Introduction
Esophageal squamous cell carcinoma (ESCC) is a
common form of aggressive malignancy (1), with a poor prognosis and high mortality
rate, 455,800 new esophageal cancer cases and 400,200 deaths
occurred in 2012 worldwide and most of the histological subtype
were ESCC (2). Predisposing factors
for ESCC include smoking and alcohol (3). The development and progression of ESCC
is very complex, with multiple genetic mutations and disorders
involved, such as mutations in TP53, CDKN2A or a loss of RB gene
(4,5). Currently, surgery, radiotherapy and
chemotherapy are commonly used singly or in combination to treat
ESCC. However, these methods may also damage healthy cells and have
limited efficacy in cancer infiltration and metastasis (6,7).
Therefore, novel and curative agents for ESCC are urgently
required. Gene therapy, which delivers therapeutic genes into cells
to alter the gene expression of a patient, may be a promising
strategy for cancer treatment (8).
Numerous cancer gene therapy approaches have reported significant
progress, including gene immunization, suppressor genes or gene
replacements, gene directed enzyme-prodrug or suicide gene
therapies and oncolytic virus therapies (9–11).
In its mature form, melittin is a 26 amino acid
peptide with strong hemolytic and antimicrobial activity, and is
the principal active component of bee venom (12). Melittin is in routine use as a
non-steroidal anti-inflammatory reagent for the relief of pain and
the treatment of chronic inflammatory diseases (13,14).
Moreover, melittin has demonstrated the ability to inhibit cancer
cell proliferation and induce apoptosis (15,16).
Compared with other pro-apoptotic proteins such as p53, Bak and
Bax, melittin is not selective, damaging both healthy and cancer
cells. To overcome the non-specific cytotoxicity of melittin,
targeted expression is a necessary prerequisite for successful
cancer gene therapy (17).
Human telomerase reverse transcriptase (hTERT) is
the catalytic subunit of telomerase and is the rate-limiting step
in the activation of telomerase (18). Furthermore, it has been reported that
hTERT expression is common to most cancer cells (18).
The present study constructed a recombinant plasmid
(pcTERT-melittin) containing the hTERT promoter followed by
melittin, and investigated the ability of this pcTERT-melittin
construct to act as a therapeutic agent able to inhibit
proliferation and induce apoptosis in TE1 cells.
Materials and methods
Materials
The human esophageal carcinoma TE1 and normal
esophageal epithelial cells Het-1a cell lines were obtained from
the American Type Culture Collection. DMEM and FBS were obtained
from Hyclone (Cytiva). BEGM BulletKit containing basal medium
(BEBM) plus additives (growth factors, hEGF, VEGF, hFGF, IGF-1;
FBS, hydrocortisone, ascorbic acid, heparin and transferrin) (BEGM
SingleQuots) was obtained Lonza Group, Ltd. Opti-MEM reduced serum
medium, pcDNA3.1+ plasmid, TRIzol RNA Isolation Reagents
and Lipofectamine® 2000 transfection reagent were
purchased from Invitrogen (Thermo Fisher Scientific, Inc.). Takara
Ex Taq DNA Polymerase was from Takara Biotechnology. The Cell
Counting Kit (CCK)-8 (cat. no. C0037), reactive oxygen species
(ROS) assay kit (cat. no. S0033S), mitochondrial membrane potential
(∆Ψm) assay kit with JC-1 (cat. no. C2006), caspase-3 activity
assay kit (cat. no. C1115) and caspase-9 activity assay kit (cat.
no. C1158) were purchased from Beyotime Institute of Biotechnology.
Annexin V-FITC staining kit and complete protease inhibitor
cocktail tablets were purchased from Roche Diagnostics.
Antibodies against tubulin (1:1,000; cat. no.
sc-80005) and horseradish peroxidase-conjugated secondary
antibodies to rabbit, mouse and goat primary antibodies were
purchased from Santa Cruz Biotechnology, Inc. Antibodies against
poly(ADP-ribose) polymerase 1 (PARP) (1:1,000; cat. no. 9542),
cleaved caspase-9 (1:1,000; cat. no. 20750), caspase-3 (1:1,000;
cat. no. 9662), cleaved caspase-3 (1:1,000; cat. no. 9661) and Bax
(1:100; cat. no. 2772) were purchased from Cell Signaling
Technology, Inc. Antibodies against CDK4 (1:1,000; cat. no. A0366),
CDK6 (1:10,00l; cat. no. A1545), cyclin D1 (1:1,000; cat. no.
A19038) and p53 (1:1,000; cat. no. A11232) were purchased from
Abclonal Biotech Co., Ltd.
EasySee® western blotting kit was
purchased from Beijing Transgen Biotech Co., Ltd. Transwell
chambers were purchased from Corning, Inc. A Primescript™ Reverse
Transcription reagent kit was purchased from Takara Biotechnology
Co., Ltd. SYBR® premix ex Taq™ II, ROX plus reagent kit
was purchased from Roche Diagnostics. All other chemicals were
analytical reagent grade.
Construction of pcTERT-melittin
vector
A recombinant plasmid encoding melittin was
constructed using a pcDNA3.1+ plasmid containing the
hTERT promoter. The enhancer sequence of pcDNA3.1+ was
first amplified using PCR. The pcDNA3.1 plasmid was used as
template. The thermocycling conditions were as follows: 30 cycles
at 95°C for 10 sec, 56°C for 30 sec and 72°C for 1 min. Forward
Primer: 5′-CTGACGCGTGGAGTTCCGCGTTAC−3′; Reverse Primer:
5′-CGCGCTAGCCAAAACAAACTCCCATTG−3′)with the MluI and
NheI restriction sites engineered at the ends of primer,
then four different hTERT promoter DNAs, each with NheI and
HindIII restriction sties, were amplified from human genomic
DNA. The melittin coding gene was synthesized by Sangon Biotech
Co., Ltd. The enhancer, hTERT promoter and melittin gene sequence
(ATGGGCATCGGCGCCATCCTGAAGGTGCTGAGCACCGGCCTGCCCGCCCTGATCAGCTGGATCAAGAGAAAGAGACAGGAGTAA)
were inserted into the pcDNA3.1+ consecutively to
produce a recombinant plasmid (pcTERT-melittin). The recombinant
plasmid containing only the enhancer and hTERT promoter was named
‘pcTERT’.
Cell culture
TE1 cells were cultivated in DMEM supplemented with
10% FBS, while Het-1a cells were cultured with bronchial epithelial
cell medium (BEGM BulletKit) containing basal medium (BEBM) plus
additives (BEGM SingleQuot), and incubated at 37°C with 5%
CO2. The cells were passaged every ~2 days. TE1 cells
were transfected with recombinant plasmid pcTERT-melittin or pcTERT
(2 µg plasmid for 12-well plate and 4 µg plasmid for 6-well plate)
at 60% of plate confluence using Lipofectamine 2000, following
manufacturer's instructions. Following transfection, the cells were
observed using a phase-contrast microscope (Olympus Corporation)
(magnification, ×200). For the cell proliferation assay, the
measurement of ∆Ψm using tetraethylbenzimidazolylcarbocyanine
iodide (JC-1) staining, intracellular ROS quantification assays,
Annexin-V-FITC apoptosis assay, cell cycle assay and cell migration
assay, the cells were detected immediately after transfection. For
the caspase-3 and caspase-9 activity assay and western blot
analysis, after the transfection, cells were lysed and proteins
were detected immediately. For the RNA extraction and reverse
transcription-quantitative (RT-q) PCR assay, after the
transfection, RNA was extracted immediately and reverse-transcribed
to cDNA, then stored at −80°C for subsequent PCR assays.
Agarose gel electrophoresis
The construction of pcTERT-melittin was identified
with agarose gel electrophoresis. In brief, after plasmids digested
with restriction endonucleases (HindIII and XbaI) for
8 h at 37°C, samples were analyzed with a 1% agarose gel at 80 V at
37°C. The pcTERT-melittin plasmid was sent to Sangon Biotech Co.,
Ltd to investigate whether the sequence of melittin had the correct
reading frames inserted.
Cell proliferation assay
Determination of the number of viable cells was
performed using a TransDetect CCK-8 assay according the
manufacturer's instructions. TE1 and Het-1a cells were plated in
quadruplicate at a concentration of 5×103 per well in
96-well cell culture plates. Cells were incubated overnight at 37°C
with 5% CO2 and then transfected with the recombinant
plasmids for 24–72 h. After treatment with 10 µl CCK-8 solution at
37°C for 1 h, the plates were measured using a microplate
spectrophotometer at 450 nm. The survival rate was calculated using
the formula: Survival rate=(mean absorption of treated group/mean
absorption of control group)x100%.
Measurement of ∆Ψm using
tetraethylbenzimidazolylcarbocyanine iodide (JC-1) staining. The
change in ∆Ψm is a key feature of early stage apoptosis (19). The change in ∆Ψm was monitored using
the mitochondrial membrane potential (∆Ψm) assay kit containing the
cell-permeable cationic JC-1 dye and fluorescence microscopy
according to the manufacturer's instructions. This procedure did
not require fixation. In brief, the TE1 cells were plated into
12-well plates at 2×105 cells/well and incubated at 37°C
with 5% CO2 overnight. A total of 3×105 TE1
cells were transfected with 2 µg pcTERT-melittin or pcTERT for 24
h, then the medium was replaced with DMEM containing JC-1 and
incubated for 20 min at 37°C with 5% CO2. After
incubation, the medium was removed and cells were washed with
ice-cold JC-1 buffer twice. The cells were observed using
fluorescent microscopy, five random fields were observed at a
magnification of ×200. Red emission was characteristic of normal
polarized mitochondria, while green fluorescence was observed with
a depolarized mitochondrial membrane, which is characteristic of
early stage apoptosis (20).
Intracellular ROS quantification
assays
Intracellular ROS levels were assessed by detecting
the oxidative conversion of dichlorodihydrofluorescein diacetate
(DCFH-DA) to 2′,7′-dichlorofluorescein in TE1 cells according to
the ROS assay kit manufacturer's instructions. TE1 cells were
transfected with 2 µg recombinant plasmids for 48 h, then harvested
with trypsinization. Then, 1×105 cells were incubated
with 5 µl DCFH-DA in 1 ml DMEM for 20 min at 37°C. The cells were
then washed three times and re-suspended in PBS, followed via
assessment using a fluorescence microplate at excitation and
emission wavelengths of 488 and 525 nm, respectively.
Annexin-V-FITC apoptosis assay
The quantification of apoptotic TE1 cells was
performed using flow cytometry with Annexin V-FITC and PI dyes,
according to the manufacturer's instructions. A total of
7.5×105 TE1 cells in 6-well plates were transfected with
4 µg recombinant plasmids pcTERT-melittin or pcTERT for 24, 48 and
72 h, before being harvested via trypsinization and washed with
PBS. Subsequently, 70% cold ethanol was used to fix the TE1 cells
for 30 min at 4°C. Pellets were re-suspended and incubated in
Annexin V-FITC labelling solution at 15–25°C for 15 min. PI was
added at 4°C and stained for 5 min. The apoptotic cells were then
analyzed using a flow cytometer (BD Calibur; Becton-Dickinson and
Company). Data were analyzed with Cell Quest data acquisition and
analysis software (Becton-Dickinson and Company).
Cell cycle assay
TE1 cell cycle distribution was measured using a
flow cytometer by quantifying the PI labeled DNA content. After
transfection with recombinant plasmids for 48 h, TE1 cells were
collected and fixed with 70% cold ethanol at 4°C for 30 min, then
washed with PBS. Pellets were re-suspended and incubated in PBS
containing RNaseA (50 µg/ml), Triton (0.2%) and PI (20 µg/ml) at
4°C for 30 min in the dark. Cell cycle analysis was performed using
a flow cytometer (BD Calibur; Becton-Dickinson and Company). Data
were analyzed with Cell Quest data acquisition and analysis
software (Becton-Dickinson and Company) after 1 h.
Cell migration assay
Transwell chambers (6.5 mm diameter inserts; 8.0-µm
pore size) were used to measure cell migration. After transfection
with recombinant plasmids for 48 h, TE1 cells were harvested with
trypsinization. Then, 1×105 cells were re-suspended in
200 µl serum-free DMEM medium and cultivated in the upper well of a
24-well Transwell chamber without Matrigel, while 400 µl DMEM
containing 10% FBS was plated in the lower chamber. For the upper
well with Matrigel (wells precoated with Matrigel for 10 min at
37°C), 1.5×105 cells were seeded. After seeding for 15
h, the cells in the top chamber were removed. Chambers were washed
with PBS three times. Migrating cells in the lower chamber were
fixed with 4% paraformaldehyde for 15 min at 37°C, washed with PBS
and stained with 2% crystal violet at 37°C for 15 min. TE1 cells in
the lower chamber were counted using light microscopy
(magnification, ×200).
RNA extraction and reverse
transcription-quantitative (RT-q)PCR
A total of 3×105 TE1 cells were
transfected with 2 µg plasmids for 48 and 72 h in 12-well plates
before RNA extraction. Total RNA was extracted using a
TRIzol® reagent. Briefly, 500 µl TRIzol reagent was
added to the well, before cells were transferred to tubes and then
incubated for 5 min at room temperature. The tubes were shaken for
30 sec, then 100 µl chloroform was added and left to stand for 10
min at 37°C. The samples were centrifuged at 12,000 × g at 4°C for
15 min, and the upper layer of supernatant was collected. An equal
volume of isopropanol (~100 µl) was added to the supernatant. After
standing for 10 min at 37°C, the samples were centrifuged at 12,000
× g for 30 min at 4°C. The RNA pellet was washed with 75% ethanol
and dissolved in DEPC treated water. Then, 1 µg total RNA was
reverse-transcribed to cDNA using the following temperature
protocol, 42°C for 30 min and 85°C for 5 sec using a Primescript™
RT reagent kit. qPCR was performed using a SYBR® premix
ex Taq™ II, ROX plus reagent kit, conducted in the Step One plus™
Real-Time PCR system (Thermo Fisher Scientific, Inc.). The primers
was as follows: Bax: Forward, primer, 5′-GGCAACTTCAACTGGGGC−3′;
reverse, primer, 5′-CCACCCTGGTCTTGGATCC-3′; and Bcl-2 forward,
primer, 5′-AGGATTGTGGCCTTCTTTGA-3′; Reverse primer,
5′-TCAGGTACTCAGTCATCCAC-3′. The PCR protocol was as follows:
Initial denaturation at 94°C for 10 min, followed by 40 cycles at
90°C for 5 sec and 60°C for 30 sec. Products were verified using
melting curve analysis. Each sample was calculated from threshold
cycle numbers and GAPDH was monitored as the internal control.
Fold-changes in target gene mRNA expression were determined using
the 2−ΔΔCq method (21).
Caspase-3 and caspase-9 activity
assay
The activities of caspase-3 and caspase-9 in TE1
cells were measured using a caspase activity assay kit following
the manufacturer's instructions. Cells were transfected with
pcTERT-melittin or pcTERT for 24–72 h, digested with trypsin
(0.25%) for 1 min at 37°C, the cells were collected and mixed with
lysis buffer (1% Triton X-100, 50 mM HEPES, 50 mM sodium
pyrophosphate, 100 mM sodium fluoride, 10 m EDTA and 10 mM sodium
vanadate) at 37°C for 5 min. After incubating at 0°C on ice for 20
min, the samples were centrifuged at 12,000 × g at 4°C for 30 min.
The supernatant was incubated with reaction buffer at 37°C for 11
h, then the absorbance measured at 450 nm using a microplate
reader.
Western blot analysis
For western blot analysis, TE1 cells were lysed on
ice with ice-cold lysis buffer (1% Triton X-100; 50 mmol/l HEPES;
50 mmol/l sodium pyrophosphate; 100 mmol/l sodium fluoride; 10
mmol/l EDTA; 10 mmol/l sodium vanadate) containing protease
inhibitors cocktail. The lysates were centrifuged for 15 min at
15,000 × g at 4°C. The supernatant was analyzed with a BCA protein
assay kit to confirm protein concentration. Subsequently, 50 µg
proteins were boiled at 100°C for 5 min, then separated on 8–15%
SDS-PAGE, before being transferred onto a PVDF membrane. Membranes
were blocked with 5% fat-free milk in TBS-0.1% Tween-20 (TBST)
buffer at room temperature for 1 h, and then incubated with the
primary antibodies overnight at 4°C. Membranes were washed with
TBST buffer three times, followed by incubation with the
corresponding secondary antibody at 37°C for 1 h and washing with
TBST buffer three times. The protein-antibody bound bands were
visualized using an EasySee® western blotting kit and
the signal strength of each protein was normalized against the
corresponding control (signal strength of tubulin) and analyzed
with ImageJ software version 1.52 (National Institutes of
Health).
Statistical analysis
Data are presented as the mean ± SEM. Data analysis
for comparison between pcTERT group and pcTERT-melittin group was
performed using SPSS version 19.0 (IBM Corp). Data were analyzed
with two-tailed unpaired Student's t-tests and one-way ANOVA
followed by the Least Significant Difference post hoc test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Construction of recombinant plasmid
pcTERT-melittin
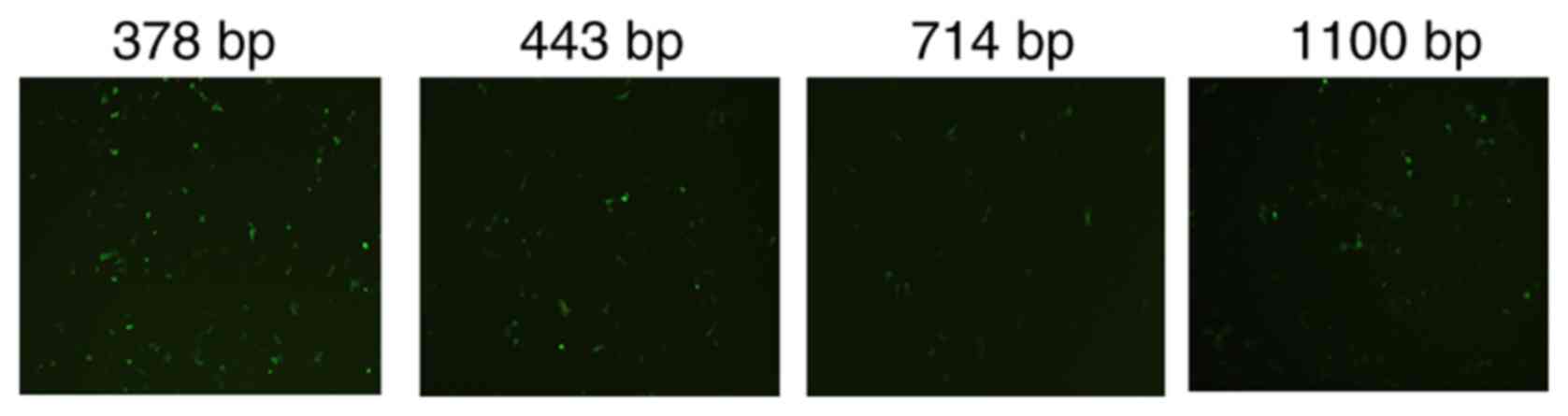
The activity of different hTERT promoters was
assessed by inserting a green fluorescence protein expressed
sequence EGFP after the hTERT promoter. The 378 bp promoter
demonstrated the greatest level of transcription compared with the
443 bp, 714 bp promoter and 1,100 bp promoters, and was used to
construct the pcTERT-melittin plasmid (Fig. 1). Agarose gel electrophoresis was
performed after pcTERT-melittin plasmid was digested with
restriction endonucleases (HindIII and XbaI), and the
100 bp fragment of melittin was observed (Fig. S1A). The sequence of melittin was
also demonstrated to be inserted with correct reading frames via
gene sequencing (Fig. S1B).
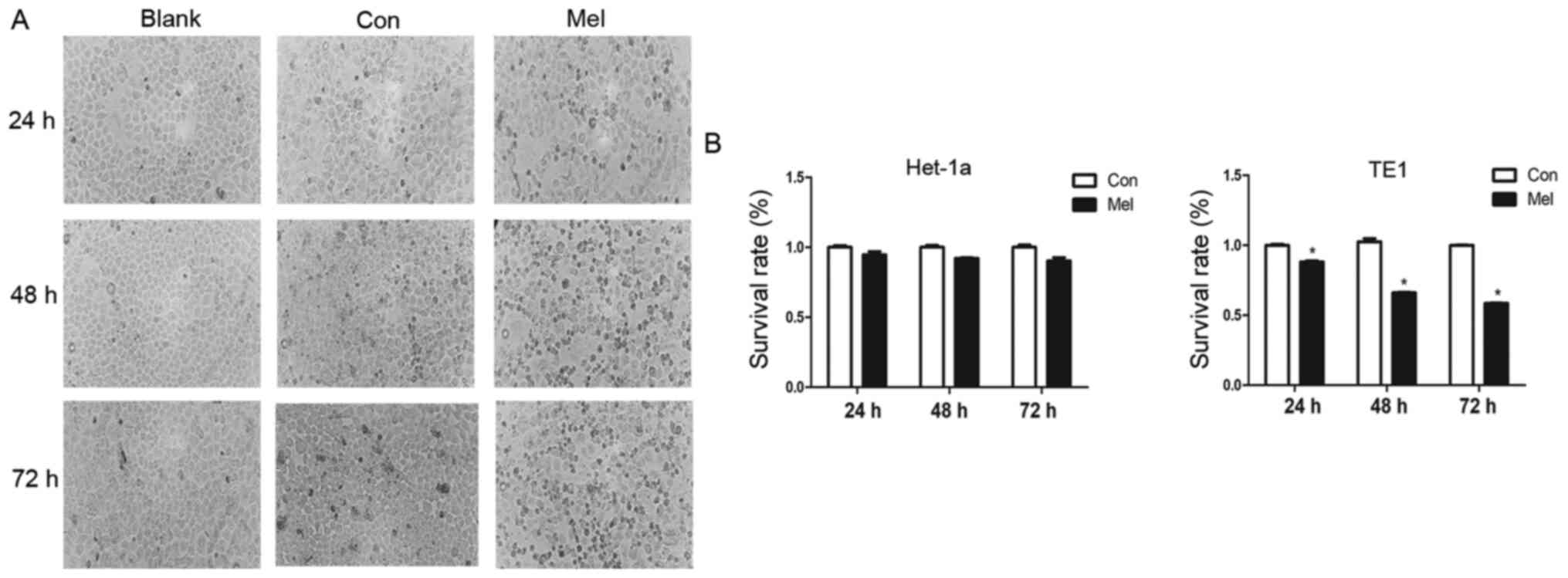
Proliferation is inhibited by
pcTERT-melittin in TE1 cells
Morphological changes typically occur during cell
apoptosis (19). To investigate the
effects of pcTERT-melittin on TE1 cells, the morphological changes
of transfected TE1 cells were observed using phase-contrast
microscopy. Phase-contrast micrographs demonstrated that TE1 cells
exposed to pcTERT-melittin exhibited cell shrinkage, apoptotic
vacuoles, membrane blebbing and formed floating cells in a
time-dependent manner, indicating that pcTERT-melittin induced
apoptosis in TE1 cells (Fig. 2A).
Cells transfected with pcTERT and untreated cells displayed normal
morphology with distinct cell borders (Fig. 2A).
A cell proliferation assay was conducted in TE1
cells exposed to recombinant plasmids for 24, 48 and 72 h using a
TransDetect CCK-8 assay. pcTERT-melittin induced a significant
decrease in cell survival in TE1 cells. The viable cell percentage
in pcTERT-melittin treated cell populations was 88.1% of the
control after transfection for 24 h, 66% of the control after 48 h
and 58.6% of the control after 72 h in TE1 cells (Fig. 2B). However, no significant effects on
cell viability were found between pcTERT-melittin treated cells and
controls in Het-1a cells. This suggested that pcTERT-melittin
treatment inhibited cell proliferation in a time-dependent manner
compared with controls in TE1 cells.
Subsequently, pcTERT and pcTERT-melittin plasmids
were used to transfect Het-1a and TE1 cells, and the mRNA
expression of melittin was measured using RT-qPCR. The mRNA
expression of melittin could only be detected in TE1 cells
transfected with pcTERT-melittin plasmid for 48 h, but not in
Het-1a cells transfected with pcTERT and pcTERT-melittin plasmid
(using the same primer pair) or TE1 cells transfected with pcTERT
(Fig. S2).
pcTERT-melittin induces apoptosis in
TE1 cells associated with damage to mitochondrial membranes and the
production of ROS
∆Ψm is a characteristic parameter for monitoring
mitochondria-dependent cell apoptosis, since reduction of the ∆Ψm
increases the generation of ROS, leading to the release of
pro-apoptotic factors such as cytochrome c (22). The current study assessed the
influence of recombinant plasmids on ∆Ψm in living cells using a
fluorescence microscope with the fluorescent dye JC-1. JC-1 is a
cationic dye that accumulates in the lumen of mitochondria,
producing red fluorescence in normally polarized mitochondria. As
the Ψm deceases, JC-1 becomes monomeric, showing green fluorescence
(20). Green fluorescence of JC-1
was observed in TE1 cells treated with pcTERT-melittin, which was
reflective of JC-1 existing in a monomeric state, and suggested a
reduction in ∆Ψm (Fig. 3A).
Moreover, pcTERT treated TE1 cells and untreated cells both
exhibited red cell-staining, indicating normal ∆Ψm. The
mitochondrial depolarization observed in pcTERT-melittin treated
TE1 cells suggested that pcTERT-melittin induces early stage
apoptosis.
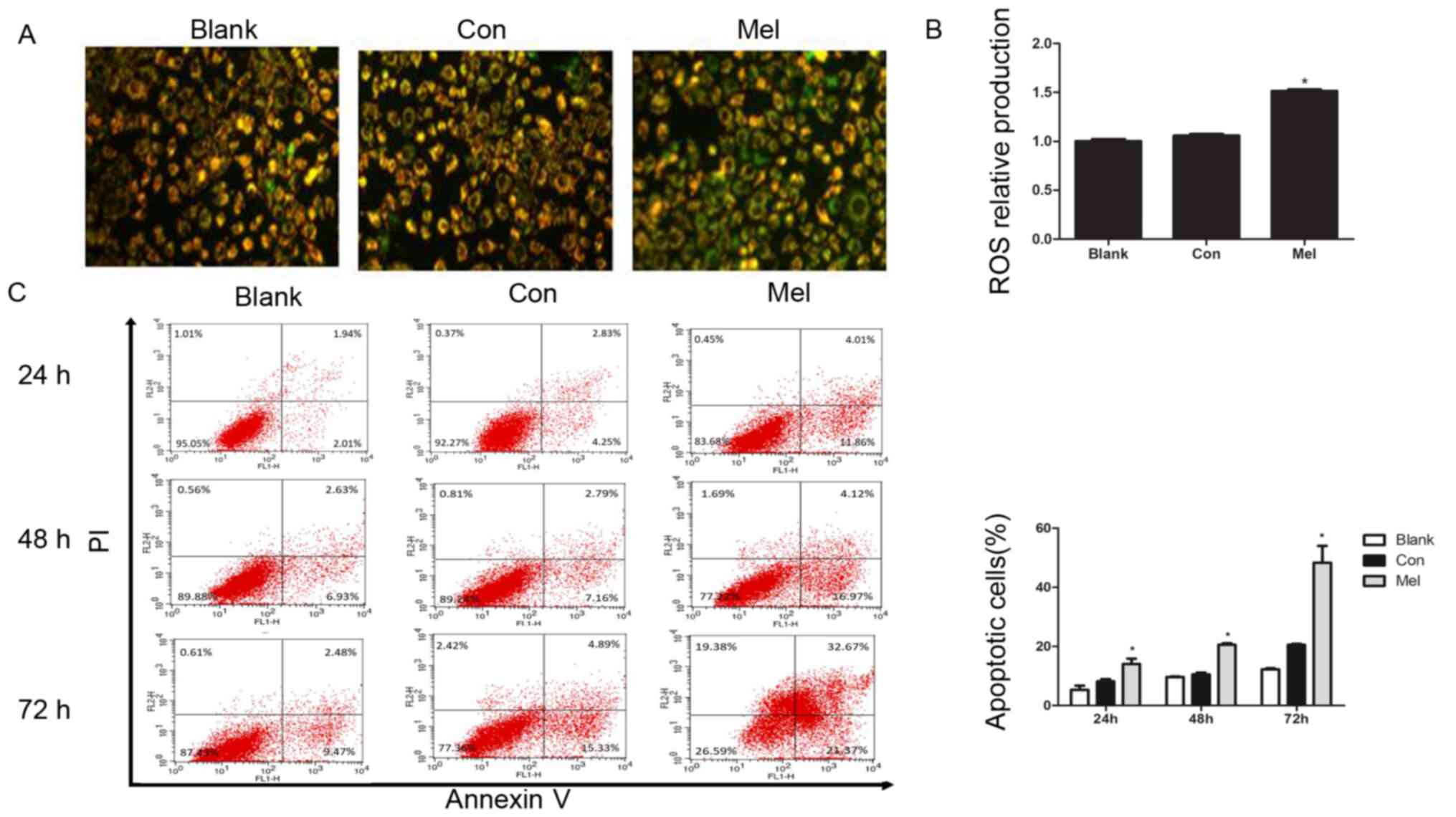 | Figure 3.Transfection of pcTERT-Mel decreases
mitochondrial membrane potential and increases ROS production in
TE1 cells, leading to apoptosis. (A) Cells were stained with
tetraethylbenzimidazolylcarbocyanine iodide and visualized using a
fluorescence microscope at 24 h post-transfection. pcTERT treated
cells and untreated cells stained red suggested normal high
membrane potentials. pcTERT-Mel treatment caused a significant loss
of red fluorescence and an increase of green fluorescence,
indicating the loss of mitochondrial membrane potential, which was
associated with apoptosis (original magnification, ×200). (B) ROS
production was detected with a ROS assay kit. Increased ROS
production was observed in pcTERT-melittin treated cells with a
fluorescence microplate at excitation and emission wavelengths of
488 and 525 nm, respectively. (C) Quantification of the
pcTERT-melittin transfection-induced apoptosis of TE1 cells, as
assessed via flow cytometry using Annexin-V and PI staining at 24,
48 and 72 h post-transfection. The percentage of apoptotic cells
was presented as the mean ± SEM. Results are an average of three
independent experiments. *P<0.05 vs. Con group. TERT, telomerase
reverse transcriptase; Con, control; Mel, melittin; ROS, reactive
oxygen species. |
Reduction in ∆Ψm is typically associated with the
opening of mitochondrial permeability transition pores, resulting
in the release of ROS (23). It was
identified that the production of ROS was significantly increased
in pcTERT-melittin treated cells compared with controls (Fig. 3B).
After typical apoptotic morphological changes, low
survival rate and mitochondrial depolarization were observed in TE1
cells transfected with pcTERT-melittin, apoptotic cells were
counted with the Annexin V-FITC and PI double-staining method using
a flow cytometer. TE1 cells transfected with pcTERT-melittin
demonstrated a significant increase in Annexin V-positive cells
compared with pcTERT treated cells (Fig.
3C). After transfection with pcTERT-melittin for 24 h,
apoptotic TE1 cells were significantly higher (14.08±2.53%)
compared with the controls (8.15±1.12%). At 48 h post-transfection,
the percentage of apoptotic cells that had been transfected with
pcTERT-melittin increased to 20.56±0.76% compared with the pcTERT
group (10.56±0.86%). After transfection for 72 h, the number of
apoptotic TE1 cells was 48.36±8.04% in the pcTERT-melittin group
compared with 21.05±1.17% in the pcTERT group.
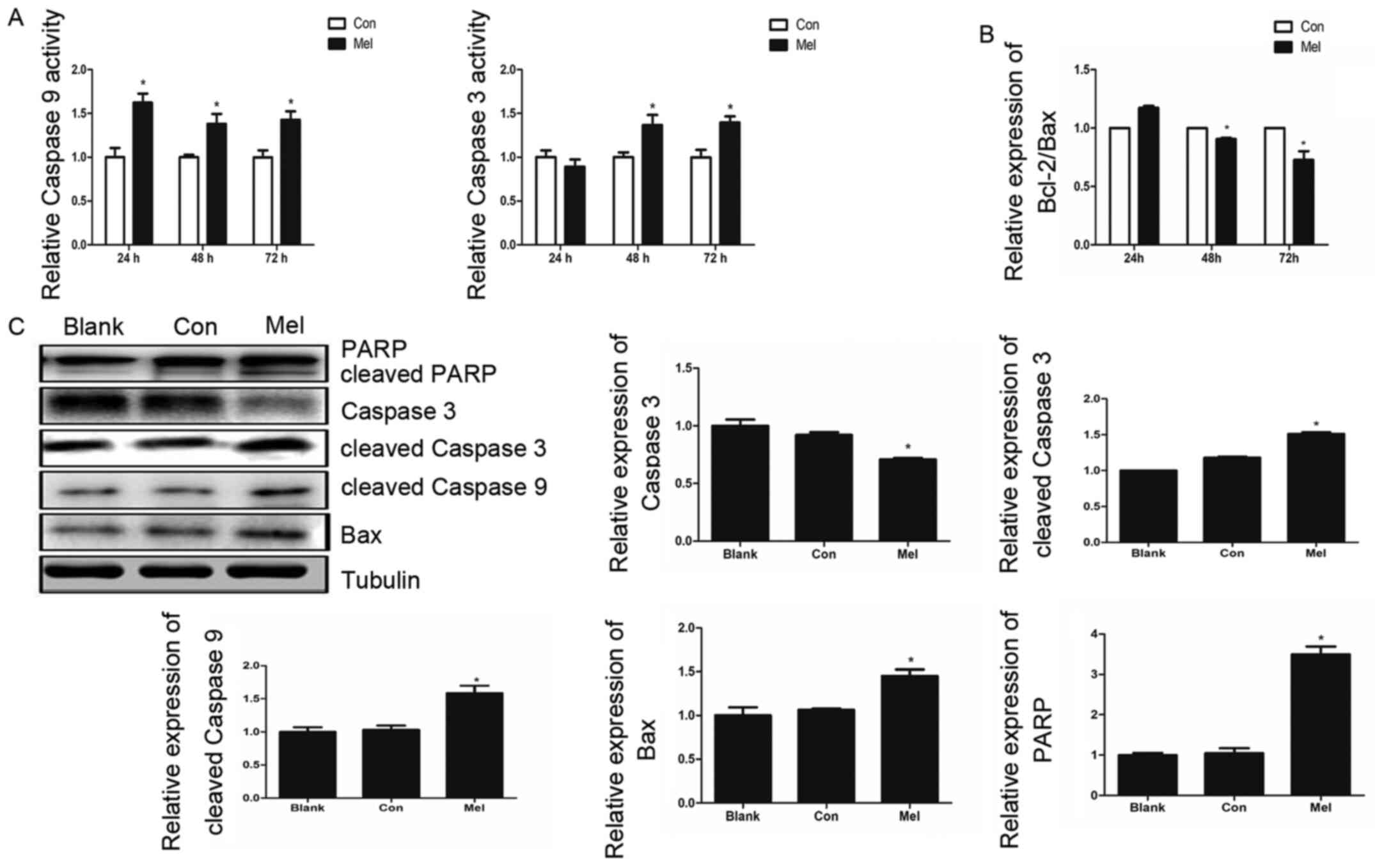
pcTERT-melittin induces apoptosis in
TE1 cells via a mitochondrial pathway
The Annexin V-FITC and PI staining assay results
demonstrated that the recombinant plasmid can induce apoptosis in
TE1 cells, thus decreasing ∆Ψm, which suggested that
pcTERT-melittin induces apoptosis via the mitochondrial pathway.
Caspase-9 and caspase-3 serve crucial roles in the apoptotic
mitochondrial pathway (24).
Therefore, caspase activity assays were conducted to investigate
the associated caspase activities in TE1 cells. Caspase-3 activity
increased 1.4 fold after pcTERT-melittin transfection for 48 and 72
h, and the activity of caspase-9 was significantly upregulated in
pcTERT-melittin treated cells after 24–72 h, indicating the
activation of caspase-9 of pcTERT-melittin group appears earlier
compared with caspase-3 activation of the pcTRET-melittin group
(Fig. 4A).
The expression levels of Bax and Bcl-2, which are
upstream of caspase-3 and caspase-9 in the apoptotic mitochondrial
pathway (25), were investigated
using RT-qPCR. The results demonstrated that the relative
expression of Bcl-2/Bax ratio in pcTERT-melittin treated TE1 cells
was significantly lower compared with controls, in a time-dependent
manner (Fig. 4B). Moreover, after 48
h transfection, the expression of pro-apoptotic cleaved caspase-9
was upregulated, followed by an increase in cleaved caspase-3 in
pcTERT-melittin treated TE1 cells, compared with controls (Fig. 4C). As the downstream substrate of
caspase-3, the expression of cleaved-PARP also increased when
caspase-3 was cleaved in pcTERT-melittin treated TE1 cells,
compared with controls.
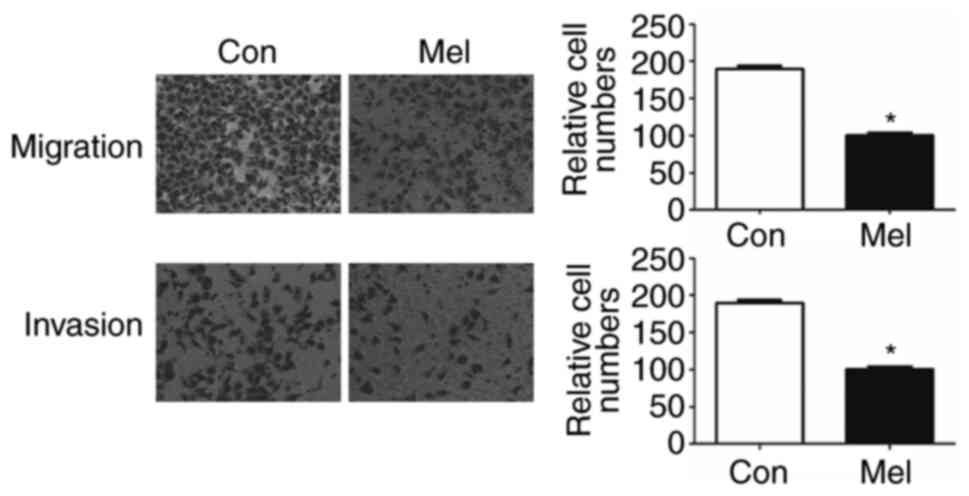
pcTERT-melittin transfection inhibits
TE1 cell migration
To identify whether transfection of pcTERT-melittin
altered the migration of TE1 cells, a Transwell assay was conducted
to assess cell migration. Migration and invasion of TE1 cells to
the lower Transwell chamber were significantly decreased in
pcTERT-melittin treated TE1 cell compared with pcTERT treated cells
after transfection for 48 h (Fig.
5). Thus, transfection of pcTERT-melittin could inhibit TE1
cell migratory and invasive ability.
pcTERT-melittin transfection induces
cell cycle arrest in TE1 cells
As a highly-ordered and tightly regulated process,
normal cell cycle progression is vital to the maintenance of cell
structure and integrity (26).
Multiple checkpoints determining extracellular growth signals, cell
size and DNA integrity are involved in cell cycle progression, and
thus dysregulation of the cell cycle is a typical features of
cancer development (27). To
determine whether cell proliferation inhibition and the
pro-apoptotic ability of pcTERT-melittin are associated with an
abnormal cell cycle, the distribution of the cell cycle was
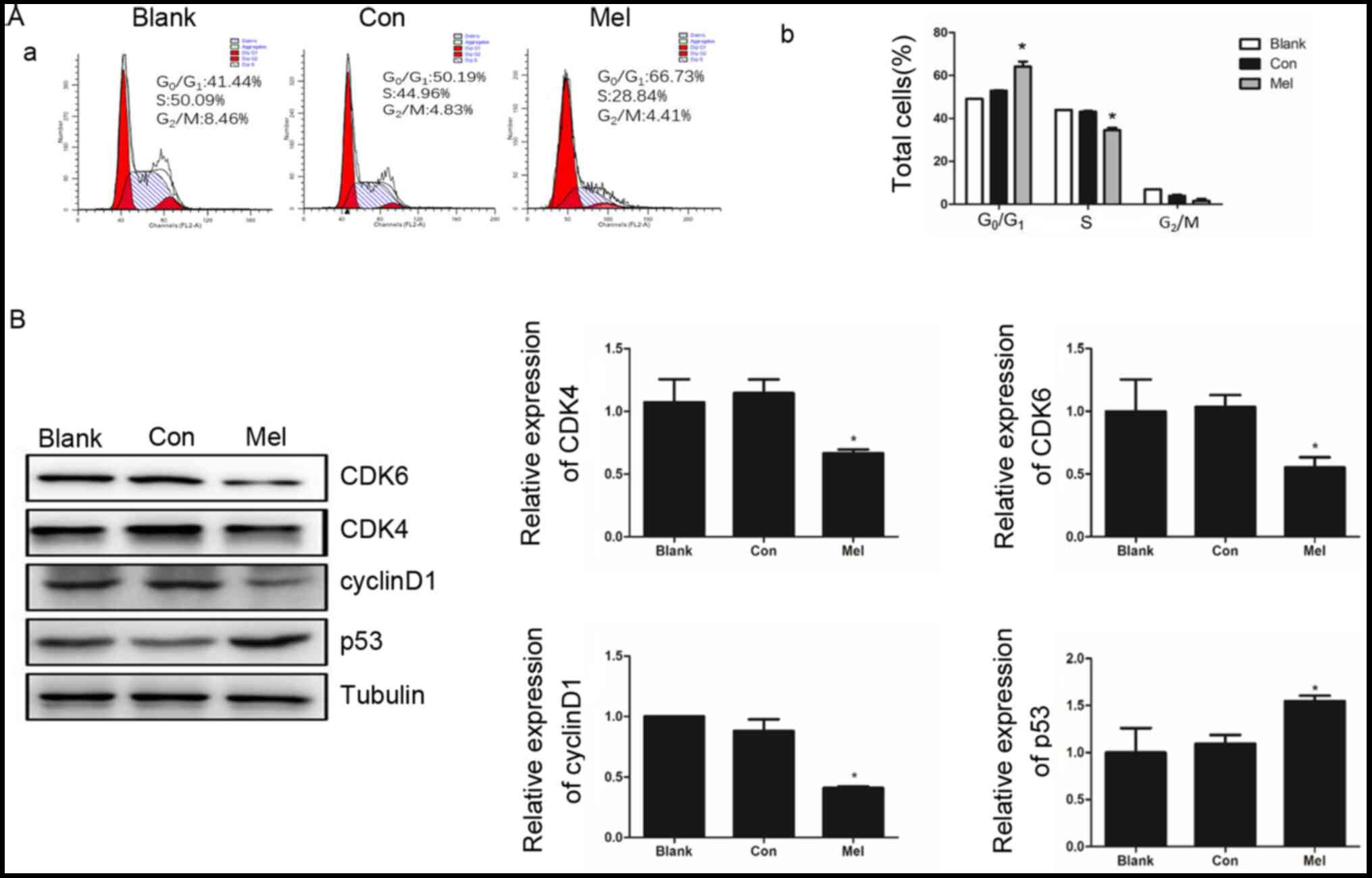
assessed using flow cytometry in TE1 cells. After transfection for
48 h the percentage of TE1 cells at phase
G0/G1 was significantly higher in
pcTERT-melittin treated cells (66.73±0.43%) compared with the
pcTERT group (50.19±0.74%) (Fig.
6A). As the cells in pcTERT-melittin group were arrested at
G0/G1 phase, the percentage of S phase cells
of pcTERT-melittin group was significant decreased (Fig. 6A).
The cyclin-dependent kinases, CDK4 and CDK6, are
important regulators of retinoblastoma phosphorylation in phase
G1/S of the cell cycle for promoting cell proliferation
(28). The activation of p53 is also
detected as a clear biological outcome of cell cycle arrest
(29). It was identified that the
expression levels of CDK4 and CDK6 were significantly decreased in
pcTERT-melittin treated cells, which was accompanied with a
significantly downregulation of cyclin D1. Furthermore, p53 was
upregulated in pcTERT-melittin treated cells after transfection for
48 h, compared with the control cells (Fig. 6B).
Discussion
Melittin is an effective anti-inflammatory compound
with activity 100 times stronger compared with that of
hydrocortisone (30). Previously,
the antineoplastic activity of melittin has received increased
attention (31,32). As a frequent cancer type, esophagus
cancer is a serious threat to human health, and is a form of
gastrointestinal tumor with a high incidence rate in China
(33). The present study used TE1
cells to investigate the apoptosis inducing capacity of the
recombinant plasmid pcTERT-melittin, which contains the melittin
coding sequence regulated by the hTERT promoter.
Changes in TE1 cell morphology were observed using
phase-contrast microscopy after transfection with the recombinant
plasmid for different intervals in the present study, and it was
identified that pcTERT-melittin treated cells exhibited distinct
apoptosis compared with controls. In addition, pcTERT-melittin
transfection induced cell shrinkage, apoptotic vacuole formation
and membrane blebbing in a dose and time-dependent manner, and
further analysis with a CCK-8 assay demonstrated proliferation
inhibition by pcTERT-melittin in TE1 cells.
There are significant changes in mitochondrial
structure during the early stage of cell apoptosis, including
increasing membrane permeability and decreasing mitochondrial
membrane potential (34). Multiple
biochemical changes accompany the decrease of ∆Ψm, such as the
release of cytochrome c into the cytoplasm and augmented ROS
production (35). The present study
used JC-1 staining to detect ∆Ψm changes, which indicated the early
stage apoptosis of TE1 cells after transfection with
pcTERT-melittin. The collapse of ∆Ψm and the increased production
of ROS suggested that the intrinsic pathway of apoptosis is the
main mechanism via which pcTERT-melittin induces apoptosis.
In the current study, quantifying apoptotic cells
using a flow cytometer demonstrated that the percentage of
apoptotic cells was significantly increased by pcTERT-melittin.
Cytochrome c can bind to the apoptotic protease activating
factor-1, recruiting and activating caspase-9 via the caspase
recruitment domain, and thus initiating the cascade reaction
(36). After transfection with
pcTERT-melittin, a significant increase in cleaved caspase-9
expression was identified in TE1 cells. PARP is associated with
numerous cell processes, including DNA repair and cell apoptosis
(37). Moreover, cleavage of PARP is
a sensitive molecular switch in apoptosis (38). In the present study, caspase-3 was
significantly activated, which was followed by cleavage of PARP in
pcTERT-melittin treated TE1 cells.
The invasion and metastasis of ESCC is not only a
common postoperative occurrence, but also causes the loss of
potential operation opportunities for numerous patients (39). In the present study, pcTERT-melittin
treated TE1 cells demonstrated a significantly reduced ability to
migrate and invade compared with controls, indicating the potential
inhibitory effects of pcTERT-melittin transfection on cancer cell
migration and invasion.
The aberrant activity of various cell cycle proteins
such as p53 and ras may result in uncontrolled tumor cell
proliferation, which is a characteristic feature of cancer
(26). In the current study, TE1
cells were arrested in the G1 phase followed by the
downregulation of CDK4, CDK6 and cyclin D1 expression levels. p53
is a pivotal protein associated with apoptosis and cell cycle
arrest. When p53 is activated, p21 is highly induced, with p21
binding to the cyclin D/CDK4 complexes that cause G1
arrest (40). The pro-apoptotic
protein Bax can also be activated by p53 in the cytosol (41). The present results demonstrated that
the expression levels of p53 and Bax were increased in TE1 cells
after transfection for 48 h.
In conclusion, the present study constructed a
recombinant plasmid pcTERT-melittin, which contained the melittin
coding sequence regulated by the hTERT promoter. It was
demonstrated that the plasmid pcTERT-melittin plasmid induced
apoptosis in TE1 cells via the mitochondrial pathway, and that did
not affect healthy esophageal cells. pcTERT-melittin also inhibited
TE1 cell migration and invasion, suggesting it may be a potential
therapeutic agent in cancer therapy. However, the present study has
some limitations, more proteins related to apoptosis, such as
proteins related to ER stress or death receptor pathway were not
investigated and the effect of pcTERT-melittin in vivo was
also not investigated. Future studies will conduct RNA-sequencing
and proteomics methods to investigate the detailed underlying
molecular mechanisms, and will construct transplantation tumor
models in mice to assess the effect of pcTERT-melittin on tumors
in vivo.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was funded by the National Natural
Science Foundation of China (grant nos. 81370497 and 81500635) and
the Science and Technology Department of Jilin Province (grant no.
20190304067YY).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding authors on reasonable
request.
Authors' contributions
PJ and JM contributed to the study conception and
design. CZ, YW, PJ and JM wrote and revised the manuscript. CZ, YL,
WZ, BX, JL, YM, YT, WW and QZ performed the experiments. PJ, WW,
WY, YL and CZ were responsible for the acquisition and analysis of
data. All authors approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Pakzad R, Mohammadian-Hafshejani A,
Khosravi B, Soltani S, Pakzed I, Mohammadian M, Salehiniya H and
Momenimovahed Z: The incidence and mortality of esophageal cancer
and their relationship to development in Asia. Ann Transl Med.
4:292016.PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Abnet CC, Arnold M and Wei WQ:
Epidemiology of esophageal squamous cell carcinoma.
Gastroenterology. 154:360–373. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu X, Zhang M, Ying S, Zhang C, Lin R,
Zheng J, Zhang G, Tian D, Guo Y, Du C, et al: Genetic alterations
in esophageal tissues from squamous dysplasia to carcinoma.
Gastroenterology. 153:166–177. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Testa U, Castelli G and Pelosi E:
Esophageal cancer: Genomic and molecular characterization, stem
cell compartment and clonal evolution. Medicines (Basel). 4:672017.
View Article : Google Scholar
|
|
6
|
Chen M, Shen M, Lin Y, Liu P, Liu X, Li X,
Li A, Yang R, Ni W, Zhou X, et al: Adjuvant chemotherapy does not
benefit patients with esophageal squamous cell carcinoma treated
with definitive chemoradiotherapy. Radiat Oncol. 13:1502018.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wu SX, Wang LH, Luo HL, Xie CY, Zhang XB,
Hu W, Zheng AP, Li DJ, Zhang HY, Xie CH, et al: Randomised phase
III trial of concurrent chemoradiotherapy with extended nodal
irradiation and erlotinib in patients with inoperable oesophageal
squamous cell cancer. Eur J Cancer. 93:99–107. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Amer MH: Gene therapy for cancer: Present
status and future perspective. Mol Cell Ther. 2:272014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li J, Liang H, Liu J and Wang Z: Poly
(amidoamine) (PAMAM) dendrimer mediated delivery of drug and
pDNA/siRNA for cancer therapy. Int J Pharm. 546:215–225. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kim HJ, Kim A, Miyata K and Kataoka K:
Recent progress in development of siRNA delivery vehicles for
cancer therapy. Adv Drug Deliv Rev. 104:61–77. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou Z, Liu X, Zhu D, Wang Y, Zhang Z,
Zhou X, Qiu N, Chen X and Shen Y: Nonviral cancer gene therapy:
Delivery cascade and vector nanoproperty integration. Adv Drug
Deliv Rev. 115:115–154. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Orsolic N: Bee venom in cancer therapy.
Cancer Metastasis Rev. 31:173–194. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rajasekaran G, Dinesh Kumar S, Nam J, Jeon
D, Kim Y, Lee CW, Park IS and Shin SY: Antimicrobial and
anti-inflammatory activities of chemokine CXCL14-derived
antimicrobial peptide and its analogs. Biochim Biophys Acta
Biomembr. 1861:256–267. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sobral F, Sampaio A, Falcão S, Queiroz MJ,
Calhelha RC, Vilas-Boas M and Ferreira IC: Chemical
characterization, antioxidant, anti-inflammatory and cytotoxic
properties of bee venom collected in Northeast Portugal. Food Chem
Toxicol. 94:172–177. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lim HN, Baek SB and Jung HJ: Bee venom and
its peptide component melittin suppress growth and migration of
melanoma cells via inhibition of PI3K/AKT/mTOR and MAPK Pathways.
Molecules. 24:9292019. View Article : Google Scholar
|
|
16
|
Rady I, Siddiqui IA, Rady M and Mukhtar H:
Melittin, a major peptide component of bee venom, and its
conjugates in cancer therapy. Cancer Lett. 402:16–31. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gajski G and Garaj-Vrhovac V: Melittin: A
lytic peptide with anticancer properties. Environ Toxicol
Pharmacol. 36:697–705. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Leao R, Apolonio JD, Lee D, Figueiredo A,
Tabori U and Castelo-Branco P: Mechanisms of human telomerase
reverse transcriptase (hTERT) regulation: Clinical impacts in
cancer. J Biomed Sci. 25:222018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ly JD, Grubb DR and Lawen A: The
mitochondrial membrane potential (deltapsi(m)) in apoptosis; an
update. Apoptosis. 8:115–128. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Perelman A, Wachtel C, Cohen M, Haupt S,
Shapiro H and Tzur A: JC-1: Alternative excitation wavelengths
facilitate mitochondrial membrane potential cytometry. Cell Death
Dis. 3:e4302012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rao XY, Huang XL, Zhou XC and Lin X: An
improvement of the 2ˆ(-delta delta CT) method for quantitative
real-time polymerase chain reaction data analysis. Biostat
Bioinforma Biomath. 3:71–85. 2013.PubMed/NCBI
|
|
22
|
Yang Y, Karakhanova S, Hartwig W, D'Haese
JG, Philippov PP, Werner J and Bazhin AV: Mitochondria and
mitochondrial ros in cancer: Novel targets for anticancer therapy.
J Cell Physiol. 231:2570–2581. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Song SB, Jang SY, Kang HT, Wei B, Jeoun
UW, Yoon GS and Hwang ES: Modulation of mitochondrial membrane
potential and ROS generation by nicotinamide in a manner
independent of SIRT1 and mitophagy. Mol Cells. 40:503–514.
2017.PubMed/NCBI
|
|
24
|
Lin M, Tang S, Zhang C, Chen H, Huang W,
Liu Y and Zhang J: Euphorbia factor L2 induces apoptosis in A549
cells through the mitochondrial pathway. Acta Pharm Sin B. 7:59–64.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wu R, Tang S, Wang M, Xu X, Yao C and Wang
S: MicroRNA-497 induces apoptosis and suppresses proliferation via
the Bcl-2/Bax-Caspase9-Caspase3 Pathway and Cyclin D2 Protein in
HUVECs. PLoS One. 11:e01670522016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Luch A: Cell cycle control and cell
division: Implications for chemically induced carcinogenesis.
Chembiochem. 3:506–516. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Otto T and Sicinski P: Cell cycle proteins
as promising targets in cancer therapy. Nature reviews. Cancer.
17:93–115. 2017.PubMed/NCBI
|
|
28
|
Sherr CJ, Beach D and Shapiro GI:
Targeting CDK4 and CDK6: From discovery to therapy. Cancer Discov.
6:353–367. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen J: The cell-cycle arrest and
apoptotic functions of p53 in tumor initiation and progression.
Cold Spring Harb Perspect Med. 6:a0261042016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lee G and Bae H: Anti-inflammatory
applications of melittin, a major component of bee venom: Detailed
mechanism of action and adverse effects. Molecules. 21:6162016.
View Article : Google Scholar
|
|
31
|
Kong GM, Tao WH, Diao YL, Fang PH, Wang
JJ, Bo P and Qian F: Melittin induces human gastric cancer cell
apoptosis via activation of mitochondrial pathway. World J
Gastroenterol. 22:3186–3195. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Soliman C, Eastwood S, Truong VK, Ramsland
PA and Elbourne A: The membrane effects of melittin on gastric and
colorectal cancer. PLoS One. 14:e02240282019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Guo LD, Chen XJ, Hu YH, Yu ZJ, Wang D and
Liu JZ: Curcumin inhibits proliferation and induces apoptosis of
human colorectal cancer cells by activating the mitochondria
apoptotic pathway. Phytother Res. 27:422–430. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bishayee K, Ghosh S, Mukherjee A,
Sadhukhan R, Mondal J and Khuda-Bukhsh AR: Quercetin induces
cytochrome-c release and ROS accumulation to promote apoptosis and
arrest the cell cycle in G2/M, in cervical carcinoma:
Signal cascade and drug-DNA interaction. Cell Prolif. 46:153–163.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hu Q, Wu D, Chen W, Yan Z, Yan C, He T,
Liang Q and Shi Y: Molecular determinants of caspase-9 activation
by the Apaf-1 apoptosome. Proc Natl Acad Sci USA. 111:16254–16261.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bai P: Biology of poly(ADP-Ribose)
polymerases: The factotums of cell maintenance. Mol Cell.
58:947–958. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Los M, Mozoluk M, Ferrari D, Stepczynska
A, Stroh C, Renz A, Herceg Z, Wang ZQ and Schulze-Osthoff K:
Activation and caspase-mediated inhibition of PARP: A molecular
switch between fibroblast necrosis and apoptosis in death receptor
signaling. Mol Biol Cell. 13:978–988. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jiang Y, Zhang J, Zhao J, Li Z, Chen H,
Qiao Y, Chen X, Liu K and Dong Z: TOPK promotes metastasis of
esophageal squamous cell carcinoma by activating the
Src/GSK3β/STAT3 signaling pathway via γ-catenin. BMC Cancer.
19:12642019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Giono LE and Manfredi JJ: The p53 tumor
suppressor participates in multiple cell cycle checkpoints. J Cell
Physiol. 209:13–20. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
McCurrach ME, Connor TM, Knudson CM,
Korsmeyer SJ and Lowe SW: Bax-deficiency promotes drug resistance
and oncogenic transformation by attenuating p53-dependent
apoptosis. Proc Natl Acad Sci USA. 94:2345–2349. 1997. View Article : Google Scholar : PubMed/NCBI
|