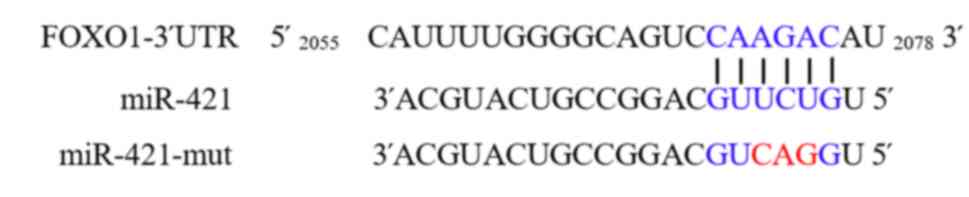Introduction
Lung cancer is the leading cause of
cancer-associated mortality worldwide (1), accounting for more deaths than breast,
prostate and colon cancer combined (2–4).
Although numerous studies have investigated the role of microRNAs
(miRNAs/miRs) in patients with lung cancer, there has been
relatively little progress in developing novel therapeutic
strategies (5–8).
miRNAs are 20–22 nucleotides in length and serve as
negative regulators of gene expression by binding to the
complementary 3′-untranslated regions (UTRs) of their target genes
(9). Previous studies have revealed
that miRNAs participate in specific biological processes, including
the cell cycle, aging and death (10,11), and
serve as hallmarks of several pathological conditions, including
neoplasms (12,13). It has been hypothesized that miRNAs
may serve critical roles in the progression of cancer, cancer
metastasis, angiogenesis (14) and
drug resistance (15).
Numerous studies have revealed that miR-421 may be
associated with several types of cancer, including neuroblastoma
(16) and gastric cancer (17,18). It
has been revealed that miR-421 enhanced the proliferation and
metastasis of gastric cancer cells by targeting claudin-11
(19). In addition, miR-421 induced
cisplatin resistance by targeting E-cadherin and caspase-3
(20), and miR-421 promoted breast
cancer progression by inhibiting caspase-10 (21). In lung cancer, miRNA sequencing
revealed that the expression levels of various miRNAs were
aberrantly upregulated, including miR-421 (22); however, the effect and mechanism of
miR-421 in lung cancer cells remains unclear.
Several signaling pathways have important roles in
cancer; for example, the AKT/glycogen synthase kinase (GSK)-3β
pathway was discovered to regulate tumor cell proliferation and
apoptosis (23). Upon activation of
this pathway, various downstream molecules associated with the cell
cycle, including forkhead box O1 (FOXO1), p21, retinoblastoma (Rb)
and cyclin D, are regulated, resulting in cell proliferation
(24,25). As a member of the FOX family, FOXO1
is a transcription factor that has been reported to serve a role in
apoptosis and cancer development (26). FOXO1 has also been suggested to serve
important roles in cell cycle regulation, apoptosis, proliferation
and immune responses in various types of cancer (27).
The current study aimed to determine whether miR-421
modulated the viability of lung cancer cells by targeting FOXO1 and
various other downstream molecules. The expression levels of
miR-421 were analyzed in seven different lung cancer cell lines,
the target gene of miR-421 was identified and verified, and the
effect of miR-421 on the viability of lung cancer cells was
determined. In addition, the potential mechanisms of action were
investigated. The results of the current study may provide novel
insights into the role of miR-421 in lung cancer and its possible
molecular mechanisms.
Materials and methods
Cell lines and reagents
Seven lung cancer cell lines (A549; hLAMP, cat. no.
XY-XB-2164; Calu-3; NCI-H1975; NCI-H1299; NCI-H1650; and NCI-H460)
and human bronchial epithelial cell line (BEAS-2B) were purchased
from Shanghai Oulu Biological Technology Co., Ltd. Moloney murine
leukemia virus reverse transcriptase (MMLV-RT) was obtained from
Toyobo Life Science. Anti-FOXO1 (cat. no. ab52874) was purchased
from Abcam. The following primary antibodies were purchased from
Santa Cruz Biotechnology, Inc.: anti-p21 (cat. no. sc-6246),
anti-phosphorylated (p)-AKT (cat. no. sc-81433), anti-p-GSK-3β
(cat. no. sc-81496), anti-p-Rb (cat. no. sc-377528), anti-cyclin D1
(cat. no. sc-8396), anti-Rb (cat. no. sc-102), anti-AKT (cat. no.
sc-81434), anti-GSK3β (cat. no. sc-81462) and anti-β-actin (cat.
no. sc-81178). The horseradish peroxidase (HRP)-conjugated rabbit
anti-mouse antibody (cat. no sc-358914) was also purchased from
Santa Cruz Biotechnology, Inc. LightSwitch™ luciferase assay
reagents were obtained from Promega Corporation. miR-421-mimics,
miR-421-inhibitors and their negative controls (NCs) were purchased
from Guangzhou RiboBio Co., Ltd.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from A549, hLAMP, Calu-3,
NCI-H1975, NCI-H1299, NCI-H1650, NCI-H460 and BEAS-2B cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). Total RNA was reverse transcribed into cDNA using MMLV-RT at
42°C for 50 min. qPCR was subsequently performed to detect the
expression levels of miR-421 and SYBR Green I was provided by
Thermo Fisher Scientific, Inc.[Mo, 2014 #10303]. Each PCR cycle
involved denaturation (95°C, 30 sec), annealing (60°C, 30 sec) and
extension (72°C, 30 sec) for 40 cycles. The specific primers for
miR-421 and small nuclear RNA U6 were designed by Guangzhou RiboBio
Co. Ltd. as follows: miR-421, forward 5′-GGCCGCGATCAACAGACAT-3′,
reverse 5′-CCAGTGCAGGGTCCGAGGTA-3′; and small nuclear RNA U6,
forward 5′-TGGCACCCAGCACAATGAA-3′ and reverse
5′-CTAAGTCATAGTCCGCCTAGAAGCA-3′. Expression levels were quantified
using the 2−ΔΔCq method (28). Small nuclear RNA U6 was used as the
loading control and for normalization.
Bioinformatics analysis
The target gene of miR-421 was predicted using
TargetScan 7.2 software (http://www.targetscan.org/vert_72; Whitehead Institute
for Biomedical Research).
Cell transfection
A549 cells (3×105) were transfected with
100 nmol miR-421-mimics, miR-421-inhibitors or their NCs (Guangzhou
RiboBio Co., Ltd.) using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. After 48 h, transfected cells were
subsequently used for the MTT assay and western blotting.
Luciferase reporter assay
The miR-421-mimics-NC, miR-421-mimics,
miR-421-mutant (mut, synthesized by Guangzhou RiboBio Co., Ltd) or
miR-421-inhibitor were co-transfected at 37°C using Lipofectamine
2000 into 5×105 A549 cells with the FOXO1-3′-UTR
(inserted into the pGL3-control luciferase reporter plasmid;
Promega Corporation) for 24 h. Luciferase assays were performed
using the LightSwitch™ reagents according to the manufacturer's
protocol. The relative firefly and Renilla luciferase
activities were determined using a luminometer (Promega
Corporation). The relative luciferase activity was calculated as
relative light units (Renilla luciferase/firefly luciferase)
to determine whether FOXO1 was a target gene of miR-421.
MTT assay
Following transfection, A549 cells
(1×104) were cultured at 37°C in 96-well plates. After
1, 2, 3, 4 and 5 days of incubation, an MTT assay was performed to
detect the viability of A549 cells as described by van Tonder et
al (29). Briefly, following the
incubation, 20 µl MTT solution (5 mg/ml) was added to each well and
incubated at 37°C for 4 h. The medium was subsequently aspirated
and 150 µl DMSO was added to each well. The plates were placed on a
shaker for 10 min and the absorbance was then determined at 570 nm
using a reference wavelength of 630 nm on a microplate reader
(Thermo Fisher Scientific, Inc.).
Colony formation assay
A549 cells transfected with miR-421-mimics or
miR-421 mimics-NC were digested into single cells using 0.25%
trypsin and plated at a density of 200 cells/well into
6-well-culture plates, which were incubated with 5% CO2
at 37°C for 7–14 days. When a colony was observed, the supernatant
was discarded and the cell culture was terminated. Subsequently,
samples were washed three times with PBS and fixed with 1 ml 4%
methanol for 10 min at room temperature. The colonies were
subsequently stained with 0.1% hematin for 10 min at room
temperature and washed with distilled water. Colonies including
>50 cells were counted under a microscope (IX71; Olympus
Corporation).
Western blotting
The expression levels of FOXO1, AKT, p-AKT, GSK-3β,
p-GSK-3β, p21, Rb, p-Rb and cyclin D1 in transfected cells were
analyzed using western blotting. Briefly, total protein was
extracted from cells using RIPA lysis buffer (Sigma-Aldrich; Merck
KGaA). Total protein was quantified using a bicinchoninic acid
protein assay kit and 20 µg protein/lane was separated via 12%
SDS-PAGE. The separated proteins were subsequently transferred onto
PVDF membranes and blocked with 5% skimmed milk at room temperature
for 2 h. The membranes were then incubated with the primary
antibodies (1:3,000) at 4°C overnight. Following the primary
antibody incubation, the membranes were incubated with the
HRP-conjugated secondary antibody (1:6,000) at room temperature for
2 h. Protein expression was quantitatively assessed using a HRP-ECL
scanner (Lenovo, Beijing, People's Republic of China).
Statistical analysis
Statistical analysis was performed using SPSS 15.0
software (SPSS Inc.). Three independent experiments were performed
and data were presented as the means ± SD. An unpaired Student's
t-test was used to determine the statistical differences between
two groups, whereas a one-way ANOVA followed by Tukey's post hoc
test was performed to determine statistical differences between
>2 groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
miR-421 expression levels are
upregulated in lung cancer cell lines
Compared with the control cell line BEAS-2B, miR-421
expression levels were significantly upregulated in the lung cancer
cell lines (P<0.01), particularly in A549 (11.19±0.85), hLAMP
(15.38±1.24), NCI-H1650 (16.41±2.97) and NCI-H460 (15.44±0.94)
cells (Table I). Among the
aforementioned lung cancer cell lines, miR-421 expression levels in
the A549 cells were neither the highest nor the lowest. For this
reason, A549 cells were selected for further experimentation.
 | Table I.MicroRNA-421 expression levels in lung
cancer cell lines, as determined using reverse
transcription-quantitative PCR. |
Table I.
MicroRNA-421 expression levels in lung
cancer cell lines, as determined using reverse
transcription-quantitative PCR.
| Cell line | 2−ΔΔCq
value |
|---|
| BEAS-2B | 1.00±0.00 |
| A549 |
11.19±0.85a |
| hLAMP |
15.38±1.24a |
| Calu-3 | 6.12
±0.86a |
| NCl-H1975 |
5.57±0.12a |
| NCl-H1299 |
8.05±1.07a |
| NCl-H1650 |
16.41±2.97a |
| NCl-H460 |
15.44±0.94a |
FOXO1 is a target gene of miR-421
The results of the bioinformatics analysis using
TargetScan revealed a complementary binding site between miR-421
and the 3′-UTR of FOXO1 (Fig. 1).
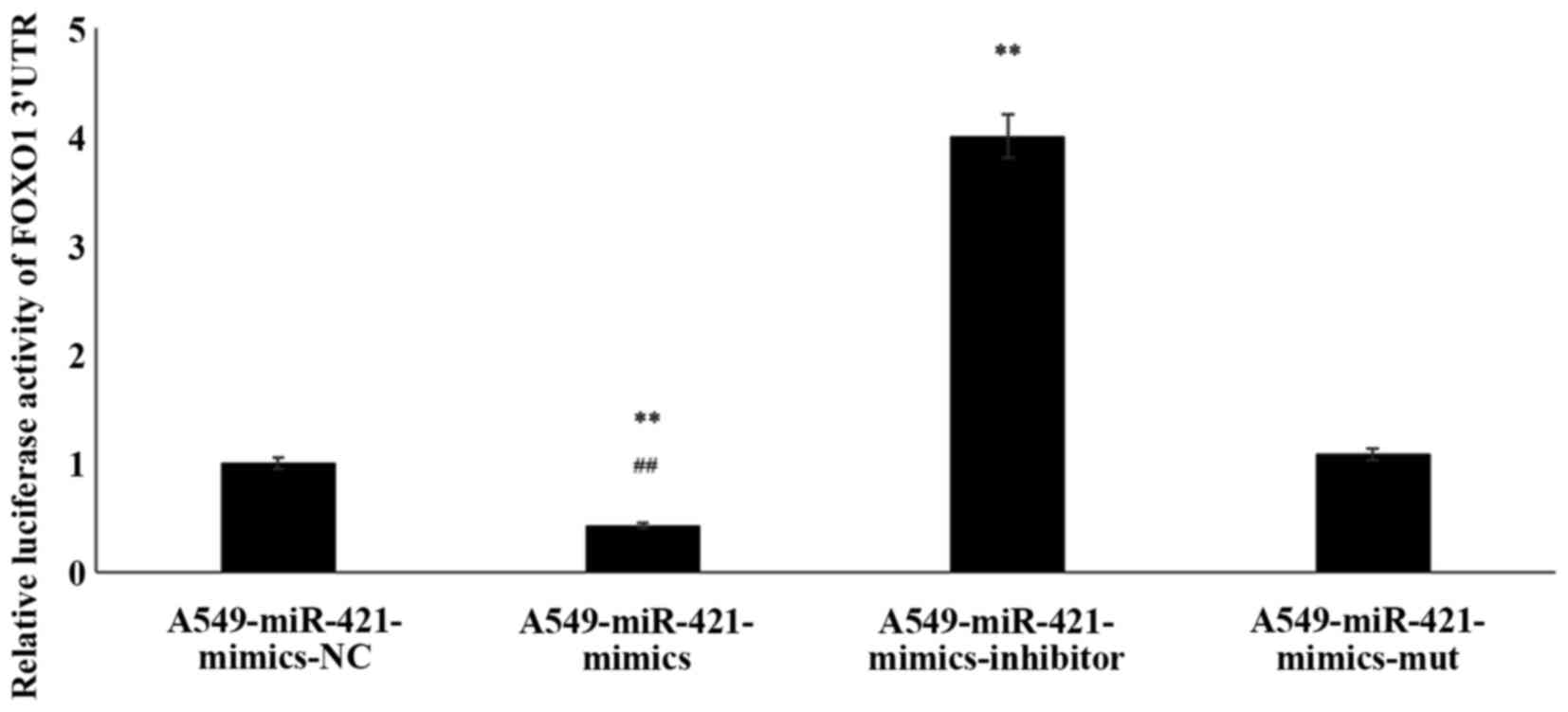
Thus, to verify the interaction between FOXO1 and miR-421, a
luciferase assay was performed. Compared with the
A549-miR-421-mimics-NC group, in the A549-miR-421-mimics group, the
relative luciferase activity of the FOXO1 3′-UTR was significantly
decreased (1.00±0.00 vs. 0.43±0.03; P<0.01; Fig. 2; Table
II). Post-transfection with miR-421-mut, the relative
luciferase activity of the FOXO1 3′-UTR remained at similar levels
compared with the A549-miR-421-mimics-NC group (1.08±0.01 vs.
1.00±0.00; P=0.71). Meanwhile, in the A549-miR-421-inhibitor group,
the relative luciferase activity of the FOXO1 3′-UTR were
significantly increased compared with in the A549-miR-421-mimics-NC
group (4.01±0.07 vs. 1.00±0.00; P<0.01). Thus, the results of
the TargetScan analysis and luciferase reporter assay indicated
that FOXO1 may be a target gene of miR-421.
| Table II.Verification of the miR-421 target
gene FOXO1 using a luciferase reporter assay. |
Table II.
Verification of the miR-421 target
gene FOXO1 using a luciferase reporter assay.
| Group | RLuc/FLuc |
|---|
|
A549-miR-421-mimics-negative control | 1.00±0.00 |
|
A549-miR-421-mimics |
0.43±0.03a |
| A549-
miR-421-inhibitor |
4.01±0.07a |
|
A549-miR-421-mutant | 1.08±0.01 |
miR-421 increases A549 cell
viability
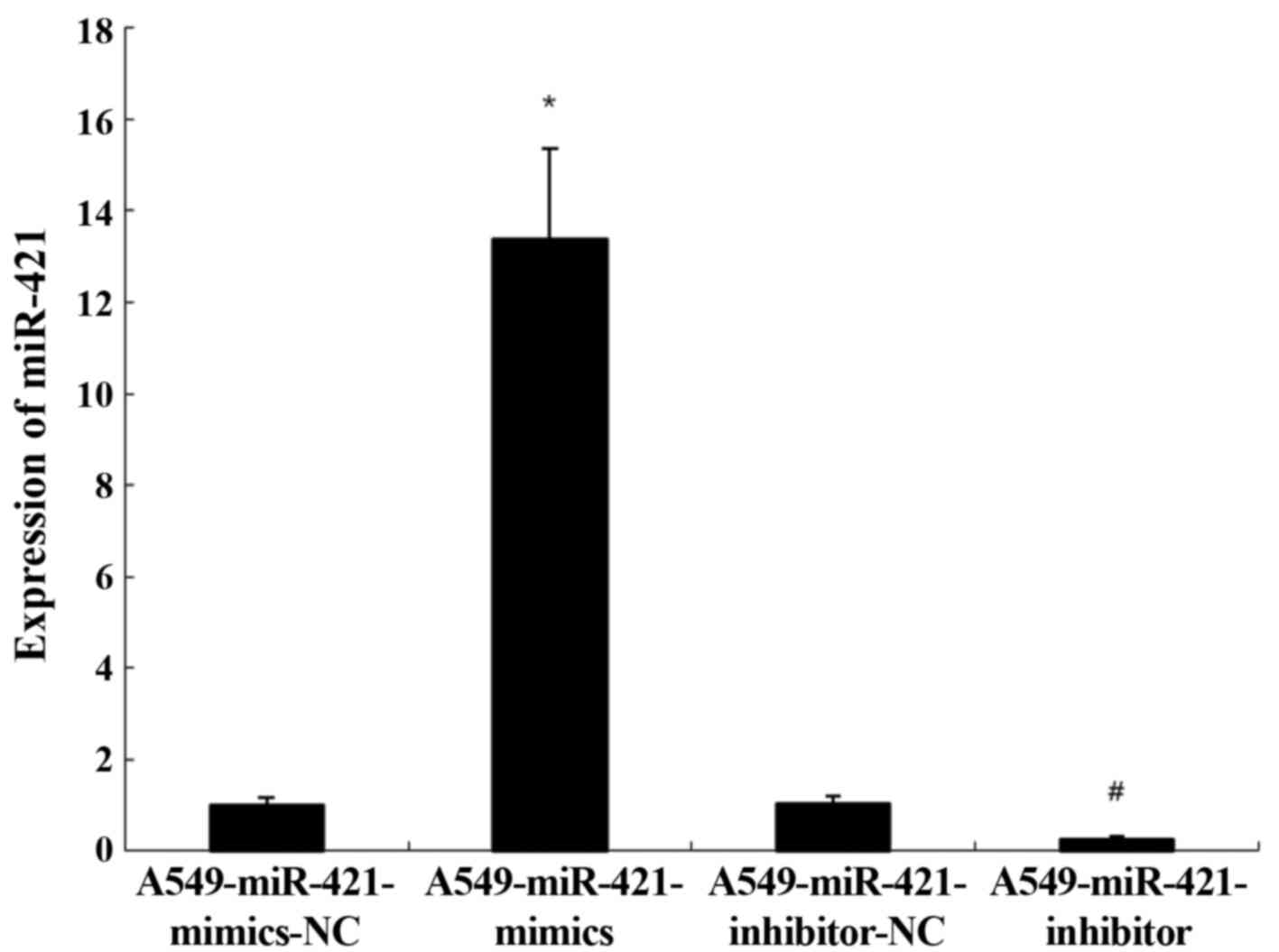
To determine whether transfection with the
miR-421-mimics and inhibitor was successful, RT-qPCR was performed.
In cells transfected with the miR-421-mimics, the expression levels
of miR-421 were significantly upregulated compared with the
A549-miR-421-mimics-NC group (13.37±1.21 vs. 1.00±0.00; P<0.05;
Fig. 3). Conversely, in cells
transfected with the miR-421-inhibitor, the expression levels of
miR-421 were significantly downregulated compared with the
A549-miR-421-inhibitors-NC group (0.26±0.05 vs. 1.03±0.03;
P<0.05; Fig. 3)
The viability of the A549-miR-421-mimics group was
significantly increased compared with in the miR-421-mimics-NC
group [optical density (OD)570 5.72±0.02 vs. 2.97±0.04;
P<0.01]; Table III at 72 h
post-transfection. By contrast, in the A549-miR-421-inhibitor
group, cell viability was significantly decreased compared with in
the A549-miR-421 inhibitor NC group (OD570 1.64±0.01 vs.
3.02±0.04; P<0.01) at 72 h.
| Table III.A549 cell viability following
transfection with miR-421-mimics or miR-421-inhibitor. |
Table III.
A549 cell viability following
transfection with miR-421-mimics or miR-421-inhibitor.
| Time, day |
A549-miR-421-mimics-NC,
OD570 |
A549-miR-421-mimics, OD570 |
A549-miR-421-inhibitor-NC,
OD570 |
A549-miR-421-inhibitor,
OD570 |
|---|
| 0 | 0.02±0.000 | 0.03±0.00 | 0.01±0.00 | 0.03±0.00 |
| 1 | 1.39±0.004 |
1.57±0.01a | 1.67±0.01 |
1.11±0.01b |
| 2 | 2.54±0.010 |
3.94±0.04a | 2.09±0.01 |
1.31±0.01b |
| 3 | 2.97±0.040 |
5.72±0.02a | 3.02±0.04 |
1.64±0.01b |
| 4 | 5.70±0.030 |
10.81±0.01a | 5.94±0.15 |
1.99±0.01b |
| 5 | 8.10±0.030 |
17.79±0.01a | 8.06±0.04 |
4.98±0.02b |
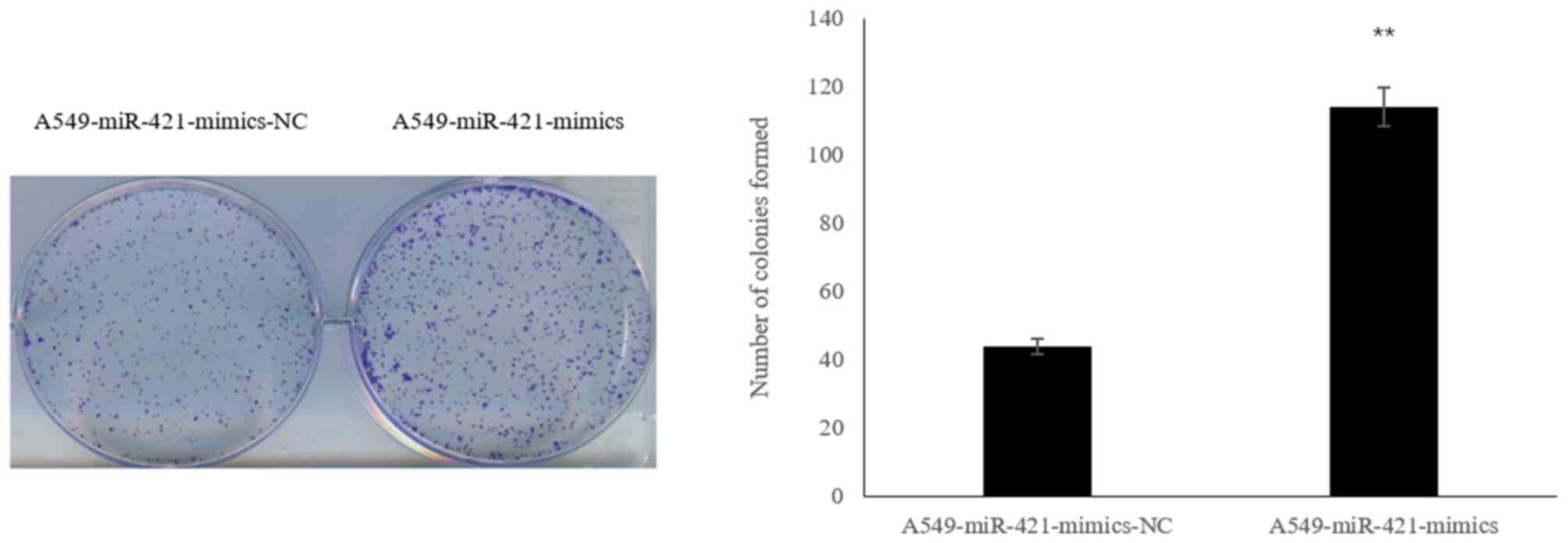
Following transfection of A549 cells with the
miR-421-mimics, the number of colonies formed was significantly
increased compared with in the A549-miR-421-mimics-NC group
(113.96±7.22 vs. 44.07±5.10; P<0.01; Fig. 4).
miR-421 downregulates FOXO1 expression
levels
To investigate whether miR-421 affected the
expression levels of FOXO1, western blotting was performed.
Compared with in the A549-miR-421-mimics-NC group, the expression
levels of FOXO1 and p21 were downregulated, whereas the expression
levels of p-AKT, p-GSK-3β, p-Rb and cyclin D1 were upregulated in
the A549-miR-421-mimics group (P<0.05 Fig. 5). Compared with the
A549-miR-421-inhibitor-NC group, the expression levels of FOXO1 and
p21 were upregulated, whereas the expression levels of p-AKT,
p-GSK-3β, p-Rb and cyclin D1 were downregulated in the
A549-miR-421-mimics group (P<0.05 Fig. 5). These results indicated that
miR-421 may downregulate the expression levels of FOXO1 and p21,
and upregulate p-AKT, p-GSK-3β, p-Rb and cyclin D1 expression
levels in A549 cells.
 | Figure 5.Western blot analysis of protein
expression levels involved in the AKT/GSK-3β pathway. In A549 cells
transfected with the miR-421-mimics, the expression levels of FOXO1
and p21 were downregulated, whereas the expression levels of cyclin
D1 and the ratios p-AKT/AKT, p-GSK-3β/GSK-3β and p-Rb/Rb were
increased. For a, c, e, g, I and k:, P<0.05 vs.
A549-miR-421-mimics-NC. For b, d, f, h, j and : P<0.05 vs.
A549-miR-421-inhibitor-NC. miR, microRNA; NC, negative control;
FOXO1, forkhead box O1; p-, phosphorylated; GSK-3β, glycogen
synthase kinase 3β; Rb, retinoblastoma. |
Discussion
The present study demonstrated that miR-421 miR-421
promoted the viability of A549 lung cancer cells by targeting
FOXO1. Previous studies reported that miR-421 is upregulated in
neuroblastoma (16), gastric cancer
(17,18) and lung adenocarcinoma (22). The results from RT-qPCR analysis also
revealed that the expression levels of miR-421 were significantly
upregulated in the lung cancer cell lines, which is consistent with
the results of above studies. In particular, in A549 cells, miR-421
expression levels were ~11-fold higher compared with in the control
cell line BEAS-2B. Among the seven lung cancer cell lines analyzed,
miR-421 expression levels in A549 cells were neither the highest
nor the lowest; therefore, A549 cells were selected for further
analysis. However, the role of mir-421 in lung cancer cell lines in
not clear. In neuroblastoma, miR-421 substantially enhance cell
proliferation, cell-cycle progression, migration and invasion of
neuroblastoma cells through upregulation of P18 and P21 (16). Furthermore, miR-421 level is
correlated with lymph node metastasis and prognosis of gastric
carcinoma (17). miR-421 may
therefore serve important roles in lung cancer cells by similar
mechanisms.
Firstly, we studied the target gene of miR-421.
TargetScan is a commonly used software for the prediction of miRNA
target genes (30). The results of
the present study revealed that FOXO1 may be a target gene of
miR-421. Luciferase reporter assays are commonly performed to
verify the results of TargetScan (31). In the present study, the results
demonstrated that miR-421 bound to the 3′-UTR of FOXO1 and
inhibited the expression of the reporter gene. Therefore, these
findings indicated that FOXO1 may be a target gene of miR-421. The
effect of miR-421 and its target gene, FOXO1, were further analyzed
using A549 cells. Then, an MTT assay was subsequently performed to
determine the effects of miR-421 on the viability of A549 cells.
Post-transfection with the miR-421-mimics, the OD values across 5
days were increased compared with in the A549-miR-421-mimics-NC
group. Finally, we studied the molecular mechanisms of miR-421 on
the viability of A549 cells. The effect may be associated with the
downregulated expression levels of FOXO1. In addition, p21, GSK-3β,
p-Rb and cyclin D1 expression levels in A549 cells transfected with
miR-421-mimics were determined. The results revealed that the
expression levels of FOXO1 and p21 were significantly
downregulated, whereas the expression levels of p-AKT, p-GSK-3β,
p-Rb and cyclin D1 were significantly upregulated by the miR-421
mimic. These proteins are downstream molecules of FOXO1 (23). The results from the present study
indicated that AKT/GSK-3β pathway was activated, resulting in the
change of p-AKT, p-GSK-3β, p-Rb and cyclin D1 protein expression.
These proteins are strongly associated with cell cycle progression
and apoptosis (32–34). Furthermore, the A549 cell cycle
transitioned from G1 to S phase, which stimulates
proliferation and inhibits apoptosis (35).
In conclusion, the current study provided novel
evidence to suggest that upregulated expression levels of miR-421
may significantly suppress FOXO1 expression, resulting in the
enhanced viability of A549 cells in vitro. The regulation of
miR-421 via FOXO1 may provide novel insight into the
pathophysiology of lung cancer and a potential therapeutic target
for the treatment of patients with lung cancer.
Acknowledgements
The authors would like to thank Mr. Liu Ming (School
of Medicine and Pharmacy, Ocean University of China, Qingdao,
Shandong, China) for his help revising the manuscript.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
XM wrote the manuscript. XM, PQ, BW, FL and HL
performed the experiments and analyzed the data. HL and FL designed
the study. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhou C, Zhu Y, Lu B, Zhao W and Zhao X:
Survivin expression modulates the sensitivity of A549 lung cancer
cells resistance to vincristine. Oncol Lett. 16:5466–5472.
2018.PubMed/NCBI
|
|
2
|
Chen Z, Wang J, Bai Y, Wang S, Yin X,
Xiang J, Li X, He M, Zhang X, Wu T, et al: The associations of
TERT-CLPTM1L variants and TERT mRNA expression with the prognosis
of early stage non-small cell lung cancer. Cancer Gene Ther.
24:20–27. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Joshi M, Ayoola A and Belani CP:
Small-cell lung cancer: An update on targeted. Adv Exp Med Bio.
779:385–404. 2013. View Article : Google Scholar
|
|
4
|
Carter BW, Halpenny DF, Ginsberg MS,
Papadimitrakopoulou VA and de Groot PM: Immunotherapy in non-small
cell lung cancer treatment: Current status and the role of imaging.
J Thorac Imaging. 32:300–312. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li L, Sun Y, Feng M, Wang L and Liu J:
Clinical significance of blood-based miRNAs as biomarkers of
non-small cell lung cancer. Oncol Lett. 15:8915–8925.
2018.PubMed/NCBI
|
|
6
|
Lu J, Zhan Y, Feng J, Luo J and Fan S:
MicroRNAs associated with therapy of non-small cell lung cancer.
Int J Biol Sc. 14:390–397. 2018. View Article : Google Scholar
|
|
7
|
Guarize J, Bianchi F, Marino E, Belloni E,
Vecchi M, Donghi S, Lo Iacono G, Casadio C, Cuttano R, Barberis M,
et al: MicroRNA expression profile in primary lung cancer cells
lines obtained by endobronchial ultrasound transbronchial needle
aspiration. J Thorac Dis. 10:408–415. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Iqbal MA, Arora S, Prakasam G, Calin GA
and Syed MA: MicroRNA in lung cancer: Role, mechanisms, pathways
and therapeutic relevance. Mol Aspects Med. 70:3–20. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Seok H, Ham J, Jang ES and Chi SW:
MicroRNA Target Recognition: Insights from Transcriptome-Wide
Non-Canonical Interactions. Mol Cells. 39:375–381. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Isik M, Blackwell TK and Berezikov E:
MicroRNA mir-34 provides robustness to environmental stress
response via the DAF-16 network in C. elegans. Sci Rep.
6:367662016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wojciechowska A, Braniewska A and
Kozar-Kamińska K: MicroRNA in cardiovascular biology and disease.
Adv Clin Exp Med. 26:865–874. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tran N: Cancer Exosomes as miRNA
Factories. Trends Cancer. 2:329–331. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang JK, Wang Z and Li G: MicroRNA-125 in
immunity and cancer. Cancer Lett. 454:134–145. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen YL, Xu QP, Guo F and Guan WH:
MicroRNA-302d downregulates TGFBR2 expression and promotes
hepatocellular carcinoma growth and invasion. Exp Ther Med.
13:681–687. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
An X, Sarmiento C, Tan T and Zhu H:
Regulation of multidrug resistance by microRNAs in anti-cancer
therapy. Acta Pharm Sin B. 7:38–51. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li Y, Li W, Zhang JG, Li HY and Li YM:
Downregulation of tumor suppressor menin by miR-421 promotes
proliferation and migration of neuroblastoma. Tumour Biol.
35:10011–10017. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu H, Gao Y, Song D, Liu T and Feng Y:
Correlation between microRNA-421 expression level and prognosis of
gastric cancer. Int J Clin Exp Pathol. 8:15128–15132.
2015.PubMed/NCBI
|
|
18
|
Wu J, Li G, Yao Y, Wang Z, Sun W and Wang
J: MicroRNA-421 is a new potential diagnosis biomarker with higher
sensitivity and specificity than carcinoembryonic antigen and
cancer antigen 125 in gastric cancer. Biomarkers. 20:58–63. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang P, Zhang M, Liu X and Pu H:
MicroRNA-421 promotes the proliferation and metastasis of gastric
cancer cells by targeting claudin-11. Exp Ther Med. 14:2625–2632.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ge X, Liu X, Lin F, Li P, Liu K, Geng R,
Dai C, Lin Y, Tang W, Wu Z, et al: MicroRNA-421 regulated by
HIF-1alpha promotes metastasis, inhibits apoptosis, and induces
cisplatin resistance by targeting E-cadherin and caspase-3 in
gastric cancer. Oncotarget. 7:24466–24482. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hu TB, Chen HS, Cao MQ, Guo FD, Cheng XY,
Han ZB and Li MQ: MicroRNA-421 inhibits caspase-10 expression and
promotes breast cancer progression. Neoplasma. 65:49–54. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cinegaglia NC, Andrade SC, Tokar T,
Pinheiro M, Severino FE, Oliveira RA, Hasimoto EN, Cataneo DC,
Cataneo AJ, Defaveri J, et al: Integrative transcriptome analysis
identifies deregulated microRNA-transcription factor networks in
lung adenocarcinoma. Oncotarget. 7:28920–28934. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen Z, Yu W, Zhou Q, Zhang J, Jiang H,
Hao D, Wang J, Zhou Z, He C and Xiao Z: A Novel lncRNA IHS promotes
tumor proliferation and metastasis in HCC by Regulating the ERK-
and AKT/GSK-3β-Signaling Pathways. Mol Ther Nucleic Acids.
16:707–720. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kim SA, Kang OH and Kwon DY:
Cryptotanshinone Induces Cell Cycle Arrest and Apoptosis of NSCLC
Cells through the PI3K/Akt/GSK-3β Pathway. Int J Mol Sci.
19:27392018. View Article : Google Scholar
|
|
25
|
Chen Y, Liu X, Wang H, Liu S, Hu N and Li
X: Akt regulated phosphorylation of GSK-3β/Cyclin D1, p21 and p27
contributes to cell proliferation through cell cycle progression
from G1 to S/G2M Phase in low-dose arsenite exposed HaCat cells.
Front Pharmacol. 10:11762019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Coomans de Brachene A and Demoulin JB:
FOXO transcription factors in cancer development and therapy. Cell
Mol Life Sci. 73:1159–1172. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen D, Yang Y and Yang P: Quxie capsule
inhibits colon tumor growth partially through foxo1-mediated
apoptosis and immune modulation. Integr Cancer Ther.
18:15347354198463772019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
van Tonder A, Joubert AM and Cromarty AD:
Limitations of the
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide
(MTT) assay when compared to three commonly used cell enumeration
assays. BMC Res Notes. 8:472015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Riffo-Campos AL, Riquelme I and
Brebi-Mieville P: Tools for sequence-based miRNA target prediction:
What to choose? Int J Mol Sci. 17:19872016. View Article : Google Scholar
|
|
31
|
Xie D, Shang C, Zhang H, Guo Y and Tong X:
Up-regulation of miR-9 target CBX7 to regulate invasion ability of
bladder transitional cell carcinoma. Med Sci Monit. 21:225–230.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zuryn A, Litwiniec A, Gagat M, Drzewucka
J, Gackowska L and Grzanka A: The influence of arsenic trioxide on
the cell cycle, apoptosis and expression of cyclin D1 in the Jurkat
cell line. Acta Histochem. 116:1350–1358. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gongpan P, Lu Y, Wang F, Xu Y and Xiong W:
AS160 controls eukaryotic cell cycle and proliferation by
regulating the CDK inhibitor p21. Cell cycle (Georgetown Tex.).
15:1733–1741. 2016. View Article : Google Scholar
|
|
34
|
Fukuda Y, Kanbe M, Watanabe M, Dan K,
Matsuzaki K, Kitanaka S and Miyata S: 3EZ,20Ac-ingenol, a catalytic
inhibitor of topoisomerases, downregulates p-Akt and induces DSBs
and apoptosis of DT40 cells. Arch Pharm Res. 36:1029–1038. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mao Z, Zhou J, Luan J, Sheng W, Shen X and
Dong X: Tamoxifen reduces P-gp-mediated multidrug resistance via
inhibiting the PI3K/Akt signaling pathway in ER-negative human
gastric cancer cells. Biomed Pharmacother. 68:179–183. 2014.
View Article : Google Scholar : PubMed/NCBI
|