Introduction
Gastric cancer (GC) is the third leading cause of
cancer-associated death and the fourth most prevalent malignancy
worldwide, accounting for 8% of total cancer incidence and 10% of
cancer-associated deaths. There were ~21,320 cases of GC (13,020
men and 8,300 women) and 10,540 patients died from GC in 2012 in
the United States (1). Patients with
advanced GC have a poor prognosis (2,3). The
identification of non-invasive tumor markers that can detect the
early stages of cancer, or assess the risk of cancer development in
individuals may decrease the mortality rate of GC (4). However, the mechanism underlying GC
development is not yet fully understood, and further research is
required in order to clarify its pathogenesis and potential
molecular targets.
The gene encoding CD90 (also known as THY1 or Thy-1
Cell Surface Antigen), is located on human chromosome 11q23.3, and
encodes a glycosylphosphatidylinositol-linked cell surface
glycoprotein (5–8). CD90 regulates key biological processes,
such as cell apoptosis, migration, adhesion, cell-cell and
cell-matrix interactions and T-cell activation (5–8).
Furthermore, numerous reports have demonstrated that CD90 has key
functions in regulating cell proliferation, angiogenesis and
metastasis in cancer (9–11). CD90 is a prognostic marker in certain
types of cancer (12,13). CD90 is upregulated in GC tissues and
modulates the expression levels of osteonectin to inhibit the
apoptosis of GC cells (14).
However, the precise mechanism underlying the effects of CD90 in GC
remains unclear.
Non-cancerous cells produce energy primarily via
oxidative phosphorylation in mitochondria. Tumor cells, even under
aerobic conditions, generate energy primarily through glycolytic
pathways (15). During glycolysis,
glucose is converted to lactic acid (LD) (16,17). A
large number of intermediate products promote cell proliferation.
As a result, glucose consumption increases, leading to anaerobic
glycolysis (18). The metabolic
rates of glucose, lactate, pyruvate, glutamine and fatty acids in
tumor cells are notably higher than those in normal cells. Their
complex metabolic properties allow tumor cells to generate ATP,
maintaining a balance of redox states, and providing energy for
survival, growth and proliferation (19,20).
Song et al (21) identified
intermediate products of aerobic glycolysis pathways, such as
increased expression levels of α-ketoglutarate and fumaric acid in
cancer tissues, indicating that abnormal changes in glucose
metabolism could be used to distinguish between GC cells and normal
cells. Similarly, previous studies have demonstrated abnormal
glucose metabolism is present in GC cells (22,23).
The present study aimed to investigate the effect of
CD90 on biological behaviors, such as proliferation, cell
senescence, and invasion and migration ability in GC cells. Changes
in LD concentration, lactate dehydrogenase (LDH) levels, ATP
concentrations and the NADPH/NADP+ ratio were assessed
following induction of CD90 overexpression in GC cells. The present
study also investigated the mechanism underlying the effects of
CD90 in GC.
Materials and methods
Cell culture and transfection
The human GC cell line AGS was acquired from the
Chinese Academy Medical Science (Beijing, China). Cells were
cultured in Ham's/F-12 (HyClone; Cytiva) with 10% FBS (Gibco;
Thermo Fisher Scientific, Inc.), 100 µg/ml streptomycin and 100
U/ml penicillin (GE Healthcare Life Sciences), at 37°C with 5%
CO2. The coding region of the CD90 gene was generated
via PCR using the following primer pair:
5′-ATACTCGAATGAACCTGGCCATCAGCAT-3′ and
5′-GCGGAATTCTCACAGGGACATGAAATCCG-3′. PCR was performed under the
following conditions: One cycle for 5 min at 94°C; 30 cycles for 45
sec at 94°C, 45 sec at 55°C and 90 sec at 72°C; and final extension
for 10 min at 72°C. The fragments were cloned into the TA vector
(Promega Corporation) and used to transform E. coli JM109
(Takara Biotechnology Co., Ltd.) according to the manufacturers'
instructions. Following selection and propagation, the pure plasmid
DNA was prepared using standard methods (24). The DNA fragments (498 bp) were
removed from the TA vector using restriction enzyme digestion with
XhoI and EcoR1 (Promega Corporation) and then
subcloned into the pEGFP-N1 vector. The brief steps are as follows:
The pEGFP-N1 vector was also treated with restriction enzyme
digestion with XhoI and EcoR1 (Promega Corporation),
the linear pEGFP-N1 vector was purified, and then the DNA fragments
(498 bp) were linked with it to obtain the pEGFP vector with this
DNA fragment. The fusion sequences were verified via DNA sequencing
using ABI 3730 (Sigma-Aldrich; Merck KGaA). In order to establish a
stable CD90-expressing cell line, plasmid pEGFP-N1/CD90 or control
empty vector pEGFP-N1 was transfected into AGS cells using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol and as
previously described (14,25,26). In
order to obtain stable CD90-overexpressing cells, 1,000 µg/ml G418
(Sigma-Aldrich; Merck KGaA) was added 48 h after transfection; this
level was sustained for >14 days. The maintenance concentration,
500 µg/ml G418, was used throughout the cell culture. CD90
overexpression was verified using western blotting.
Cell Counting Kit-8 (CCK-8) assay
Cell proliferation was detected using a CCK-8 assay
(7 Sea Biotech Co., Ltd.) according to the manufacturer's
instructions. Cell suspension (200 µl) was seeded in 96-well cell
culture plates at a density of 1,000 cells/well and incubated at
37°C for 6 days. Cell density was measured every 24 h. A total of
20 µl CCK-8 solution was added to each well and incubated for 2 h
at 37°C. Finally, the absorbance was measured at 450 nm using a
Paradigm Detection System (Beckman Coulter, Inc.).
β-Galactosidase staining
Cells were washed three times in a 6-well plate with
1X PBS and fixed using β-galactosidase staining stationary solution
(cat. no. C0602; Beyotime Institute of Biotechnology) at room
temperature for 15 min. The plate was rinsed twice with 1X PBS, and
1 ml staining solution (10 µl each of β-galactosidase staining
solution A and B, 930 µl β-galactosidase staining solution C and 50
µl X-gal solution) was added. The plate was incubated overnight at
37°C without CO2. Cellular senescence was detected using
a Senescence-associated β-Galactosidase Staining kit (cat. no.
C0602; Beyotime Institute of Biotechnology). Images were captured
using a light microscope (magnification, ×10; cat. no. CKX41;
Olympus Corporation). Blue β-galactosidase positive cells were
considered to be senescent.
Cell invasion and migration
assays
A Corning Matrigel invasion chamber in 24-well
plates with 8-µm pores (Corning Life Sciences) and standard
Transwell inserts (6.5 mm) were used to perform the cell invasion
assays. In total, 20 µl Matrigel was used for per well. A total of
2.5×104 cells were suspended in 500 µl of serum-free
medium (Hyclone; Cyvita) and added to the top chamber. A total of
750 µl Ham's/F-12 growth medium (Hyclone; Cyvita) with 20% FBS was
added to the lower chamber. Cells were then allowed to invade onto
the lower surface of the upper chamber following incubation in a
humidified incubator with 5% CO2 at 37°C, for 48 h. The
cells were fixed with 4% paraformaldehyde for 10 min at room
temperature, and stained with 0.1% crystal violet at room
temperature for 10 min. The cells on the upper surface of the upper
chamber were removed using a cotton swab. Images were captured
using a light microscope at ×100 magnification. Cells were counted
from at least five randomly selected fields for each experiment.
Falcon Cell Culture Inserts with 8-µm pores (Corning Life Sciences)
were used to perform cell migration assay using the aforementioned
method, but without Matrigel coating on the upper surface of the
Transwell filters.
LD assay
A total of ~5×105 cells were seeded and
cultured for 48 h for each group. The LD content in the cells and
media were detected. Ultrasonic fragmentation (40 kHz, for 5 sec
and stopped for 1 sec, on ice) was used to produce the cell
homogenate, and reagents were added according to the manufacturer's
instructions (Nanjing Jiancheng Bioengineering Institute).
Absorbance was measured at 721 nm using the Paradigm Detection
system (Beckman Coulter, Inc.). Finally, the protein concentration
was determined to calibrate the LD value in each group.
LDH detection
A total of ~1×106 cells were seeded and
cultured at 37°C for 48 h for each group. The LDH content in the
cells and media were detected. Ultrasonic fragmentation (40 kHz for
5 sec and stopped for 1 sec on ice) was used to produce the cell
homogenate, and then reagents were added according to the
manufacturer's instructions (Nanjing Jiancheng Bioengineering
Institute). Absorbance was measured at 440 nm using the Paradigm
Detection system. Finally, the protein concentration was determined
and used to calibrate the LD value in each group.
NADP+/NADPH assay
In order to extract NADP+, the Coenzyme
II NADP (H) content test kit was used (cat. no. A115-1-1; Nanjing
Jiancheng Bioengineering Institute). In total, 1 ml of acidic
extract was added to 5,000,000 cells. Cells were then sonicated (at
40% for 5 sec and stopped for 1 sec on ice) for 1 min, boiled for 5
min, cooled in an ice bath and centrifuged at 10,000 × g at 4°C for
10 min. The supernatant was retained and an equal volume of alkali
extract was added to neutralize it. The solution was mixed and
centrifuged at 10,000 × g for 10 min, followed by the addition of
1–7 reagents from the aforementioned kit then mixing. Absorbance
was measured at 570 nm. The NADP+ content was calculated
as: 4.570×(ΔA-0.062)/Cpr, where ΔA is the absorbance value of
measuring tube-contrast tube absorbance value, and Cpr is the
sample protein concentration. In order to extract NADPH, 1 ml
alkali extract was added to 5,000,000 cells. Cells were sonicated
for 1 min (40 kHz for 5 sec and stopped for 1 sec on ice), boiled
for 5 min, cooled in an ice bath and centrifuged at 10,000 × g at
4°C for 10 min. The supernatant was retained, and the same volume
of acid extract as the supernatant was added to neutralize it. The
solution was mixed and centrifuged at 10,000 × g 4°C for 10 min,
followed by the addition of 1–7 reagents from the aforementioned
kit and mixing. Absorbance was measured at 570 nm as
aforementioned. The NADPH content was calculated as:
7.200×(ΔA-0.072)/Cpr, where ΔA is the absorbance value of measuring
tube-contrast tube absorbance value, and Cpr is the sample protein
concentration.
ATP detection
An ATP content test kit (cat. no. S0026; Beyotime
Institute of Biotechnology) was used to detect the ATP content
according to the manufacturer's instructions. Relative fluorescence
unit values were determined using a Luminometer (Beckman Coulter,
Inc.). A standard curve was constructed and the ATP concentration
of the sample was calculated. Protein concentrations were then
assessed. The concentration of ATP was divided by the protein
concentration and the units were converted to nmol/mg protein.
Western blotting analysis
CD90-overexpressing AGS cells (AGS-CD90) and control
AGS cells (AGS/vector) were lysed in RIPA buffer (CoWin
Biosciences), and a bicinchoninic acid kit (Thermo Fisher
Scientific, Inc.) was used to determine the protein concentration.
A total 50 µg sample protein was added to 5X loading buffer, boiled
for 5 min, then separated using 10% SDS-PAGE gel and transferred
onto nitrocellulose membranes (HyClone; Cytiva). The nitrocellulose
membranes were incubated at 4°C with TBST ((25 mM Tris-HCl, 150 mM
NaCl, pH 7.5 and 0.05% Tween-20) containing 5% non-fat milk,
followed by overnight incubation at 4°C with the following primary
antibodies: Rabbit anti-CD90, anti-PI3K, anti-PDK1, mouse anti-AKT,
rabbit anti-phosphorylated (p)-AKT-Ser473, anti-HIF-1α, mouse
anti-MDM2, mouse anti-PTEN, rabbit anti-p53, mouse anti-Sirtuin
(SIRT) 1, mouse anti-SIRT 2, rabbit anti-SIRT 3 and mouse anti-SIRT
6 (all 1:1,000; cat. nos. YT0783, YT3709, YT3645, YT0173, YP0006,
YT2133, YT2692, YT5752, YT3528, YM1416, YM1271, YT4304 and YM1274,
respectively; all ImmunoWay Biotechnology Company). After washing
three times with PBS and incubation with labeled secondary
antibodies, including anti-rabbit secondary antibody (1:3,000; cat.
no. sc-2004) and anti-mouse secondary antibody (1:3,000; cat. no.
sc-2005) (both Santa Cruz Biotechnology, Inc.), signals were
visualized using an enhanced chemiluminescence detection system
(EMD Millipore). Rabbit anti-β-tubulin (1:5,000; cat. no. T0028;
Affinity Biosciences) was used as a loading control.
Statistical analysis
The data are presented as the mean ± standard error
of the mean from three independent experiments. Differences in
parametric variables were analyzed using one-way ANOVA followed by
Student-Newman-Keuls post hoc test, and differences in
non-parametric variables between groups were analyzed by
Kruskal-Wallis test followed by Bonferroni's. Differences of
non-parametric variables were analyzed using Mann Whitney U tests.
Differences between two quantitative variable groups were analyzed
using unpaired Student's t-tests and GraphPad Prism version 5.01
(GraphPad Software). All other statistical analyses was performed
using SPSS (version 11.0; SPSS, Inc.). P<0.05 was considered to
indicate a statistically significant difference.
Results
CD90 promotes proliferation of GC
cells
A previous study demonstrated that expression levels
of CD90 are higher in GC tissues compared with adjacent normal
tissues (9). In order to investigate
the effect and potential mechanism of CD90 in GC, a
CD90-overexpressing AGS cell line was established, using G418 to
select the positive clones. Western blotting confirmed that CD90
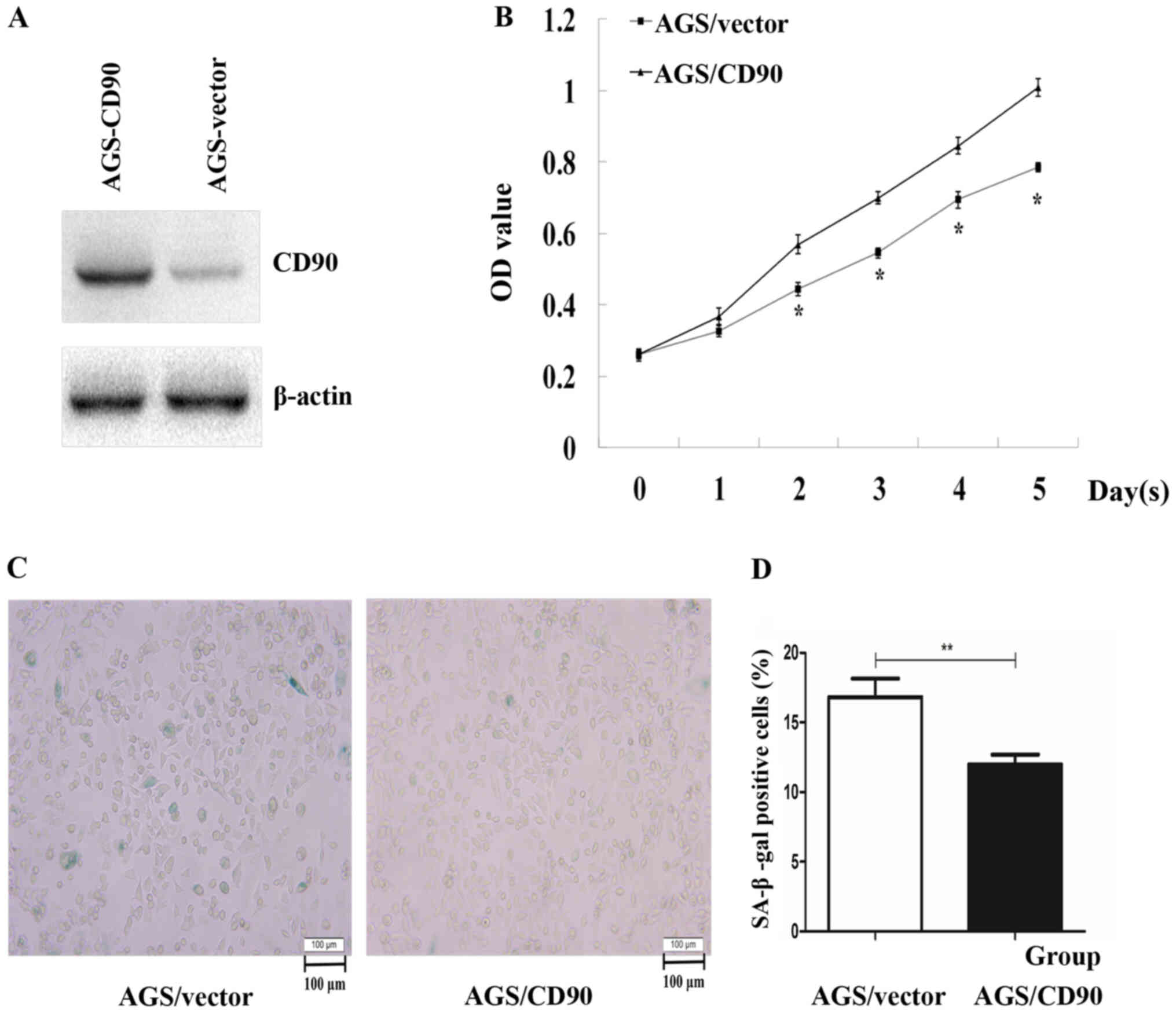
was stably and highly expressed in AGS-CD90 cells (Fig. 1A). In order to verify whether CD90
influences the proliferation rate of GC cells, a CCK-8 assay was
performed. The results demonstrated that the cell proliferation
rate was significantly increased in AGS-CD90 cells compared with
that in control cells (P<0.05). These results indicated that
CD90 promotes proliferation of GC cells (Fig. 1B).
CD90 inhibits senescence of GC
cells
Senescence represents stable cell cycle arrest and
serves a key role in tumor development and suppression. The
senescence-specific marker β-galactosidase catalyzes the production
of a blue-green product of X-Gal, and those cells stained with
β-galactosidase appear blue-green in color when observed under a
microscope according to the instructions of the β-galactosidase
staining kit (cat. no. C0602; Beyotime Institute of Biotechnology).
The role of CD90 in cell senescence was assessed using
β-galactosidase staining in the present study. The percentage of
senescent AGS-CD90 cells was 12.14%, while that in the control
group was 17.09%. The proportion of AGS-CD90 cells that were
positive for β-galactosidase staining was significantly smaller
than that of the control cells (P<0.05). This indicated that
CD90 inhibits the senescence of AGS cells (Fig. 1C and D).
CD90 promotes the invasion and
migration abilities of GC cells
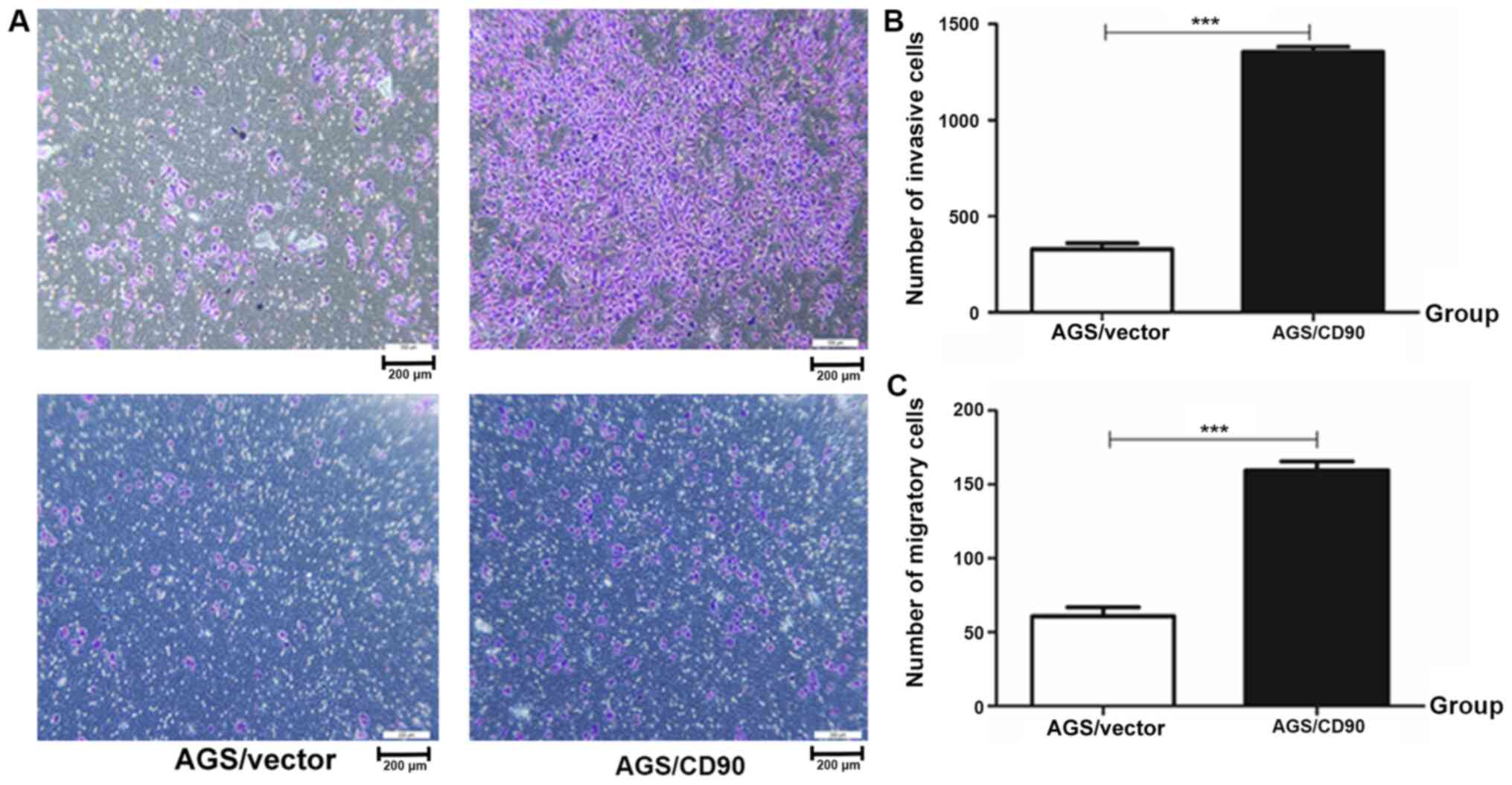
Invasion and migration ability experiments were
performed. The results demonstrated that the number of invasive
cells in the AGS-CD90 group was 1,397, while that in the control
group was 406. The number of migratory cells in the AGS-CD90 group
was 163, whereas that in the control group was 61. The number of
AGS-CD90 cells passing through matrix (cell invasion test) or
polycarbonate membrane (cell migration test) in the low nutritional
environment was higher than that of AGS-vector cells (P<0.01).
These results demonstrated that CD90 promoted the invasion and
migration abilities of GC cells (Fig.
2).
CD90 affects the LD concentration of
GC cells
LD is a key metabolite in aerobic glycolysis in
tumor cells. The level of LD is associated with tumor metastasis
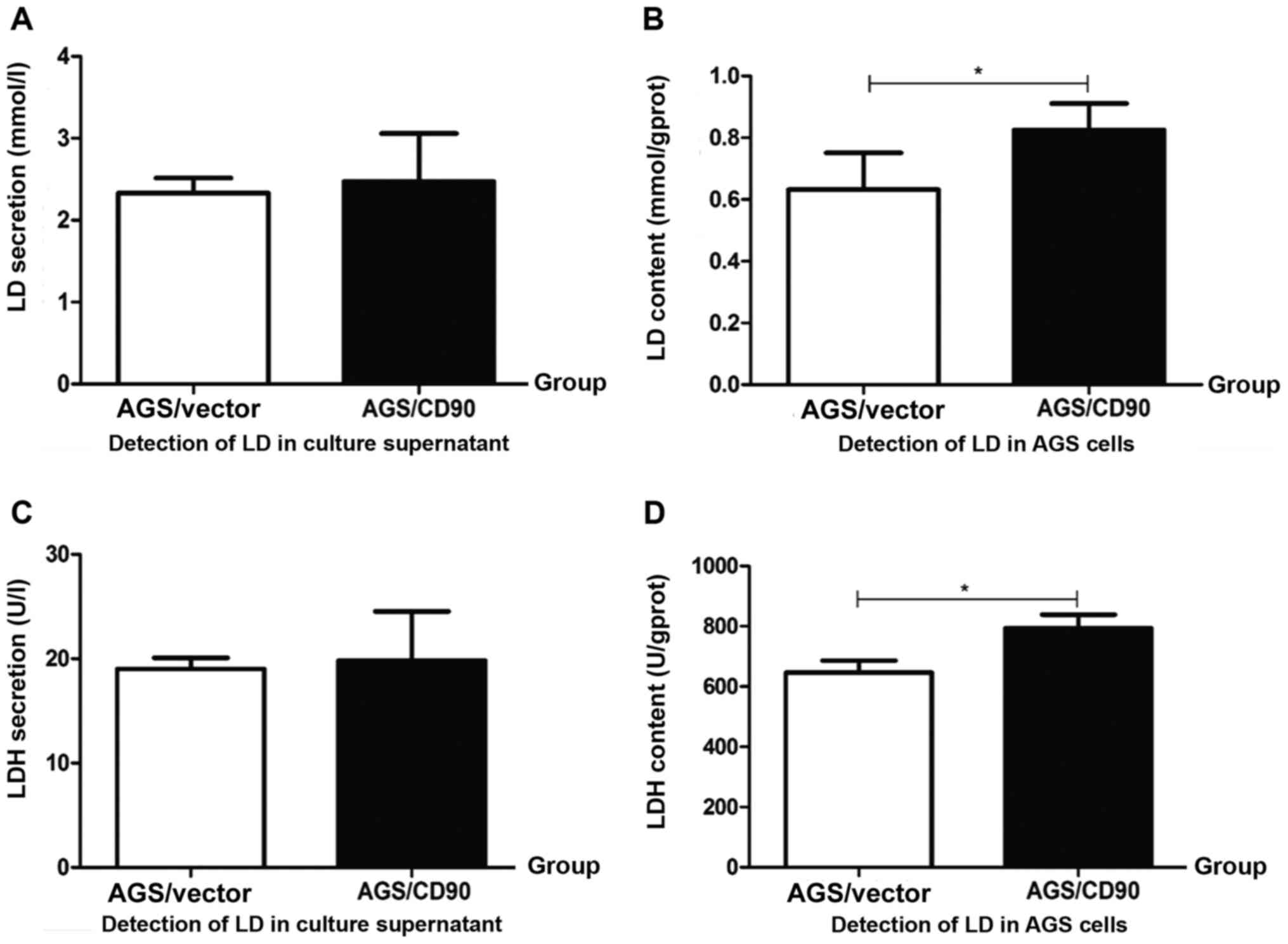
and poor prognosis (27,28). LD concentration was detected in
AGS-CD90, AGS-vector cells and culture supernatant in the present
study. There was no significant difference in the LD concentrations
between the AGS-CD90 and AGS-vector cell culture supernatant.
However, the intracellular LD concentration of AGS-CD90 cells was
higher than that of the AGS-vector cells. These results indicated
that CD90 overexpression resulted in the accumulation of LD in GC
cells (Fig. 3A and B).
CD90 upregulates LDH concentration in
GC cells
Pyruvate is degraded to LD under the action of LDH
and cannot enter the tricarboxylic acid cycle (29). Therefore, it was hypothesized that
the level of LDH in AGS-CD90 cells would be altered in the present
study. LDH concentration was detected in the cells and culture
supernatant of the AGS-CD90 and AGS-vector groups. The
intracellular LDH concentration was increased in AGS-CD90 cells
compared with that in AGS-vector cells. However, there was no
significant difference in LDH concentration in the cell culture
supernatant of the AGS-CD90 and AGS-vector groups (Fig. 3C and D).
CD90 increases the
NADPH/NADP+ ratio of GC cells
NADP+ is a coenzyme of dehydrogenases and
produces NADPH. Therefore, changes to the NADPH/NADP+
ratio are associated with the pentose phosphate pathway and
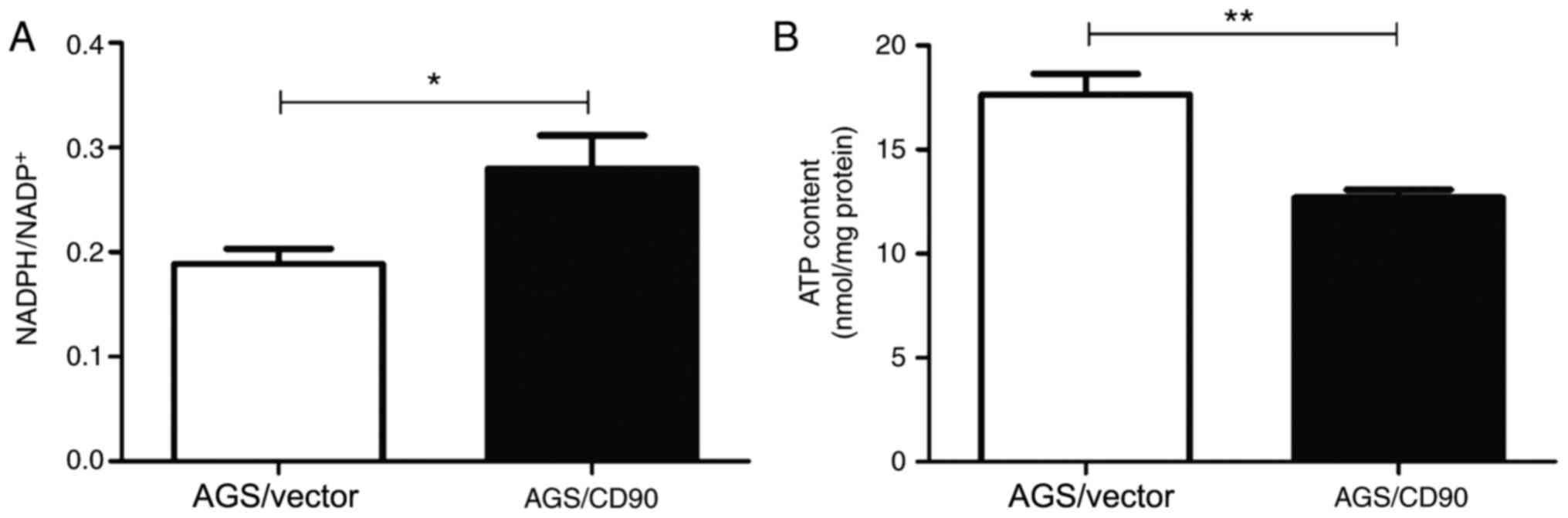
biosynthesis. The present study compared the intracellular
NADPH/NADP+ ratios of AGS-CD90 and AGS-vector cells. The
results demonstrated that the NADPH/NADP+ ratio in
AGS-CD90 cells was higher than that in control cells. The present
results also indicated that CD90 influenced levels of NADPH and
NADP+ in GC cells (Fig.
4A).
CD90 decreases the ATP concentration
of GC cells
NADPH/NADP+ is associated with
intracellular ATP content. In order to investigate whether CD90
affects the ATP levels of GC cells, ATP concentration was detected
in the cells and culture supernatant of the AGS-CD90 and AGS-vector
groups. The ATP concentration was lower in AGS-CD90 cells compared
with that in the AGS-vector cells. This indicated that CD90
decreases ATP concentration in GC cells (Fig. 4B).
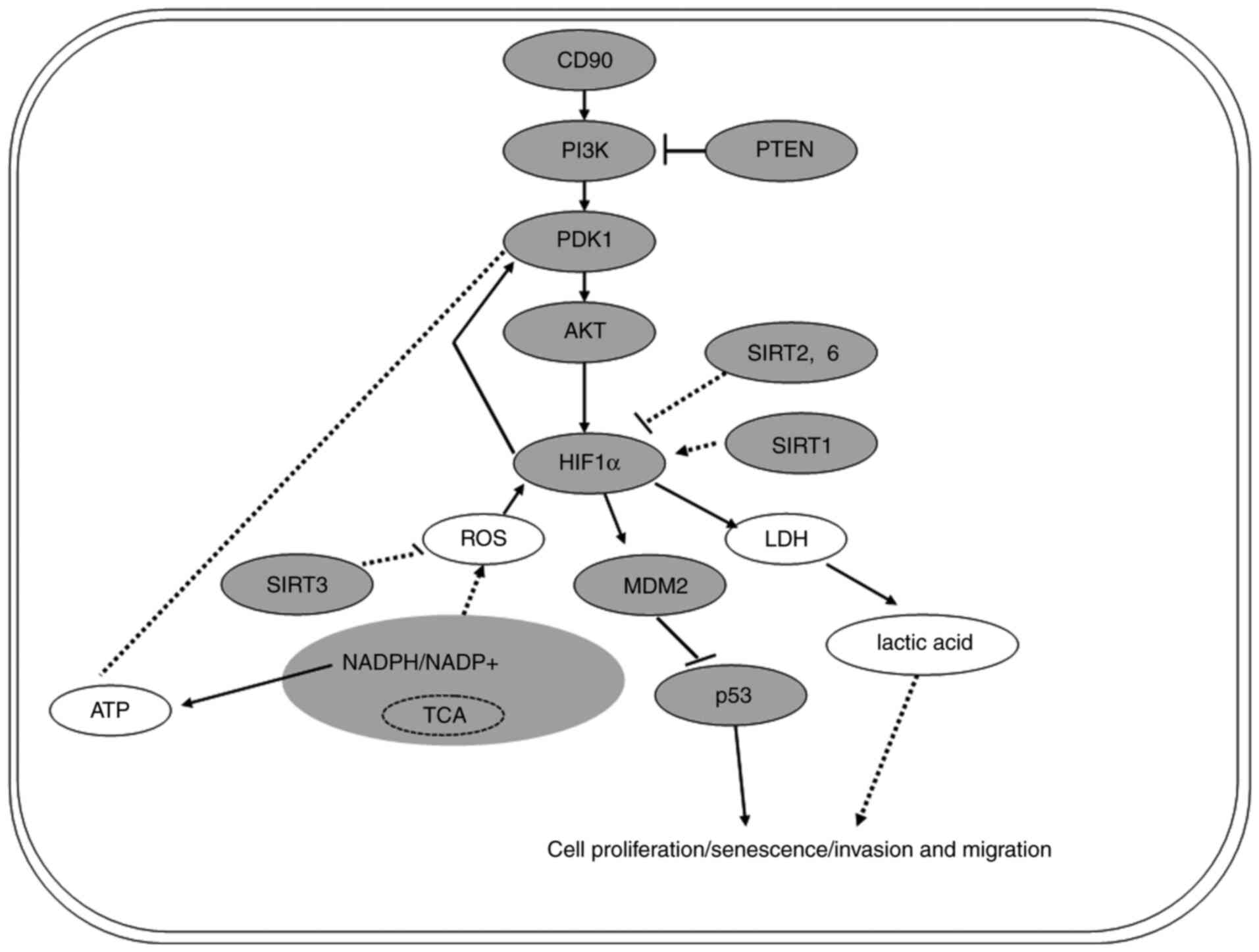
CD90 affects the biological behavior
and levels of energy metabolism of GC cells by regulating the
PI3K/AKT/HIF-1α signaling pathway
Western blotting was performed to investigate the
mechanism by which CD90 affects biological behavior and energy
metabolism in GC cells. The results demonstrated that levels of
PI3K, PDK1, p-AKT-Ser473, HIF-1α and MDM2 were upregulated in
AGS-CD90 cells compared with control cells. In contrast, levels of
PTEN and p53 were downregulated in AGS-CD90 cells (Fig. 5). The results of the present study
demonstrated that CD90 affects the activity of the PI3K/AKT/HIF-1α
signaling pathway. In addition, expression levels of SIRT1 were
upregulated in AGS-CD90 cells compared with control cells.
Meanwhile, SIRT2, SIRT3 and SIRT6 levels were downregulated in
AGS-CD90 cells (Fig. 5). These
results indicate that CD90 affects the levels of LD and ATP via
regulation of levels of SIRTs and HIF-1α.
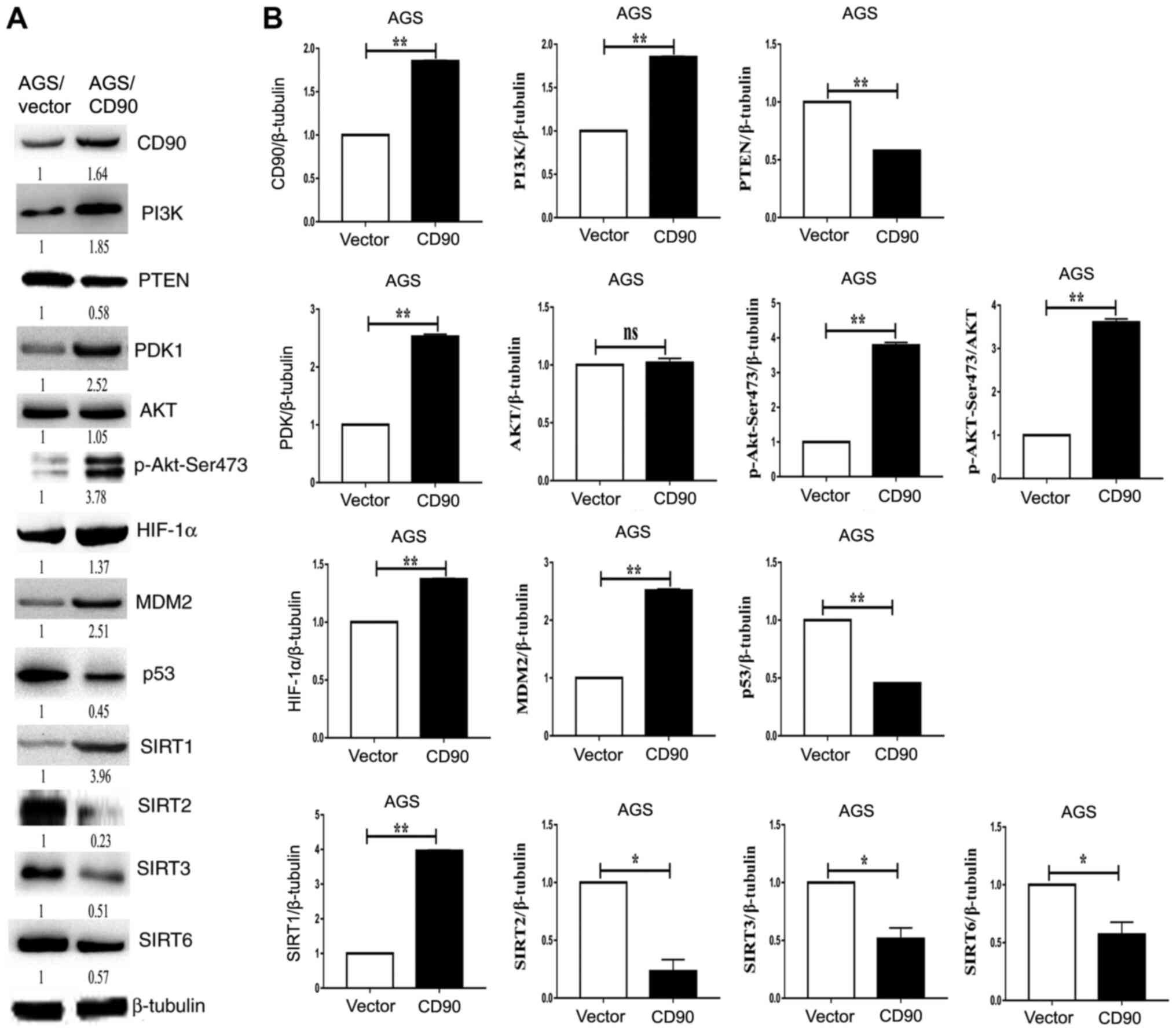 | Figure 5.Western blotting analysis of the
PI3K/AKT/HIF-1α signaling pathway. (A) Levels of PI3K, PTEN, PDK1,
AKT, p-AKT-Ser473, HIF-1α, MDM2, P53, SIRT2, SIRT3 and SIRT6 were
assessed via western blotting. Left, AGS-vector cells; right,
AGS-CD90 cells. (B) Densitometric analysis of the western blotting
results. *P<0.05, **P<0.01. HIF-1α, hypoxia-inducible factor
1α; SIRT, Sirtuin; Vector, gastric cancer cells transfected with
pEGFP-N1; CD90, gastric cancer cells transfected with
pEGFP-N1/CD90. |
Discussion
GC is the fourth most frequently occurring
malignancy worldwide (1,30), and is the second leading cause of
cancer-associated mortality (31).
East Asia accounts for ~60% of GC cases worldwide, and 41% of cases
occur in China alone (1). Although
progress in early diagnosis and adjuvant therapy has improved
outcomes for patients with GC, radical surgery remains to be the
primary treatment approach. GC can relapse with local or distant
metastasis following radical gastrectomy; therefore, outcomes
remain unsatisfactory. The pathogenesis of GC is a multi-factor and
multi-step process, involving microorganisms, proto-oncogenes,
tumor suppressor genes and immunological factors (32–35).
Helicobacter pylori is considered to be a notable cause of
GC but does not account for all cases (36). Thus, the molecular basis of GC
remains unclear.
Energy metabolism in cancer cells is an area of
interest in research. The Warburg effect indicates that cancer
cells have faults in mitochondrial oxidative phosphorylation
(37). Even in the presence of
plentiful oxygen, cancer cells rely primarily on anaerobic
glycolysis in the cytosol as a source of ATP to support cell
proliferation (38). Normal cells do
not tend to use the aerobic glycolysis pathway. Aerobic glycolysis
occurs most frequently in proliferative cells, such as tumor cells,
which use glucose via the glycolysis pathway, even in aerobic
conditions, which requires overexpression of glucose transporters
and glycolytic enzymes, resulting in the secretion of LD (39–41).
A previous study verified that abnormally high
expression levels of CD90 are present in GC tissues, and as such,
constructed an AGS cell line that stably overexpresses CD90,
demonstrating that CD90 promotes proliferation and inhibits
apoptosis (14). The present study
investigated the effect of CD90 on the biological behavior and
energy metabolism of GC cells. CD90 promoted cell proliferation and
the invasion and migration abilities of AGS cells, and inhibited
their senescence. Overexpression of CD90 promoted the accumulation
of LD and ATP, upregulated the LDH concentration and increased the
intracellular NADPH/NADP+ ratio in AGS cells. The
association between metabolic levels of lactate, ATP and malignant
tumors has been confirmed by previous studies (42). For example Choi et al
(43) demonstrated that elevated
glycolysis, combined with increased LD production and secretion, is
a key metabolic feature of neuroendocrine prostate cancer (NEPC).
Certain oncogenes or tumor suppressor genes serve key roles in the
regulation of metabolism and biological behavior of malignant tumor
cells. Monocarboxylate transporter 4 (MCT4) is a plasma membrane LD
transporter. The expression level of MCT4 can be inhibited using
antisense oligonucleotides; this decreases LD secretion and glucose
metabolism, thus affecting the proliferation of NEPC cells
(43). Similarly, the present study
demonstrated that CD90 serves a key role in the metabolism and
biological behavior of GC cells. It has also been demonstrated that
LD secreted by tumors is associated with a decrease of macrophages
in head and neck squamous cell carcinoma, and induces polarization
of M2-like macrophages (44). These
previous studies also demonstrate the key role of LD in the
pathogenesis of malignant tumors. LD, NADPH, ATP and other
metabolites serve an important role in the occurrence and
progression of malignant tumor cells. Circulating melanoma cells,
metastases and primary tumor cells exhibit elevated levels of
reactive oxygen species (ROS). Compared with these cells, a highly
metastatic type of cancer experiences reversible metabolic changes,
increased mitochondrial NADPH levels and ROS stress resistance;
inhibition of these effects decreases metastasis of tumor cells
(39,45). ATP is released into the extracellular
matrix via exocytosis or hemichannels (46). In order to amplify TGF-β1-induced
migration of lung cancer cells, autocrine signaling via exocytosis
of ATP and activation of P2 receptors is required (47). Alvarez et al (48) demonstrated that hemichannels activate
the downstream region of the αVβ3 integrin-PI3K-PLCγ-IP3R pathway
and are responsible for Thy-1-induced, hemichannel-mediated and
Syndecan-4-modulated ATP release, which transactivates P2X7Rs to
induce Ca(2+) entry into cells. The present study demonstrated that
CD90 affects the levels of LD, ATP and other metabolites of GC
cells, and influences the invasion and metastasis ability of GC
cells, thus clarifying the function and mechanism of CD90 in
GC.
In order to investigate the mechanism underlying the
effects of CD90 on the biological behavior and energy metabolism of
GC cells, key molecules involved in the PI3K/AKT/HIF-1α signaling
pathway were assessed. It was demonstrated that CD90 upregulated
the levels of PI3K, PDK1, AKT, p-AKT-Ser473, HIF-1α and MDM2, and
downregulated the levels of PTEN and p53 (Fig. 6). Hyper-activation of PI3K/AKT
signaling is common in numerous types of malignant tumor and
promotes cell proliferation (49).
Studies have demonstrated that metastasis suppressor protein 1 is
regulated by PTEN to suppress invasion, migration and
epithelial-to-mesenchymal transition of gastric carcinoma cells by
inactivating PI3K/AKT signaling (50,51).
Dupuy et al (52)
demonstrated that pyruvate dehydrogenase kinase 1 phosphorylates
pyruvate dehydrogenase, thus serving a key role in reprogramming
metabolism. HIF-1α is another key molecule in the PI3K/AKT/HIF-1α
signaling pathway. Studies have demonstrated that dimerization of
the HIF-1α and -β subunits forms the HIF-1 complex (53,54).
This complex alters the transcription of associated genes such that
tumor cells under hypoxic conditions generate energy to maintain
growth (55,56). In tumor cells, mutated p53 cannot
interact with BCL2 and therefore cannot promote mitochondrial
membrane permeation. Therefore, in addition to the effect on
transcriptional activity, the p53 mutant can also promote the
survival of cancer cells via direct action on mitochondrial
function (57). Furthermore, the
Sirtuin histone deacetylase family has been identified as a key
regulatory factor in metabolism of cancer cells (58,59). In
summary, the PI3K/AKT/HIF-1α signaling pathway serves a key role in
regulating the metabolism and biological behavior of malignant
tumor cells (Fig. 6).
The present study demonstrated that CD90 affects the
metabolism and biological behavior of GC cells via the
PI3K/AKT/HIF-1α signaling pathway. The interaction between CD90 and
this pathway, and the mechanism underlying its regulation, are not
yet fully understood. Further investigations are thus required in
order to elucidate this.
The present study demonstrated that CD90 disrupts
the biological behavior and metabolic processes of GC cells,
potentially via the PI3K/AKT/HIF-1α signaling pathway.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Sciences Foundation of China (grant no. 81672685) and the
Science and Technology Foundation Survey Project of Ministry of
Science and Technology of China (grant nos. 2018FY100900 and
2018FY10090004).
Availability of data and materials
Not applicable.
Authors' contributions
LG performed the cell proliferation, invasion and
migration experiments, as well as the lactic acid and lactate
dehydrogenase experiments. JL performed the cell senescence,
NADPH/NADP+ ratio, and ATP concentration experiments.
JH, LL and ZH analyzed the PI3K/AKT/HIF-1α signaling pathway. CY
and XJ also performed the NADPH/NADP+ ratio and ATP
concentration experiments. GL and YZ were responsible for the
design and writing of the manuscript. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
HIF-1
|
hypoxia inducible factor 1
|
|
PDK1
|
pyruvate dehydrogenase kinase 1
|
|
LD
|
lactic acid
|
|
LDH
|
lactate dehydrogenase
|
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cunningham D, Allum WH, Stenning SP,
Thompson JN, Van de Velde CJ, Nicolson M, Scarffe JH, Lofts FJ,
Falk SJ, Iveson TJ, et al: Perioperative chemotherapy versus
surgery alone for resectable gastroesophageal cancer. N Engl J.
355:11–20. 2006. View Article : Google Scholar
|
|
3
|
Ueda T, Volinia S, Okumura H, Shimizu M,
Taccioli C, Rossi S, Alder H, Liu CG, Oue N, Yasui W, et al:
Relation between microRNA expression and progression and prognosis
of gastric cancer: A microRNA expression analysis. Lancet Oncol.
11:136–146. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cancer Genome Atlas Research Network, .
Comprehensive molecular characterization of gastric adenocarcinoma.
Nature. 513:202–209. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jeng CJ, McCarroll SA, Martin TF, Floor E,
Adams J, Krantz D, Butz S, Edwards R and Schweitzer ES: Thy-1 is a
component common to multiple populations of synaptic vesicles. J
Cell Biol. 140:685–698. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kawamura K, Hiroshima K, Suzuki T, Chai K,
Yamaguchi N, Shingyoji M, Yusa T, Tada Y, Takiguchi Y, Tatsumi K,
et al: CD90 is a diagnostic marker to differentiate between
malignant pleural mesothelioma and lung carcinoma with
immunohistochemistry. Am J Clin Pathol. 140:544–549. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shaikh MV, Kala M and Nivsarkar M: CD90 a
potential cancer stem cell marker and a therapeutic target. Cancer
Biomark. 16:301–307. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kumar A, Bhanja A, Bhattacharyya J and
Jaganathan BG: Multiple roles of CD90 in cancer. Tumour Biol.
37:11611–11622. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhu J, Thakolwiboon S, Liu X, Zhang M and
Lubman DM: Overexpression of CD90 (Thy-1) in pancreatic
adenocarcinoma present in the tumor microenvironment. PLoS One.
9:e1155072014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lung HL, Cheung AK, Cheng Y, Kwong FM, Lo
PH, Law EW, Chua D, Zabarovsky ER, Wang N, Tsao SW, et al:
Functional characterization of THY1 as a tumor suppressor gene with
antiinvasive activity in nasopharyngeal carcinoma. Int J Cancer.
127:304–312. 2010.PubMed/NCBI
|
|
11
|
Tang KH, Dai YD, Tong M, Chan YP, Kwan PS,
Fu L, Qin YR, Tsao SW, Lung HL, Lung ML, et al: A CD90(+)
tumor-initiating cell population with an aggressive signature and
metastatic capacity in esophageal cancer. Cancer Res. 73:2322–2332.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pirozzi G, Tirino V, Camerlingo R, La
Rocca A, Martucci N, Scognamiglio G, Franco R, Cantile M, Normanno
N and Rocco G: Prognostic value of cancer stem cells,
epithelial-mesenchymal transition and circulating tumor cells in
lung cancer. Oncol Rep. 29:1763–1768. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lu JW, Chang JG, Yeh KT, Chen RM, Tsai JJ
and Hu RM: Overexpression of Thy1/CD90 in human hepatocellular
carcinoma is associated with HBV infection and poor prognosis. Acta
Histochem. 113:833–838. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhu GC, Gao L, He J, Long Y, Liao S, Wang
H, Li X, Yi W, Pei Z, Wu M, et al: CD90 is upregulated in gastric
cancer tissues and inhibits gastric cancer cell apoptosis by
modulating the expression level of SPARC protein. Oncol Rep.
34:2497–2506. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Martinez-Outschoorn UE, Peiris-Pagés M,
Pestell RG, Sotgia F and Lisanti MP: Cancer metabolism: A
therapeutic perspective. Nat Rev Clin Oncol. 14:1132017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Koppenol WH, Bounds PL and Dang CV: Otto
Warburg's contributions to current concepts of cancer metabolism.
Nat Rev Cancer. 11:325–337. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Le A, Cooper CR, Gouw AM, Dinavahi R,
Maitra A, Deck LM, Royer RE, Vander Jagt DL, Semenza GL and Dang
CV: Inhibition of lactate dehydrogenase A induces oxidative stress
and inhibits tumor progression. Proc Natl Acad Sci USA.
107:2037–2042. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cairns RA, Harris IS and Mak TW:
Regulation of cancer cell metabolism. Nat Rev Cancer. 11:85–95.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Doherty JR and Cleveland JL: Targeting
lactate metabolism for cancer therapeutics. J Clin Invest.
123:3685–3692. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yuan LW, Yamashita H and Seto Y: Glucose
metabolism in gastric cancer: The cutting-edge. World J
Gastroenterol. 22:2046–2059. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Song H, Wang L, Liu HL, Wu XB, Wang HS,
Liu ZH, Li Y, Diao DC, Chen HL and Peng JS: Tissue metabolomic
fingerprinting reveals metabolic disorders associated with human
gastric cancer morbidity. Oncol Rep. 26:431–438. 2011.PubMed/NCBI
|
|
22
|
Wu H, Xue R, Tang Z, Deng C, Liu T, Zeng
H, Sun Y and Shen X: Metabolomic investigation of gastric cancer
tissue using gas chromatography/mass spectrometry. Anal Bioanal
Chem. 396:1385–1395. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cai Z, Zhao JS, Li JJ, Peng DN, Wang XY,
Chen TL, Qiu YP, Chen PP, Li WJ, Xu LY, et al: A combined
proteomics and metabolomics profiling of gastric cardia cancer
reveals characteristic dysregulations in glucose metabolism. Mol
Cell Proteomics. 9:2617–2628. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sambrook J and Russell DW: Molecular
Cloning: A Laboratory Manual. 3rd edition. Cold Spring Harbor
Laboratory Press; New York, NY: 2001
|
|
25
|
He J, Zhu G, Gao L, Chen P, Long Y, Liao
S, Yi H, Yi W, Pei Z, Wu M, et al: Fra-1 is upregulated in gastric
cancer tissues and affects the PI3K/Akt and p53 signaling pathway
in gastric cancer. Int J Oncol. 47:1725–1734. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liao S, Xiao S, Zhu G, Zheng D, He J, Pei
Z, Li G and Zhou Y: CD38 is highly expressed and affects the
PI3K/Akt signaling pathway in cervical cancer. Oncol Rep.
32:2703–2709. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Baumann F, Leukel P, Doerfelt A, Beier CP,
Dettmer K, Oefner PJ, Kastenberger M, Kreutz M, Nickl-Jockschat T,
Bogdahn U, et al: Lactate promotes glioma migration by
TGF-beta2-dependent regulation of matrix metalloproteinase-2. Neuro
Oncol. 11:368–380. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yokota H, Guo J, Matoba M, Higashi K,
Tonami H and Nagao Y: Lactate, choline, and creatine levels
measured by vitro 1H-MRS as prognostic parameters in patients with
non-small-cell lung cancer. J Magn Reson Imaging. 25:992–999. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Warburg OH: The classic: The chemical
constitution of respiration ferment. Clin Orthop Relat Res.
468:2833–2839. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kamangar F, Dores GM and Anderson WF:
Patterns of cancer incidence, mortality, and prevalence across five
continents: Defining priorities to reduce cancer disparities in
different geographic regions of the world. J Clin Oncol.
24:2137–2150. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jin EH, Sung JK, Lee SI and Hong JH:
Mitochondrial NADH dehydrogenase subunit 3 (MTND3) polymorphisms
are associated with gastric cancer susceptibility. Int J Med Sci.
15:1329–1333. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Buglioni S, Melucci E, Sperati F, Pallocca
M, Terrenato I, De Nicola F, Goeman F, Casini B, Amoreo CA, Gallo
E, et al: The clinical significance of PD-L1 in advanced gastric
cancer is dependent on ARID1A mutations and ATM expression.
Oncoimmunology. 7:e14576022018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Malfertheiner P: Helicobacter pylori
treatment for gastric cancer prevention. N Engl J Med.
378:1154–1156. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pormohammad A, Ghotaslou R, Leylabadlo HE,
Nasiri MJ, Dabiri H and Hashemi A: Risk of gastric cancer in
association with Helicobacter pylori different virulence factors: A
systematic review and meta-analysis. Microb Pathog. 118:214–219.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Infection with Helicobacter pylori. IARC
Monogr Eval Carcinog Risks Hum. 61:177–240. 1994.PubMed/NCBI
|
|
37
|
Ma B, Cheng H, Mu C, Geng G, Zhao T, Luo
Q, Ma K, Chang R, Liu Q, Gao R, et al: The SIAH2-NRF1 axis
spatially regulates tumor microenvironment remodeling for tumor
progression. Nat Commun. 10:10342019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Warburg O: On the origin of cancer cells.
Science. 123:309–314. 1956. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Vander Heiden MG and Deberardinis RJ:
Understanding the intersections between metabolism and cancer
biology. Cell. 168:657–669. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Altman BJ, Stine ZE and Dang CV: From
Krebs to clinic: Glutamine metabolism to cancer therapy. Nat Rev
Cancer. 16:619–634. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Vander Heiden MG, Cantley LC and Thompson
CB: Understanding the warburg effect: The metabolic requirements of
cell proliferation. Science. 324:1029–1033. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hu LP, Zhang XX, Jiang SH, Tao LY, Li Q,
Zhu LL, Yang MW, Huo YM, Jiang YS, Tian GA, et al: Targeting
purinergic receptor P2Y2 prevents the growth of pancreatic ductal
adenocarcinoma by inhibiting cancer cell glycolysis. Clin Cancer
Res. 25:1318–1330. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Choi SYC, Ettinger SL, Lin D, Xue H, Ci X,
Nabavi N, Bell RH, Mo F, Gout PW, Fleshner NE, et al: Targeting
MCT4 to reduce lactic acid secretion and glycolysis for treatment
of neuroendocrine prostate cancer. Cancer Med. 7:3385–3392. 2018.
View Article : Google Scholar
|
|
44
|
Ohashi T, Aoki M, Tomita H, Akazawa T,
Sato K, Kuze B, Mizuta K, Hara A, Nagaoka H, Inoue N and Ito Y:
M2-like macrophage polarization in high lactic acid-producing head
and neck cancer. Cancer Sci. 108:1128–1134. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Piskounova E, Agathocleous M, Murphy MM,
Hu Z, Huddlestun SE, Zhao Z, Leitch AM, Johnson TM, DeBerardinis RJ
and Morrison SJ: Oxidative stress inhibits distant metastasis by
human melanoma cells. Nature. 527:186–191. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Fang WG and Tian XX: Identification of a
new pro-invasion factor in tumor microenvironment: Progress in
function and mechanism of extracellular ATP. Beijing Da Xue Xue Bao
Yi Xue Ban. 49:188–195. 2017.(In Chinese). PubMed/NCBI
|
|
47
|
Takai E, Tsukimoto M, Harada H, Sawada K,
Moriyama Y and Kojima S: Autocrine regulation of TGF-beta1-induced
cell migration by exocytosis of ATP and activation of P2 receptors
in human lung cancer cells. J Cell Sci. 125:5051–5060. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Alvarez A, Lagos-Cabre R, Kong M, Cárdenas
A, Burgos-Bravo F, Schneider P, Quest AF and Leyton L:
Integrin-mediated transactivation of P2X7R via
hemichannel-dependent ATP release stimulates astrocyte migration.
Biochim Biophys Acta. 1863:2175–2188. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yao H, Su S, Xia D, Wang M, Li Z, Chen W,
Ren L and Xu L: F-box and leucine-rich repeat protein 5 promotes
colon cancer progression by modulating PTEN/PI3K/AKT signaling
pathway. Biomed Pharmacother. 107:1712–1719. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang H, Zhao Y, Cao L, Zhang J, Wang Y and
Xu M: Metastasis suppressor protein 1 regulated by PTEN suppresses
invasion, migration, and EMT of gastric carcinoma by inactivating
PI3K/AKT signaling. J Cell Biochem. 120:3447–3454. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Matsuoka T and Yashiro M: The role of
PI3K/Akt/mTOR signaling in gastric carcinoma. Cancers (Basel).
6:1441–1463. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Dupuy F, Tabariès S, Andrzejewski S, Dong
Z, Blagih J, Annis MG, Omeroglu A, Gao D, Leung S, Amir E, et al:
PDK1-dependent metabolic reprogramming dictates metastatic
potential in breast cancer. Cell Metab. 22:577–589. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Semenza GL and Wang GL: A nuclear factor
induced by hypoxia via de novo protein synthesis binds to the human
erythropoietin gene enhancer at a site required for transcriptional
activation. Mol Cell Biol. 12:5447–5454. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wang GL and Semenza GL: Purification and
characterization of hypoxia-inducible factor 1. J Biol Chem.
270:1230–1237. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Sáenz-de-Santa-María I,
Bernardo-Castiñeira C, Secades P, Bernaldo-de-Quirós S, Rodrigo JP,
Astudillo A and Chiara MD: Clinically relevant HIF-1α-dependent
metabolic reprogramming in oropharyngeal squamous cell carcinomas
includes coordinated activation of CAIX and the miR-210/ISCU
signaling axis, but not MCT1 and MCT4 upregulation. Oncotarget.
8:13730–13746. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Devignes CS, Aslan Y, Brenot A, Devillers
A, Schepers K, Fabre S, Chou J, Casbon AJ, Werb Z and Provot S: HIF
signaling in osteoblast-lineage cells promotes systemic breast
cancer growth and metastasis in mice. Proc Natl Acad Sci USA.
115:E992–E1001. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Vyas S, Zaganjor E and Haigis MC:
Mitochondria and Cancer. Cell. 166:555–566. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Bringman-Rodenbarger LR, Guo AH, Lyssiotis
CA and Lombard DB: Emerging roles for SIRT5 in metabolism and
cancer. Antioxid Redox Signal. 28:677–690. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
De Matteis S, Granato AM, Napolitano R,
Molinari C, Valgiusti M, Santini D, Foschi FG, Ercolani G,
Vespasiani Gentilucci U, Faloppi L, et al: Interplay between
SIRT-3, metabolism and its tumor suppressor role in hepatocellular
carcinoma. Dig Dis Sci. 62:1872–1880. 2017. View Article : Google Scholar : PubMed/NCBI
|