Introduction
Primary liver cancer is a common type of cancer
worldwide with high morbidity and mortality (1). In China, primary liver cancer is the
second leading cause of cancer-related death, and the incidence and
mortality rates have continued to rise over the decades (2). Primary liver cancer mainly occurs in
the context of long-term chronic liver disease, particularly in
hepatitis B and C virus infections and cirrhosis (3). Primary liver cancer is characterized as
an asymptomatic disease in early stages and is often detected when
at advanced stages. Due to being diagnosed in the advanced stages,
primary liver cancer is often incurable (4). With the development of medical science
multiple advanced treatments, such as radiofrequency ablation,
immunotherapy and molecular-targeted therapy have made significant
progress in clinical practice (5).
However, new methods for early diagnosis and for therapies are
urgently required.
Phospholipid scramblase 1 (PLSCR1) is a
Ca2+-binding type II endofacial plasma membrane protein
that serves an important role in the transbilayer movement of
phosphatidylserine and other amino phospholipids (6). These movements are often associated
with critical cellular processes, including cell activation, cell
proliferation, cell injury responses, tumour suppression,
transcriptional regulation and apoptosis (7–12). The
expression of PLSCR1 can be regulated by interferons, epidermal
growth factor (EGF) and other cytokines (13). Increasing evidence suggests that
PLSCR1 also interacts with several protein kinases that contribute
to cell signalling pathways, particularly the EGF receptor (EGFR)
signalling pathway (13,14). These studies suggest that PLSCR1 may
serve an important role in tumorigenesis. PLSCR1 has been
identified to be highly expressed in colorectal carcinoma and
breast carcinoma, suggesting that PLSCR1 is closely correlated with
tumour initiation and progression (15–17).
However, to the best of our knowledge there are no previous studies
on the biological functions and expression levels of PLSCR1 in
primary liver cancer.
The present study investigated the expression of
PLSCR1 in patients with primary liver cancer and compared
biological behaviours among HepG2 cells with different expression
levels of PLSCR1. The results of the present study may clarify the
association between PLSCR1 and human primary liver cancer.
Materials and methods
Patient information and tissue
samples
A total of 44 patients (male=31, female=11; age
range, 30–80 years) diagnosed with primary liver cancer at of Wuxi
No. 2 People's Hospital between January 2010 and December 2014 were
included in the present study. The inclusion criteria were as
follows: i) Diagnosed to be primary liver cancer treatable with
surgical resection; and ii) all the patients had complete
information for clinicopathological and prognostic characteristics.
Tumour tissue samples and adjacent noncancerous tissue samples were
obtained during surgery and normal tissue samples were obtained
from 10 normal individuals recruited (male=6, female=4; age range,
45–77 years) from the same hospital within the same time frame.
Normal individuals were defined as patients with cholelithiasis
requiring surgical treatment and preoperative laboratory tests,
their imaging examination did not reveal any hepatic lesions. None
of the patients received radiotherapy, chemotherapy or other
related anti-tumour therapies prior to surgery. Clinicopathological
features, including tumour size, differentiation grade and clinical
stage, were collected from pathological reports and checked
independently by two observers and finally incorporated in Tables I and II. The present study was approved by the
Ethics Committee of Wuxi No. 2 People's Hospital (approval no.
2009-Y-22) and written informed consent was obtained from all the
patients prior to surgery and from the normal individuals.
 | Table I.Protein levels of PLSCR1 in primary
liver cancer tissue samples, adjacent non-cancerous liver tissue
samples and normal liver tissue samples. |
Table I.
Protein levels of PLSCR1 in primary
liver cancer tissue samples, adjacent non-cancerous liver tissue
samples and normal liver tissue samples.
|
|
| PLSCR1
expression |
|
|---|
|
|
|
|
|
|---|
| Variable | Cases, n | Negative, n (%) | Positive, n (%) | P-value |
|---|
| Primary liver
cancer | 44 | 5 (11.36) | 39 (88.64) | 0.082a |
| Adjacent
non-cancerous | 44 | 29 (65.91) | 15 (34.09) |
|
| Normal | 10 | 9 (90) | 1 (10) |
<0.001b |
| Table II.Associations between PLSCR1
expression and clinicopathological features in 44 primary liver
cancer patients. |
Table II.
Associations between PLSCR1
expression and clinicopathological features in 44 primary liver
cancer patients.
|
|
| PLSCR1
expression |
|
|---|
|
|
|
|
|
|---|
| Variable | Cases, n | Negative, n
(%) | Positive, n
(%) | P-value |
|---|
| Age, years |
|
<60 | 34 | 3 (8.82) | 31 (91.18) | 0.328 |
|
≥60 | 10 | 2 (20) | 8 (80) |
|
| Sex |
|
Male | 33 | 3 (9.09) | 30 (90.91) | 0.411 |
|
Female | 11 | 2 (18.18) | 9 (81.82) |
|
| Tumour size,
cm |
| ≤3 | 12 | 2 (16.67) | 10 (83.33) | 0.497 |
|
>3 | 32 | 3 (9.38) | 29 (90.62) |
|
|
Differentiation |
|
High-moderate | 41 | 5 (12.20) | 36 (87.80) | 0.521 |
|
Poor | 3 | 0 (0) | 3 (100) |
|
| HBs-Ag |
|
Positive | 35 | 1 (2.86) | 34 (97.14) | 0.004 |
|
Negative | 9 | 4 (44.44) | 5 (55.56) |
|
| AFP, µg/l |
|
>400 | 20 | 2 (10) | 18 (90) | 0.795 |
|
≤400 | 24 | 3 (12.5) | 21 (87.5) |
|
| Lymphatic
metastasis |
|
Positive | 11 | 0 (0) | 11 (100) | 0.411 |
|
Negative | 33 | 5 (15.15) | 28 (84.85) |
|
| TNM stage |
| I | 3 | 2 (66.67) | 1 (33.33) | 0.045 |
| II | 19 | 2 (10.53) | 17 (89.47) |
|
|
III | 21 | 1 (4.76) | 20 (95.24) |
|
| IV | 1 | 0 (0) | 1 (100) |
|
Immunohistochemistry and staining
assessment
Immunohistochemical staining was performed on serial
4-µm tissue sections from formalin-fixed (10% formalin for 24 h),
paraffin-embedded human primary liver cancer and adjacent normal
samples. Dehydration was performed using a descending alcohol
series (70, 95 and 100% ethanol). Firstly, paraffin sections were
baked in an oven at 60°C for 1 h. The paraffin slides were
deparaffinized and washed three times with PBS. The sections were
subsequently immersed in 0.01 mmol/l EDTA, microwaved at 98°C for
25 min for antigen retrieval and allowed to cool to room
temperature (RT). Each section was then washed with PBS, followed
by an incubation for 10 min with 0.3% H2O2 to
inhibit endogenous peroxidase activity. Each section was then
washed with PBS and incubated with 20% goat serum (Thermo Fisher
Scientific, Inc.) for 30 min at room temperature and a diluted
rabbit anti-PLSCR1 polyclonal antibody (1:1,000; cat. no. ab250216,
Abcam) for 2 h at 37°C and 2 h at RT, successively. After being
washed with PBS, the slides were incubated with horseradish
peroxidase-labeled anti-rabbit IgG secondary antibody (goat,
1:2,000; cat. no. A-1178, Thermo Fisher Scientific, Inc.) for 30
min at RT. Subsequently, the slides were washed with PBS, incubated
with EnVision (Dako; Agilent Technologies, Inc.) for 30 min at RT
and then thoroughly washed with PBS. Subsequently, the sections
were developed with diaminobenzidine tetrahydrochloride, washed in
running water and counterstained with haematoxylin for 2 min at RT.
Representative samples were selected and images captured. Finally,
the sections were observed under an Olympus light microscope (BX61;
Olympus Corporation; magnification, ×400). Five random fields were
selected and evaluated by two blinded experienced pathologists from
Wuxi No. 2 People's Hospital (Wuxi, China). The following
definition of immunostaining intensity from Kuo et al
(17) was used: Negative, no brown
particles in the plasma membrane; positive, light-brown particles,
moderately brown particles or deep brown particles in the plasma
membrane.
Cell lines and cell culture
HepG2 cell lines (purchased from the Cell Bank of
the Chinese Academy of Sciences) were cultured in DMEM (Gibco;
Thermo Fisher Scientific, Inc.) supplemented with 10% FBS (Gibco;
Thermo Fisher Scientific, Inc.). All cells were maintained at 37°C
in a humidified incubator with 5% CO2. STR profiling was
performed to ensure the cell identity.
Establishment of cell lines with
stable PLSCR1 up- or downregulation
HepG2 cells were seeded in 6-well plates 24 h prior
to viral infection at a density of 2×105 cells per well
and incubated at 37°C with 5% CO2 overnight. The cells
were transiently transfected with synthesized PLSCR1-specific short
hairpin (sh)RNAs (sh-PLSCR1, Applied Biological Materials, Inc.), a
PLSCR1-expressing lentivirus (lv-PLSCR1, Applied Biological
Materials, Inc.) or the corresponding negative control (shnon or
Lvnon, respectively, from Applied Biological Materials, Inc.) using
Lipofectamine 2000® (Invitrogen; Thermo Fisher
Scientific, Inc.) after removing the growth medium. Subsequently, 1
µl of polybrene (5 µg/ml) was added to each well. The medium was
changed 24 h later and replaced with 2 ml of complete medium, and
the incubation was continued. Stable PLSCR1-expressing cell lines
were harvested following transfection. Reverse
transcription-quantitative (RT-q) PCR and western blotting were
performed to investigate the expression levels of PLSCR1. Three
days later, well-growing monoclonal strains were selected for
frozen storage and subsequent experiments.
RT-qPCR
Transfected cancer cells were processed with
TRIzol® (Life Technologies; Thermo Fisher Scientific,
Inc.). Total RNA was reverse transcribed into cDNA using the
PrimeScript™ RT reagent kit (Takara Bio, Inc.) at 50°C for 45 min.
All the experiments were performed following manufacturer's
protocols. qPCR was subsequently performed using the SYBR Green I
Master mix kit (Invitrogen; Thermo Fisher Scientific, Inc.) and a
7500 Real-Time PCR System (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The PCR thermocycling conditions were: 95°C for
2 min; and 40 cycles of 95°C for 10 sec, 60°C for 30 sec and 72°C
for 30 sec. The sequences of the PCR primers were: PLSCR1-Forward,
5′-CACCCATGTCTACCAAAGTT-3′ and PLSCR1-Reverse,
3′-CTCTCAAAATTCCAGTCCAG-5′; β-actin forward,
5′-GAAGATCAAGATCATTGCTCCT-3′ and β-actin reverse,
5′-TACTCCTGCTTGCTGATCCA-3′. The relative gene expression levels
were calculated using the 2−ΔΔCq method (18) and normalized to the internal
reference gene β-actin. Each sample was run in triplicate and
independently repeated three times.
Western blotting
Total protein was extracted from transfected cancer
cells using lysis buffer (Beyotime Institute of Biotechnology), and
protein concentrations were measured with a Bicinchoninic Acid
Protein Assay kit (Beyotime Institute of Biotechnology). A total of
30 µl protein extract was separated using 10% SDS-PAGE gels and
transferred onto a PVDF membrane. The membranes were blocked with
5% non-fat milk for 60 min at RT, followed by incubation with
primary antibodies against PLSCR1 (1:500; cat. no. ab250216;
Abcam), EGFR (1:500; cat. no. ab52894; Abcam), and β-actin (1:500;
cat. no. ab8227; Abcam) overnight at 4°C, then treated with a
Horseradish peroxidase-labeled goat anti-rabbit secondary antibody
(1:1,000; cat. no. ab150077; Abcam) for 2 h at RT. The protein
bands were visualized using an enhanced chemiluminescence system
(Beyotime Institute of Biotechnology). β-actin was used as a
loading control for normalization and images were analyzed using
ImageJ software version 1.52 (National Institutes of Health).
CCK8 assay
Transfected cancer cells (8×103 per well)
were seeded in 96-well plates in quintuplicate and incubated at
37°C. DMEM was removed 24 h later and replaced with 100 µl of fresh
medium. A Cell Counting Kit-8 (CCK8; Dojindo Molecular Technologies
Inc.) assay was used to evaluate cell proliferation according to
the manufacturer's instructions. A CCK8 solution (10 µl) was added
to each well, and the cells were incubated for 4 h at 37°C in a
CO2 incubator. The absorbance at 450 nm was determined
by an ELISA plate reader.
Wound heal assay
Cells (3×105/well) were seeded and grown
in 6-well plates until confluent (90%) cell monolayers were formed.
Then, a sterile 10-µl pipette tip was used to make a uniform
scratch along the bottom of each well. The horizontal distance
between the sides of the scratch was measured. The cells were
washed with PBS three times to remove detached cells and incubated
with serum-free medium at 37°C for 24 h after replacement of the
culture medium. Then, wound size was observed and images were
captured under an inverted fluorescence microscope (Nikon
Corporation; magnification: ×20).
Transwell migration and invasion
assays
Transwell chambers were used for transfected cancer
cell migration and invasion assays. The 24-well culture inserts (BD
Biosciences) with 8-µm pores coated with Matrigel (1 mg/ml; BD
Biosciences) were used. The membranes of upper chamber were coated
with Matrigel (BD) and then incubated for 6 h at 37°C. For the
migration assay, 2×105 transfected cells suspended in
200 µl of serum-free medium were seeded in the upper chambers.
Medium containing 20% FBS was added to the lower chamber. For the
invasion assay, a Chemicon Cell Invasion Assay kit (Chemicon
International; Thermo Fisher Scientific, Inc.) was used. The
non-migrating/invading cells were removed from the upper surface of
the membrane with cotton swabs following an incubation for 24 h at
37°C, and the migrating/invading cells were stained with 0.1%
crystal violet for 30 min at RT and images captured with an
inverted fluorescence microscope (magnification: ×20; Nikon
Corporation).
Adhesion assays
A 96-well plate was coated with laminin (Ln, 20
µg/well) or fibronectin (Fn, 10 µg/well) for 2 h at 37°C, washed
with PBS twice and sealed with 1% BSA (Thermo Fisher Scientific,
Inc.). A single-cell suspension (3×105 cancer cells/ml)
was prepared with transfected cells and serum-free medium. Then,
200 µl of suspension was added to the coated plates. The cells were
incubated at 37°C with 5% CO2 for 2 h and then the
non-adherent cells were gently washed off. Each well was fixed with
4% formaldehyde for 10 min at RT and stained with 0.5%
hexamethylpararosaniline for 20 min at RT. Subsequently, 100 µl of
10% SDS was added to each well. An ELISA plate reader was used to
assess the absorbance at 570 nm.
Statistical analysis
SPSS 18.0 software (SPSS, Inc.) was used to perform
all statistical analyses. Continuous variables are presented as the
mean ± SD and comparisons between groups were performed using
one-way ANOVAs followed by post-hoc Tukey's tests. Categorical data
are expressed as a proportion (%) and were compared using the
χ2 test, McNemar's test or Fisher's exact test.
Bonferroni's corrections were used to reduce type I errors as the
primary liver cancer data in Table I
had been analysed more than once. P<0.05 was considered to
indicate a statistically significant difference.
Results
Expression of PLSCR1 in primary liver
cancer tissue samples and the associations of PLSCR1 levels with
clinicopathological variables
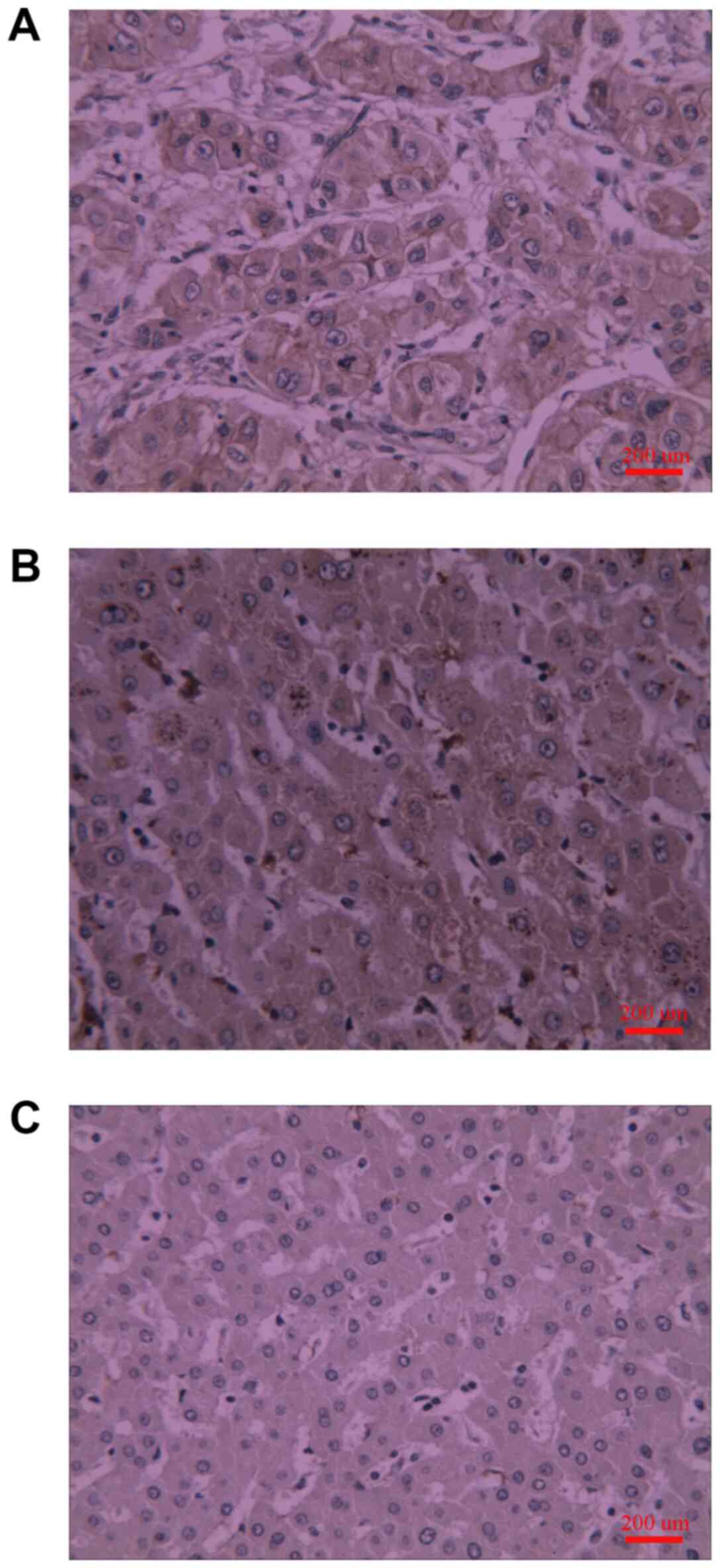
Fig. 1 and Table I show that PLSCR1 protein was
detected in liver tissue samples through immunohistochemical
staining. The PLSCR1 protein was mainly located at the cancer cell
membrane (Fig. 1). The expression of
PLSCR1 in the primary liver cancer sample group was significantly
higher compared with the normal group, while no significant
difference was observed between primary liver cancer tissue samples
and the adjacent non-cancerous liver tissue samples (Fig. 1 and Table
I). As shown in Table II,
PLSCR1 expression was observed in almost all primary liver cancer
patients (88.64%, 39/44 patients), whereas the positive rate of
PLSCR1 expression in the normal group was only 10%. The positive
rate of PLSCR1 expression increased with the increase of tumour
size, although the difference was not statistically significant
(83.33% vs. 90.62%). The expression of PLSCR1 in primary liver
cancer was correlated with the hepatitis B virus (HBV) infection
status. Furthermore, it was identified that the positive rate of
PLSCR1 expression in TNM stage III/IV samples was significantly
higher compared with that in stage I/II samples, suggesting that
the expression level of PLSCR1 is correlated with clinical stage.
However, no correlations were identified between PLSCR1 expression
and other clinicopathological characteristics, including patient
sex, tumour size, tumour differentiation, α fetoprotein level and
lymph node metastasis status.
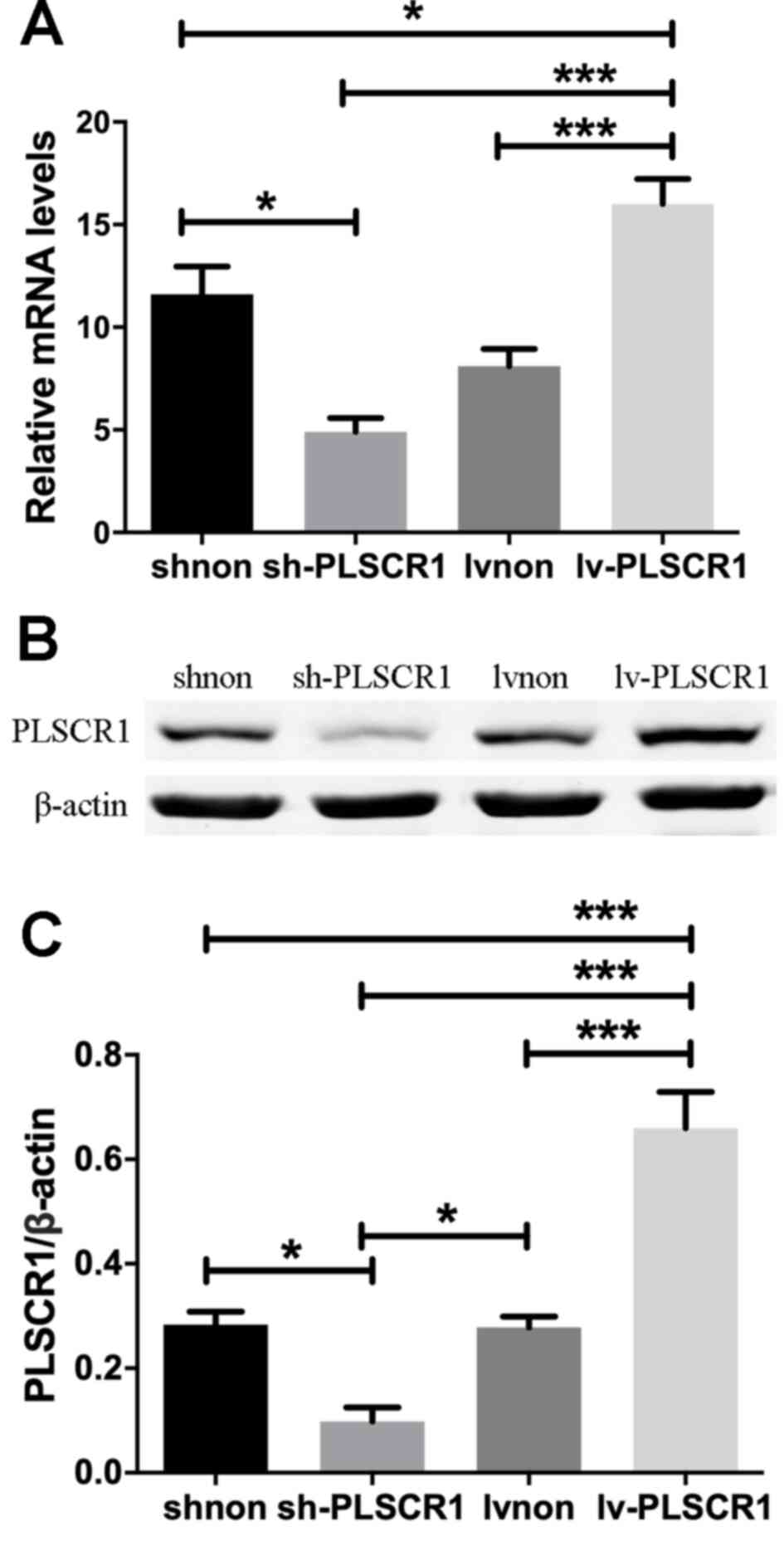
PLSCR1 expression in transfected
cancer cells
Following transfection with sh-PLSCR1, Lv-PLSCR1 or
the corresponding negative control, the expression levels of PLSCR1
in the various groups was assessed using RT-qPCR and western
blotting. The sh-PLSCR1 cell lines expressed the lowest levels of
PLSCR1 at both the mRNA and protein levels, while significantly
increased expression levels of PLSCR1 was observed in the Lv-PLSCR1
group compared with the other three groups (Fig. 2).
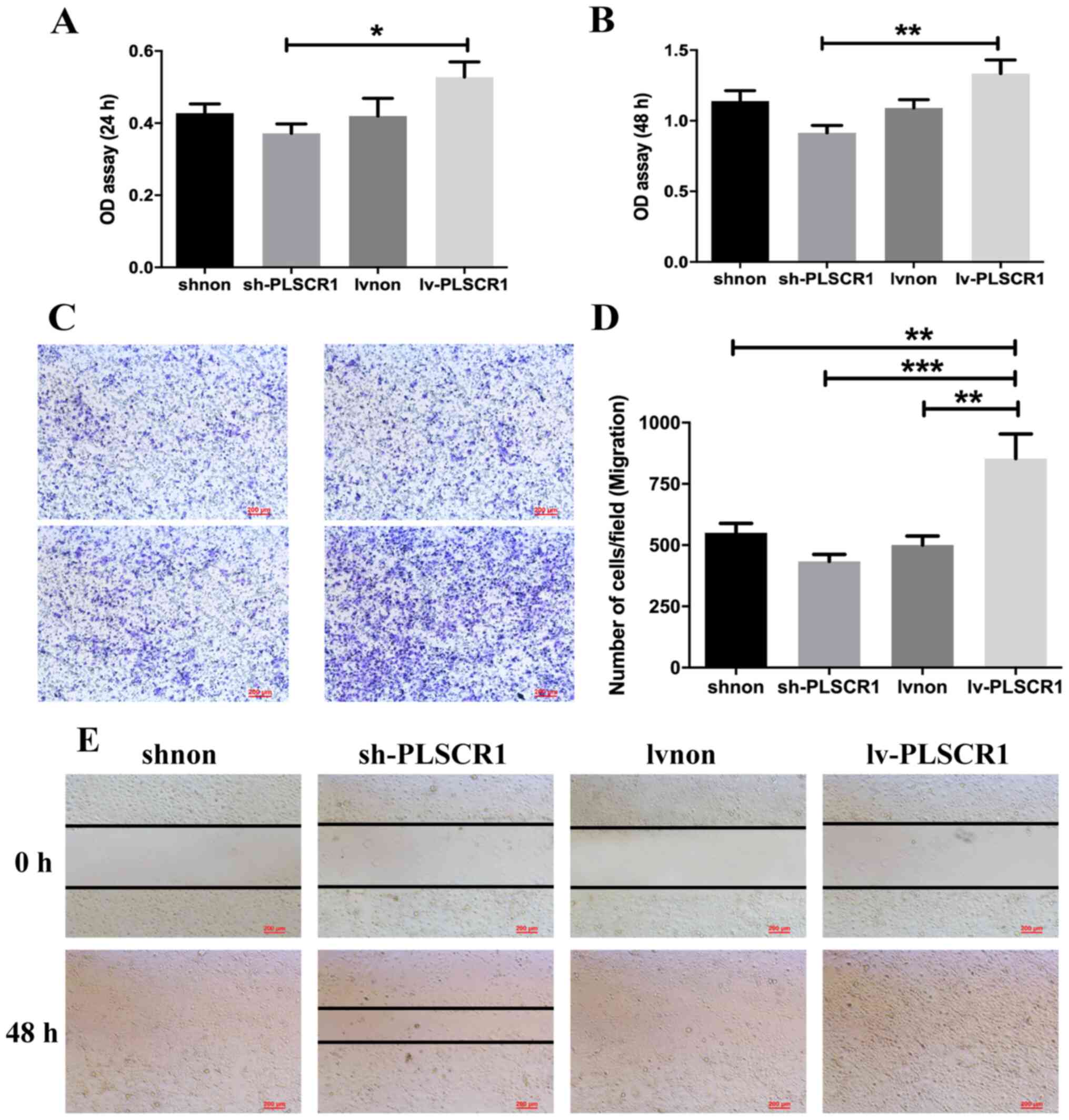
Effects of PLSCR1 expression on cell
proliferation, adhesion, migration and invasion
Following transfection, PLSCR1 was overexpressed in
the Lv-PLSCR1 cells, and the expression levels of PLSCR1 were
reduced in the sh-PLSCR1 cells as aforementioned. To investigate
the effects of PLSCR1 on the biological behaviour of HepG2 cells, a
series of assays including CCK8, adhesion and Transwell migration
and invasion assays were performed. As shown in Fig. 3A and B, the CCK8 assays indicated
that compared with the Lv-PLSCR1 cells, the sh-PLSCR1 cells
exhibited significantly inhibited proliferation at 24 and 48 h.
Representative micrographs of Transwell filters are shown in
Fig. 3C. Transwell migration and
wound heal assays demonstrated that PLSCR1 overexpression resulted
in an increased number of migrating Lv-PLSCR1 cells and promoted
the migration of cancer cells (Fig.
3C-E). In addition, the number of invasive HepG2 cells was
significantly decreased in the sh-PLSCR1 group compared with the
Lv-PLSCR1 group (Fig. 4A and B). Ln
and Fn adhesion assays were performed to assess HepG2 cell adhesion
following transfection. These assays demonstrated that HepG2 cell
adhesion was inhibited in the sh-PLSCR1 group at 48 h (Fig. 4C and D). Collectively, the data
suggested that PLSCR1 upregulation may promote HepG2 cell
proliferation, migration and invasion and that PLSCR1
downregulation can inhibit these effects.
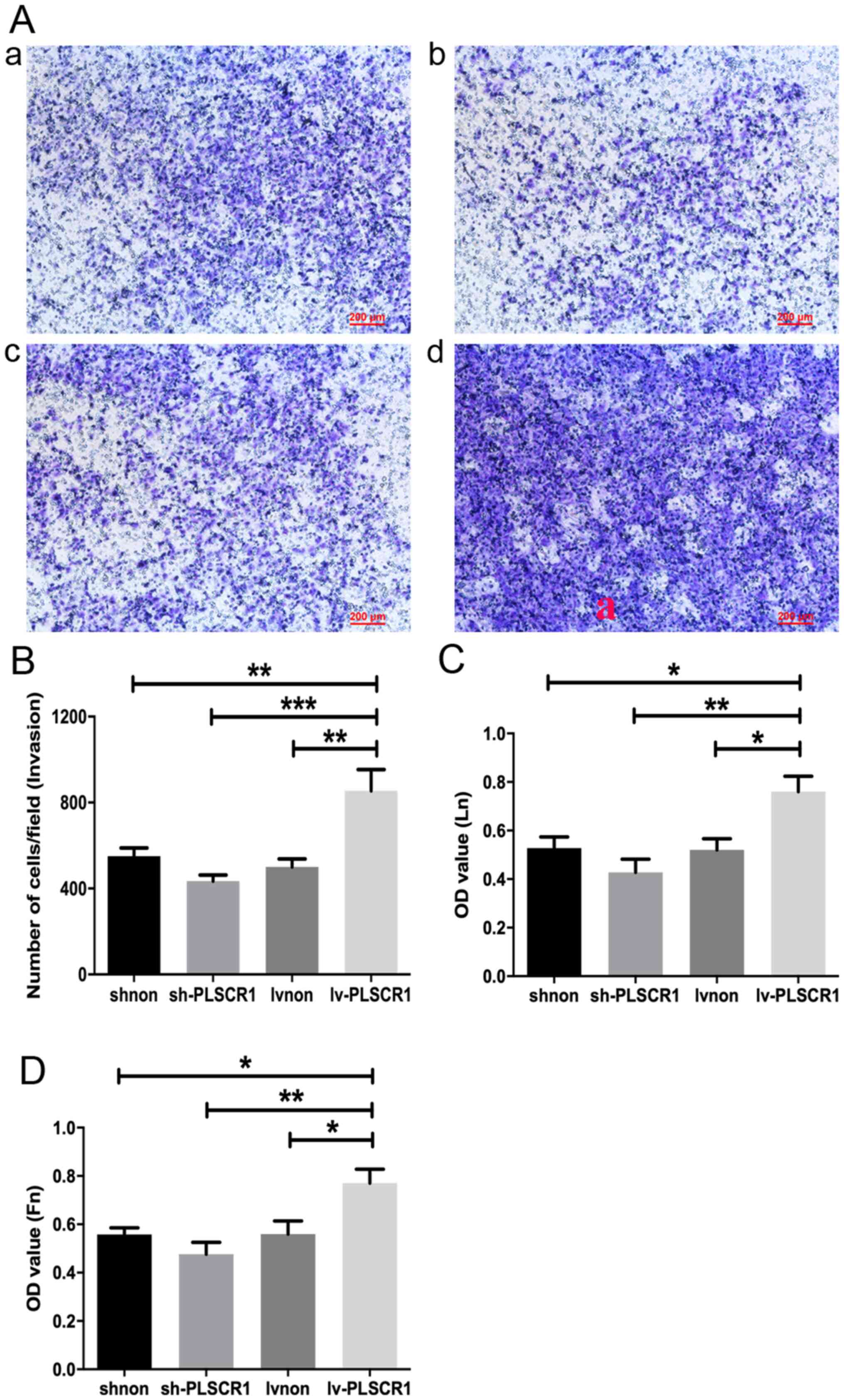 | Figure 4.Lentivirus-mediated overexpression of
PLSCR1 promotes HepG2 cell invasion and adhesion in vitro.
(A) Representative images of cell invasion determined by Transwell
invasion assays of cells transfected with (a) shnon, (b) sh-PLSCR1,
(c) Lvnon or (d) Lv-PLSCR1. (B) Quantification of the Transwell
invasion assays. (C) Ln and (D) Fn adhesion assays. Data are
presented as the mean ± SD. *P<0.05, **P<0.01, ***P<0.001.
OD, optical density; PSCR1, phospholipid scramblase 1; sh, short
hairpin RNA; lv, lentivirus; Ln, laminin; Fn, fibronectin. |
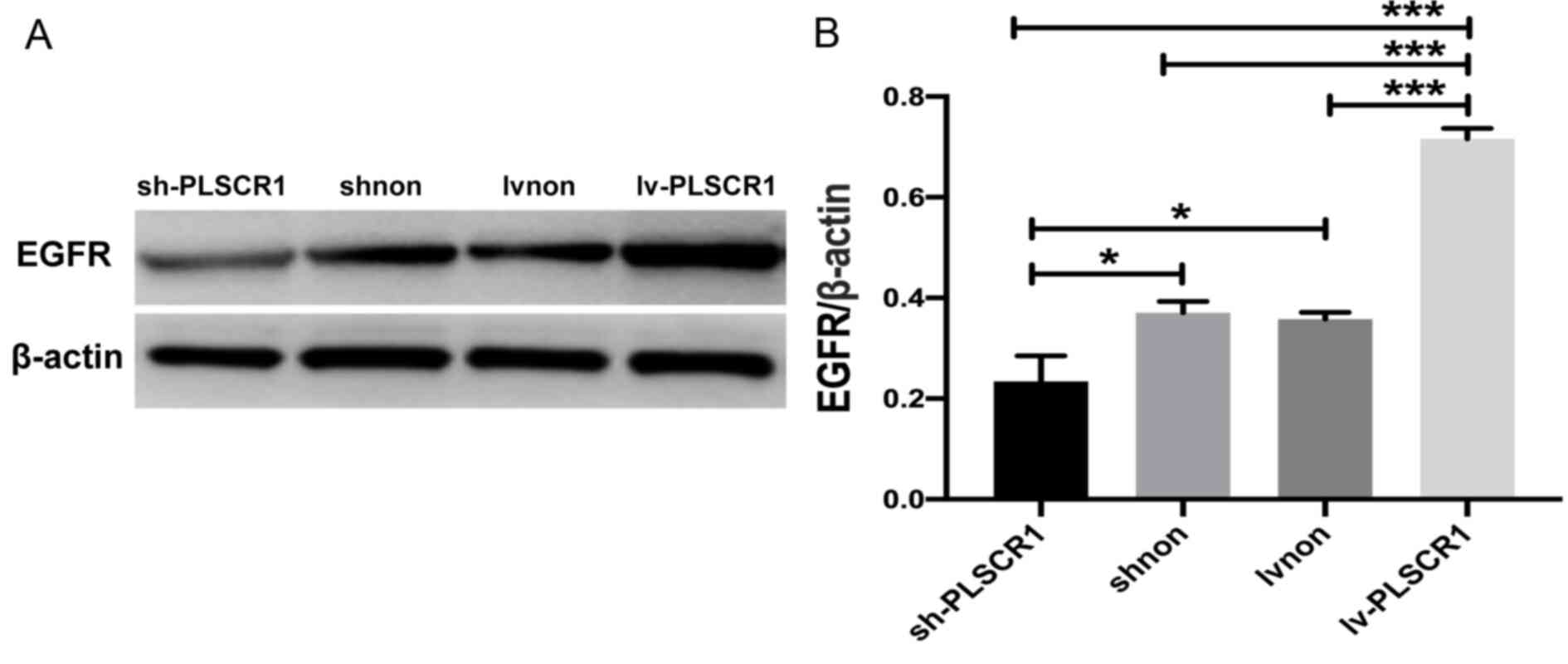
Suppression of PLSCR1 expression
blocks the EGFR signalling pathway
As shown in Fig. 5,
expression of EGFR was decreased in cancer cells following PLSCR1
inhibition but increased in the PLSCR1-overexpression cells.
Discussion
Primary liver cancer is considered a serious health
problem of global importance (19).
The majority of patients are asymptomatic in the early stage but
are often diagnosed in advanced stages, which are resistant to
therapies (4,20). Surgery is an optimal approach for
primary liver cancer treatment, but the majority of patients may
not be able to benefit from surgery due to a high rate of
recurrence (21). Therefore, early
diagnosis and the identification of new therapeutic targets will
benefit patients with primary liver cancer.
As aforementioned, increasing evidence suggests that
PLSCR1 serves important roles in cell proliferation,
differentiation, apoptosis and cancer pathogenesis and progression
(17). Studies have demonstrated
that PLSCR1 overexpression is associated with the differentiation
of human myeloid leukaemia cells into granulocytes and inhibits the
growth of ovarian cancer cells (9,22). Other
studies have revealed that suppression of PLSCR1 inhibits the
growth and metastasis of colorectal cancer (16,23).
Nevertheless, the roles of PLSCR1 in the pathogenesis and
progression of primary liver cancer remain to be elucidated.
In the present study, there was a strong correlation
between PLSCR1 expression and tumor stage. Univariate analysis with
the Cox regression model demonstrates that increased PLSCR1
expression indicates a poor prognosis for patients with colorectal
cancer (16,17). Based on these results and previous
studies, it was hypothesized that PLSCR1 may serve a vital role
behind the tumorigenesis of primary liver cancer. The present study
also demonstrated that the expression of PLSCR1 in primary liver
cancer was associated with the HBs-Ag status. Yuan et al
(24) reported an important role for
PLSCR1 in the host defence against HBV infection and the
development of primary liver cancer in patients with HBV, which are
consistent with the findings of the present study.
To evaluate the biological function of PLSCR1, siRNA
and lentiviral technologies were used to regulate the expression of
PLSCR1 and the changes in expression were verified by RT-qPCR and
western blotting. The results demonstrated that proliferation,
adhesion, migration and invasion were blocked following the
suppression of PLSCR1 expression and promoted following
upregulation of PLSCR1 expression, suggesting that PLSCR1 could be
a potential therapeutic target for hepatocellular carcinoma.
Previous studies have indicated that the EGFR signalling pathway is
inhibited in cells with knocked down PLSCR1 expression (9,22).
Stimulation of EGFR-expressing cells via EGF causes PLSCR1 to
interact with activated EGFR and SHC Adaptor Protein 1 (Shc), which
results in the tyrosine phosphorylation of PLSCR1 (25). PLSCR1 is a substrate of multiple
kinases, involved in cell proliferation, differentiation or
apoptosis including tyrosine-protein kinase ABL1, proto-oncogene
tyrosine-protein kinase Src (Src) and protein kinase (14,26–28). In
EGF-treated cells, activated Src kinase causes the tyrosine
phosphorylation of PLSCR1, which is required for the interaction of
PLSCR1 with Shc in response to EGF stimulation (14). In the present study, the production
of EGFR was decreased following suppression of PLSCR1 expression
and increased following upregulation of PLSCR1. Therefore,
suppression of PLSCR1 expression may block the EGFR signalling
pathway and inhibit the proliferation, adhesion, migration and
invasion of HepG2 cells. However, a limitation of the present study
is that the downstream proteins inside the EGFR signal penetration
pathway were not measured, which would further support the
reduction in EGFR signalling. Lack of in vivo experiments is
another limitation of the present study and which should be further
investigated.
In summary, the present study indicated that PLSCR1
was highly expressed in primary liver cancer and associated with
the tumour stage. Downregulating the expression of PLSCR1
significantly inhibited the proliferation, adhesion, migration and
invasion of cancer cells. PLSCR1 overexpression may serve important
roles in tumorigenesis and tumour progression, and suppressing
PLSCR1 expression may be beneficial for the treatment of primary
liver cancer. However, further research needs to be conducted to
explore the potential molecular mechanisms of the signalling
pathways involved.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This work was supported by i) Gastroenterology of
key disciplines from Wuxi (grant no. DXK002), ii) Academician
Workstation of No. 2 People's Hospital of Wuxi City (grant no.
CYR1705), iii) Wuxi Medical Key Talent Project (grant no. ZDRC029),
iv) Wuxi Sci-Tech Development Fund (grant no. CSE31N1603), v)
Technical achievements and appropriate technology promotion project
of Wuxi Municipal Commission of Health and Family Planning (grant
no. T201603), vi) Wuxi municipal commission of health and family
planning project (grant no. MS201720), vii) Wuxi municipal
commission of health and family planning project (grant no.
MS201814), viii) Wujin district Changzhou Sci-Tech Development Fund
(grant no. WS201811), ix) Jiangsu commission of health project
(grant no. LGY2018021) and x) Jiangsu University clinical medical
science and technology development fund (grant no.
JLY20180090).
Availability of data and materials
The datasets used and analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LG, YZ and QX designed the study, performed the
experiments and analysed the data statistically. LG wrote the
manuscript. JH, PH and GW performed data collection. LZ and JL made
substantial contributions to the conception of this study. JN and
HT conducted data processing and statistical analyses, and made
critical revisions. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
Wuxi No. 2 People's Hospital, and written informed consent was
obtained from all the patients prior to surgery.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Grandhi MS, Kim AK, Ronnekleiv-Kelly SM,
Kamel IR, Ghasebeh MA and Pawlik TM: Hepatocellular carcinoma: From
diagnosis to treatment. Surg Oncol. 25:74–85. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhu Y, Tang H, Zhang L, Gong L, Wu G, Ni J
and Tang X: Suppression of miR-21-3p enhances TRAIL-mediated
apoptosis in liver cancer stem cells by suppressing PI3K/Akt/Bad
cascade via regulating PTEN. Cancer Manag Res. 11:955–968. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Balogh J, Victor D III, Asham EH,
Burroughs SG, Boktour M, Saharia A, Li X, Ghobrial RM and Monsour
HP Jr: Hepatocellular carcinoma: A review. J Hepatocell Carcinoma.
3:41–53. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gomes MA, Priolli DG, Tralhao JG and
Botelho MF: Hepatocellular carcinoma: Epidemiology, biology,
diagnosis, and therapies. Rev Assoc Med Bras (1992). 59:514–524.
2013.(In English, Portuguese). View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cidon EU: Systemic treatment of
hepatocellular carcinoma: Past, present and future. World J
Hepatol. 9:7972017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wiedmer T, Zhao J, Nanjundan M and Sims
PJ: Palmitoylation of phospholipid scramblase 1 controls its
distribution between nucleus and plasma membrane. Biochemistry.
42:1227–1233. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhou Q, Zhao J, Wiedmer T and Sims PJ:
Normal hemostasis but defective hematopoietic response to growth
factors in mice deficient in phospholipid scramblase 1. Blood.
99:4030–4038. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huang Y, Zhao Q, Zhou CX, Gu ZM, Li D, Xu
HZ, Wiedmer T, Sims PJ, Zhao KW and Chen GQ: Antileukemic roles of
human phospholipid scramblase 1 gene, evidence from inducible
PLSCR1-expressing leukemic cells. Oncogene. 25:6618–6627. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Silverman RH, Halloum A, Zhou A, Dong B,
Al-Zoghaibi F, Kushner D, Zhou Q, Zhao J, Wiedmer T and Sims PJ:
Suppression of ovarian carcinoma cell growth in vivo by the
interferon-inducible plasma membrane protein, phospholipid
scramblase 1. Cancer Res. 62:397–402. 2002.PubMed/NCBI
|
|
10
|
Francis VG, Padmanabhan P and Gummadi SN:
Snail interacts with hPLSCR1 promoter and down regulates its
expression in IMR-32. Biochem Biophys Res Commun. 450:172–177.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Vinnakota JM and Gummadi SN: Two c-Myc
binding sites are crucial in upregulating the expression of human
phospholipid scramblase 1 gene. Biochem Biophys Res Commun.
469:412–417. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhou Q, Ben-Efraim I, Bigcas J, Junqueira
D, Wiedmer T and Sims PJ: Phospholipid scramblase 1 binds to the
promoter region of the inositol 1,4,5-triphosphate receptor type 1
gene to enhance its expression. J Biol Chem. 280:35062–35068. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sahu SK, Gummadi SN, Manoj N and Aradhyam
GK: Phospholipid scramblases: An overview. Arch Biochem Biophys.
462:103–114. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nanjundan M, Sun J, Zhao J, Zhou Q, Sims
PJ and Wiedmer T: Plasma membrane phospholipid scramblase 1
promotes EGF-dependent activation of c-Src through the epidermal
growth factor receptor. J Biol Chem. 278:37413–37418. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Choi HJ, Lui A, Ogony J, Jan R, Sims PJ
and Lewis-Wambi J: Targeting interferon response genes sensitizes
aromatase inhibitor resistant breast cancer cells to
estrogen-induced cell death. Breast Cancer Res. 17:62015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cui W, Li SY, Du JF, Zhu ZM and An P:
Silencing phospholipid scramblase 1 expression by RNA interference
in colorectal cancer and metastatic liver cancer. Hepatob Pancreat
Dis. 11:393–400. 2012. View Article : Google Scholar
|
|
17
|
Kuo YB, Chan CC, Chang CA, Fan CW, Hung
RP, Hung YS, Chen KT, Yu JS, Chang YS and Chan EC: Identification
of phospholipid scramblase 1 as a biomarker and determination of
its prognostic value for colorectal cancer. Mol Med. 17:41–47.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Singal AG and El-Serag HB: Hepatocellular
carcinoma from epidemiology to prevention: Translating knowledge
into practice. Clin Gastroenterol Hepatol. 13:2140–2151. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Finn RS: Emerging targeted strategies in
advanced hepatocellular carcinoma. Semin Liver Dis. 33 (Suppl
1):S11–S19. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Brito AF, Abrantes AM, Tralhão JG and
Botelho MF: Targeting hepatocellular carcinoma: What did we
discover so far? Oncol Rev. 10:3022016.PubMed/NCBI
|
|
22
|
Nakamaki T, Okabe-Kado J,
Yamamoto-Yamaguchi Y, Hino Ki, Tomoyasu S, Honma Y and Kasukabe T:
Role of MmTRA1b/phospholipid scramblase1 gene expression in the
induction of differentiation of human myeloid leukemia cells into
granulocytes. Exp Hematol. 30:421–429. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fan CW, Chen CY, Chen KT, Shen CR, Kuo YB,
Chen YS, Chou YP, Wei WS and Chan EC: Blockade of phospholipid
scramblase 1 with its N-terminal domain antibody reduces
tumorigenesis of colorectal carcinomas in vitro and in vivo. J
Transl Med. 10:2542012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yuan Y, Tian C, Gong Q, Shang L, Zhang Y,
Jin C, He F and Wang J: Interactome map reveals phospholipid
scramblase 1 as a novel regulator of hepatitis B virus × protein. J
Proteome Res. 14:154–163. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yokoyama A, Yamashita T, Shiozawa E,
Nagasawa A, Okabe-Kado J, Nakamaki T, Tomoyasu S, Kimura F,
Motoyoshi K, Honma Y and Kasukabe T: MmTRA1b/phospholipid
scramblase 1 gene expression is a new prognostic factor for acute
myelogenous leukemia. Leuk Res. 28:149–157. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sun J, Zhao J, Schwartz MA, Wang JY,
Wiedmer T and Sims PJ: c-Abl tyrosine kinase binds and
phosphorylates phospholipid scramblase 1. J Biol Chem.
276:28984–28990. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kowalczyk JE, Beręsewicz M, Gajkowska B
and Zabłocka B: Association of protein kinase C delta and
phospholipid scramblase 3 in hippocampal mitochondria correlates
with neuronal vulnerability to brain ischemia. Neurochem Int.
55:157–163. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhao KW, Li X, Zhao Q, Huang Y, Li D, Peng
ZG, Shen WZ, Zhao J, Zhou Q, Chen Z, et al: Protein kinase Cdelta
mediates retinoic acid and phorbol myristate acetate-induced
phospholipid scramblase 1 gene expression: Its role in leukemic
cell differentiation. Blood. 104:3731–3738. 2004. View Article : Google Scholar : PubMed/NCBI
|