Introduction
The cancer stem cell (CSC) hypothesis states that
cancer originates from a small fraction of cancer cells termed CSCs
that exhibit the abilities of self-renewal and pluripotency
(1). Based on studies of leukemia
initiation, the CSC hypothesis has been used to explain the
development of a variety of solid tumors, such as lung and brain
tumors (2–4). CSCs are often identified by cell
surface markers, such as CD44 and CD24 (5,6);
however, the role of these markers remains poorly understood.
CD133, a cell surface marker, is a five-transmembrane domain
glycoprotein present in lipid rafts, the cholesterol-rich domains
on cell membranes (7). Although the
precise functions of CD133 remain unclear, it has been identified
as a candidate marker for CSCs in various types of solid tumors,
including colorectal cancer (CRC) (8–10).
The metastatic routes of cancer include
hematogenous, lymphatic and disseminated metastasis (11). These routes are important metastatic
pathways in CRC, and the liver and lungs exhibit high frequencies
of hematogenous metastasis (12). In
Japan, ~17% of all colorectal cancer cases present with distant
metastasis; 55.8% of them are liver metastases, 12.2% are lung
metastases, and 22.9% are peritoneal dissemination (13). Hypoxia is a significant factor that
contributes to the metastatic progression of cancer (14). A number of solid tumors often develop
hypoxia due to insufficient blood flow during tumor growth, which
leads to hematogenous metastasis as cancer cells form new vascular
networks to survive and attempt to escape the unfavorable
environment (15). Hypoxia-inducible
factor (HIF)-1α is a key molecule induced under hypoxic conditions
to fulfill this function (16).
HIF-1 is a central transcription factor that enhances the
transcription of various genes under hypoxic conditions and induces
angiogenesis, cell proliferation, survival and migration, thus
promoting cancer invasion and metastasis (17).
The epithelial-mesenchymal transition (EMT) is a
normal physiological process during which epithelial cells lose
their polarity and cell-cell adhesion, gain migratory and invasive
properties, and transform into mesenchymal cells (18). The EMT is essential during organ
formation and tissue growth; in cancer cells, this process enables
them to invade the surrounding tissues and metastasize to distant
sites, resulting in hematogenous metastasis (19). HIF-1α enhances the EMT in cancer
cells (14), and hypoxia triggers
the EMT in CRC cells (20).
Disseminated metastasis is a type of metastasis that
occurs when cancer cells on the primary tumor surface detach from
the tumor and adhere to other serosal surfaces such as the
peritoneum; cell adhesion molecules are involved in disseminated
metastasis (21). The integrin
family of cell adhesion molecules comprises transmembrane receptors
that facilitate cell-extracellular matrix attachment (22). Integrins have α and β subunits and
serve an important role in cancer cell attachment to the peritoneum
to induce peritoneal dissemination (21). In an animal model of surgical trauma,
blocking β1 integrin reduced the number of tumor cells that
attached to the peritoneum and subsequently impaired tumor
outgrowth (23). This suggests an
important role for β1 integrin in the adherence and proliferation
of tumor cells in the peritoneum.
Our previous study demonstrated that the
proliferative capacity of CD133+ cells was higher
compared with that of CD133− cells, whereas resistance
to chemotherapy in CD133− cells was higher compared with
that of CD133+ cells in CRC (24). In addition, patients with CRC with
liver metastasis without CD133 expression exhibited a significantly
shorter overall and disease-free survival time (25). Thus, CD133 is not only an important
CSC marker, but also serves an important role in cell proliferation
and metastasis. Compared with CD133− pancreatic cancer
cells, CD133+ pancreatic cancer cells are more likely to
induce the EMT in a hypoxic environment (26,27). In
addition, CD133 overexpression enhances the expression of HIF-1α in
head and neck squamous cell carcinoma (28), suggesting that CSCs may promote the
EMT. However, the separation of a cancer cell line into
CD133+ and CD133− cells, and a direct
comparison of their EMT-associated abilities to investigate the
involvement of CSCs in this process has not yet been reported.
Therefore, the present study isolated CD133+ and
CD133− cell populations from a single CRC cell line LoVo
and examined their EMT-associated abilities under hypoxia.
A previous study on the association between CD133
and peritoneal dissemination revealed that CD133+ cells
of an ovarian cancer cell line were more likely to adhere to
peritoneal mesothelial cells and cause peritoneal dissemination
compared with CD133− cells (29). By contrast, CD133 has been
demonstrated to not be associated with peritoneal dissemination in
CRC (30), and the role of CD133 in
peritoneal dissemination remains unclear. Our previous study
reported that the expression levels of β1 integrin in
CD133− cells were higher compared with those in
CD133+ cells (24). Our
other previous study analyzed the association between CD133
expression and postoperative recurrence of CRC in patients with
peritoneal metastasis and reported that the recurrence rate in the
CD133− group was higher compared with that in the
CD133+ group (31).
Therefore, we hypothesized that CD133+ cells may be
important for distant metastasis due to the involvement of the EMT,
whereas CD133− cells may be important for peritoneal
metastasis due to the involvement of β1 integrin.
Materials and methods
Cell culture and reagents
The human CRC cell line LoVo (Japanese Cancer
Research Resource Bank) was cultured in RPMI-1640 medium
(Sigma-Aldrich; Merck KGaA) supplemented with 10% fetal bovine
serum (Gibco; Thermo Fisher Scientific, Inc.) and a 1%
antibiotic/antimycotic solution (Gibco; Thermo Fisher Scientific,
Inc.) in a 5% CO2 incubator at 37°C (normoxia). For
hypoxia, the cells were cultured in 1% O2, 5%
CO2 and 94% N2 at 37°C in a humidified
atmosphere using a multi-gas incubator (Juji Field, Inc.).
Patients and tissue samples
Primary and metastatic tumor tissue samples were
collected from 88 consecutive patients with CRC and synchronous
liver metastasis between 1998 and 2010, and from 58 consecutive
patients with CRC and synchronous peritoneal metastasis between
1997 and 2017. These patients underwent complete resection of the
primary and metastasized tumors at the University of Tokyo Hospital
(Tokyo, Japan). None of the patients underwent preoperative chemo-
and/or radiotherapy. The study was approved by the Ethics Committee
of The University of Tokyo [approval no. 3252-(8)], and written informed consent was
obtained from all participants.
Cell isolation by magnetic cell
sorting
CD133+ and CD133− cells were
isolated from a single CRC cell line as previously described
(24). Briefly, LoVo cells
(5×107 cells in 500 µl) were labeled with a
biotin-conjugated anti-human CD133 monoclonal antibody
(CD133/1-Biotin; dilution, 1:20; cat. no. 130-091-895; Miltenyi
Biotec), washed twice with an isolation buffer and incubated with a
microbead-conjugated biotin monoclonal antibody (dilution, 1:10;
cat. no. 130-091-895; Miltenyi Biotec). The cells were isolated
using a magnetic-activated cell sorting system (Miltenyi Biotec)
according to the manufacturer's instructions. Magnetically labeled
cells were passed through a LS column with a magnetic field.
Labeled cells were isolated as the CD133+ population via
positive selection, and unlabeled cells were isolated as the
CD133− population via negative selection. Purity of the
CD133+ and CD133− cells was confirmed by flow
cytometry as described below.
Flow cytometric analysis
HIF-1α and vimentin expression was assessed by flow
cytometry as previously described (32) with minor modifications. Briefly,
isolated cells were harvested, fixed with 4% paraformaldehyde in
phosphate-buffered saline (PBS) for 30 min at 4°C, permeabilized
with 0.1% Tween-20 in PBS for 10 min at 4°C and stained using
anti-HIF-1α (dilution, 1:50; cat. no. IC1935P; R&D Systems,
Inc.) or anti-vimentin (dilution, 1:100; cat. no. 562337; BD
Pharmingen) phycoerythrin-conjugated antibodies for 30 min at 4°C.
Cell surface E-cadherin (dilution, 1:100; cat. no. 562870),
N-cadherin (dilution, 1:100; cat. no. 561554) and β1 integrin
(dilution, 1:25; cat. no. 561795) (all from BD Pharmingen)
expression levels were assessed by incubating the cells with the
antibodies for 30 min at 4°C and analyzing them using a BD
FACScalibur flow cytometer (BD Biosciences) as previously described
(20). The data were analyzed using
Cell Quest software (version 3.0, BD Biosciences) and are presented
as the mean fluorescence intensity.
Immunofluorescence analysis
LoVo cells (8×103 cells in 500 µl/well)
were plated in 24-well plates for 24 h prior to incubation under
hypoxic conditions. Then, the cells were fixed with 4%
paraformaldehyde and permeabilized with 1% Triton X-100 in PBS for
10 min at 20–25°C, followed by incubation with 3% bovine serum
albumin (Miltenyi Biotec) in PBS for blocking. The cells were
incubated overnight at 4°C with a mouse polyclonal antibody against
β-catenin (dilution, 1:500; cat. no. 610154; BD Pharmingen), washed
and incubated with an Alexa Fluor 488-conjugated anti-mouse
secondary antibody (dilution, 1:500; cat. no. 1907294, Thermo
Fisher Scientific, Inc.) for 1 h at 20–25°C. Cell nuclei were
stained using 4′,6-diamidino-2-phenylindole (Vector Laboratories,
Inc.). Images were observed and acquired using a BZ-8100 confocal
laser microscope (Keyence Corporation) at ×400 magnification in 10
fields per sample.
Migration assay
The chemokinetic activity of cells was evaluated
using a modified Boyden chamber assay as previously described
(20,33). Briefly, a polycarbonate filter with a
collagen type I coating and 8-µm pores (Ieda Trading Corporation)
was placed on a 96-well plate (Ieda Trading Corporation) containing
RPMI-1640 medium supplemented with 5% fetal bovine serum loaded in
the lower chamber. The cell suspension was prepared in RPMI-1640
medium containing 0.1% bovine serum albumin. LoVo cells
(5×105 cells in 200 µl/well) were loaded into the upper
chamber. Prior to the assay, the cells were precultured in either
hypoxic or normoxic conditions for 24 h. Two chambers were
prepared, and cells preincubated under hypoxia were incubated for
another 12 h under hypoxia, whereas those preincubated in normoxia
were incubated for another 12 h under normoxia. Subsequently, the
filters were fixed with 99% methanol and stained using a Diff-Quick
staining kit (Sysmex Corporation). The upper side of the filter was
scraped with a polyethylene blade to eliminate non-migratory cells.
The number of cells that migrated to the lower side of the filter
was measured by recording the staining intensity using a 96-well
microplate reader at 595 nm, which was defined as the migration
index.
Immunohistochemistry analysis
For analysis of CD133 expression, consecutive 3-µm
formalin-fixed paraffin-embedded sections were stained
immunohistochemically as previously described (25,31).
Following incubation with a mouse monoclonal anti-CD133 antibody
(dilution, 1:100; cat. no. 130-090-422; Miltenyi Biotec), the
Histofine SAB-PO (M) kit (Nichirei Corporation) was used to prevent
non-specific binding, secondary antibody treatment and signal
amplification. For chromogen development, the slides were incubated
in 2% 3,3′-diaminobenzidine tetrahydrochloride and 50 mM Tris
buffer (pH 7.6) containing 0.03% hydrogen peroxide. Meyer's
hematoxylin (Sigma-Aldrich; Merck KGaA) was used 30 sec for
counterstaining at 35–40°C.
A total of 1,000 tumor cells were counted manually
using ImageJ software (National Institutes of Health, Bethesda, MD,
USA). Positive CD133 expression was defined as the presence of
CD133 staining in >5% cancer cells of the primary tumor samples
(25,31). All images were evaluated
independently at ×400 magnification by two investigators who were
blinded to the clinical details of the specimens.
Statistical analyses
Data are presented as the mean ± SD. For the in
vitro experiments, HIF-1α and EMT-related marker expression
levels were compared using the unpaired two-tailed Student's
t-test. Statistical significance of migratory differences among
multiple groups was determined using two-way ANOVA followed by
Tukey's post hoc test. For the clinical study, associations between
patient characteristics and CD133 expression were determined using
the χ2 or Fisher's exact test. Statistical analyses of
tissue samples were performed using the paired Student's t-test.
All analyses were performed using JMP Pro 14.0 software (SAS
Institute). P<0.05 was considered to indicate a statistically
significant difference.
Results
HIF-1α levels and EMT markers in
CD133+ and CD133− cells under hypoxia
First, the present study investigated whether the
CD133+ CRC cells exhibited enhanced HIF-1α and
EMT-related marker expression under hypoxia. Magnetic-activated
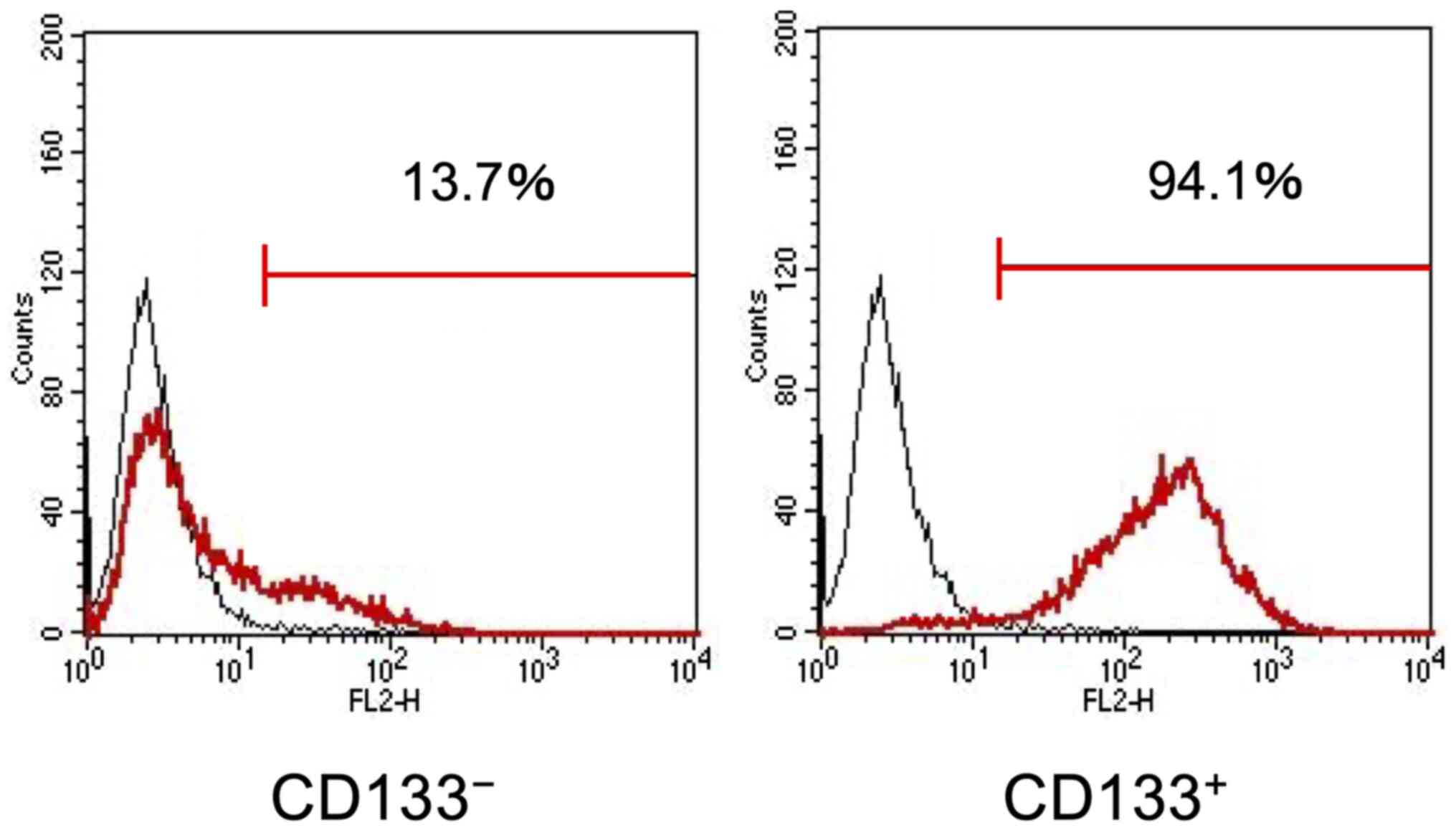
cell sorting was used to obtain the CD133+ and
CD133− LoVo cell subpopulations with purities of 94.1%
(Fig. 1). Flow cytometry analysis
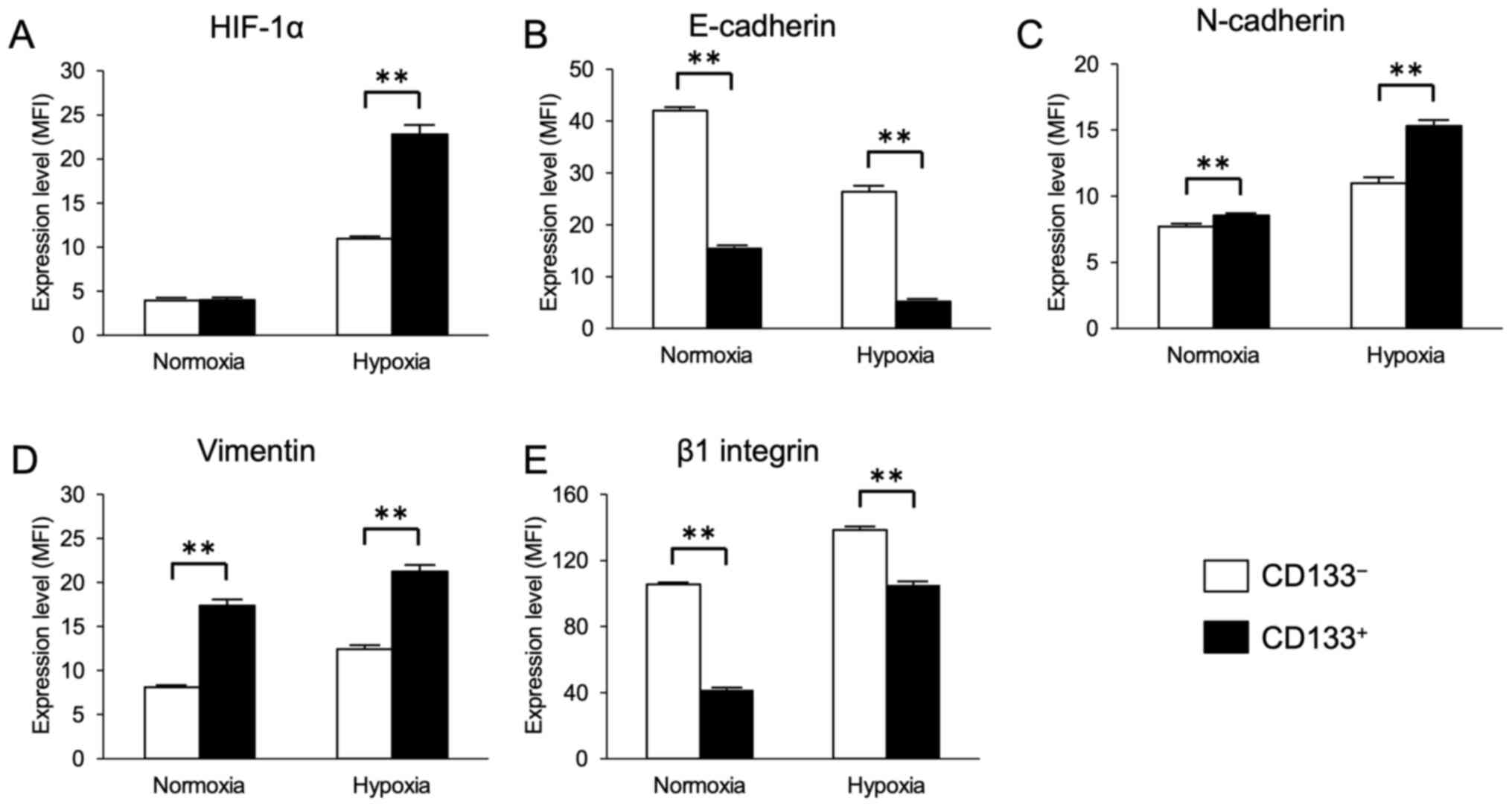
results demonstrated that the cell surface expression levels of
HIF-1α appeared to be upregulated in the CD133+ and
CD133− cells after 48-h exposure to hypoxia, and that
the expression levels of HIF-1α in the CD133+ cells were
significantly higher compared with those in the CD133−
cells (Fig. 2A).
The expression levels of the EMT markers E-cadherin,
N-cadherin and vimentin were next evaluated in the
CD133+ and CD133− cells under hypoxia. The
cell surface levels of E-cadherin expression appeared to be
decreased, whereas the levels of N-cadherin and vimentin appeared
to be increased in the CD133+ and CD133−
cells under hypoxia compared with those in the corresponding cells
under normoxia. Furthermore, under both normoxic and hypoxic
conditions, the expression levels of E-cadherin in the
CD133+ cells were significantly lower compared with
those in the CD133− cells (Fig. 2B), and the expression levels of
N-cadherin and vimentin in the CD133+ cells were
significantly higher compared with those in the CD133−
cells (Fig. 2C and D). Similarly,
the expression levels of β1 integrin appeared to be increased in
the CD133+ and CD133− cells under hypoxia
compared with those in the corresponding cells under normoxia.
However, under both normoxic and hypoxic conditions, the cell
surface levels of β1 integrin expression in the CD133+
cells were significantly lower compared with those in the
CD133− cells (Fig. 2E).
The flow cytometry histogram plots for each protein under hypoxia
are presented in Fig. S1.
Nuclear β-catenin is considered to be a marker of
the EMT (34). In the present study,
β-catenin was localized in the cytoplasm under normoxia in the
CD133+ and CD133− cells. Nuclear
translocation of β-catenin was observed only in the
CD133+ cells under hypoxia (Fig. S2).
Migratory ability of the
CD133+ and CD133− cells under hypoxia
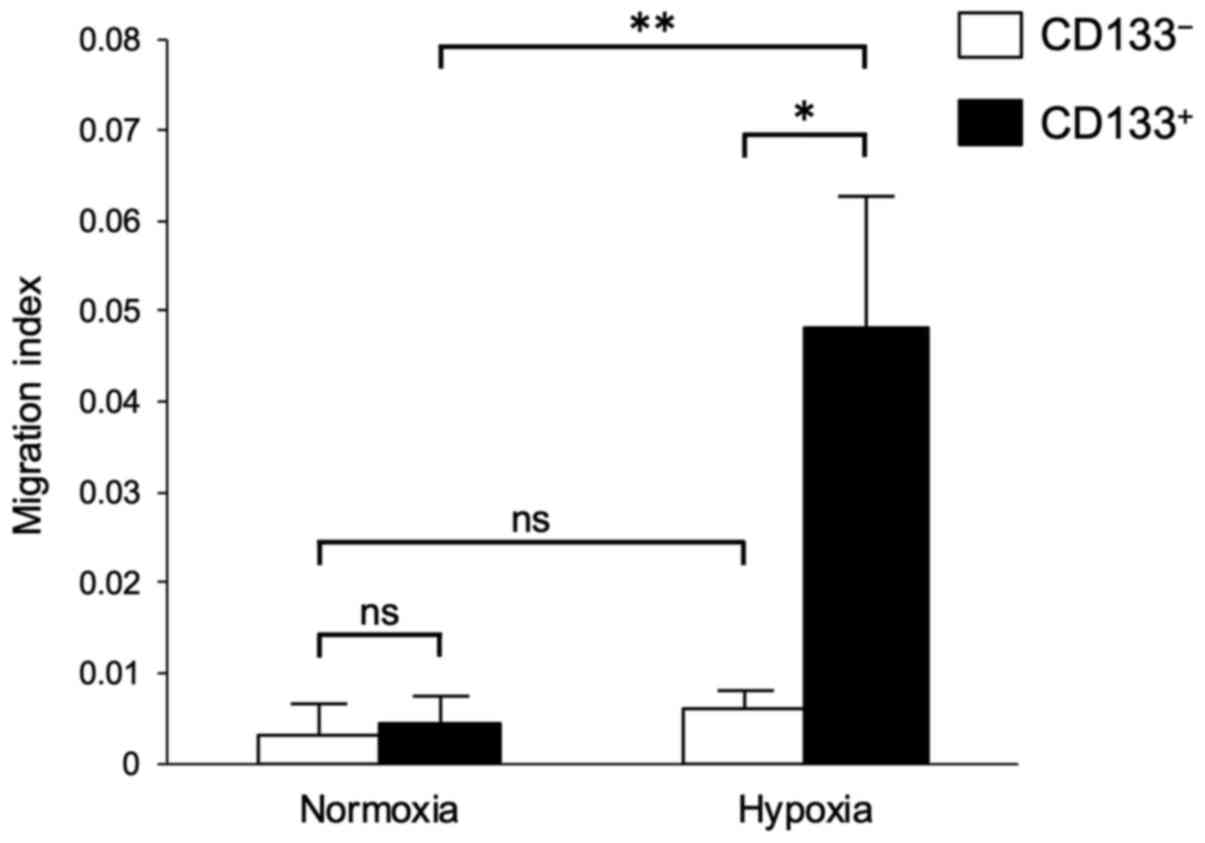
Migration assays were performed to determine the
hypoxia-induced differences in the migratory abilities of the
CD133+ and CD133− cells. No significant
differences were observed in the migratory abilities of the
CD133− cells under normoxia and hypoxia (Fig. 3). However, the migratory ability of
the CD133+ cells under hypoxia was significantly higher
compared with those of the CD133+ cells under normoxia
and the CD133− cells under hypoxia (Fig. 3).
Recovery of E-cadherin expression
after reoxygenation
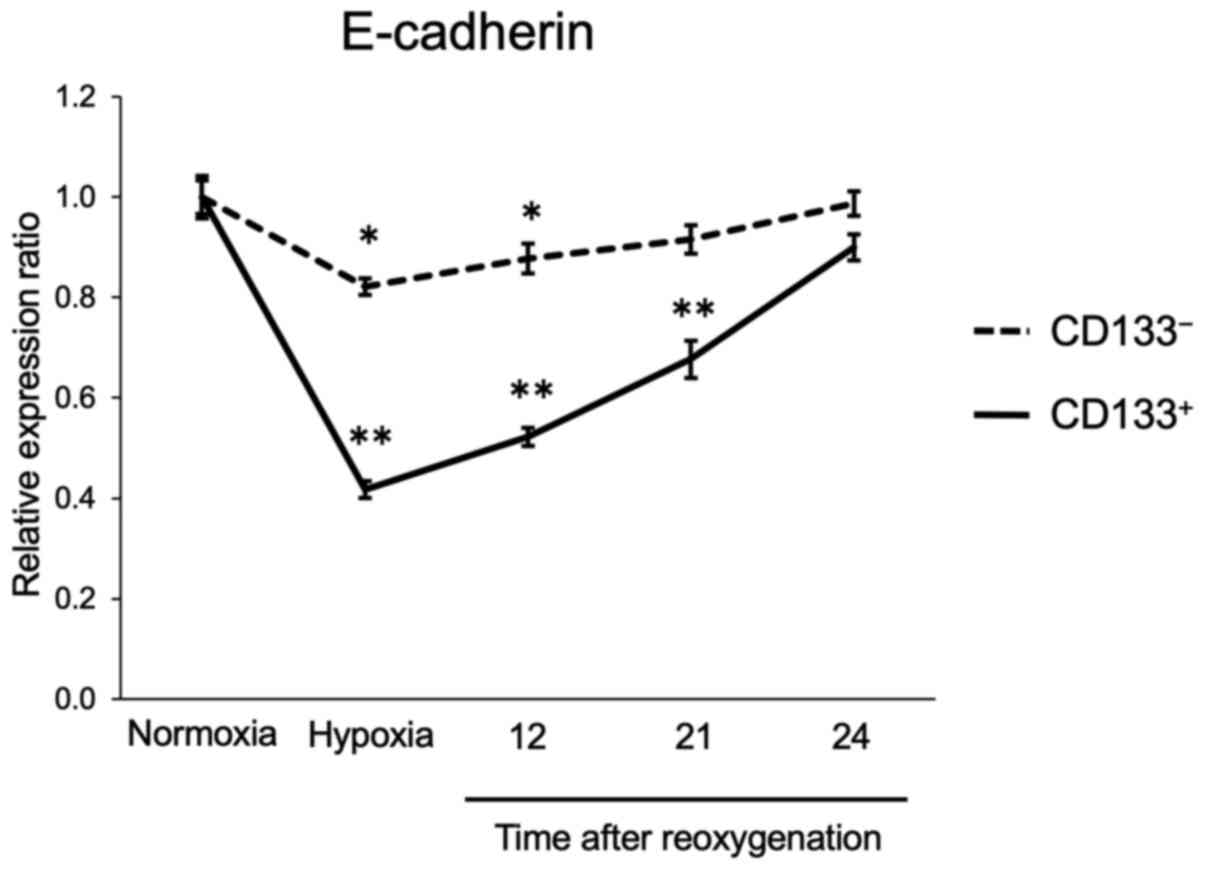
The expression of E-cadherin in the
CD133+ and CD133− cells subjected to
reoxygenation after hypoxia was next assessed. The flow cytometry
analysis results demonstrated that E-cadherin expression levels in
the CD133− cells were reduced to 82% of those in
normoxia following 24-h hypoxia; after reoxygenation, E-cadherin
levels recovered gradually and almost reached pre-hypoxia levels
within 24 h. In the CD133+ cells, E-cadherin expression
levels were reduced to 42% of those under normoxia after 24-h
hypoxia, and were restored to the pre-hypoxia levels within 24 h
(Fig. 4). The flow cytometry
histogram plots of the time course experiment are presented in
Fig. S3. These results suggested
that the CD133+ cells could recover their adhesive
ability as rapidly as the CD133− cells following
reoxygenation.
Expression of CD133 in primary tumors
and synchronous liver or peritoneal metastasis
To assess the differences in CD133 expression in
different patterns of tumor metastasis, CD133 expression levels
were determined in tumor tissues from patients with CRC with liver
or peritoneal metastasis. Immunohistochemical staining revealed
CD133 expression on the luminal cell surface of CRC tissues
(Fig. S4).
The associations between patient clinicopathological
characteristics and CD133 expression are presented in Table I. CD133 expression exhibited no
association with any clinicopathological factor, with the exception
of histopathological differentiation in individuals with
synchronous peritoneal metastasis.
 | Table I.Clinicopathological characteristics
of the patients enrolled in this study. |
Table I.
Clinicopathological characteristics
of the patients enrolled in this study.
|
| Primary tumor with
liver metastasis | Primary tumor with
peritoneal metastasis |
|---|
|
|
|
|
|---|
|
Characteristics | CD133+,
n (%) | CD133−,
n (%) | P-value | CD133+,
n (%) | CD133−,
n (%) | P-value |
|---|
| Total, n | 44 | 40 |
| 33 | 25 |
|
| Age, median years
(range) | 61 (41–81) | 59 (33–79) | 0.449 | 64 (41–80) | 68 (40–88) | 0.261 |
| Sex |
|
| 0.730 |
|
| 0.673 |
|
Male | 27 (61.4) | 26 (65.0) |
| 14 (42.4) | 12 (48.0) |
|
|
Female | 17 (38.6) | 14 (35.0) |
| 19 (57.6) | 13 (52.0) |
|
| Site of primary
tumor |
|
| 0.368 |
|
| 0.847 |
|
Right | 10 (22.7) | 6 (15.0) |
| 18 (54.5) | 13 (52.0) |
|
|
Left | 34 (77.3) | 34 (85.0) |
| 15 (45.5) | 12 (48.0) |
|
|
Differentiation |
|
| 0.084 |
|
| 0.037 |
|
High | 20 (45.5) | 14 (35.0) |
| 14 (42.4) | 6
(24.0) |
|
|
Moderate | 24 (54.5) | 22 (55.0) |
| 16 (48.5) | 10 (40.0) |
|
|
Other | 0 (0.0) | 4
(10.0) |
| 3
(12.0) | 9
(36.0) |
|
| T category |
|
| 0.896 |
|
| 0.105 |
|
T1-3 | 28 (63.7) | 26 (65.0) |
| 8
(24.2) | 2 (8.0) |
|
| T4 | 16 (36.3) | 14 (35.0) |
| 25 (75.8) | 23 (92.0) |
|
| N category |
|
| 0.641 |
|
| 0.308 |
| N0 | 13 (29.6) | 10 (25.0) |
| 9
(27.3) | 4
(16.0) |
|
|
N1-3 | 31 (70.4) | 30 (75.0) |
| 24 (72.7) | 21 (84.0) |
|
| Peritoneal cancer
index |
|
|
|
|
| 0.915 |
|
<10 |
|
|
| 22 (66.7) | 17 (68.0) |
|
|
≥10 |
|
|
| 11 (33.3) | 8
(32.0) |
|
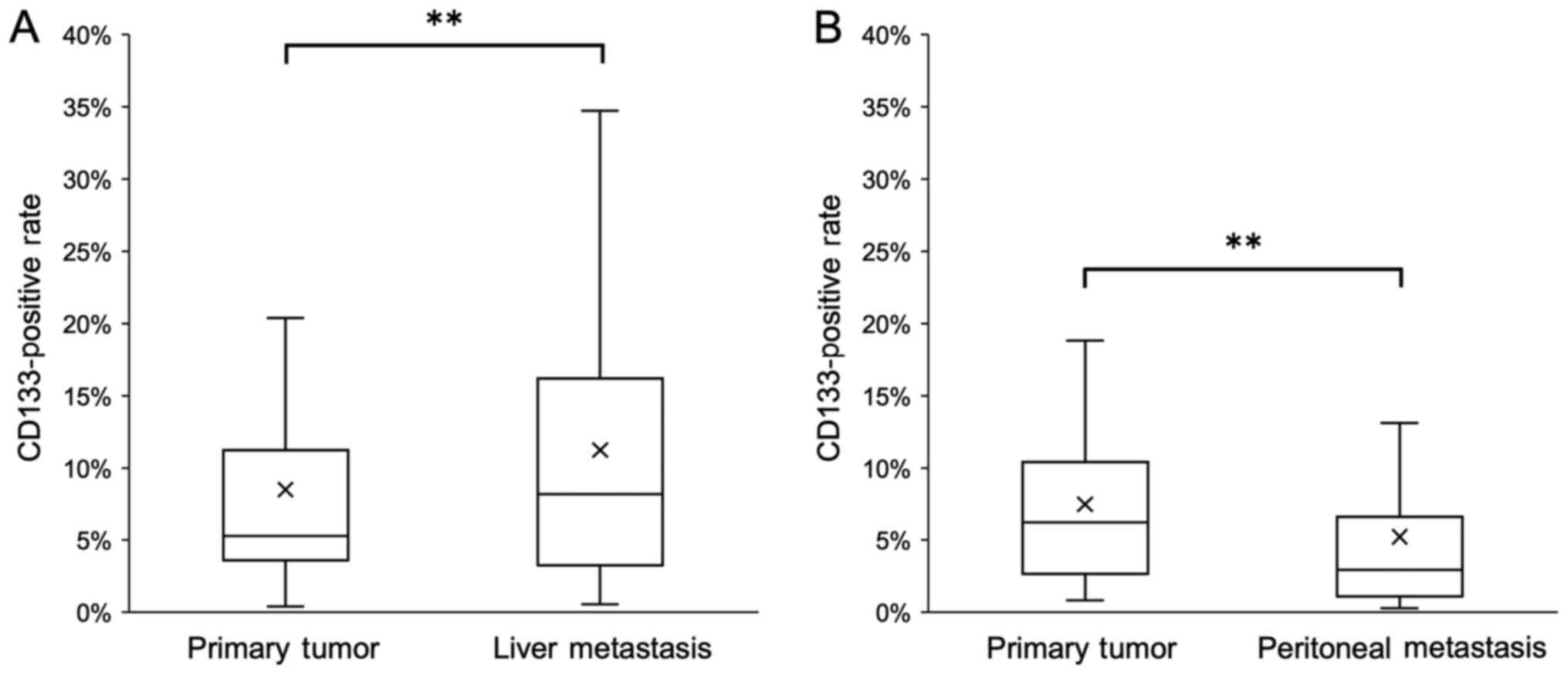
The percentage of CD133+ cells in liver
metastatic tissues was significantly higher compared with that in
the corresponding primary tumors (Fig.
5A). By contrast, the percentage of CD133+ cells in
peritoneal metastatic tissues was significantly lower compared with
that in the corresponding primary tumors (Fig. 5B).
Discussion
A hypoxic environment is essential for cancer cells
to acquire stem cell properties, and HIF pathways contribute to the
proliferation of such CSCs (35).
CD133, one of the CSC markers, has been reported to interact with
HIF-1α. For example, Maeda et al (26) have reported that CD133 knockdown in
pancreatic cancer cells inhibits HIF-1α expression. By contrast,
under hypoxic conditions, HIF-1α expression levels in glioma cells
increase, resulting in an increased proportion of CD133+
cells (36). These results suggest
that CD133 expression may be associated with that of HIF-1α, and
that these two molecules may promote each other's expression.
The present study is the first to report the
separation of a single CRC cell line into CD133+ and
CD133− cells and to directly compare the expression of
HIF-1α and EMT-associated proteins in these two populations under
hypoxic conditions. In the present study, HIF-1α expression in
hypoxic CD133+ LoVo cells was higher compared with that
in hypoxic CD133− LoVo cells. These results suggested
that CD133 may promote HIF-1α expression in CRC cells.
Nuclear translocation of β-catenin was observed in
the CD133+ cells only under hypoxia in the present
study, and significant upregulation of the levels of EMT markers
under hypoxia was observed in the CD133+ cells compared
with those in CD133− cells. Under hypoxic conditions,
HIF-1α promotes the nuclear translocation of β-catenin by
activating the Wnt signaling pathway (37). Combining the nuclear translocation of
β-catenin with HIF-1α activates the transcription of HIF-1α target
genes such as VEGF and erythropoietin, which promotes the EMT
(38). Therefore, the results of the
present study suggested that the promotion of β-catenin nuclear
translocation via CD133 and hypoxia-induced HIF-1α expression may
be the intermediary steps that enhance the expression of EMT
markers in CRC cell lines.
In the present study, although increased migration
was observed in the CD133+ cells under hypoxia compared
with that in the CD133− cells, no changes in the
migratory ability were observed in the CD133− cells.
Ding et al (39) have
reported that CD133 knockdown reduces the migratory ability of
pancreatic cancer cells, and that CD133 expression contributes to
the promotion of migration. Additionally, promotion of the EMT
enhances the invasive and migratory abilities of cancer cells
(40). In the present study, hypoxia
appeared to increase the expression of EMT markers in the
CD133+ cells and enhance the migratory ability of these
cells. Collectively, these results suggested that the promotion of
the EMT induced the migration of CRC cells.
The EMT and the mesenchymal-epithelial transition
(MET) serve important roles in cancer metastasis; although the EMT
induces distant metastasis from primary lesions, the MET causes
cancer cells in the metastatic site to revert to epithelial cells,
re-expressing E-cadherin (40). This
is considered to result in re-establishing adhesion between the
cancer cells and the surrounding tissues (41), leading to the formation of
macrometastasis (40). Accordingly,
the MET following reoxygenation is also considered to enable cancer
cells after undergoing the EMT under hypoxia to reverse the EMT and
form colonies under normoxic conditions in distant organs (42). However, the MET in CD133+
cells has not yet been reported. In the present study, although the
levels of E-cadherin expression in the CD133+ cells
appeared to be downregulated by exposure to hypoxia compared with
those under normoxia, they returned to pre-hypoxia levels 24 h
after reoxygenation. Thus, the CD133+ CRC cells not only
possessed high EMT ability, but may also achieve the MET after
reoxygenation. This phenomenon may enable them to form
macrometastasis at a metastatic site.
Integrins, which are cell adhesion molecules,
facilitate the attachment between cancer cells and the
extracellular matrix (22). The
HIF-1 pathway is one of the signaling pathways that regulate
integrin functions under hypoxic conditions (43). Hypoxia induces the transcription of
various integrin family molecules in a number of cancer cell lines,
including colorectal and breast cancer (44–46).
Previous studies have reported that integrin expression induces
CD133 expression and increases the ratio of CD133+ cells
in colorectal and liver cancer cell lines (47,48).
Thus, although the expression of a variety of integrin family
molecules contributes to CD133 expression levels, the mechanisms
underlying the interplay between CD133 and integrins remain
unclear. Our previous study demonstrated that β1 integrin
expression levels and the adhesive ability of the CD133−
LoVo cells were higher compared with those in the CD133+
LoVo cells (24). In the present
study, compared with those in the CD133+ cells, the
CD133− cells exhibited higher expression levels of β1
integrin under both normoxic and hypoxic conditions. These results
suggested that the adhesive ability of the CD133− cells
may be stronger compared with that of the CD133+ cells,
which are considered to be CSCs.
Adhesive ability serves an important role in the
peritoneal dissemination of cancer cells. Specifically, cancer
cells released from the primary lesion need to adhere to the
peritoneal surface to penetrate the peritoneum and form a colony
(21). Cell adhesion molecules, such
as β1 integrin and E-cadherin, assist in the adhesion of cancer
cells to the peritoneal surface and facilitate the interaction of
cancer cells with the extracellular matrix (21). Cancer cells with strong peritoneal
metastatic capability can upregulate the expression of β1 integrin
in gastric (49), ovarian (50) and pancreatic (51) cancer. In addition, blocking β1
integrin reduces the number of tumor cells adhering to the
peritoneum ex vivo (23). In
the present study, β1 integrin expression levels in the
CD133− cells were higher compared with those in the
CD133+ cells. These results suggest that
CD133− cells with high β1 integrin expression may be
involved in the occurrence of peritoneal metastasis.
Accordingly, we hypothesized that the
CD133+ cells may induce hematogenous metastasis owing to
their high EMT and MET recovery abilities, whereas
CD133− cells may induce peritoneal metastasis due to
their high β1 integrin content. In our previous study, positive
CD133 expression was defined as the presence of CD133 staining in
>5% of the tumor cells, and the results demonstrated that CD133
positivity in liver metastasis was slightly higher compared with
that in primary lesions (25).
However, the specific ratio of CD133+ cells could not be
evaluated in the previous study. In the present study, the
percentages of CD133+ cells in primary tumor tissues and
liver metastatic lesions were compared; the percentage of
CD133+ cells in metastatic liver tissues was
significantly higher compared with that in the corresponding
primary tumors. A previous in vivo study used a mouse liver
metastasis model to demonstrate the high liver metastatic potential
of CD133+ human lung cancer cells compared with
CD133− cells (52). In a
clinical sample-based study, CD133 mRNA expression levels in
metastatic liver lesions were significantly higher compared with
those in colorectal tumors (53),
thus corroborating the results of the present study. However, not
all liver metastatic cells were CD133+ in the present
study. The CD133+ cells may have undergone MET
transformation at the metastatic site and differentiation to
CD133− cells after liver metastasis occurred, as
reported previously (24). Thus, the
number of CD133− cells may increase with tumor growth.
This is supported by the observation that CD133 protein levels in
colorectal metastatic liver tissues decrease as the tumor grows
(54).
Our previous study demonstrated that CD133
expression levels in peritoneal metastatic tissues were lower
compared with those in primary lesions, which was contrary to the
expression pattern observed in individuals with liver metastasis
(31). However, the specific
positivity ratio was not evaluated. In the present study,
significantly lower CD133 expression levels were identified in
peritoneal metastatic tissues compared with those in the
corresponding primary tumors. Although a CD133+ ovarian
cancer cell line has been reported to be more likely to adhere to
peritoneal mesothelial cells and cause peritoneal dissemination
compared with CD133− cells (29), another study has demonstrated that
CD133 expression levels in CRC with peritoneal metastasis are lower
compared with those in CRC with liver metastasis (30). The present results also suggested
that the CD133− cells contributed to peritoneal
metastasis in CRC.
In conclusion, the results of the present study
demonstrated that the CD133+ CRC cells exhibited a high
EMT potential under hypoxic conditions, which may lead to the
development of liver metastasis. By contrast, CD133−
cells may potentiate the development of peritoneal metastasis via
expression of β1 integrin.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This study was supported by Grants-in-Aid for
Scientific Research (grant nos. 17K10620, 7K 10621, 17K10623,
18K07194, 19K09114 and 19K09115) and Challenging Research
(Exploratory) grant (grant no. 20K21626) from the Japan Society for
The Promotion of Science, and the Project for Cancer Research and
Therapeutic Evolution from the Japan Agency for Medical Research
and Development. (grant no. JP 19cm0106502).
Availability of data and materials
The datasets generated and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
KK and SI conceived of the study and contributed to
the design of the study. MO, HSh, HNa and JK carried out the
experiments with support from KK and HSo. JK, HNa and KH
contributed to sample preparation. MO, KK, HSo, HSh, HNa, HNo, KS,
MK, KM, SE, YI, HI, YY, HA and SI contributed to the interpretation
of the results. MO drafted the manuscript and designed the figures.
KK, HSo, HSh, JK, HNa, HNo, KS, MK, KM, SE, YI, HI, YY, HA, KH and
SI managed the literature search and revised the manuscript
critically. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
The University of Tokyo [approval no. 3252-(8)], and written informed consent was
obtained from all participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer, and cancer stem cells. Nature.
414:105–111. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bonnet D and Dick JE: Human acute myeloid
leukemia is organized as a hierarchy that originates from a
primitive hematopoietic cell. Nat Med. 3:730–737. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kim CF, Jackson EL, Woolfenden AE,
Lawrence S, Babar I, Vogel S, Crowley D, Bronson RT and Jacks T:
Identification of bronchioalveolar stem cells in normal lung and
lung cancer. Cell. 121:823–835. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Singh SK, Hawkins C, Clarke ID, Squire JA,
Bayani J, Hide T, Henkelman RM, Cusimano MD and Dirks PB:
Identification of human brain tumour initiating cells. Nature.
432:396–401. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li F, Tiede B, Massagué J and Kang Y:
Beyond tumorigenesis: Cancer stem cells in metastasis. Cell Res.
17:3–14. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li C, Heidt DG, Dalerba P, Burant CF,
Zhang L, Adsay V, Wicha M, Clarke MF and Simeone DM: Identification
of pancreatic cancer stem cells. Cancer Res. 67:1030–1037. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shmelkov SV, St Clair R, Lyden D and Rafii
S: AC133/CD133/Prominin-1. Int J Biochem Cell Biol. 37:715–719.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hermann PC, Huber SL, Herrler T, Aicher A,
Ellwart JW, Guba M, Bruns CJ and Heeschen C: Distinct populations
of cancer stem cells determine tumor growth and metastatic activity
in human pancreatic cancer. Cell Stem Cell. 1:313–323. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
O'Brien CA, Pollett A, Gallinger S and
Dick JE: A human colon cancer cell capable of initiating tumour
growth in immunodeficient mice. Nature. 445:106–110. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ricci-Vitiani L, Lombardi DG, Pilozzi E,
Biffoni M, Todaro M, Peschle C and De Maria R: Identification and
expansion of human colon-cancer-initiating cells. Nature.
445:111–115. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Paduch R: The role of lymphangiogenesis
and angiogenesis in tumor metastasis. Cell Oncol (Dordr).
39:397–410. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Japanese Society for Cancer of the Colon
and Rectum: Japanese Classification of Colorectal, Appendiceal, and
Anal Carcinoma: The 3d English Edition [Secondary Publication]. J
Anus Rectum Colon. 3:175–195. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hashiguchi Y, Muro K, Saito Y, Ito Y,
Ajioka Y, Hamaguchi T, Hasegawa K, Hotta K, Ishida H, Ishiguro M,
et al Japanese Society for Cancer of the Colon Rectum, : Japanese
Society for Cancer of the Colon and Rectum (JSCCR) guidelines 2019
for the treatment of colorectal cancer. Int J Clin Oncol. 25:1–42.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gilkes DM, Semenza GL and Wirtz D: Hypoxia
and the extracellular matrix: Drivers of tumour metastasis. Nat Rev
Cancer. 14:430–439. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pennacchietti S, Michieli P, Galluzzo M,
Mazzone M, Giordano S and Comoglio PM: Hypoxia promotes invasive
growth by transcriptional activation of the met protooncogene.
Cancer Cell. 3:347–361. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Semenza GL and Wang GL: A nuclear factor
induced by hypoxia via de novo protein synthesis binds to the human
erythropoietin gene enhancer at a site required for transcriptional
activation. Mol Cell Biol. 12:5447–5454. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Semenza GL: Targeting HIF-1 for cancer
therapy. Nat Rev Cancer. 3:721–732. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chaffer CL and Weinberg RA: A perspective
on cancer cell metastasis. Science. 331:1559–1564. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hongo K, Tsuno NH, Kawai K, Sasaki K,
Kaneko M, Hiyoshi M, Murono K, Tada N, Nirei T, Sunami E, et al:
Hypoxia enhances colon cancer migration and invasion through
promotion of epithelial-mesenchymal transition. J Surg Res.
182:75–84. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
de Cuba EM, Kwakman R, van Egmond M, Bosch
LJ, Bonjer HJ, Meijer GA and te Velde EA: Understanding molecular
mechanisms in peritoneal dissemination of colorectal cancer: Future
possibilities for personalised treatment by use of biomarkers.
Virchows Arch. 461:231–243. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Giancotti FG and Ruoslahti E: Integrin
signaling. Science. 285:1028–1032. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Oosterling SJ, van der Bij GJ, Bögels M,
ten Raa S, Post JA, Meijer GA, Beelen RH and van Egmond M:
Anti-beta1 integrin antibody reduces surgery-induced adhesion of
colon carcinoma cells to traumatized peritoneal surfaces. Ann Surg.
247:85–94. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hongo K, Tanaka J, Tsuno NH, Kawai K,
Nishikawa T, Shuno Y, Sasaki K, Kaneko M, Hiyoshi M, Sunami E, et
al: CD133(−) cells, derived from a single human colon cancer cell
line, are more resistant to 5-fluorouracil (FU) than CD133(+)
cells, dependent on the β1-integrin signaling. J Surg Res.
175:278–288. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kishikawa J, Kazama S, Oba K, Hasegawa K,
Anzai H, Harada Y, Abe H, Matsusaka K, Hongo K, Oba M, et al: CD133
expression at the metastatic site predicts patients' outcome in
colorectal cancer with synchronous liver metastasis. Ann Surg
Oncol. 23:1916–1923. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Maeda K, Ding Q, Yoshimitsu M, Kuwahata T,
Miyazaki Y, Tsukasa K, Hayashi T, Shinchi H, Natsugoe S and Takao
S: CD133 modulate HIF-1alpha expression under hypoxia in EMT
phenotype pancreatic cancer stem-like cells. Int J Mol Sci.
17:10252016. View Article : Google Scholar
|
|
27
|
Salnikov AV, Liu L, Platen M, Gladkich J,
Salnikova O, Ryschich E, Mattern J, Moldenhauer G, Werner J,
Schemmer P, et al: Hypoxia induces EMT in low and highly aggressive
pancreatic tumor cells but only cells with cancer stem cell
characteristics acquire pronounced migratory potential. PLoS One.
7:e463912012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen YS, Wu MJ, Huang CY, Lin SC, Chuang
TH, Yu CC and Lo JF: CD133/Src axis mediates tumor initiating
property and epithelial-mesenchymal transition of head and neck
cancer. PLoS One. 6:e280532011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mitsui H, Shibata K, Suzuki S, Umezu T,
Mizuno M, Kajiyama H and Kikkawa F: Functional interaction between
peritoneal mesothelial cells and stem cells of ovarian yolk sac
tumor (SC-OYST) in peritoneal dissemination. Gynecol Oncol.
124:303–310. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Neumann J, Löhrs L, Albertsmeier M, Reu S,
Guba M, Werner J, Kirchner T and Angele M: Cancer stem cell markers
are associated with distant hematogenous liver metastases but not
with peritoneal carcinomatosis in colorectal cancer. Cancer Invest.
33:354–360. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nagata H, Ishihara S, Kishikawa J, Sonoda
H, Murono K, Emoto S, Kaneko M, Sasaki K, Otani K, Nishikawa T, et
al: CD133 expression predicts post-operative recurrence in patients
with colon cancer with peritoneal metastasis. Int J Oncol.
52:721–732. 2018.PubMed/NCBI
|
|
32
|
Kawai K, Tsuno NH, Kitayama J, Okaji Y,
Yazawa K, Asakage M, Hori N, Watanabe T, Takahashi K and Nagawa H:
Epigallocatechin gallate, the main component of tea polyphenol,
binds to CD4 and interferes with gp120 binding. J Allergy Clin
Immunol. 112:951–957. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Iida Y, H Tsuno N, Kishikawa J, Kaneko K,
Murono K, Kawai K, Ikeda T, Ishihara S, Yamaguchi H, Sunami E, et
al: Lysophosphatidylserine stimulates chemotactic migration of
colorectal cancer cells through GPR34 and PI3K/Akt pathway.
Anticancer Res. 34:5465–5472. 2014.PubMed/NCBI
|
|
34
|
Banyard J and Bielenberg DR: The role of
EMT and MET in cancer dissemination. Connect Tissue Res.
56:403–413. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Keith B and Simon MC: Hypoxia-inducible
factors, stem cells, and cancer. Cell. 129:465–472. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Soeda A, Park M, Lee D, Mintz A,
Androutsellis-Theotokis A, McKay RD, Engh J, Iwama T, Kunisada T,
Kassam AB, et al: Hypoxia promotes expansion of the CD133-positive
glioma stem cells through activation of HIF-1alpha. Oncogene.
28:3949–3959. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Vu T and Datta PK: Regulation of EMT in
colorectal cancer: a culprit in metastasis. Cancers (Basel).
9:1712017. View Article : Google Scholar
|
|
38
|
Valenta T, Hausmann G and Basler K: The
many faces and functions of β-catenin. EMBO J. 31:2714–2736. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ding Q, Miyazaki Y, Tsukasa K, Matsubara
S, Yoshimitsu M and Takao S: CD133 facilitates
epithelial-mesenchymal transition through interaction with the ERK
pathway in pancreatic cancer metastasis. Mol Cancer. 13:152014.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Thiery JP: Epithelial-mesenchymal
transitions in tumour progression. Nat Rev Cancer. 2:442–454. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bukholm IK, Nesland JM and Børresen-Dale
AL: Re-expression of E-cadherin, alpha-catenin and beta-catenin,
but not of gamma-catenin, in metastatic tissue from breast cancer
patients [seecomments]. J Pathol. 190:15–19. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Manzo G: Similarities between embryo
development and cancer process suggest new strategies for research
and therapy of tumors: a new point of view. Front Cell Dev Biol.
7:202019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ata R and Antonescu CN: Integrins and cell
metabolism: an intimate relationship impacting cancer. Int J Mol
Sci. 18:1892017. View Article : Google Scholar
|
|
44
|
Koike T, Kimura N, Miyazaki K, Yabuta T,
Kumamoto K, Takenoshita S, Chen J, Kobayashi M, Hosokawa M,
Taniguchi A, et al: Hypoxia induces adhesion molecules on cancer
cells: A missing link between Warburg effect and induction of
selectin-ligand carbohydrates. Proc Natl Acad Sci USA.
101:8132–8137. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ju JA, Godet I, Ye IC, Byun J, Jayatilaka
H, Lee SJ, Xiang L, Samanta D, Lee MH, Wu PH, et al: Hypoxia
selectively enhances integrin α5β1 receptor expression in breast
cancer to promote metastasis. Mol Cancer Res. 15:723–734. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ryu MH, Park HM, Chung J, Lee CH and Park
HR: Hypoxia-inducible factor-1alpha mediates oral squamous cell
carcinoma invasion via upregulation of alpha5 integrin and
fibronectin. Biochem Biophys Res Commun. 393:11–15. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wu X, Cai J, Zuo Z and Li J: Collagen
facilitates the colorectal cancer stemness and metastasis through
an integrin/PI3K/AKT/ Snail signaling pathway. Biomed Pharmacother.
114:1087082019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Chen WC, Chang YS, Hsu HP, Yen MC, Huang
HL, Cho CY, Wang CY, Weng TY, Lai PT, Chen CS, et al: Therapeutics
targeting CD90-integrin-AMPK-CD133 signal axis in liver cancer.
Oncotarget. 6:42923–42937. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Nakashio T, Narita T, Akiyama S, Kasai Y,
Kondo K, Ito K, Takagi H and Kannagi R: Adhesion molecules and
TGF-beta1 are involved in the peritoneal dissemination of NUGC-4
human gastric cancer cells. Int J Cancer. 70:612–618. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Strobel T and Cannistra SA:
Beta1-integrins partly mediate binding of ovarian cancer cells to
peritoneal mesothelium in vitro. Gynecol Oncol. 73:362–367. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Hosono J, Narita T, Kimura N, Sato M,
Nakashio T, Kasai Y, Nonami T, Nakao A, Takagi H and Kannagi R:
Involvement of adhesion molecules in metastasis of SW1990, human
pancreatic cancer cells. J Surg Oncol. 67:77–84. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhang H, Yang N, Sun B, Jiang Y, Hou C, Ji
C, Zhang Y, Liu Y and Zuo P: CD133 positive cells isolated from
A549 cell line exhibited high liver metastatic potential.
Neoplasma. 61:153–160. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Jing F, Kim HJ, Kim CH, Kim YJ, Lee JH and
Kim HR: Colon cancer stem cell markers CD44 and CD133 in patients
with colorectal cancer and synchronous hepatic metastases. Int J
Oncol. 46:1582–1588. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Huang X, Sheng Y and Guan M: Co-expression
of stem cell genes CD133 and CD44 in colorectal cancers with early
liver metastasis. Surg Oncol. 21:103–107. 2012. View Article : Google Scholar : PubMed/NCBI
|