Introduction
Breast cancer is the most commonly occurring type of
cancer among women (1). The Global
Health Organization reported that 508,000 women succumbed to breast
cancer in 2011 (2). There are a
number of factors involved in the occurrence and development of
breast cancer, such as tissue transglutaminase (TG2) and
transforming growth factor-β (TGF-β) family members (3,4).
TGF-β1 is a member of the transforming
growth factor superfamily that is widely involved in various
pathophysiological processes, such as inflammation, trauma and
organ fibrosis (5). TG2 is an enzyme
that is upregulated in epithelial malignancies and participates in
Ca2+-dependent protein post-translational modifications
and cross-linking via the acyl-transfer reaction between glutamine
and lysine residues (6). It has been
reported that TG2 serves an important role in the
epithelial-to-mesenchymal transition (EMT) (6). In addition, upregulation of TGF-β is
associated with metastasis, cell invasiveness and EMT in ovarian
cancer (7,8). However, to the best of our knowledge,
the synergistic role of TG2 and TGF-β1 in regulating the
occurrence and development of breast cancer has been reported.
Cisplatin is a broad-spectrum anticancer drug that
is commonly used in ovarian, prostate, testicular and lung cancer,
nasopharyngeal carcinoma, esophageal cancer, malignant lymphoma,
head and neck squamous cell carcinoma and thyroid cancer (9). However, whether the expression of TG2
and TGF-β1 is regulated by cisplatin remains to be
elucidated.
The present study aimed to analyze the role of TG2
and TGF-β1 in breast cancer. In addition, the present
study aimed to investigate the protein levels of TGF-β1
and TG2 in MCF-7 cells treated with cisplatin and the effect of TG2
and TGF-β1 in MCF-7 cells treated with TGF-β1
and TG2 inhibitors or TGF-β1.
Materials and methods
Tissue samples
A total of 30 pairs of breast cancer and
paracancerous tissue samples were obtained from the China-Japan
Union Hospital (Changchun, China) between March 2018 and March
2019. The median age is 38 years (range, 28–45 years). The study
was approved by the Ethics Committee of the China-Japan Union
Hospital. The samples were obtained with signed informed consent
from the patients or their family.
Cell culture
MCF-7 cells were gifted from Jilin University School
of Pharmacy. The cells were cultured in Dulbecco's modified Eagle's
medium (DMEM) supplemented with 10% FBS, 100 U/ml penicillin G and
100 µg/ml streptomycin in an incubator at 37°C and 5%
CO2. (all from Invitrogen; Thermo Fisher Scientific,
Inc.).
Reagents
All materials for the SDS-PAGE were purchased from
Bio-Rad Laboratories, Inc. The monoclonal antibody against β-actin
(1,2000; cat. no. AAPR201-100) was purchased from Sigma-Aldrich;
Merck KGaA. Rabbit polyclonal antibodies against TGF-β1
(1:2,000; cat. no. RAB-0238) and TG2 (1:2,000; cat. no. CTA-DE056)
were obtained from Cell Signaling Technology, Inc. The TG2
inhibitor (MDC) and the TGF-β1 inhibitor (ITD) were
purchased from Sigma-Aldrich; Merck KGaA.
Hematoxylin and eosin (HE)
staining
The sections (3 µm) of breast cancer tissue were
fixed in 4% paraformaldehyde followed by dehydration using a
gradient ethanol series (80 and 95%). The section was subsequently
stained with HE at 25°C (10 min). Images were captures on a fully
automatic photomicrography device (magnification ×200; five field
of views; Olympus PM-10AO; Olympus Corporation).
Immunohistochemistry
Immunohistochemical staining was performed using a SABC kit
(Biyuntian Biotechnology Co., Ltd.)
0.3% hydrogen peroxide formaldehyde solution was
added to the paraffin sections and incubated at 37°C. The sections
of breast cancer tissues and paracancerous tissues were washed with
PBS and incubated with 10% bovine serum albumin (Thermo Fisher
Scientific, Inc.) for 15 min. Rabbit anti-human TGF-β1
and TG2 polyclonal antibodies (1:300) were added and incubated at
4°C for 12 h. The next day, color rendering was performed using
3,3′-diaminobenzidine and images were captures on a fully automatic
photomicrography device (magnification ×200; five field of views;
Olympus PM-10AO; Olympus Corporation).
MTT assay
MCF-7 cells were seeded in 96-well plates at
1×103 per well, treated with 1 or 2 mg/l cisplatin
(German Pharmaceutical Co., Ltd.) and incubated in an incubator at
37°C at 5% CO2 for 24 h. Next, 10 µl MTT was added to
each well and incubated for 30 min at 37°C. The absorbance was
measured at 450 nm using a microplate reader.
Western blotting
The protein expression levels of TG2 and
TGF-β1 inMCF-7 cells were assessed by western blotting.
Tissue samples were homogenized in a PIPA buffer (Qiagen, Inc.).
Protein concentrations were determined using a bicinchoninic acid
kit (Pierce; Thermo Fisher Scientific, Inc.). Protein (20 µg/lane)
was separated using 12% SDS-PAGE and transferred to a PVDF
membrane. The membrane was incubated with primary antibodies
against TG2 (1:2,000), TGF-β1 (1:2,000) and β-actin
(1:2,000) at 4°C overnight. The membranes were incubated with
horseradish peroxidase-conjugated anti-rabbit IgG antibody (cat.
no. ZB-2301; 1:2,000; Beijing Noble Technology Co., Ltd). TG2 and
TGF-β1 were detected using ECL development solution
(Pierce; Thermo Fisher Scientific, Inc.). TG2 and TGF-β1
expression levels were determined using Quantity One v4.6.2
software (Bio-Rad Laboratories, Inc.).
Immunofluorescence
The fluorescence intensity of TG2 and
TGF-β1 in MCF-7 cells was assessed via
immunofluorescence. MCF-7 cells (5×104) were treated
with cisplatin (1 mg/l), TGF-β1 (150 µmol/l) and TG2
inhibitors (8.31 µmol/l), and incubated at 37°C and 5%
CO2 for 24 h. Rabbit monoclonal primary antibodies
against TG2 (1:300) and TGF-β1 (1:300) were added to the
cells and incubated overnight at 4°C. Following overnight
incubation with fluorescein-conjugated IgG (cat. no. ZB-2301;
Beijing Noble Technology Co., Ltd.) antibody at 4°C. TG2 and
TGF-β1 expression levels were determined using Quantity
One v4.6.2 software (Bio-Rad Laboratories, Inc.).
Molecular docking
The crystal structure of TG2 [in complex with GTP;
Protein Data Band (PDB) ID, 4PYG] and TGF-β1 (in complex
with scFv GC1009; PDB ID, 4KV5) were obtained from the PDB
(http://www.rcsb.org/pdb). The protein files of
TG2 and TGF-β1 were prepared by removing water molecules
and other ligands. Molecular docking studies and docking analysis
were performed using the PatchDock server (http://bioinfo3d.cs.tau.ac.il/PatchDock/). Analysis
and visualization of interactions in the docked complexes obtained
by PatchDock server were analyzed by PyMOL (https://pymol.en.softonic.com/).
Statistical analysis
Quantitative data are presented as the mean ±
standard deviation and were analyzed using SPSS 19.0 software (IBM
Corp.). Student's t-test was used to compare two groups; one-way
ANOVA followed by Dunnett's test was used to compare all treatment
groups against an untreated control group. The χ2 test
was used to analyze the associations between protein expression and
patient characteristics. P<0.05 was considered to indicate a
statistically significant difference.
Results
HE staining
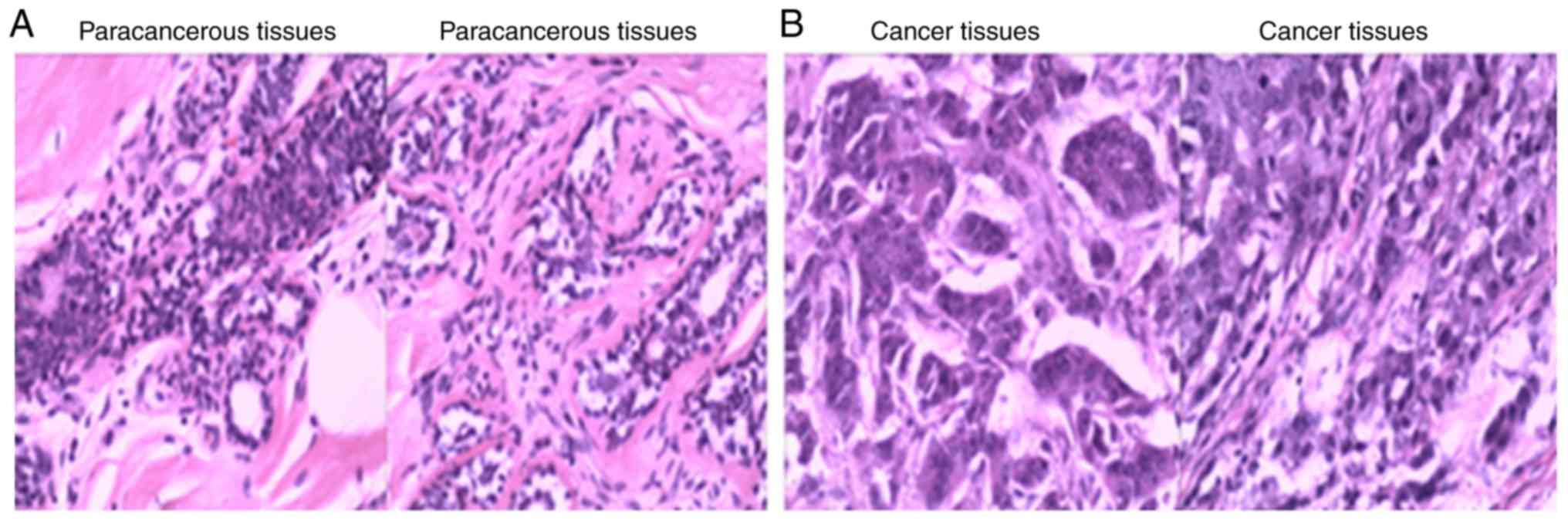
In the present study, tissues from 30 patients with
breast cancer were evaluated using HE staining (Table I). These patients were recruited at
the China-Japan Union Hospital of Jilin University. Clear
structures and contours of the tissues were detected in
paracancerous tissues (Fig. 1A).
However, in the breast cancer samples, moderately heteromorphic and
moderately differentiated cells were observed in the epithelial
tissue (Fig. 1B). In addition,
moderate atypia and interstitial fibrosis were observed in the
breast cancer tissues (Fig. 1B).
 | Table I.Associations between patient
clinicopathological characteristics and the expression of TG2 and
TGF-β1. |
Table I.
Associations between patient
clinicopathological characteristics and the expression of TG2 and
TGF-β1.
| Characteristic | TG2-positive | TG2-negative | P-value |
TGF-β1-positive |
TGF-β1-negative | P-value |
|---|
| Age, years, n |
|
35-45 | 8 | 6 |
| 10 | 5 |
|
|
45-55 | 9 | 7 |
| 10 | 5 |
|
| Stage, n |
| II | 30 |
|
| 30 |
|
|
| Infiltration depth
(%) | 17 (56.7%) | 13 (43.3%) | 0.025 | 20 (66.7%) | 10 (33.3%) | <0.001 |
| Nucleus
and nuclear membrane, n | 15 | 11 |
| 2 | 1 |
|
| Serosa,
n | 2 | 2 |
| 18 | 9 |
|
Immunohistochemical staining
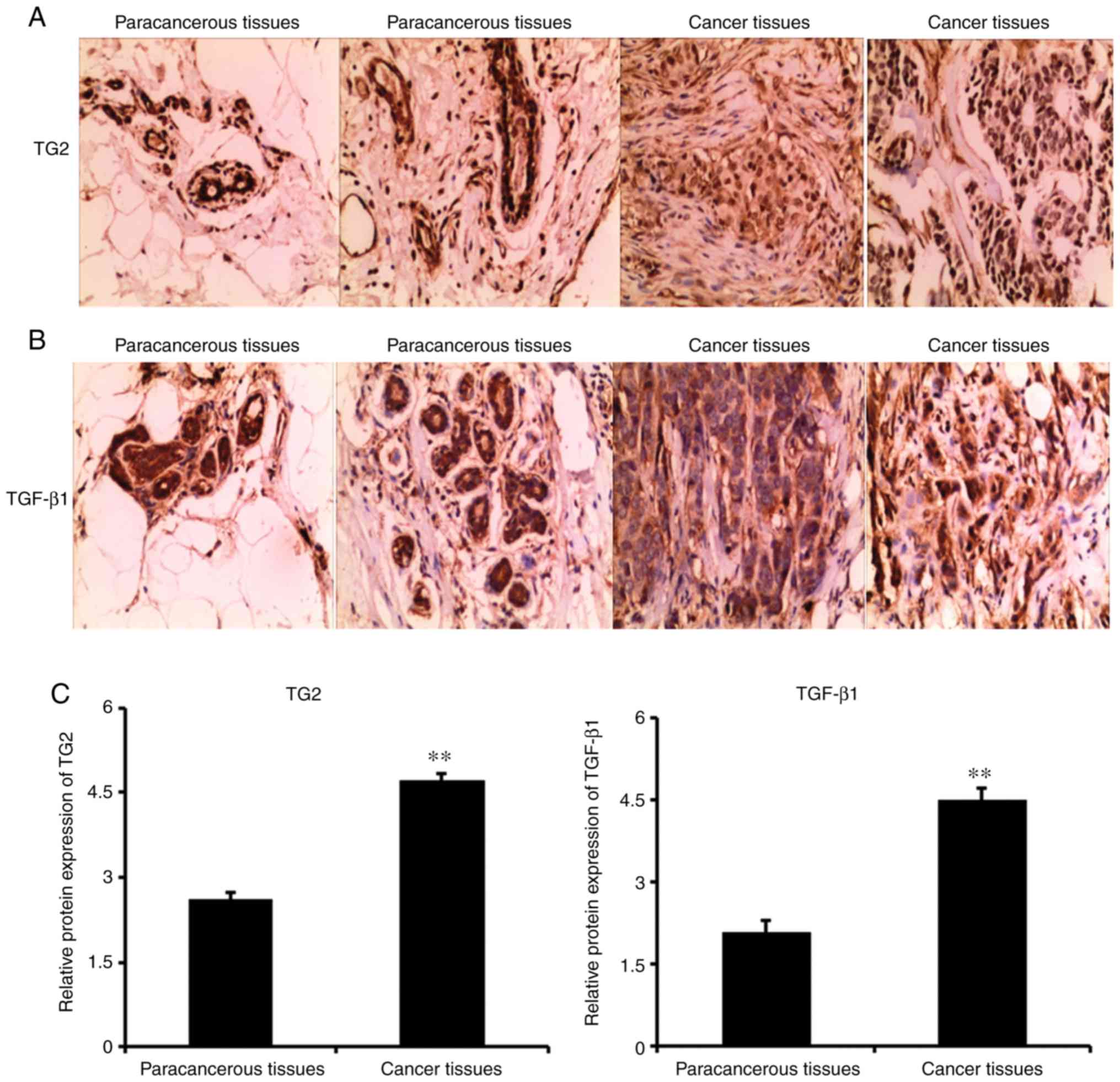
In the present study, the expression levels of TG2
and TGF-β1 were assessed via immunohistochemical
staining. The results revealed that the expression levels of TG2
and TGF-β1 in the cell membrane and cytoplasm in the
paracancerous tissues were low. However, the expression levels of
TG2 and TGF-β1 were significantly higher in the breast
cancer tissues compared with those in the paracancerous tissues
(P<0.05; Fig. 2A-C). TG2 and
TGF-β1 were primarily expressed in the tumor and
interstitial regions.
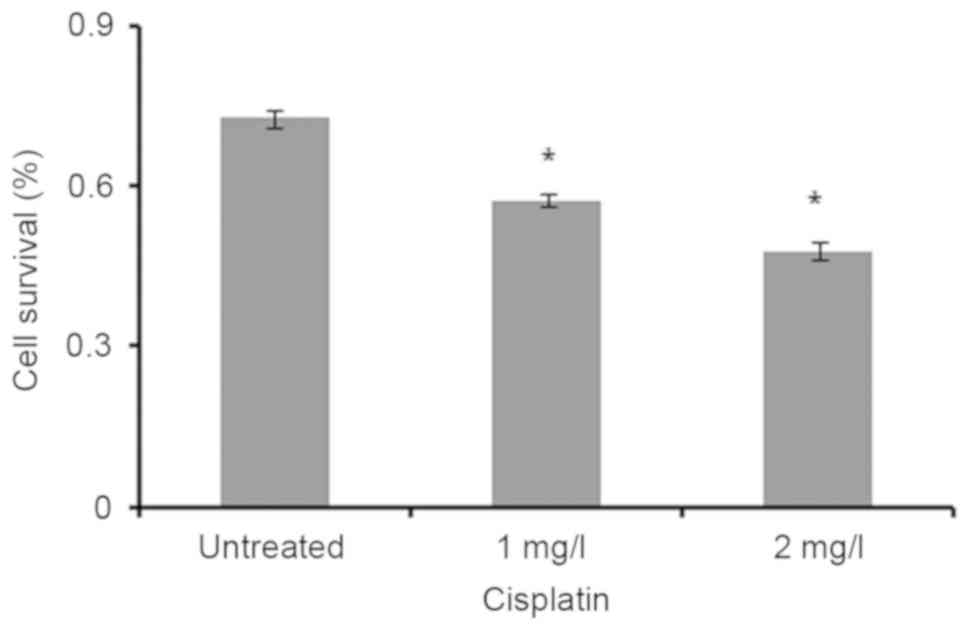
MTT
The viability of MCF-7 cells treated with cisplatin
was analyzed by MTT assay. The results revealed that cell survival
in untreated MCF-7 cells was lower compared with that in cells
treated with 1 and 2 mg/l cisplatin (P<0.05; Fig. 3).
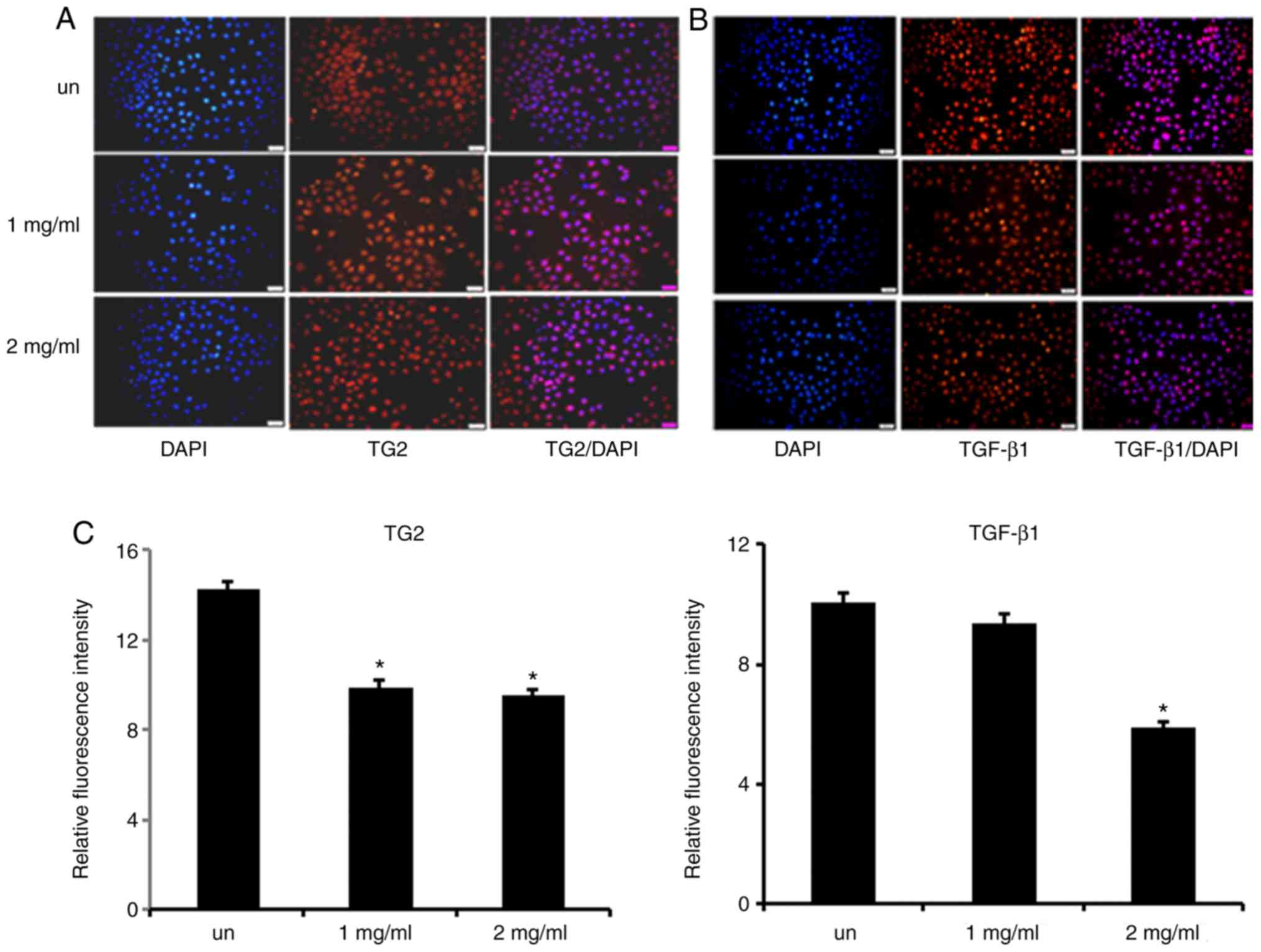
Immunofluorescence
In the present study, immunofluorescence was used to
determine the fluorescence intensity of TG2 and TGF-β1
in MCF-7 cells. The results demonstrated that the fluorescence
intensity of TG2 and TGF-β1 decreased in
cisplatin-treated MCF-7 cells compared with untreated MCF-7 cells
(P<0.05; Fig. 4A-C).
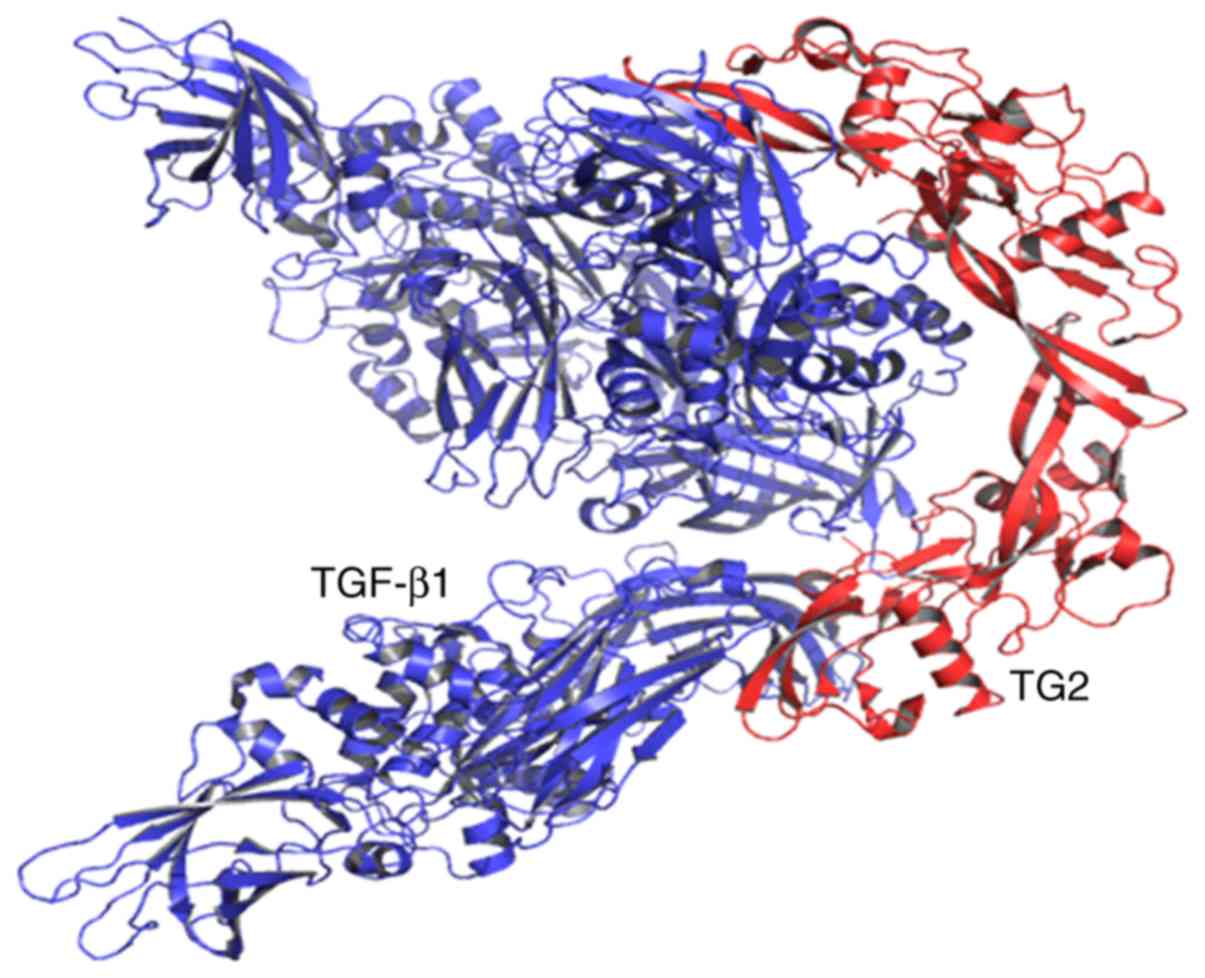
Molecular docking of TG2 with
TGF-β1
In order to further understand and characterize the
interactions between TG2 and TGF-β1, the binding
interaction of TG2 with TGF-β1 was modeled by molecular
docking. The present results found that there were ten sites that
prove TGF-β1 may be associated with TG2 (Table II; Fig.
5).
| Table II.Binding scores of junction for TG2
with TGF-β1. |
Table II.
Binding scores of junction for TG2
with TGF-β1.
| Solution no. | Score | Area (Å) |
|---|
| 1 | 20,950 | 3614.3 |
| 2 | 20,368 | 3214.5 |
| 3 | 20,160 | 3012.9 |
| 4 | 19,952 | 2918.5 |
| 5 | 19,554 | 3492.0 |
| 6 | 19,460 | 3311.2 |
| 7 | 19,428 | 2974.3 |
| 8 | 19,288 | 2760.4 |
| 9 | 19,254 | 2881.1 |
| 10 | 19,096 | 2987.4 |
TG2 and TGF-β1 fluorescence
intensity following TGF-β1 induction and TG2 or
TGF-β1 inhibitors
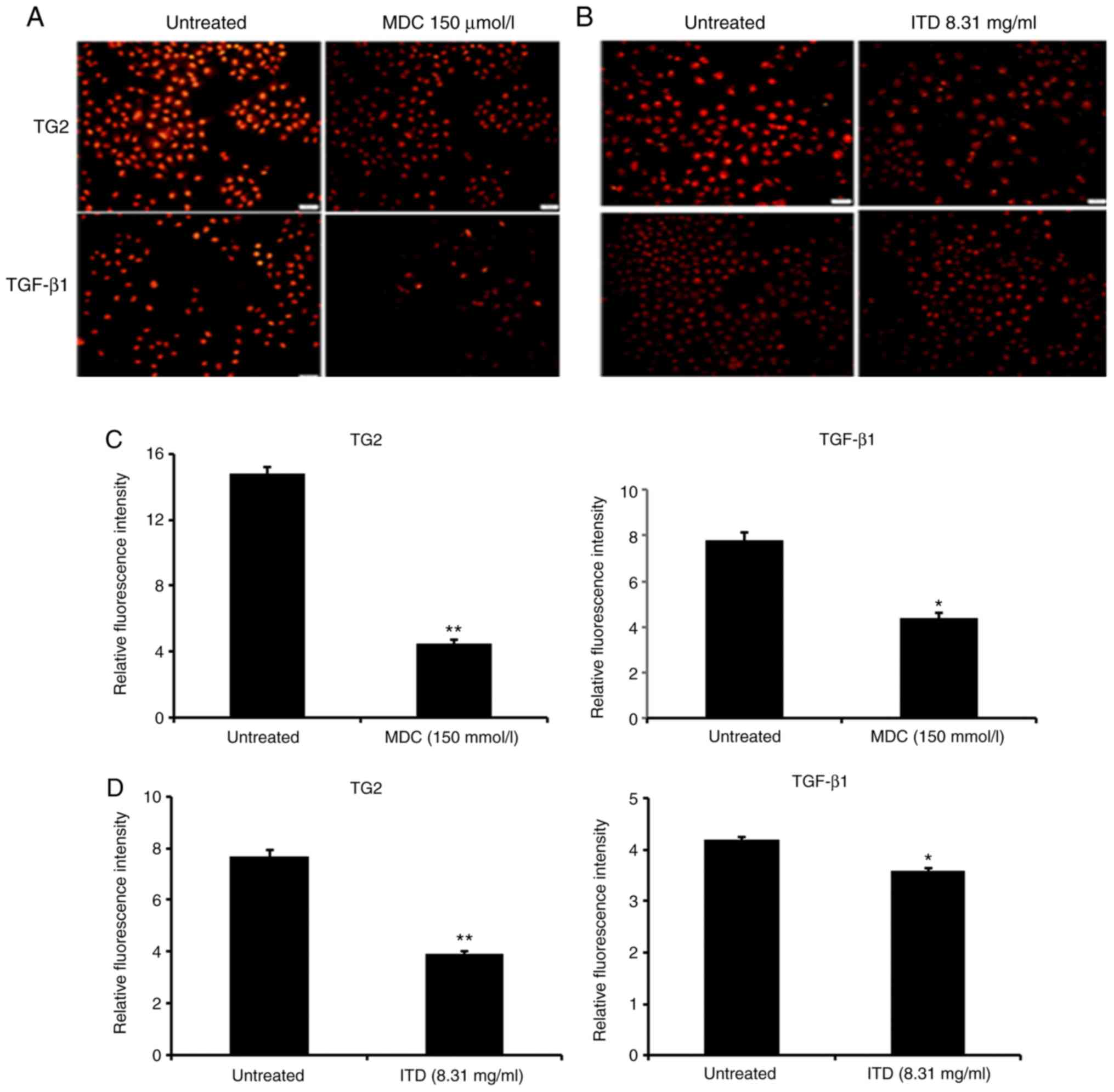
In the present study, the fluorescence intensity of
TG2 and TGF-β1 in MCF-7 cells treated with the
inhibitors of TGF-β1 and TG2 was assessed by
immunofluorescence. The results demonstrated that the fluorescence
intensity of TG2 and TGF-β1 in MCF-7 cells treated with
the TGF-β1 inhibitor (8.31 µmol/l) and the TG2 inhibitor
(150 µmol/l) was lower compared with that in untreated MCF-7 cells
(P<0.05; Fig. 6). In addition,
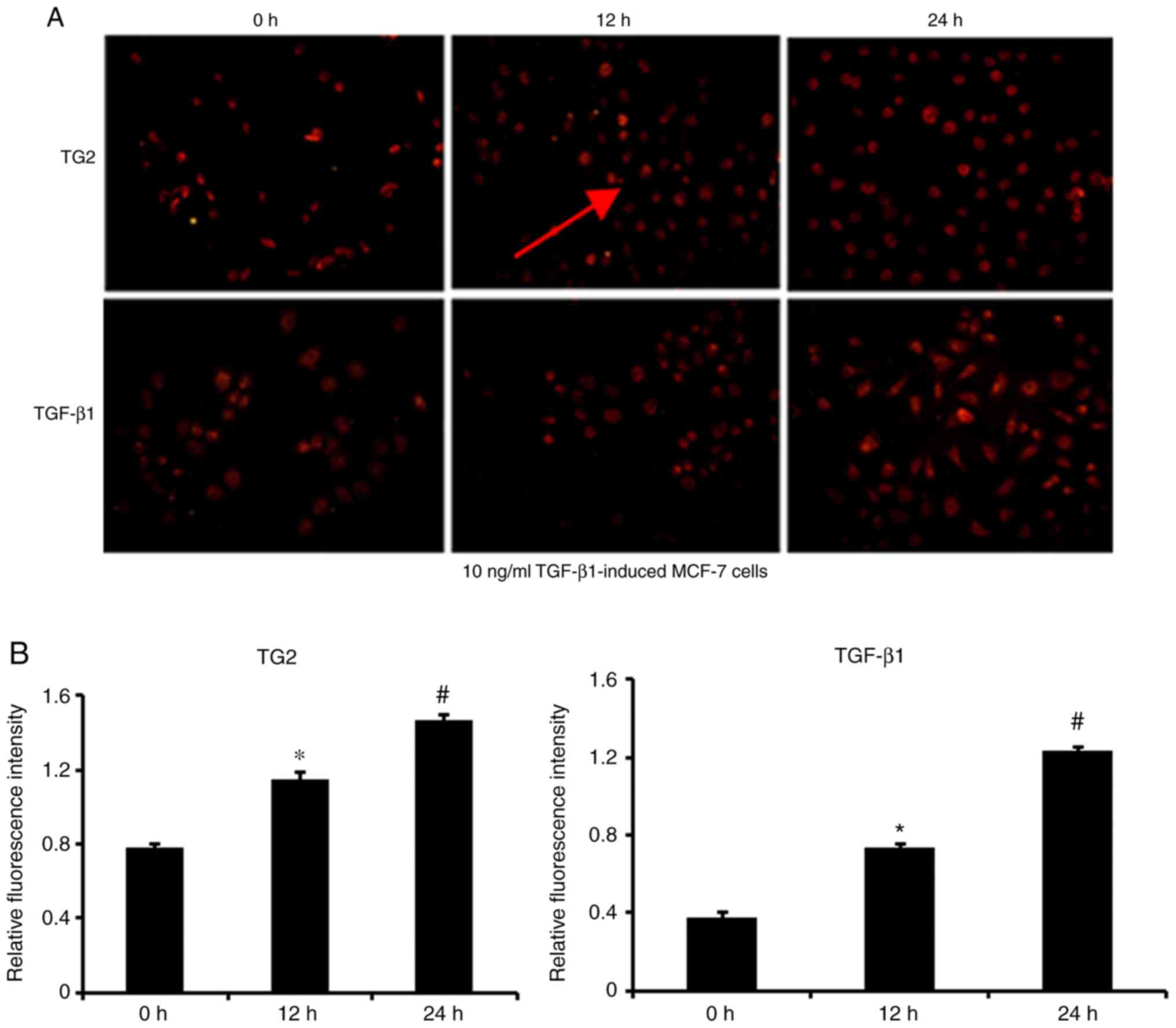
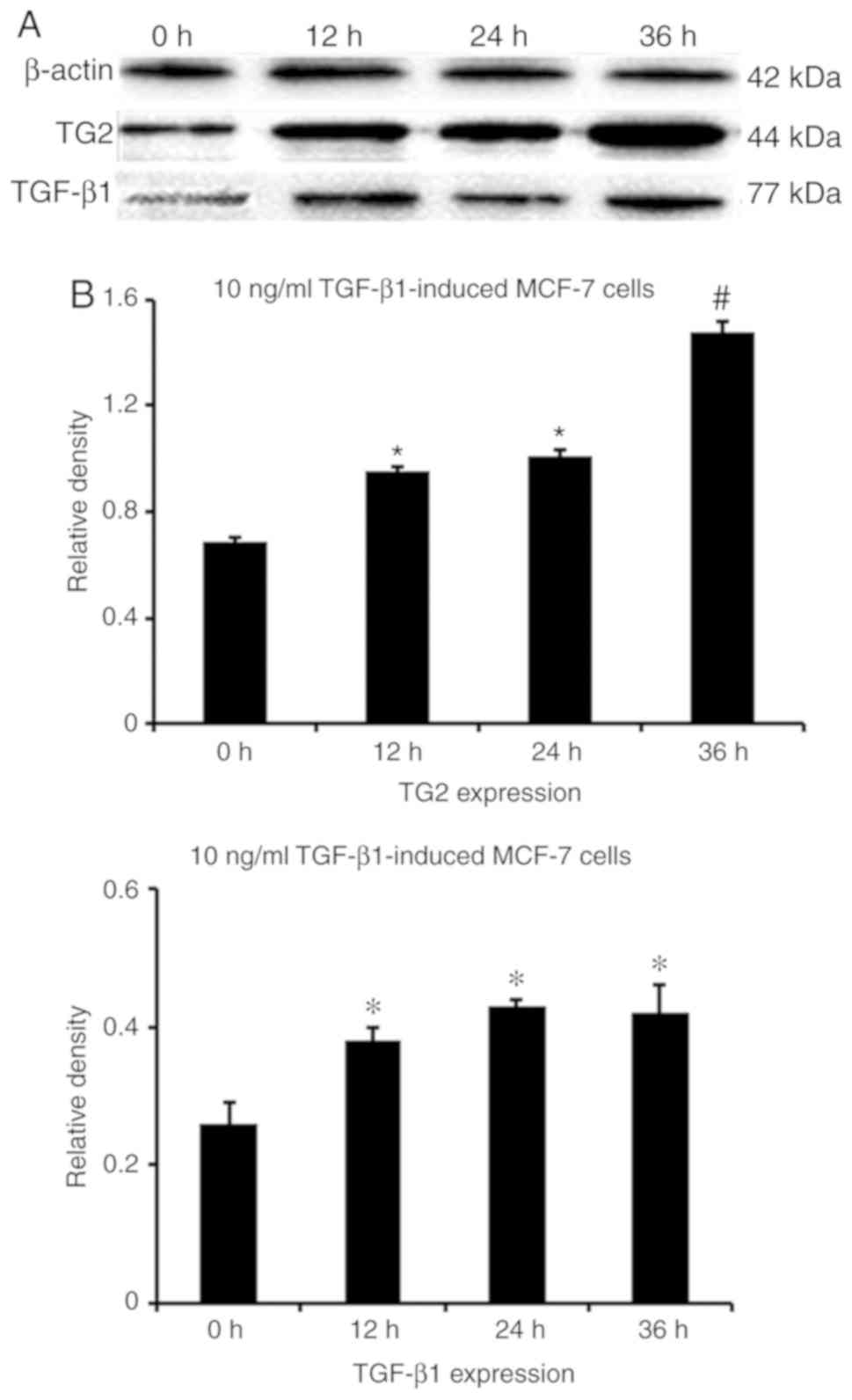
the fluorescence and protein levels of TG2 and TGF-β1 in
MCF-7 cells induced with TGF-β1 were higher compared
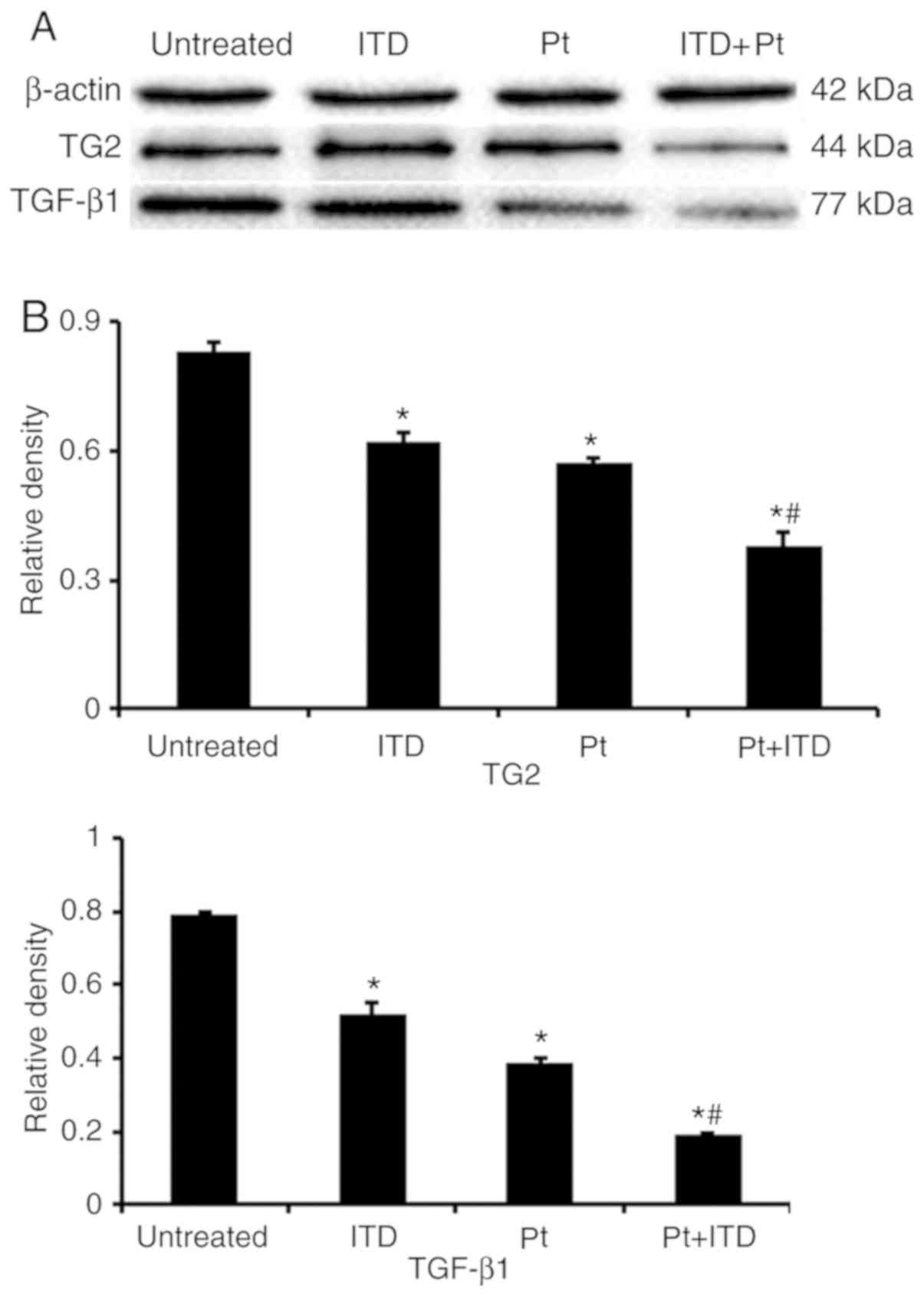
with that in untreated MCF-7 cells (P<0.001; Figs. 7 and 8). In Fig.
9, the expression levels of TG2 and TGF-β1 in MCF-7
cells treated with ITD (TGF-β1), Pt (cisplatin), and ITD
plus Pt were lower compared with those in untreated MCF-7 cells
(P<0.05 and P<0.001).
Discussion
The results of the present study demonstrated that
TG2 and TGF-β1 served an important role in the
development and progression of breast cancer. The protein levels of
TG2 and TGF-β1 were evaluated in MCF-7 cells treated
with cisplatin. In addition, the present study investigated the
role of TG2 and TGF-β1 in MCF-7 cells treated with
inhibitors of TGF-β1 and TG2.
TG2 is involved in the occurrence and development of
Parkinson's disease (10,11). A previous study have demonstrated the
association of TG2 with tumor growth, EMT and metastasis (12). TG2 also exhibits
Ca2+-independent guanosine triphosphate hydrolase,
protein kinase and disulfide isomerase activity (13). TG2 knockdown decreases the
proliferative, migratory and colony forming abilities of breast
cancer cells (14). With the
occurrence and development of cancer cachexia, the role of the
TGF-β signaling pathway changes, inducing the EMT process and
promoting tumor invasion and metastasis (15). It has been demonstrated that the
level of the TGF-β signaling pathway activity is associated with
lung fibrosis and patient mortality (16). The results of the present study
revealed that the expression levels of TG2 and TGF-β1 in
human breast cancer were higher compared with those in
paracancerous tissues. These results suggested that
TGF-β1 and TG2 may serve an important role in the
occurrence and development of breast cancer.
To further analyze the role of TGF-β1 and
TG2 in breast cancer, the present study analyzed the expression
levels of TGF-β1 and TG2 in MCF-7 cells treated with
cisplatin. Cisplatin is a selective drug used for the treatment of
prostate cancer (17). A large
proportion of patients develop resistance to cisplatin, inducing
tumor relapse and limiting its clinical usefulness (17). In the present study, the results
demonstrated that the fluorescence intensity of TG2 and
TGF-β1 in untreated MCF-7 cells was higher compared with
that in MCF-7 cells treated with cisplatin. These results suggested
that cisplatin may exert its chemotherapeutic effects in MCF-7
cells by regulating the levels of TG2 and TGF-β1
protein.
TG2 was associated with TGF-β1 in the
occurrence and development of various diseases (8). To clarify the effect of TG2 and
TGF-β1 in MCF-7 cells, the present study investigated
the effect of TG2 and TGF-β1 in the MCF-7 cells treated
with inhibitors of TGF-β1 and TG2. The results
demonstrated that the fluorescence intensity of TG2 and
TGF-β1 in MCF-7 cells treated with a TG2 or
TGF-β1 inhibitor was lower compared with that in
untreated MCF-7 cells. By contrast, the fluorescence intensity of
TG2 and TGF-β1 in MCF-7 cells treated with
TGF-β1 was higher compared with that in untreated MCF-7
cells. Therefore, TG2 and TGF-β1 may serve a role in the
occurrence and development of breast cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
Science and Technology Department of Jilin Province Project (grant
no. 20170204024YY) and the Health Department Project of Jilin
Province (grant no. 2016Q028).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HCG conceived the study and drafted the manuscript.
YC and ZHW acquired the data. YZH, YQL and GHY analyzed the data
and revised the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
China-Japan Union Hospital (Changchun, China; approval no.
2018120506). Signed informed consents were obtained from the
patients and/or the guardians.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhu W, Harvey S, Macura KJ, Euhus DM and
Aremov D: Invasive breast cancer preferably and predominantly
occurs at the interface between fibroglandular and adipose tissue.
Clin Breast Cancer. 17:e11–e18. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Alipour S, Jannat F and Hosseini L:
Teaching breast cancer screening via text messages as part of
continuing education for working nurses: A case-control study.
Asian Pac J Cancer Prev. 15:5607–5609. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Agnihotri N, Kumar S and Mehta K: Tissue
transglutaminase as a central mediator in inflammation-induced
progression of breast cancer. Breast Cancer Res. 15:2022013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang J, Xi C, Yang X, Lu X, Yu K, Zhang Y
and Gao R: LncRNA WT1-AS inhibits triple-negative breast cancer
cell migration and invasion by downregulating transforming growth
factor-β1. Cancer Biother Radiopharm. 34:671–675. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Takai E, Tsukimoto M and Kojima S:
TGF-β1 downregulates COX-2 expression leading to
decrease of PGE2 production in human lung cancer A549 cells, which
is involved in fibrotic response to TGF-β1. PLoS One.
8:e763462013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shao M, Cao L, Shen C, Satpathy M,
Chelladurai B, Bigsby RM, Nakshatri H and Matei D:
Epithelial-to-mesenchymal transition and ovarian tumor progression
induced by tissue transglutaminase. Cancer Res. 69:9192–9120. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Humbert L, Ghozlan M, Canaff L, Tian J and
Lebrun JJ: The leukemia inhibitory factor(LIF) and p21 mediate the
TGFβ tumor suppressive effects in human cutaneous melanoma. BMC
Cancer. 15:2002015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cao L, Shao M, Schilder J, Guise T,
Mohammad KS and Matei D: Tissue transglutaminase links TGF-β,
epithelial to mesenchymal transition and a stem cell phenotype in
ovarian cancer. Oncogene. 31:2521–2534. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Efferth T, Konkimalla VB, Wang YF,
Sauerbrey A, Meinhardt S, Zintl F, Mattern J and Volm M: Prediction
of broad spectrum resistanc-e of tumors towards anticancer drugs.
Clin Cancer Res. 14:2405–2412. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Segers-Nolten IM, Wilhelmus MM, Veldhuis
G, van Rooijen BD, Drukarch B and Subramaniam V: Tissue
transglutaminase modulates alpha-synuclein oligomeri-zation.
Protein Sci. 17:1395–1402. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schmid AW, Chiappe D, Pignat V, Grimminger
V, Hang I, Moniatte M and Lashuel HA: Dissecting the mechanisms of
tissue transglutaminase-induced cross-linkingof alpha-synuclein:
Implications for the pathogenesis of parkinson disease. J BiolChem.
284:13128–13142. 2009.
|
|
12
|
Assi J, Srivastava G, Matta A, Chang MC,
Walfish PG and Ralhan R: Transglutaminase 2 overexpression in tumor
stroma identifies invasive ductal carcinomas of breast at high risk
of recurrence. PLoS One. 8:e744372013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Király R, Thangaraju K, Nagy Z, Collighan
R, Nemes Z, Griffin M and Fésüs L: Isopeptidase activity of human
transglutaminase 2: Disconnection from transamidation and
characterization by kinetic parameters. Amino Acids. 48:31–40.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhao G, Zhang ZQ, Zhang B, Luo M, Sun YW
and Wu ZY: Down-Regulation of TG2 expression by RNAi inhibits HSC
proliferation and attenuates liver fibrosis. Int J Clin Exp Pathol.
4:513–520. 2011.PubMed/NCBI
|
|
15
|
Alves MJ, Figuerêdo RG, Azevedo FF,
Cavallaro DA, Neto NI, Lima JD, Matos-Neto E, Radloff K, Riccardi
DM, Camargo RG, et al: Adipose tissue fibrosis in human cancer
cachexia: The role of TGF-β pathway. BMC Cancer. 17:1902017.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang YP, Cao SY, Ma JY, Wang LQ and Wang
BF: The change and significance of coagulation activity in
bleomycin-induced lung fibrosis in rats. Zhonghua Jie He He Hu Xi
Za Zhi. 28:541–544. 2005.(In Chinese). PubMed/NCBI
|
|
17
|
Vesprini D, Narod SA, Trachtenberg J,
Crook J, Jalali F, Preiner J, Sridhar S and Bristow RG: The
therapeutic ratio is preserved for radiotherapy or cisplatin
treatment in BRCA2-mutated prostate cancers. Can Urol Assoc J.
5:E31–E35. 2011. View Article : Google Scholar : PubMed/NCBI
|