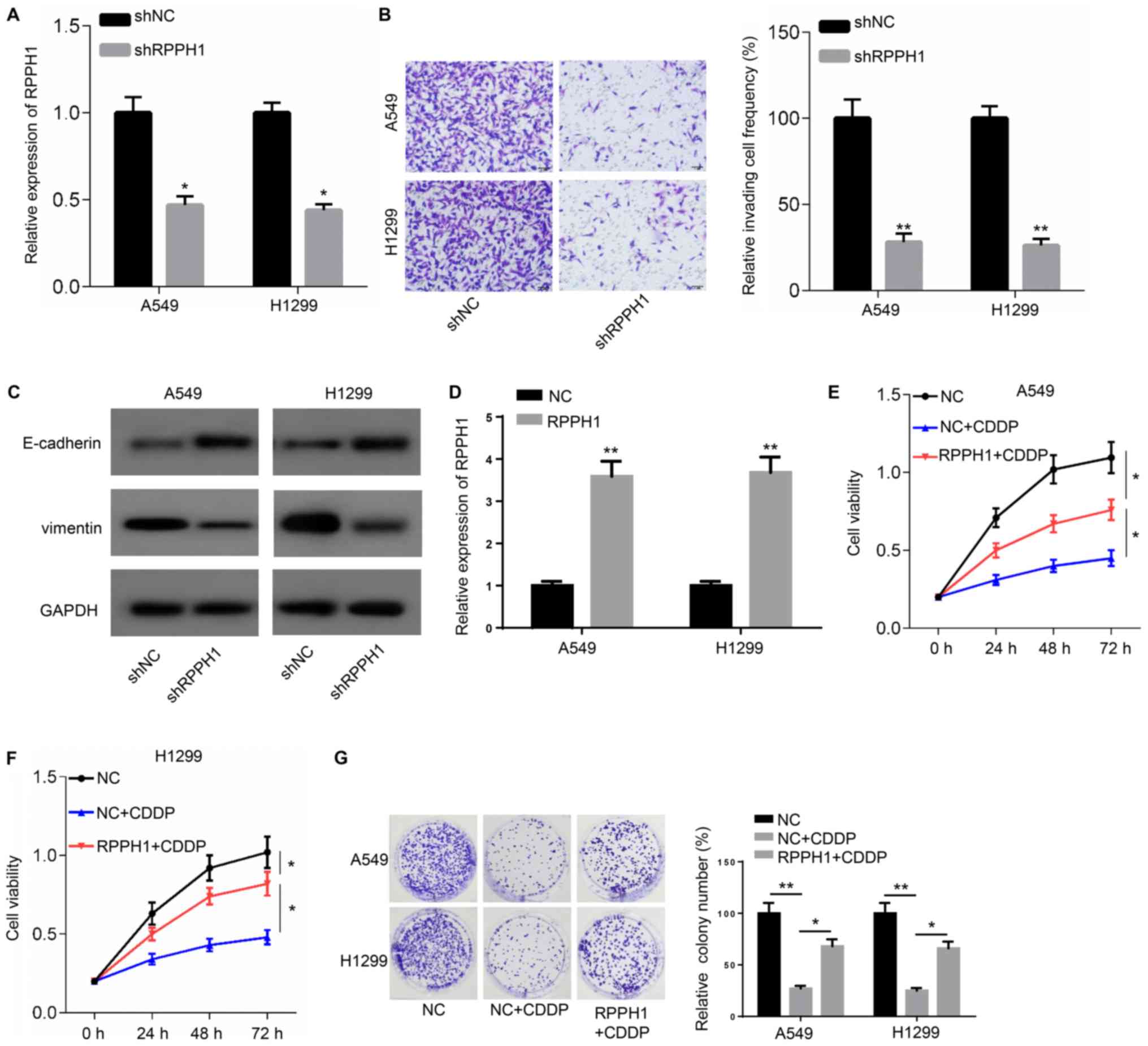
Since A549 and H1299 cell lines presented the higher
expression of RPPH1, A549 and H1299 were used in the following
experiments and were transfected with shRPPH1 or shNC. RPPH1
expression significantly declined in A549 and HI1299 cells
transfected with shRPPH1 compared with the shNC group (Fig. 2A). In addition, a significant
reduction in the number of invading cells was also observed in the
shRPPH1 group compared with the shNC group (Fig. 2B). Higher E-cadherin and lower
vimentin protein expression levels were detectable in A549 and
H1299 cells following transfection with shRPPH, compared with shNC
(Fig. 2C).
The expression of RPPH1 was significantly
upregulated in A549 and H1299 cells transfected with RPPH1
overexpression plasmid compared with the empty vector NC (Fig. 2D). Furthermore, cell viability and
colony numbers were significantly reduced in A549 and H1299 cells
in response to CDDP treatment, compared with untreated cells.
However, RRHP1 overexpression resulted in a significant increase in
cell viability and higher colony numbers in response to CDDP
compared with CDDP-treated, NC-transfected cells (Fig. 2E-G). Collectively, these observations
indicated that RPPH1 could increase the resistance of A549 and
H1299 to CDDP treatment.
miR-326 inhibitor partially rescues
the cell phenotype caused by RPPH1 knockdown
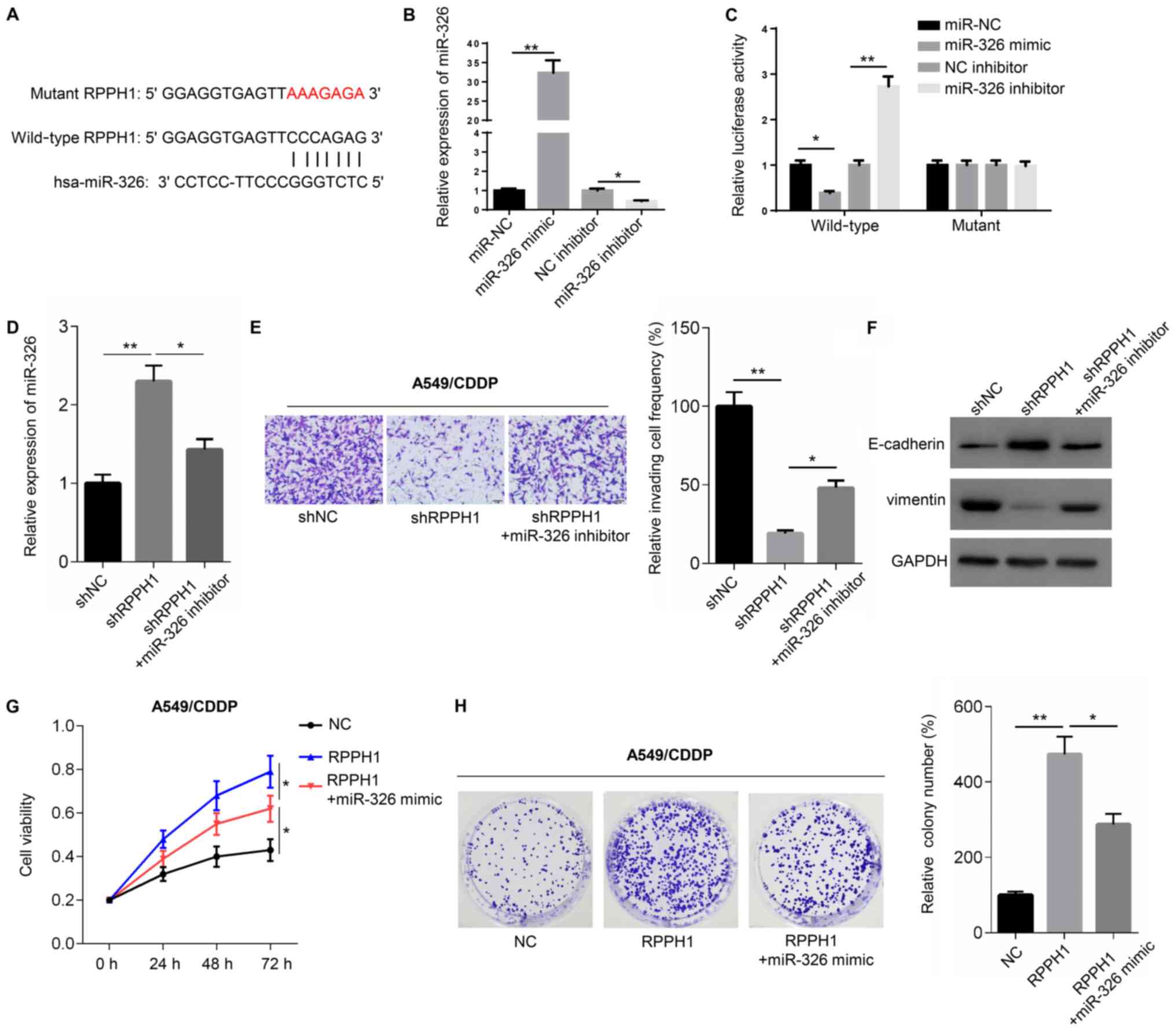
According to Starbase 2.0, RPPH1 contains a
potential binding site for miR-326 in its 3′-untranslated (3′-UTR)
region (Fig. 3A). Transfection with
miR-326 mimic significantly increased the expression of miR-326,
whereas the miR-326 inhibitor significantly decreased miR-326
expression, compared with their respective controls (Fig. 3B). To validate the potential
interaction between RPPH1 and miR-326, dual luciferase reporter
assays were performed. Compared with miR-NC, miR-326 mimic
significantly reduced the luciferase activity of cells transfected
with wild-type RPPH1 luciferase construct, but not with the mutant.
Similarly, the miR-326 inhibitor significantly increased the
luciferase activity of cells transfected with wild-type, but not
mutant, RPPH1 luciferase constructs (Fig. 3C). Moreover, compared with shNC
transfection shRPPH1-transfected A549 cells demonstrated
significantly higher miR-326 expression levels, significantly lower
numbers of invasive cells, higher E-cadherin expression levels and
reduced vimentin protein expression levels. However,
co-transfection with shRPPH1 and miR-326 inhibitor resulted in
significantly lower miR-326 expression levels, significantly higher
invasive cell frequencies, reduced E-cadherin levels and increased
vimentin protein expression, compared with shRPPH1 alone (Fig. 3D-F).
The resistance of A549 cells to CDDP was then
evaluated using CCK-8 and colony formation assays. A549 cells
transfected with the RPPH1 overexpression vector displayed
significantly higher viability and greater colony numbers, compared
with the empty vector NC (Fig. 3G and
H). However, cell viability and colony numbers were
significantly reduced in cells transfected with RPPH1 + miR-326
mimic, compared with RRPH1 alone. Altogether, these results
suggested that the role of RPPH1 may be mediated by miR-326.
miR-326 directly inhibits the Wnt
signaling pathway
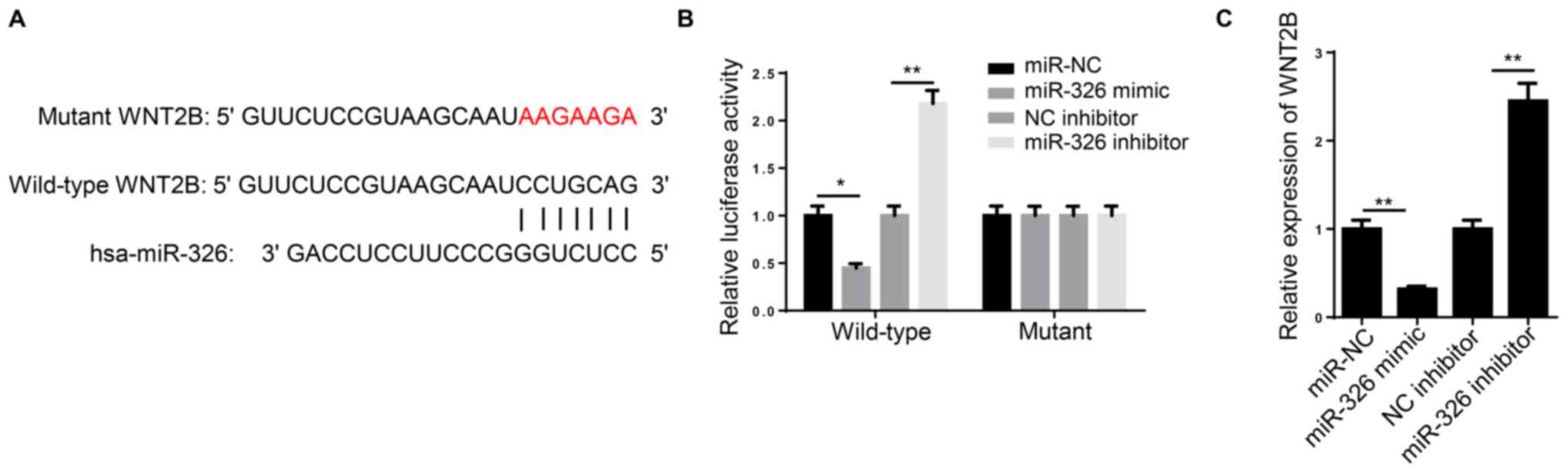
TargetScan 7.2 predicted that the miR-326 sequence
contained a binding site in the 3′-UTR region of WNT2B (Fig. 4A). In a dual luciferase reporter
assay, the luciferase activity of wild-type WNT2B, but not the
mutant construct, was significantly decreased following
co-transfection with miR-326 mimic, compared with miR-NC. By
contrast, miR-326 inhibitor transfection significantly increased
the wild-type WNT2B luciferase activity, but not that of the mutant
WNT2B (Fig. 4B).
WNT2B expression was significantly increased in A549
cells transfected with miR-326 mimic compared with miR-NC.
Conversely, transfection with miR-326 inhibitor resulted in
significantly higher WNT2B expression compared with the NC
inhibitor group (Fig. 4C). These
results indicated that WNT2B was directly inhibited by miR-326.
WNT2B partially rescues the cell
phenotype induced by miR-326 overexpression
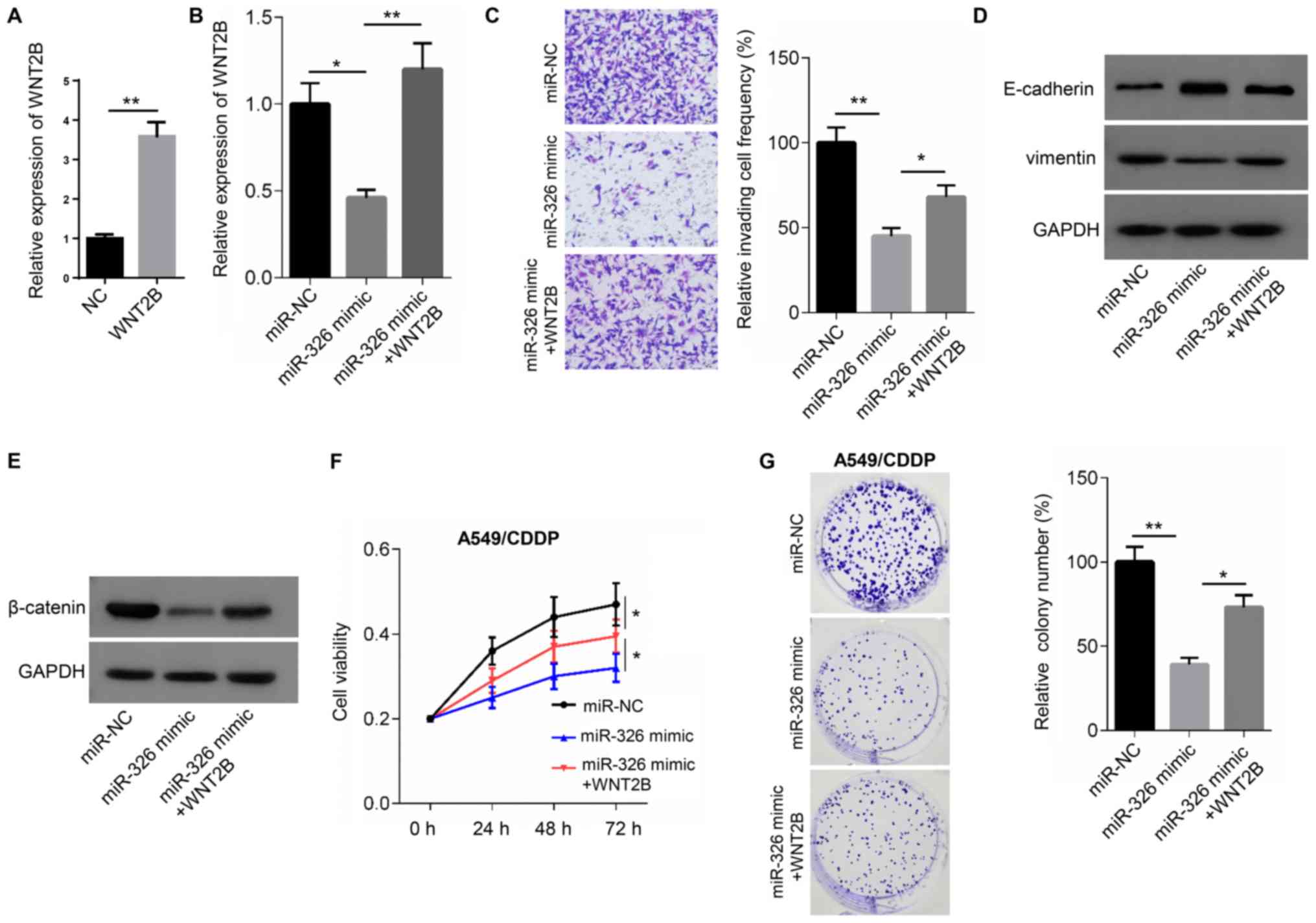
The expression of WNT2B was significantly
upregulated in A549 cells transfected WNT2B overexpression plasmid
compared with the NC group (Fig.
5A). Relative to the miR-NC group, reduced WNT2B expression
levels and invasive cell frequencies were observed in A549 cells
transfected with miR-326 mimic. Moreover, E-cadherin expression was
increased, while vimentin levels were reduced in miR-326
mimic-transfected cells compared with the miR-NC groups. However,
co-transfection with miR-326 mimic and WNT2B overexpression vector
resulted in higher WNT2B expression levels, compared with mimic
transfection alone. miR-326 mimic and WNT2B overexpression also
resulted in a significant increase in the frequency of invading
cells, a reduction in E-cadherin protein expression and an increase
in vimentin protein expression compared with the miR-326 mimic
groups (Fig. 5B-D). In addition,
lower β-catenin protein expression was observed in miR-326
mimic-transfected A549 cells compared with miR-NC. However, the
expression of β-catenin increased in cells co-transfected with
miR-326 mimic and WNT2B overexpression vector compared with cells
transfected with the mimic alone (Fig.
5E)
The aforementioned co-transfection experiments were
also carried out in the presence or absence of 5 µg/ml CDDP in
order to evaluate drug resistance in each of the groups. Cell
viability and colony numbers were significantly reduced in miR-326
mimic-transfected cells, compared with the miR-NC group. However,
co-transfection with the miR-326 mimic and WNT2B overexpression
vector significantly increased cell viability and colony numbers,
compared with transfection with the miR-326 mimic alone (Fig. 5F and G). Thus, WNT2B partially
rescued the cell phenotype induced by miR-326 overexpression.
Discussion
The present study demonstrated that RPPH1 was
upregulated in NSCLC tissues. Moreover, high RPPH1 expression was
associated with poor prognosis, including low 80-month overall
survival and advanced clinical stages. In vitro experiments
further indicated that RPPH1 could promote NSCLC progression via
regulation of miR-326 and WNT2B.
NSCLC is a major malignant tumor type and a leading
cause of cancer-related deaths worldwide, accounting for 85% of all
lung cancer cases (19). At the time
of diagnosis, most patients with NSCLC are already at a locally
advanced stage of the disease or have distantly metastatic tumors
(20). The 5-year survival rate is
particularly low in patients with advanced NSCLC, reaching only 4%
(21). NSCLC pathogenesis is not
fully understood, which may account for the lack of adequate
therapeutic strategies available to patients with this disease.
In recent years, lncRNAs have been considered to be
potential target for the treatment of NSCLC due to their regulatory
role in the development of this cancer type (22). LncRNAs are a class of RNA molecules
>200 nucleotides in length (23).
Despite their lack of protein-coding function, lncRNAs have been
demonstrated to regulate gene expression (24). LncRNAs participate in a variety of
cellular biological processes, including chromatin modification,
epigenetic regulation, cell cycle regulation, nucleoplasm
transport, transcription, translation and cell differentiation
(25,26). Therefore, dysregulation of lncRNA
function could lead to the onset and progression of diseases,
including tumors.
RPPH1 is an RNA subunit of RNase P, which is
reported to be abnormally overexpressed in the neocortex of
patients with seizures and in tumor tissues of patients with
gastric or breast cancer (10,27,28).
However, to the best of the authors' knowledge, the relationship
between RPPH1 and NSCLC has not been examined to date. In the
present study, RPPH1 was demonstrated to promote NSCLC progression
and to enhance the resistance of NSCLC cells to CDDP by regulating
the miR-326/WNT2B axis. Mechanistically, RPPH1 might enhance NSCLC
progression by competitively binding to miR-326, thereby inhibiting
the binding of miR-326 to its target gene, WNT2B. Consistent with
this hypothesis, previous studies have suggested that miR-326
expression is decreased in NSCLC primary tumor tissue and NSCLC
cells, and that miR-326 inhibits NSCLC cell migration and invasion
by suppressing the expression of the G1/S-specific cyclin-D1
oncogene (29). The present findings
suggested that RPPH1 silencing reduced the invasion potential and
EMT in NSCLC cell lines. Moreover, downregulation of miR-326
partially rescued this phenotype.
In several types of human malignancies, including
lung cancer, resistance to CDDP is a major obstacle to successful
therapy (30). The present study
indicated that RPPH1 overexpression could increase the CDDP
resistance of NSCLC cells. Moreover, miR-326 overexpression
reversed the CDDP resistance conferred by RPPH1 upregulation.
Similarly, Li et al (31)
demonstrated that miR-326 was downregulated in CDDP-resistant A549
cells compared with wild-type A549 cells. Furthermore, patients
with lung adenocarcinoma receiving CDDP chemotherapy displayed
reduced miR-326 levels in their tumor tissues (31). Consistent with these previous
findings, miR-326 acted as a tumor suppressor in NSCLC tissue in
the present study and could inhibit CDDP resistance induced by
RPPH1 overexpression.
WNT2B is a paralogue of WNT2 and is one of the key
molecules in the Wnt signaling pathway (32). WNT2B promotes the progression of head
and neck squamous cell carcinoma, malignant pleural mesothelioma,
ovarian cancer and pancreatic cancer, and enhances chemotherapy
resistance and metastasis, leading to poor prognosis (33–36). In
nasopharyngeal carcinoma cells, WNT2B expression is directly
upregulated by decreased miR-324-3p expression levels, which in
turn enhances migration, invasion and EMT (37). Furthermore, Wang et al
(16) suggested that abnormally
elevated WNT2B expression levels in NSCLC cells were the result of
miR-577 downregulation. High expression of WNT2B is associated with
a malignant phenotype in NSCLC cells, including enhanced cell
viability, migration and invasion (16). One possible underlying mechanism may
be that reduced miR-577 expression promotes the Wnt/β-catenin
pathway by regulating WNT2B expression. The aforementioned previous
studies emphasize the oncogenic role of WNT2B in human malignant
tumor types. Similarly, the present study also demonstrated that
WNT2B expression was increased in NSCLC cells through the
downregulation of miR-326 and that WNT2B could promote NSCLC cell
invasion, EMT and resistance to CDDP.
In conclusion, RPPH1 acted an oncogene in NSCLC,
which enhanced NSCLC progression and resistance to CDDP through the
miR-326/WNT2B axis. The present article provides a theoretical
framework the development of therapeutic strategies for NSCLC, with
RPPH1 as a novel molecular target for NSCLC treatment.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
YW and XW contributed to the conception and design
of the study. KC, WL, XW and YW performed the experiments. YW and
KC analyzed the data. YW and XW wrote the manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Brahmer JR, Govindan R, Anders RA, Antonia
SJ, Sagorsky S, Davies MJ, Dubinett SM, Ferris A, Gandhi L, Garon
EB, et al: The Society for Immunotherapy of Cancer consensus
statement on immunotherapy for the treatment of non-small cell lung
cancer (NSCLC). J Immunother Cancer. 6:752018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lei Z, Shi H, Li W, Yu D, Shen F, Yu X, Lu
D, Sun C and Liao K: MiR-185 inhibits non-small cell lung cancer
cell proliferation and invasion through targeting of SOX9 and
regulation of Wnt signaling. Mol Med Rep. 17:1742–1752.
2018.PubMed/NCBI
|
|
3
|
Tanaka F and Yoneda K: Adjuvant therapy
following surgery in non-small cell lung cancer (NSCLC). Surg
Today. 46:25–37. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sotgia F and Lisanti MP: Mitochondrial
markers predict survival and progression in non-small cell lung
cancer (NSCLC) patients: Use as companion diagnostics. Oncotarget.
8:68095–68107. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yin D, Lu X, Su J, He X, De W, Yang J, Li
W, Han L and Zhang E: Long noncoding RNA AFAP1-AS1 predicts a poor
prognosis and regulates non-small cell lung cancer cell
proliferation by epigenetically repressing p21 expression. Mol
Cancer. 17:922018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Guo X, Wei Y, Wang Z, Liu W, Yang Y, Yu X
and He J: LncRNA LINC00163 upregulation suppresses lung cancer
development though transcriptionally increasing TCF21 expression.
Am J Cancer Res. 8:2494–2506. 2018.PubMed/NCBI
|
|
7
|
Pei W, Dong C, Hongbing M and Yong L: Long
non-coding RNA MEG3 regulates proliferation and apoptosis in
non-small cell lung cancer via the miR-205-5p/LRP1 pathway. Rsc
Adv. 7:49710–49719. 2017. View Article : Google Scholar
|
|
8
|
Cai Y, Sun Z, Jia H, Luo H, Ye X, Wu Q,
Xiong Y, Zhang W and Wan J: Rpph1upregulates CDC42 expression and
promotes hippocampal neuron dendritic spine formation by competing
with miR-330-5p. Front Mol Neurosci. 10:272017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang P..Sun Y..Peng R..et al: Long
non-coding RNA Rpph1 promotes inflammation and proliferation of
mesangial cells in diabetic nephropathy via an interaction with
Gal-3. Cell Death Dis. 10:5262019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liang ZX, Liu HS, Wang FW, Xiong L, Zhou
C, Hu T, He XW, Wu XJ, Xie D, Wu XR and Lan P: LncRNA RPPH1
promotes colorectal cancer metastasis by interacting with TUBB3 and
by promoting exosomes-mediated macrophage M2 polarization. Cell
Death Dis. 10:8292019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lei B, He A, Chen Y, Cao X, Zhang P, Liu
J, Ma X, Qian L and Zhang W: Long non-coding RNA RPPH1 promotes the
proliferation, invasion and migration of human acute myeloid
leukemia cells through down-regulating miR-330-5p expression. EXCLI
J. 18:824–837. 2019.PubMed/NCBI
|
|
12
|
Jiang H, Liang M, Jiang Y, Zhang T, Mo K,
Su S, Wang A, Zhu Y, Huang G and Zhou R: The lncRNA TDRG1 promotes
cell proliferation, migration and invasion by targeting miR-326 to
regulate MAPK1 expression in cervical cancer. Cancer Cell Int.
19:1522019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang R, Chen X, Xu T, Xia R, Han L, Chen
W, De W and Shu Y: MiR-326 regulates cell proliferation and
migration in lung cancer by targeting phox2a and is regulated by
HOTAIR. Am J Cancer Res. 6:173–186. 2016.PubMed/NCBI
|
|
14
|
Ghaemi Z, Soltani BM and Mowla SJ:
MicroRNA-326 functions as a tumor suppressor in breast cancer by
targeting ErbB/PI3K signaling pathway. Front Oncol. 9:6532019.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu W, Zhang B, Xu N, Wang MJ and Liu Q:
MiR-326 regulates EMT and metastasis of endometrial cancer through
targeting TWIST1. Eur Rev Med Pharmacol Sci. 21:3787–3793.
2017.PubMed/NCBI
|
|
16
|
Wang B, Sun L, Li J and Jiang R: MiR-577
suppresses cell proliferation and epithelial-mesenchymal transition
by regulating the WNT2B mediated Wnt/β-catenin pathway in non-small
cell lung cancer. Mol Med Rep. 18:2753–2761. 2018.PubMed/NCBI
|
|
17
|
Detterbeck FC, Boffa DJ, Kim AW and Tanoue
LT: The eighth edition lung cancer stage classification. Chest.
151:193–203. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hai J, Zhu CQ, Wang T, Organ SL, Shepherd
FA and Tsao MS: TRIM14 is a putative tumor suppressor and regulator
of innate immune response in non-small cell lung cancer. Sci Rep.
7:396922017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Stinchcombe TE and Socinski MA: Current
treatments for advanced stage non-small cell lung cancer. Proc Am
Thorac Soc. 6:233–241. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang L, Ma L, Xu F, Zhai W, Dong S, Yin L,
Liu J and Yu Z: Role of long non-coding RNA in drug resistance in
non-small cell lung cancer. Thorac Cancer. 9:761–768. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Schmitt AM and Chang HY: Long Noncoding
RNAs in cancer pathways. Cancer Cell. 29:452–463. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Noh JH, Kim KM, McClusky WG, Abdelmohsen K
and Gorospe M: Cytoplasmic functions of long noncoding RNAs. Wiley
Interdiscip Rev RNA. 9:e14712018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen QN, Wei CC, Wang ZX and Sun M: Long
non-coding RNAs in anti-cancer drug resistance. Oncotarget.
8:1925–1936. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jiang C, Li X, Zhao H and Liu H: Long
non-coding RNAs: Potential new biomarkers for predicting tumor
invasion and metastasis. Mol Cancer. 15:622016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xia T, Liao Q, Jiang X, Shao Y, Xiao B, Xi
Y and Guo J: Long noncoding RNA associated-competing endogenous
RNAs in gastric cancer. Sci Rep. 4:60882014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Leonard L, Dachet F, Cai J, Bagla S, Balan
K, Jia H and Loeb JA: Activity-dependent human brain
coding/noncoding gene regulatory networks. Genetics. 192:1133–1148.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sun C, Huang C, Li S, Yang C, Xi Y, Wang
L, Zhang F, Fu Y and Li D: Hsa-miR-326 targets CCND1 and inhibits
non-small cell lung cancer development. Oncotarget. 7:8341–8359.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sun Y, Zheng S, Torossian A, Speirs CK,
Schleicher S, Giacalone NJ, Carbone DP, Zhao Z and Lu B: Role of
insulin-like growth factor-1 signaling pathway in
cisplatin-resistant lung cancer cells. Int J Radiat Oncol Biol
Phys. 82:e563–e572. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li J, Li S, Chen Z, Wang J, Chen Y, Xu Z,
Jin M and Yu W: MiR-326 reverses chemoresistance in human lung
adenocarcinoma cells by targeting specificity protein 1. Tumour
Biol. 37:13287–13294. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Katoh M: Differential regulation of WNT2
and WNT2B expression in human cancer. Int J Mol Med. 8:657–660.
2001.PubMed/NCBI
|
|
33
|
Li SJ, Yang XN and Qian HY: Antitumor
effects of WNT2B silencing in GLUT1 overexpressing cisplatin
resistant head and neck squamous cell carcinoma. Am J Cancer Res.
5:300–308. 2014.PubMed/NCBI
|
|
34
|
Kobayashi M, Huang CL, Sonobe M, Kikuchi
R, Ishikawa M, Kitamura J, Miyahara R, Menju T, Iwakiri S, Itoi K,
et al: Intratumoral Wnt2B expression affects tumor proliferation
and survival in malignant pleural mesothelioma patients. Exp Ther
Med. 3:952–958. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang H, Fan L, Xia X, Rao Y, Ma Q, Yang J,
Lu Y, Wang C, Ma D and Huang X: Silencing Wnt2B by siRNA
interference inhibits metastasis and enhances chemotherapy
sensitivity in ovarian cancer. Int J Gynecol Cancer. 22:755–761.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jiang H, Li F, He C, Wang X, Li Q and Gao
H: Expression of Gli1 and Wnt2B correlates with progression and
clinical outcome of pancreatic cancer. Int J Clin Exp Pathol.
7:4531–4538. 2014.PubMed/NCBI
|
|
37
|
Liu C, Li G, Yang N, Su Z, Zhang S, Deng
T, Ren S, Lu S, Tian Y, Liu Y and Qiu Y: MiR-324-3p suppresses
migration and invasion by targeting WNT2B in nasopharyngeal
carcinoma. Cancer Cell Int. 17:22017. View Article : Google Scholar : PubMed/NCBI
|