Introduction
Osteosarcoma, a common primary malignant bone
cancer, is often diagnosed in children and it is the second leading
cause of cancer-related deaths of children (1). Characterized by high malignancy and
recurrence rate, it affects the life of patients to a large extent.
Although surgical resection and neoadjuvant chemotherapy have
reduced the mortality of patients with osteosarcoma, the 5-year
survival rate is still markedly low, with 10–20% of patients with
metastasis (2,3). Therefore, it is crucial to identify new
diagnostic biomarkers and therapeutic targets, understand the
influence and mechanism of these targets on the malignant
biological behavior of osteosarcoma cells in order to improve the
clinical treatment and the prognosis of the patients (4,5).
Long-chain non-coding RNAs (lncRNAs) are RNA
molecules that exist in the nucleus or cytoplasm, with a length of
more than 200 nts and have a complex secondary structure (6). lncRNAs play an important role in
various biological and pathological processes, such as
transcriptional regulation, cell fate determination and
tumorigenesis. lncRNAs can adsorb miRNA, block miRNA, and inhibit
the binding of miRNA to its target genes (6). In addition, lncRNAs plays an important
role in the process of tumorigenesis and tumor progression
(7). It has been revealed that
numerous lncRNAs, including MEG3, HOTAIR, CCAL, tumor suppressor
candidate 7 (TUSC7) play a regulatory role in the occurrence and
progression of osteosarcoma at the transcriptional and
post-transcriptional levels (8,9), and
participate in the proliferation, invasion, and migration of the
tumor (10). lncRNA TUSC7 was
originally identified in osteosarcoma by Pasic et al
(11). The discovery of the function
of lncRNA TUSC7 as a tumor-inhibiting factor in human osteosarcoma
has attracted broad attention (12).
Several studies have revealed that the lncRNA TUSC7 exhibits a
downregulated expression in numerous malignant tumors, such as
liver cancer (13), neuroglioma
(14), colorectal cancer (15), and endometrial cancer (16). A previous study also revealed that
the expression of TUSC7 in osteosarcoma was significantly lower
than that in non-tumor tissues (12). lncRNA TUSC7 is a type of potential
tumor-inhibiting factor, which can be used as a biomarker and the
therapeutic target of osteosarcoma patients. However, how lncRNA
TUSC7 regulates the proliferation of osteosarcoma cells remains
unknown.
miRNAs are short non-coding RNAs (~21 nucleotides)
that can regulate gene expression at the post-transcriptional level
(17). Studies have found that
miR-375 can be used as a tumor therapeutic target and a tumor
diagnosis mark, and it is differentially expressed in diseases such
as esophageal, liver and cervical cancer, participating in the
progression of the disease (17–19). A
previous study revealed that miR-375 was aberrantly downregulated
in osteosarcoma cells and functioned as a tumor suppressor
(20). In addition, miR-375 has been
revealed to inhibit osteogenic differentiation (21), which is involved in the growth and
proliferation of osteosarcoma. A study reported that
calcium-binding protein S100A6 has been revealed to promote human
osteosarcoma growth by promoting cell proliferation and inhibiting
osteogenic differentiation (22).
The flavonoid phellodendron glycoside can inhibit the proliferation
of human osteosarcoma cells and stimulate osteogenic
differentiation (23). Therefore,
miR-375 may promote the growth of osteosarcoma, and its role
remains to be further clarified.
To the best of our knowledge, there is no previous
study on the regulatory relationship between miR-375 and lncRNA
TUSC7 in osteosarcoma cells. Based on the inhibitory effect of long
non-coding RNA TUSC7 on the invasion of osteosarcoma cells, it was
explored whether lncRNA TUSC7 can influence the malignant
biological behavior of the osteosarcoma cells by regulating
miR-375.
Materials and methods
Materials
Human osteosarcoma cells (HOS and MG63) and human
fetal osteoblast cells (hFOB 1.19) were purchased from ATCC.
Dulbecco's modified Eagle's Medium (DMEM) and fetal bovine serum
(FBS) were obtained from Gibco; Thermo Fisher Scientific, Inc. The
Transwell chamber was obtained from BD Biosciences. The Two-step
fluorescence quantitative PCR kit was purchased from
MedChemExpress. Penicilin-streptamycin solution (double antibiotic)
were obtained from Hyclone; GE Healthcare Life Sciences.
Consumables such as cell culture dishes and plates were purchased
from Corning Costar, Inc. TRIzol reagent was obtained from Cwbio IT
Group and Lipo3000 transfection reagent was obtained from
Invitrogen; Thermo Fisher Scientific, Inc..
Patient and tissue samples
The present study included 30 patients (17 males and
13 females; 15–24 years old) with osteosarcoma admitted to the
Orthopedic Surgery Department of the ShengLi Oilfield Central
Hospital from January 2010 to December 2019, where postoperative
tissue samples were collected to analyze the pathological results.
The samples were used to extract total RNA. All patients or their
families signed informed and written consent. The study was
approved by the ShengLi Oilfield Central Hospital Ethics Committee
(approval no. SOCH20091201A).
Culture and plasmid transfection of
osteosarcoma cells
HOS cells and MG63 cells were cultured in DMEM
containing 10% FBS and 1% penicillin/streptomycin in an 5%
CO2 and 37°C incubator. Although the levels of Lnc-TUSC7
in both HOS and MG63 cells were detected, the changes in MG63 cells
were more significant than those of HOS cells, and subsequent
experiments were performed with MG63 cells. MG63 cells were
digested with 0.25% Trypsin pancreatin (Gibco; Thermo Fisher
Scientific, Inc.) and 3×105 cells/well were seeded into
a six-well plate. Transfection was carried out after the cell
confluence reached 60–70%. A total of 40 nM pcDNA-TUSC7, negative
control plasmid pcDNA3.1, miR-375 mimics and miR-negative control
(NC) were transfected using Lipo3000 transfection reagent according
to the manufacturer's instructions. After 48 h, the total RNA of
the cells was extracted to detect the transfection efficiency.
Then, G418 (500 µg/ml, Gibco; Thermo Fisher Scientific, Inc.) was
used to select cells for 4 weeks to obtain MG63 cells with stable
expression of lncRNA TUSC7.
The primer sequences of the pcDNA-TUSC7 expression
vector was as follows: Forward,
5′-CGATCTTAATTAAGGGGTACCAAAGTCCACTCTG-3′ and reverse,
5′-TCAGTGGCGCGCCTTTTTCGTGAGTACACAATAGTCATC-3′. The miR-375 mimic
sequence was as follows: Forward,
5′-CGATCTAATTAAGGGTACCAAAGTCACTCTG-3′ and reverse,
5′-TCAGGGCGCGCCTTCGTGGTACACAATAGTCAT-3′.
Transwell assay
Osteosarcoma cells with a fusion rate of 70–80%
after transfection of MG63 cells in each group were collected,
washed with PBS, centrifuged to remove the supernatant, and this
process was repeated 2 times. Then, serum-free DMEM (Gibco; Thermo
Fisher Scientific, Inc.) was added to adjust the cell concentration
to 2.5×105 cells/ml. Subsequently, 200 µl of cell
suspension was added to the upper layer of a Transwell chamber
prepared in advance, and the chamber was placed in a 24-well
culture plate with medium containing 10% serum (Gibco; GE
Healthcare Life Sciences) mixed in advance. Then, the culture plate
was placed in a 37°C, 5% CO2 incubator for 24 h. After
the culture was finished, the number of cells migrating from
Transwell upper chamber to lower chamber membrane were observed
under a light microscope (×20). Finally, the Transwell chamber was
washed with PBS, the non-migrated cells were removed with a cotton
swab and the chamber was air-dried. Then, 4% formaldehyde solution
was added to the chamber for fixation at room temperature for 20
min, and 0.1% crystal violet was added for staining at room
temperature for 20 min. Finally, the chamber was cleaned with PBS 3
times and dried naturally. The number of cells was counted and
images were observed and captured under a light microscope
(×40).
MTT assay
Osteosarcoma cells with a cell fusion rate of 70–80%
after transfection in each group were collected and washed with PBS
solution. A small number of cells was obtained and counted, and
then the cell concentration was adjusted to 1×104
cells/ml. Diluted cell suspension (200 µl) was seeded onto 96-well
cell plates. The culture plate was placed in a 37°C, 5%
CO2 incubator for 24 h, and then 50 µl MTT solution
(Beyotime Institute of Biotechnology) was added to the cell wells,
cautiously avoiding light during this process. The cells were
cultured for another 3–4 h, the supernatant was discarded, and 150
µl DMSO solution was added to each well. The cells were shaken on a
shaker at low speed for 10 min and finally the absorbance value of
each well was detected at 490 nm with a microplate reader. Both the
48 and 72-h plates also underwent the aforementioned treatment.
Real-time qPCR assay
The transfected cells and tissues with a fusion rate
of 70–80% were collected into an EP tube, and total RNA was
extracted using TRIzol reagent (CoWin Biosciences). cDNA products
were prepared using cDNA synthesis kit (iScript™ cDNA
kit; cat. no. 1708890; Bio-Rad Laboratories), and the reaction
system was configured according to the specifications of the
quantitative kit (SYBR Green qPCR Master Mix; cat. no. HY-K0501;
MedChemExpress). The whole process (<1 h) was completed on ice.
Then the configured reaction was placed into a tube and an ABI7500
fluorescence quantitative PCR instrument (Applied Biosystems;
Thermo Fisher Scientific, Inc.) was used to detect gene expression.
The PCR reaction conditions were: 95°C pre-denaturation for 10 min;
95°C denaturation for 15 sec; 60°C annealing for 45 sec; 72°C
extension for 1 min; 40 cycles; 72°C extension for 5 min; 4°C
retention. The relative expression of the target gene was
calculated by 2−ΔΔCq method (24). The primer sequences were as follows:
lncRNA TUSC7 forward, 5′-GGAAACAGAAGGCACCTCA-3′ and reverse
5′-TCTCAGAGGTCAAACAGGCA-3′; miR-375 forward,
5′-AGCCGTCAAGAGCAATAACGAA-3′ and reverse, 5′-GTGCAGGGTCCGAGGT-3′;
and U6 forward, 5′-CGCTTCGGCAGCACATATAC-3′ and reverse,
5′-AGGGGCCATGCTAATCTTCT-3′.
Luciferase reporter gene assay
Control and miR-375 WT, lncRNA TUSC7 and miR-375 WT,
NC and miR-375 MUT, lncRNA TUSC7 and miR-375 MUT were transfected
into MG63 cells (Lipo3000; Invitrogen; Thermo Fisher Scientific,
Inc.), respectively, in accordance with the specifications of the
luciferase reporter gene test system (Promega) kit.pmirGLO
luciferase vector and luciferase reporter plasmid were constructed
by GenScript. The relative fluorescence intensity of each group
after 48 h of transfection was detected. Renilla luciferase
activity was used as a standardized method.
Statistical analysis
SPSS 17.0 statistical software (SPSS, Inc.) was used
to process the experimental data. An unpaired Student's t-test was
used for comparison of differences between two groups, and ANOVA
followed by Bonferroni test was used for comparison of differences
between multiple groups. Pearson correlation analysis was also
performed. The experimental results were expressed as the mean ±
SD, and each experiment was repeated 3 times. P<0.05 was
considered to indicate a statistically significant difference;
(*P<0.05, **P<0.01, ***P<0.001 as indicated in the figures
and legends).
Results
lncRNA TUSC7 and miR-375 are
differently expressed in osteosarcoma tissue and adjacent
tissue
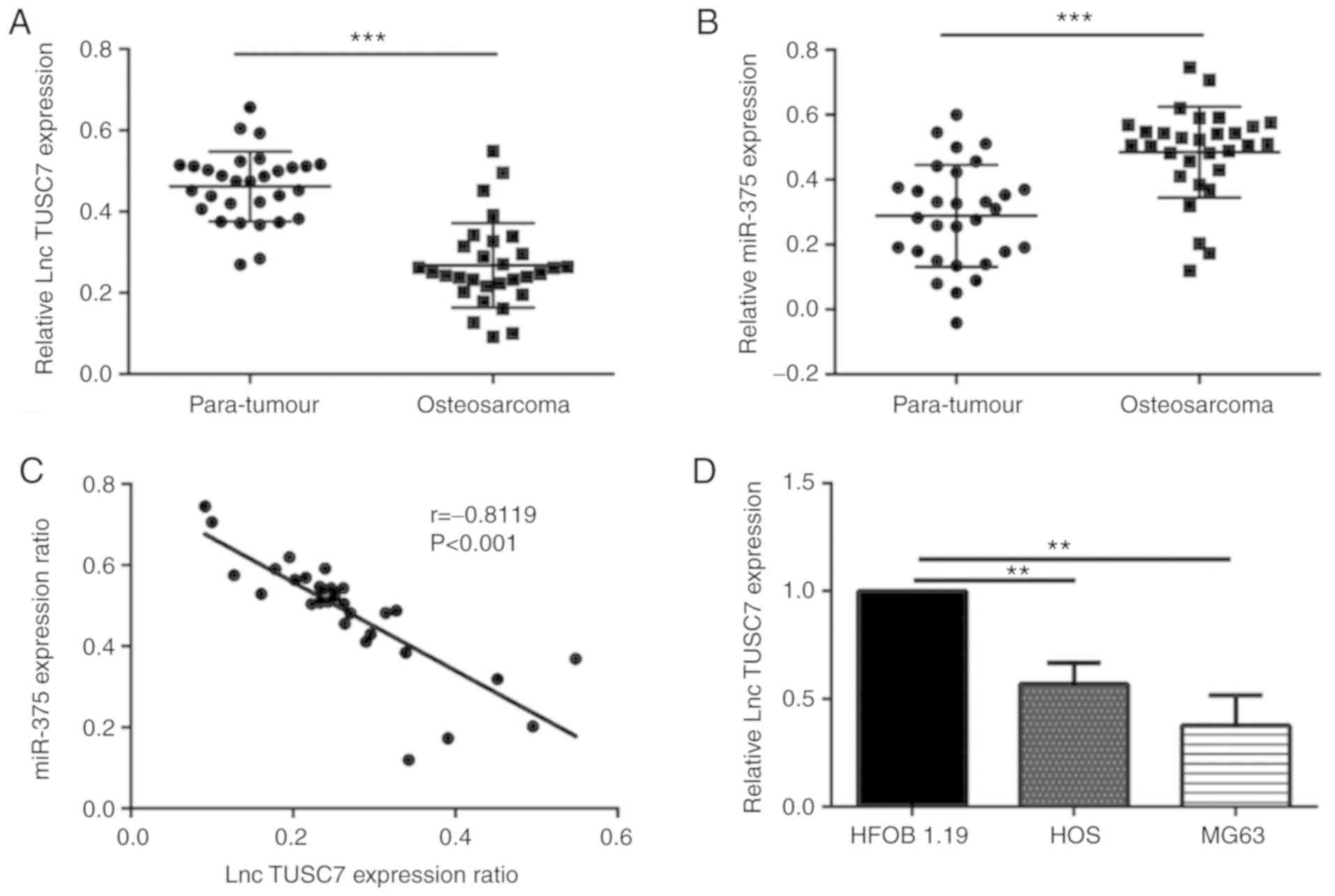
Tissue samples and para-tumor tissues were collected
from 30 patients with osteosarcoma, and qPCR was used to detect the
expression level of lncRNA TUSC7 and miR-375. It was revealed that
lncRNA TUSC7 in osteosarcoma tissue was significantly lower than
that of para-tumor tissue (P<0.001), while the level of miR-375
was significantly higher than that of the adjacent tissues
(P<0.001), and the difference was statistically significant
(Fig. 1A and B). Through correlation
analysis, it was revealed that there was a negative correlation
between lncRNA TUSC7 and miR-375, as revealed in Fig. 1C (r=−0.8119, P<0.001). To
determine the difference in the expression levels of lncRNA TUSC7,
HOS, MG63 and HFOB 1.19 cell lines were selected and the expression
of lncRNA TUSC7 was detected by RT-qPCR. The results revealed that
lncRNA TUSC7 expression in HOS and MG63 cells was significantly
lower than that in HFOB 1.19 cells, indicating that lncRNA TUSC7
expression was downregulated in HOS and MG63 cells (Fig. 1D).
Proliferation and migration abilities
of MG63 cells transfected with lncRNA TUSC7
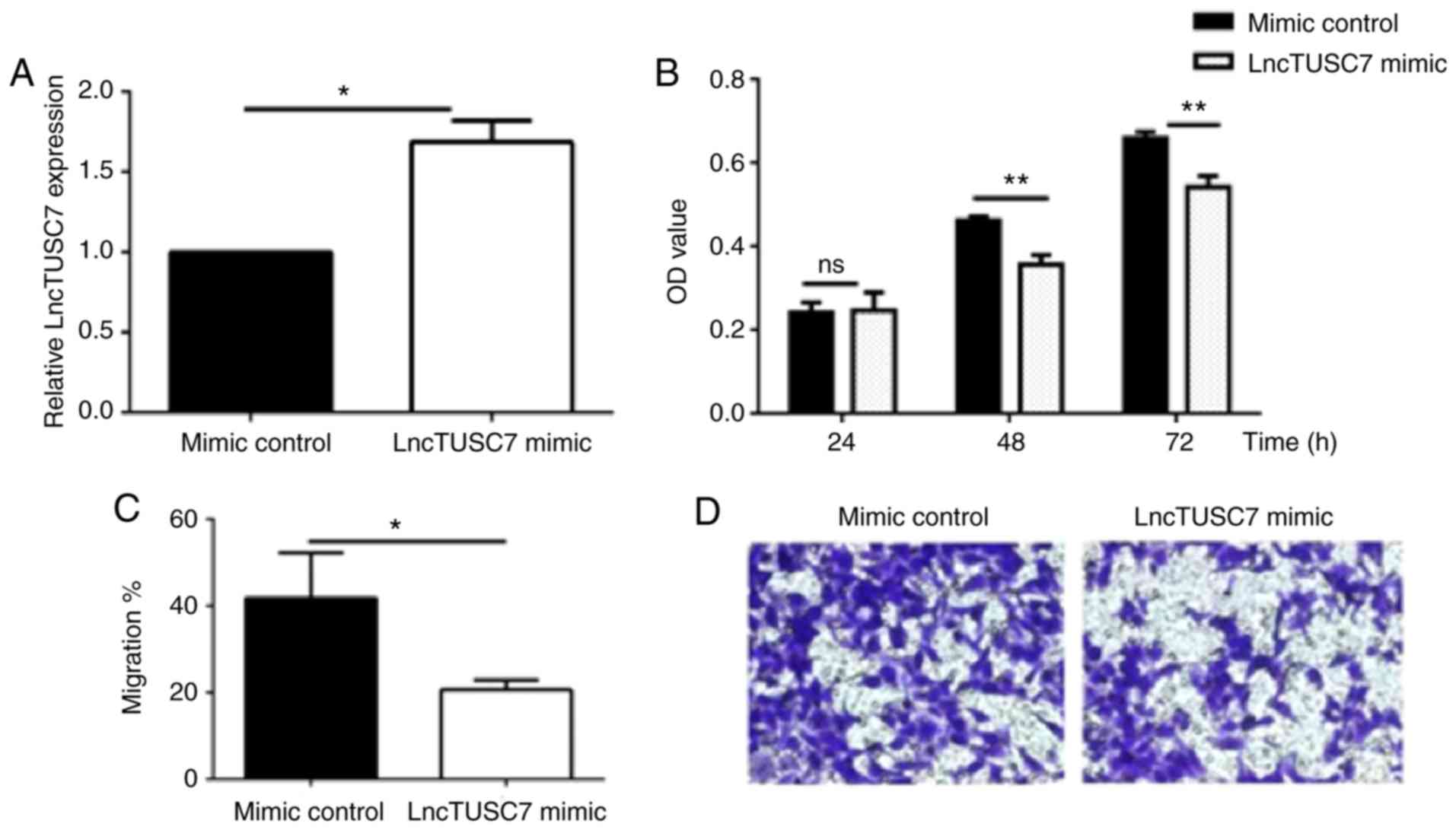
In order to further understand the effect of lncRNA
TUSC7 on the biological function of MG63, the proliferation and
migration abilities of MG63 were detected through MTT and Transwell
experiments. RT-qPCR experiments revealed that in MG63 cells
transfected with lncRNA pcDNA-TUSC7, the expression level of lncRNA
TUSC7 was significantly enhanced compared with that in the control
group, indicating that the MG63 cell line that stably expressed the
target gene was successfully transfected (Fig. 2A). In addition, MTT assays and
Transwell migration assays revealed that compared with the mimic
control group, the proliferation (Fig.
2B) and migration (Fig. 2C and
D) abilities in the lncRNA TUSC7 mimic group were significantly
decreased. The aforementioned results indicated that lncRNA TUSC7
inhibited the proliferation and migration abilities of osteosarcoma
MG63 cells to a certain extent.
Effects of miR-375 on the
proliferation and migration abilities of osteosarcoma cells
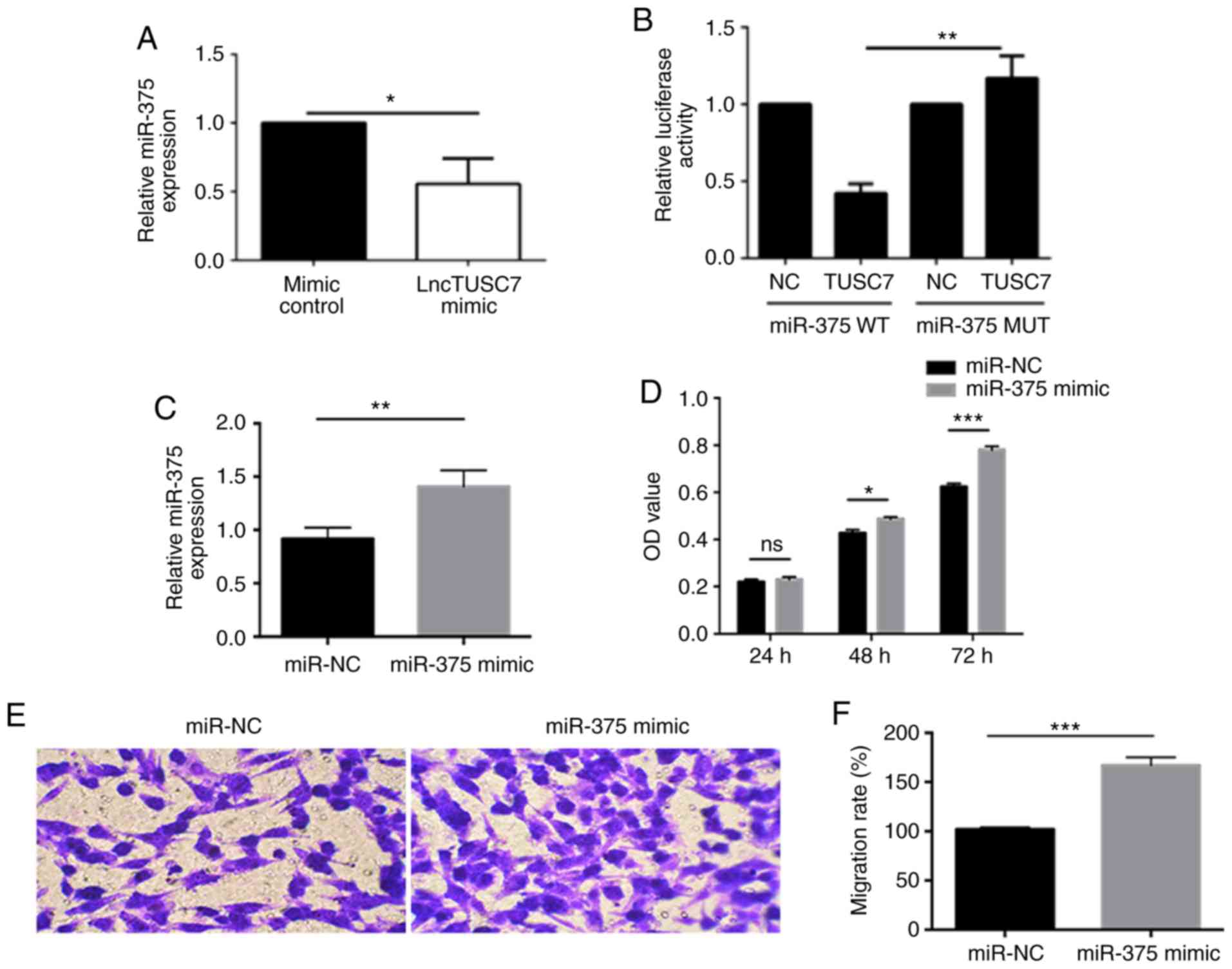
After establishing the lncRNA TUSC7-mimic
transfection model, the expression of miR-375 was detected and it
was revealed that compared with the mimic control, miR-375
expression was significantly downregulated in the lncRNA
TUSC7-mimic MG63 cells (Fig. 3A).
Concurrently, it was demonstrated that miR-375 was the target gene
of lncRNA TUSC7 through a luciferase reporter gene assay (Fig. 3B). In addition, MG63 cells were
transfected with the miR-375 mimic and it was revealed that the
expression level of miR-375 in the miR-375-mimic-transfected group
was significantly higher than that of the miR-NC-transfected group
(Fig. 3C). Furthermore, the
proliferation capacity was significantly higher in the
miR-375-mimic-transfected group than that of the miR-NC-transfected
control group (Fig. 3D). The
Transwell migration assay revealed that the miR-375-mimic group
also had higher migratory capacity compared with the miR-NC control
group (Fig. 3E and F), indicating
that miR-375 promoted the growth and migration of osteosarcoma.
lncRNA TUSC7 affects the proliferation
and migration of osteosarcoma cells through miR-375
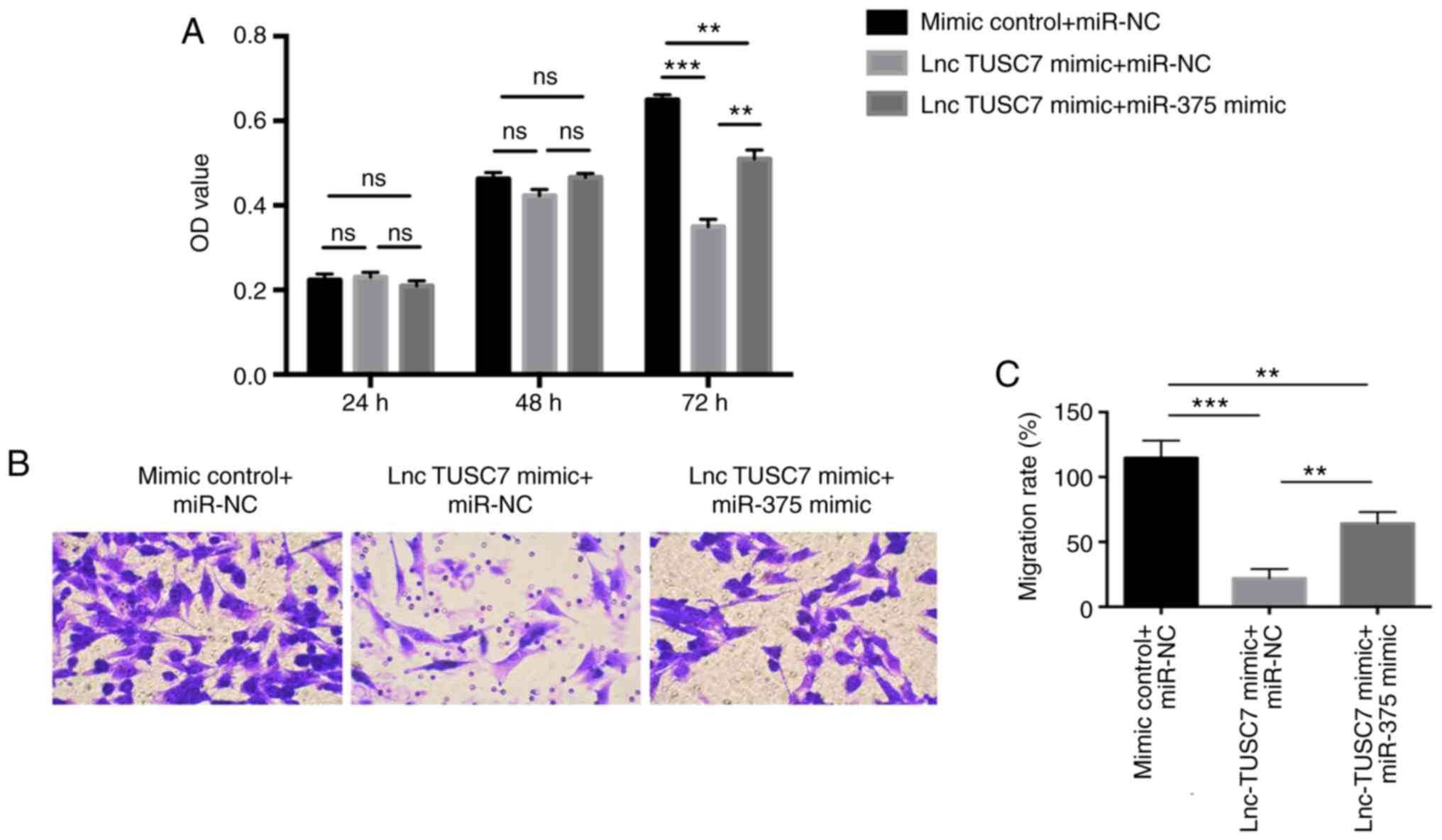
miR-375 mimic and lncRNA TUSC7 mimic were
co-transfected into MG63 osteosarcoma cells, and it was revealed
that the proliferation ability of lncRNA TUSC7 mimic + miR-375
mimic group was significantly higher than that of lncRNA TUSC7
mimic + miR-NC group, but lower than that in the control group
(mimic control+miR-NC), especially at 72 h (Fig. 4A). The same trend was also revealed
in the Transwell migration assay. The migration ability of the
lncRNA TUSC7 mimic + miR-375 mimic group was significantly higher
than that of the lncRNA TUSC7 mimic + miR-NC group (Fig. 4B and C), indicating that miR-375
mimic can partially rescue the inhibitory effect of lncRNA TUSC7 on
osteosarcoma.
Discussion
Osteosarcoma (OS) is a common primary bone neoplasm
and one of the most invasive solid malignant tumors (1). Evidently, the occurrence of
osteosarcoma is correlated with numerous factors such as race, age,
sex, various genomic changes, and the environment people live in
(25). However, to date, the cause
of the disease is still controversial (1–4). In the
present study, it was determined that lncRNA TUSC7 in osteosarcoma
tissue was significantly lower than that in the adjacent tissue,
while miR-375 level in osteosarcoma tissue was significantly higher
than that in the adjacent tissue, and there was a negative
correlation between the two. The expression differences of two
types of osteosarcoma cell lines (HOS and MG63) and the normal
osteoblast HFOB 1.19 cell line were examined. It was revealed that
the expression of lncRNA TUSC7 in osteosarcoma cells was lower than
that in normal cells, especially in MG63 cells. The present study
confirmed that lncRNA TUSC7 exhibited an antitumor effect in
osteosarcoma cell line MG63. lncRNAs are a type of RNA, defined as
regulatory transcripts that have no-protein coding potential and
are more than 200 nucleotides in length (16,26,27). By
regulating genetic expression at the transcriptional,
post-transcriptional or epigenetic levels, it has been revealed
that lncRNAs participate in cell activities and the formation of
various diseases (28). According to
the ENCODE project (29), more than
90 percent of the human genome can be transcribed, and only about
two percent of the genome are protein-coding genes (30). Therefore, it has been revealed that
most transcriptomes are of noncoding transcripts, including
microRNAs, lncRNAs, and circular RNAs (30). As oncogenes or cancer suppressor
genes, lncRNAs play an important role in the process of tumor
development (30,31).
TUSC7 consists of four exons that are located on
chromosome 3q13.3 (11). Studies
have demonstrated that TUSC7 exhibits a downregulated expression in
most cancers and plays an anticancer role (15). TUSC7 has been revealed to inhibit the
process of the epithelial-mesenchymal transition (13) in liver cancer by negatively
regulating miR-10a. The overexpression of TUSC7 inhibited the
occurrence of glioma and gastric cancer by targeting miR-23b
(14). The upregulation of TUSC7 has
been revealed to inhibit the proliferation of lung cancer,
colorectal cancer and osteosarcoma (15). These aforementioned studies revealed
that TUSC7 can play an anticancer role by sponging certain
tumor-related miRNAs. In the present study, a transfection cell
model with lncRNA TUSC7 mimic was set up to detect the influence of
lncRNA TUSC7 on the malignant biological behavior of MG63 cells. We
successfully constructed the MG63 cell line expressing lncRNA
TUSC7, and also used qPCR experiments to verify that the expression
level of lncRNA TUSC7 in the lncRNA TUSC7-mimic group was
significantly upregulated. Then, the effect of lncRNA TUSC7 on the
migration ability of the MG63 osteosarcoma cells was assessed using
Transwell migration assays. The results revealed that in comparison
with the control group, the migration ability of MG63 cells
transfected with the lncRNA TUSC7 mimic was significantly
downregulated. Similarly, the MTT assay revealed that the
proliferation ability of the lncRNA TUSC7-mimic group was
significantly lower than that of the mimic control group,
indicating that lncRNA TUSC7 could inhibit the growth of MG63
cells. Collectively the aforementioned results indicated that TUSC7
can function as a type of cancer suppressor gene in osteosarcoma
cells. TUSC7 with high expression can inhibit the proliferation
rate and the migration rate of tumor cells.
It has been demonstrated that the aberrant
expression of miR-375 is correlated with the occurrence and
progression of a variety of cancers, including colorectal (15), lung (14), oral (18,20) and
breast cancer (19), as well as
glioma (32) and gastric cancer
(33). It has been revealed that the
expression of miR-375 is increased in gastric cancer, yet miR-375
can desensitize cells to ionizing radiation and etoposide via
targeting of p53 (33). miR-375 has
been revealed to downregulate human colorectal cancer cell lines
and tissues, and inhibit the growth of colorectal cancer cells
(34–36). In addition, it has been revealed that
miR-375 inhibits osteogenic differentiation (21), and studies have confirmed that tumor
growth can be inhibited by promoting osteogenic differentiation
(22,23). Therefore, miR-375 may promote the
growth of osteosarcoma. The present study revealed that miR-375
expression in tumor tissue of osteosarcoma patients was reduced,
and miR-375 mimic could enhance the malignant biological behavior
of MG63 cells. It was revealed that miR-375 plays a role in
promoting the growth of osteosarcoma, which is consistent with the
results of some previous studies (22,23).
lncRNAs are reported to regulate gene expression,
chromosome modification, or the function of miRNAs (37) through binding to RNA-binding
proteins. In the present study it was further confirmed through a
luciferase reporter gene assay that miR-375 is the target gene of
lncRNA TUSC7, and that co-transfection of lncRNA TUSC7 mimic and
miR-375 mimic could partially rescue the inhibitory effect of
lncRNA TUSC7 on MG63 cells, confirming that lncRNA TUSC7 inhibits
the malignant biological behavior of osteosarcoma by regulating the
expression of miR-375. The specific mechanism of how lncRNA TUSC7
and miR-375 affects the biological behavior of osteosarcoma needs
to be further clarified. It is suggested that miR-375 inhibits
osteogenic differentiation and lncRNA TUSC7 may promote osteogenic
differentiation by inhibiting the expression of miR-375. In
addition, miR-375 has a variety of target genes, which involve
complex cell signaling pathway regulation processes, and requires
further exploration. In the future, extensive data using
bioinformatics will be used to confirm the binding site between
lncRNA TUSC7 and miR-375, and the mechanism will be further studied
through the method of binding site mutation. lncRNA TUSC7 may
inhibit osteosarcoma cell growth by acting on miR-375, which
provides us with a new perspective for treating patients.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
LW, JJ and GS conceived and designed the study, and
drafted the manuscript. LW, JJ, PZ and YL collected, analyzed and
interpreted the experimental data. GS revised the manuscript for
important intellectual content. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
ShengLi Oilfield Central Hospital (approval no. SOCH20091201A).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lin YH, Jewell BE, Gingold J, Lu L, Zhao
R, Wang LL and Lee DF: Osteosarcoma: Molecular pathogenesis and
iPSC modeling. Trends Mol Med. 23:737–755. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang J, Liu S, Shi J, Li J, Wang S, Liu H,
Zhao S, Duan K, Pan X and Yi Z: The role of miRNA in the diagnosis,
prognosis, and treatment of osteosarcoma. Cancer Biother
Radiopharm. 34:605–613. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wan J and Zhang X, Liu T and Zhang X:
Strategies and developments of immunotherapies in osteosarcoma.
Oncol Lett. 11:511–520. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lindsey BA, Markel JE and Kleinerman ES:
Osteosarcoma overview. Rheumatol Ther. 4:25–43. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Geller DS and Gorlick R: Osteosarcoma: A
review of diagnosis, management, and treatment strategies. Clin Adv
Hematol Oncol. 8:705–718. 2010.PubMed/NCBI
|
|
6
|
Xu S, Gong Y, Yin Y, Xing H and Zhang N:
The multiple function of long noncoding RNAs in osteosarcoma
progression, drug resistance and prognosis. Biomed Pharmacother.
127:1101412020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li Z, Shen J, Chan MT and Wu WK: TUG1: A
pivotal oncogenic long non-coding RNA of human cancers. Cell
Prolif. 49:471–475. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li N, Shi K and Li W: TUSC7: A novel tumor
suppressor long non-coding RNA in human cancers. J Cell Physiol.
233:6401–6407. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hung T and Chang HY: Long noncoding RNA in
genome regulation: Prospects and mechanisms. RNA Biol. 7:582–585.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang B, Su Y, Yang Q, Lv D, Zhang W, Tang
K, Wang H, Zhang R and Liu Y: Overexpression of long non-coding RNA
HOTAIR promotes tumor growth and metastasis in human osteosarcoma.
Mol Cells. 38:432–440. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pasic I, Shlien A, Durbin AD, Stavropoulos
DJ, Baskin B, Ray PN, Novokmet A and Malkin D: Recurrent focal
copy-number changes and loss of heterozygosity implicate two
noncoding RNAs and one tumor suppressor gene at chromosome 3q13.31
in osteosarcoma. Cancer Res. 70:160–171. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cong M, Li J, Jing R and Li Z: Long
non-coding RNA tumor suppressor candidate 7 functions as a tumor
suppressor and inhibits proliferation in osteosarcoma. Tumour Biol.
37:9441–9450. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang Y, Liu Z, Yao B, Dou C, Xu M, Xue Y,
Ding L, Jia Y, Zhang H, Li Q, et al: Long non-coding RNA TUSC7 acts
a molecular sponge for miR-10a and suppresses EMT in hepatocellular
carcinoma. Tumour Biol. 37:11429–11441. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shang C, Guo Y, Hong Y and Xue YX: Long
non-coding RNA TUSC7, a target of miR-23b, plays tumor-suppressing
roles in human gliomas. Front Cell Neurosci. 10:2352016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ren W, Chen S, Liu G, Wang X, Ye H and Xi
Y: TUSC7 acts as a tumor suppressor in colorectal cancer. Am J
Transl Res. 9:4026–4035. 2017.PubMed/NCBI
|
|
16
|
Wu X, Cai D, Zhang F, Li M and Wan Q: Long
noncoding RNA TUSC7 inhibits cell proliferation, migration and
invasion by regulating SOCS4 (SOCS5) expression through targeting
miR-616 in endometrial carcinoma. Life Sci. 231:1165492019.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Krol J, Loedige I and Filipowicz W: The
widespread regulation of microRNA biogenesis, function and decay.
Nat Rev Genet. 11:597–610. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Siow MY, Ng LP, Vincent-Chong VK,
Jamaludin M, Abraham MT, Abdul Rahman ZA, Kallarakkal TG, Yang YH,
Cheong SC and Zain RB: Dysregulation of miR-31 and miR-375
expression is associated with clinical outcomes in oral carcinoma.
Oral Dis. 20:345–351. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Luo D, Wilson JM, Harvel N, Liu J, Pei L,
Huang S, Hawthorn L and Shi H: A systematic evaluation of
miRNA:mRNA interactions involved in the migration and invasion of
breast cancer cells. J Transl Med. 11:572013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu X, Ajani JA, Gu J, Chang DW, Tan W,
Hildebrandt MA, Huang M, Wang KK and Hawk E: MicroRNA expression
signatures during malignant progression from Barrett's esophagus to
esophageal adenocarcinoma. Cancer Prev Res (Phila). 6:196–205.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Du F, Wu H, Zhou Z and Liu YU:
microRNA-375 inhibits osteogenic differentiation by targeting
runt-related transcription factor 2. Exp Ther Med. 10:207–212.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li Y, Wagner ER, Yan Z, Wang Z, Luther G,
Jiang W, Ye J, Wei Q, Wang J, Zhao L, et al: The calcium-binding
protein S100A6 accelerates human osteosarcoma growth by promoting
cell proliferation and inhibiting osteogenic differentiation. Cell
Physiol Biochem. 37:2375–2392. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang N, Ying MD, Wu YP, Zhou ZH, Ye ZM,
Li H and Lin DS: Hyperoside, a flavonoid compound, inhibits
proliferation and stimulates osteogenic differentiation of human
osteosarcoma cells. PLoS One. 9:e989732014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Prater S and Mckeon B: Cancer,
osteosarcoma. StatPearls [Internet] Treasure Island (FL):
StatPearls Publishing; 2020, https://www.ncbi.nlm.nih.gov/pubmed/31751058
|
|
26
|
Liu Y, Xing R, Zhang X, Dong W, Zhang J,
Yan Z, Li W, Cui J and Lu Y: miR-375 targets the p53 gene to
regulate cellular response to ionizing radiation and etoposide in
gastric cancer cells. DNA Repair (Amst). 12:741–750. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chang Y, Yan W, He X, Zhang L, Li C, Huang
H, Nace G, Geller DA, Lin J and Tsung A: miR-375 inhibits autophagy
and reduces viability of hepatocellular carcinoma cells under
hypoxic conditions. Gastroenterology. 143:177–187 e8. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Song W, Xie J, Li J, Bao C and Xiao Y: The
emerging roles of long noncoding RNAs in bone homeostasis and their
potential application in bone-related diseases. DNA Cell Biol.
39:926–937. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
ENCODE Project Consortium, Birney E,
Stamatoyannopoulos JA, Dutta A, Guigó R, Gingeras TR, Margulies EH,
Weng Z, Snyder M, Dermitzakis ET, et al: Identification and
analysis of functional elements in 1% of the human genome by the
ENCODE pilot project. Nature. 447:799–816. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Esteller M: Non-coding RNAs in human
disease. Nat Rev Genet. 12:861–874. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ling H, Vincent K, Pichler M, Fodde R,
Berindan-Neagoe I, Slack FJ and Calin GA: Junk DNA and the long
non-coding RNA twist in cancer genetics. Oncogene. 34:5003–5011.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chang C, Shi H, Wang C, Wang J, Geng N,
Jiang X and Wang X: Correlation of microRNA-375 downregulation with
unfavorable clinical outcome of patients with glioma. Neurosci
Lett. 531:204–208. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xu Y, Deng Y, Yan X and Zhou T: Targeting
miR-375 in gastric cancer. Expert Opin Ther Targets. 15:961–972.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Elshafei A, Shaker O, Abd El-Motaal O and
Salman T: The expression profiling of serum miR-92a, miR-375, and
miR-760 in colorectal cancer: An Egyptian study. Tumour Biol.
39:10104283177057652017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cui F, Wang S, Lao I, Zhou C, Kong H,
Bayaxi N, Li J, Chen Q, Zhu T and Zhu H: miR-375 inhibits the
invasion and metastasis of colorectal cancer via targeting SP1 and
regulating EMT-associated genes. Oncol Rep. 36:487–493. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xu L, Li M, Wang M, Yan D, Feng G and An
G: The expression of microRNA-375 in plasma and tissue is matched
in human colorectal cancer. BMC Cancer. 14:7142014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Arab K, Park YJ, Lindroth AM, Schäfer A,
Oakes C, Weichenhan D, Lukanova A, Lundin E, Risch A, Meister M, et
al: Long noncoding RNA TARID directs demethylation and activation
of the tumor suppressor TCF21 via GADD45A. Mol Cell. 55:604–614.
2014. View Article : Google Scholar : PubMed/NCBI
|