Introduction
Gastric cancer (GC) is the third leading cause of
cancer-associated mortality and the fifth most frequent type of
cancer worldwide. According to the latest statistics, there were
more than 1,000,000 new cases of GC and an estimated 783,000 deaths
from GC worldwide in 2018 (1–3). Risk
factors, including smoking and atrophic gastritis, have been
identified to contribute to the incidence of GC (4). Despite improvements in the diagnosis
and treatment of GC, the 5-year survival rate for patients with GC
remains unsatisfactory (5).
Chemotherapeutic cisplatin (CDDP) is one of the main options for
the treatment of patients with various types of cancer (6). However, CDDP treatment often results in
the development of chemoresistance, leading to therapeutic failure
(7). Therefore, it is important to
understand the molecular regulatory mechanism underlying
chemoresistance and to identify effective tumor biomarkers to
improve the prognosis of patients with GC.
Long non-coding RNAs (lncRNAs) are a type of
transcript, with a length of >200 nucleotides, that lack
protein-coding ability (8,9). Previous studies have reported that
lncRNAs are involved in the regulation of human cancer by acting as
oncogenes or tumor suppressor genes. For example, Wu et al
(10) revealed that lncRNA MEG3
inhibited the progression of prostate cancer via binding to
microRNA (miRNA/miR)-9-5p and regulating quaking I-5 protein
expression. Moreover, Gao et al (11) indicated that lncRNA NORAD facilitated
the progression of non-small cell lung cancer by enhancing cell
proliferation and glycolysis. Additionally, Xu et al
(12) demonstrated that lncRNA
ROR1-antisense RNA 1 (AS1) sponged miR-375 to aggravate lung
adenocarcinoma metastasis and to enhance epithelial-mesenchymal
transition, while Zhang et al (13) reported that the lncRNA
FOXD2-AS1/miR-25-3p/Sema4c axis increased colorectal cancer cell
migration and invasion.
lncRNAs are reported to be involved in the
regulation of CDDP resistance in GC. For instance, lncRNA MALAT1
enhances autophagy-associated CDDP resistance in GC by sponging
miR-20b and by upregulating autophagy protein 5 (14). Furthermore, prostate
cancer-associated ncRNA transcripts 1 promotes CDDP resistance in
GC by regulating the miR-128/zinc-finger E-box binding homeobox 1
axis (15), while OIP5-AS1
suppression increases CDDP sensitivity by targeting the
miR-377-3p/fos-like antigen 2 axis in osteosarcoma (16). MCM3AP-AS1 has been reported to
promote tumorigenesis and progression in various types of cancer,
such as pancreatic cancer, hepatocellular carcinoma and breast
cancer (17–19). However, the regulatory effects of
lncRNA MCM3AP-AS1 on CDDP resistance in GC remain to be
determined.
The present study aimed to investigate the role of
MCM3AP-AS1 and the regulatory mechanisms associated with this
lncRNA in CDDP-resistant GC cells, which may provide a novel
biomarker for the diagnosis and treatment of GC.
Materials and methods
Cell lines and cell culture
GC cell lines (AGS, MKN45, NCI-N87 and SNU638), the
normal gastric mucosa epithelial GES-1 cell line and 293T cells
were obtained from The Cell Bank of Type Culture Collection of the
Chinese Academy of Sciences. Cells were incubated in RPMI-1640
medium supplemented with 10% FBS (both purchased from Thermo Fisher
Scientific, Inc.) and 1% penicillin-streptomycin (Thermo Fisher
Scientific, Inc.). CDDP-resistant GC cell lines (NCI-N87/CDDP and
AGS/CDDP) were induced in RPMI-1640 medium with increasing
concentrations (0–10 µg/ml) of CDDP at 37°C for more than 6 months.
Cell incubation conditions included a humidified environment with
5% CO2 at 37°C.
Cell transfection
Specific short hairpin RNAs (shRNAs) against
MCM3AP-AS1 (shMCM3AP-AS1#1; 5′-GGGAGUAAGUGAAAGUAAU-3′ and
shMCM3AP-AS1#2; 5′-GCUUCGAUGUGUUACUUAA-3′) were used to knock down
MCM3AP-AS1 expression. A sh-negative control (shNC; 10 nM;
5′-UUCUCCGAACGUGUCACGU-3′) was used as the NC. shRNAs were used for
stable transfection. The pcDNA3.1 plasmids targeting FOXC1 or
MCM3AP-AS1 were used to overexpress FOXC1 or MCM3AP-AS1,
respectively, with empty pcDNA3.1 as the NC. miR-138 mimics
(5′-AGCUGGUGUUGUGAAUCAGGCCG-3′) and miR-138 inhibitor
(5′-CGGCCUGAUUCACAACACCAGCU-3′) were utilized to overexpress and
knock down miR-138 expression, respectively. NC mimics (NC
miR-mimics; 10 nM; 5′-ACUCUAUCUGCACGCUGACUU-3′) and NC inhibitor
(NC miR-inhibitor; 10 nM; 5′-CAGUACUUUUGUGUAGUACAA-3′) served as
scrambled NCs. The aforementioned vectors were provided by Shanghai
GenePharma Co., Ltd.. Transfection was conducted using
Lipofectamine® 2000 for 48 h at 37°C (Invitrogen; Thermo
Fisher Scientific, Inc.). Cells were subjected to subsequent
experimentation 48 h following transfection.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from NCI-N87 and AGS cells
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). A PrimeScript RT reagent kit (cat. no. RR047A;
Takara Biotechnology Co., Ltd.) or a TaqMan™ Advanced miRNA cDNA
Synthesis kit (cat. no. 4366596; Thermo Fisher Scientific, Inc.)
were used to reverse transcribe RNAs into cDNA according to the
manufacturer's protocol. RT-qPCR was performed on the ABI 7500
real-time PCR System (Applied Biosystems; Thermo Fisher Scientific,
Inc.) using a SYBR-Green PCR Master Mix kit (Takara Biotechnology
Co., Ltd.). The following thermocycling conditions were used:
Pre-denaturation at 95°C for 15 sec, denaturation at 94°C for 30
sec, annealing at 60°C for 20 sec, and extension at 72°C for 40 sec
for 40 cycles. The expression levels of genes were calculated
utilizing the 2−ΔΔCq method (20), with GAPDH or U6 as the internal
controls The primer (Shanghai GeneChem) sequences used in the
present study were as follows: MCM3AP-AS1 forward,
5′-CCTATCCCTTTCTCTAAGA-3′, reverse, 5′-ACTTCTGCAAAAACGTGCTG-3′;
miR-138 forward, 5′-GTTAGGGCAGGTGGGATG-3′, reverse,
5′-TGTATGCGGCTGGTAAGTAG-3′; FOXC1 forward,
5′-AACATCGCCTGCGTTATCCTC-3′, reverse, 5′-ACGTCCCGGATGATCCCAA-3′; U6
forward, 5′-TTATGGGTCCTAGCCTGAC-3′, reverse,
5′-CACTATTGCGGGTCTGC-3′; and GAPDH forward,
5′-CCACATCGCTCAGACACCAT-3′ and reverse,
5′-ACCAGGCGCCCAATACG-3′.
Cell viability and drug-sensitivity
assay
The viability of NCI-N87/CDDP and AGS/CDDP cells was
assessed using a Cell Counting Kit-8 (CCK-8; Dojindo Molecular
Technologies, Inc.) assay. Cells (1×104 cells/well) were
seeded into 96-well plates. Following incubation at 37°C for 0, 24,
48 and 72 h, each well was supplemented with 10 µl CCK-8 reagent
and the cells were incubated for 4 h at room temperature to examine
cell viability. Finally, cell viability was represented by
absorbance values at 450 nm, which were measured using the MRX II
microplate reader (Dynex Technologies).
To determine the IC50, NCI-N87/CDDP and
AGS/CDDP cells were treated with varying concentrations (0, 1, 2,
4, 8, 16 and 32 µg/ml) of CDDP for 48 h.
RNA pull-down
An RNA pull-down assay was conducted using the
Pierce Magnetic RNA-Protein Pull-Down kit (cat. no. 20164; Pierce;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. GC cells were transfected with 50 µM
biotinylated-MCM3AP-AS1 (Biotin-MCM3AP-AS1) and biotinylated NC.
Cells were collected 48 h after transfection and subsequently lysed
in RNase-free cell lysis solution (1 mM HEPES, 200 mM NaCl, 1%
Triton X-100, 10 mM MgCl2, 200 U/ml RNase Inhibitor) at
4°C. Cell lysates were incubated with M-280 streptavidin magnetic
beads (Invitrogen; Thermo Fisher Scientific, Inc.) overnight at
4°C, according to the manufacturer's protocol. Next, the beads were
washed with high salt buffer (1% Triton X-100; 0.1% SDS; 20 mM
Tris-HCl, pH 8.0; 2 mM EDTA; 500 mM NaCl). After washing and
centrifugation (1,500 × g; 10 min, 4°C), the pellet was lysed with
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.), the binding of miR-138 to MCM3AP-AS1 was assessed via
RT-qPCR, as aforementioned.
Wound healing assay
NCI-N87 and AGS cells (5×104 cells) were
plated in 6-well plates and incubated with RPMI-1640 medium without
serum for 24 h at 37°C. When cells reached 100% confluence, they
were used forthe wound healing assay. A 200-µl pipette tip was used
to generate scratches. The wound width at 0 and 24 h was observed
under an light microscope (magnification, ×200; Olympus
Corporation) and measured using ImageJ software version 1.8
(National Institutes of Health).
Transwell assay
NCI-N87 and AGS cells (1×104 cells)
suspended in serum-free medium were added into the upper chambers,
which were precoated with Matrigel® at 37°C for 1 h. The
lower chamber was filled with RPMI-1640 medium (600 µl) containing
10% FBS. After 48 h of incubation at 37°C, invasive cells were
fixed with methanol for 20 min at room temperature and stained with
0.1% crystal violet for 20 min at room temperature. The number of
invasive cells was counted under a light microscope (magnification,
×200; Olympus Corporation).
Luciferase reporter assay
The starBase 2.0 database (http://starbase.sysu.edu.cn/) was used to predict the
potential target genes of MCM3AP-AS1. The pmirGLO vectors (Shanghai
GenePharma Co., Ltd.) of MCM3AP-AS1-wild-type (Wt) or
MCM3AP-AS1-mutant (Mut) reporters were co-transfected with NC
mimics, miR-138 mimics, NC inhibitor or miR-138 inhibitor into 293T
cells using Lipofectamine 2000. In addition, pmirGLO-FOXC1-Wt or
pmirGLO-FOXC1-Mut reporters were co-transfected with NC mimics,
miR-138 mimics, NC inhibitor or miR-138 inhibitor into 293T cells
using Lipofectamine 2000. After 48 h, the relative luciferase
activity was measured using a Dual Luciferase Reporter assay system
(Promega Corporation) and normalized to Renilla luciferase
activity.
Western blot analysis
The proteins from transfected AGS/CDDP cells were
isolated using RIPA lysis buffer (Beyotime Institute of
Biotechnology) and then quantified using a BCA Protein Assay kit
(Pierce; Thermo Fisher Scientific, Inc.). After separating via 10%
SDS-PAGE (Bio-Rad Laboratories, Inc.), a total of 30 µg proteins
were transferred onto PVDF membranes (Invitrogen; Thermo Fisher
Scientific, Inc.). After blocking with 5% skimmed milk for 2 h at
room temperature, membranes were incubated with primary antibodies
against FOXC1 (1:1,000; cat. no. ab227977; Abcam) and GAPDH
(1:1,000; cat. no. ab9485; Abcam) overnight at 4°C. Subsequently,
HRP-conjugated goat anti-rabbit IgG secondary antibodies (1:1,000;
cat. no. ab205718; Abcam) were incubated with the membranes for 2 h
at room temperature. The blots were visualized using an ECL
detection system (Thermo Fisher Scientific, Inc.).
Nuclear/cytoplasmic RNA
fractionation
Nuclear and cytoplasmic fractions were acquired from
NCI-N87/CDDP and AGS/CDDP cells using nucleoplasmic fractionation
buffer (140 mmol/l NaCl, 1.5 mmol/l MgCl2, 10 mmol/l
Tris-HCl pH 8.5, 0.5% NP-40). First, cell pellet was resuspended
through nucleoplasmic fractionation buffer and incubated for 5 min
on ice. After centrifugation, the supernatant and pellet were
collected as the cytoplasmic and nuclear fractions, respectively.
RNA was isolated from nuclear/cytoplasmic fractions, and RT-qPCR
was then used to assess the levels of MCM3AP-AS1, GAPDH and U6.
GAPDH served as the cytoplasmic endogenous control, and U6 served
as the nuclear endogenous control.
Statistical analysis
Data are presented as the mean ± SD, and were
analyzed using SPSS 20.0 Software (IBM Corp.). Each experiment was
repeated ≥3 times. A one-way ANOVA followed by Tukey's post hoc
test and unpaired Student's t-test were used to compare the
differences among >2 and 2 groups, respectively. P<0.05 was
considered to indicate a statistically significant difference.
Results
MCM3AP-AS1-knockdown weakens CDDP
resistance in GC cells
To examine the biological role of MCM3AP-AS1 in the
CDDP resistance of GC cells, CDDP-resistant NCI-N87 cells
(NCI-N87/CDDP) and AGS cells (AGS/CDDP) were established. As
presented in Fig. 1A, the
IC50 of CDDP was significantly increased in NCI-N87/CDDP
and AGS/CDDP cells compared with in their parental cell lines.
Moreover, MCM3AP-AS1 expression was upregulated in CDDP-resistant
GC cells compared with in their parental cell lines (Fig. 1B). Subsequently, NCI-N87/CDDP and
AGS/CDDP cells were transfected with shMCM3AP-AS1#1/2. The
transfection efficiency was confirmed via RT-qPCR (Fig. 1C). Wound healing, Transwell and CCK-8
assay results indicated that the knockdown of MCM3AP-AS1
significantly inhibited the migration, invasion and proliferation
of NCI-N87/CDDP and AGS/CDDP cells (Fig.
1D-F). In addition, it was found that the IC50 in
NCI-N87/CDDP and AGS/CDDP cells was significantly decreased
following MCM3AP-AS1-knockdown (Fig.
1G). These findings indicated that MCM3AP-AS1-knockdown
potentiated the CDDP sensitivity of GC cells.
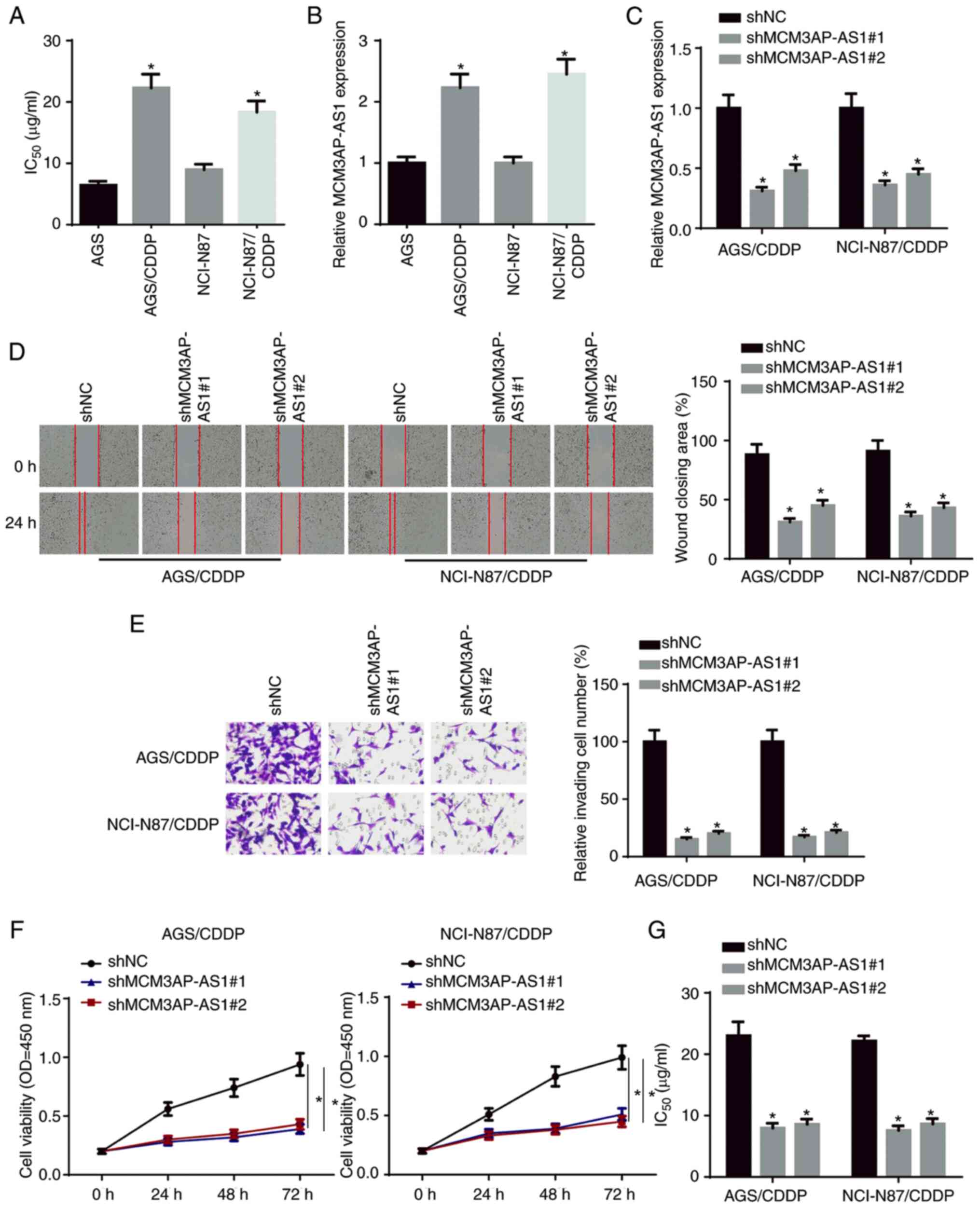 | Figure 1.MCM3AP-AS1-knockdown weakens CDDP
resistance in GC cells. (A) The IC50 was tested in
NCI-N87, NCI-N87/CDDP, AGS and AGS/CDDP. (B) MCM3AP-AS1 expression
in GC cells and CDDP-resistant GC cells was measured by RT-qPCR.
(C) The knockdown efficiency of MCM3AP-AS1 in NCI-N87/CDDP and
AGS/CDDP cells was assessed by RT-qPCR. (D) Migratory
(magnification, ×200) and (E) invasive abilities (magnification,
×200) of CDDP-resistant GC cells were detected after repressing
MCM3AP-AS1 by wound healing and Transwell assays, respectively. (F)
Cell viability of NCI-N87/CDDP and AGS/CDDP cells was measured
after repressing MCM3AP-AS1 by Cell Counting Kit-8 assay. (G) The
IC50 of NCI-N87/CDDP and AGS/CDDP was tested after
repressing MCM3AP-AS1 by drug-sensitivity assay. *P<0.05 vs.
AGS, NCI-N87 or shNC. MCM3AP-AS1, MCM3AP antisense RNA 1; CDDP,
cisplatin; GC, gastric cancer; RT-qPCR, reverse
transcription-quantitative PCR; sh, short hairpin RNA; NC, negative
control; OD, optical density. |
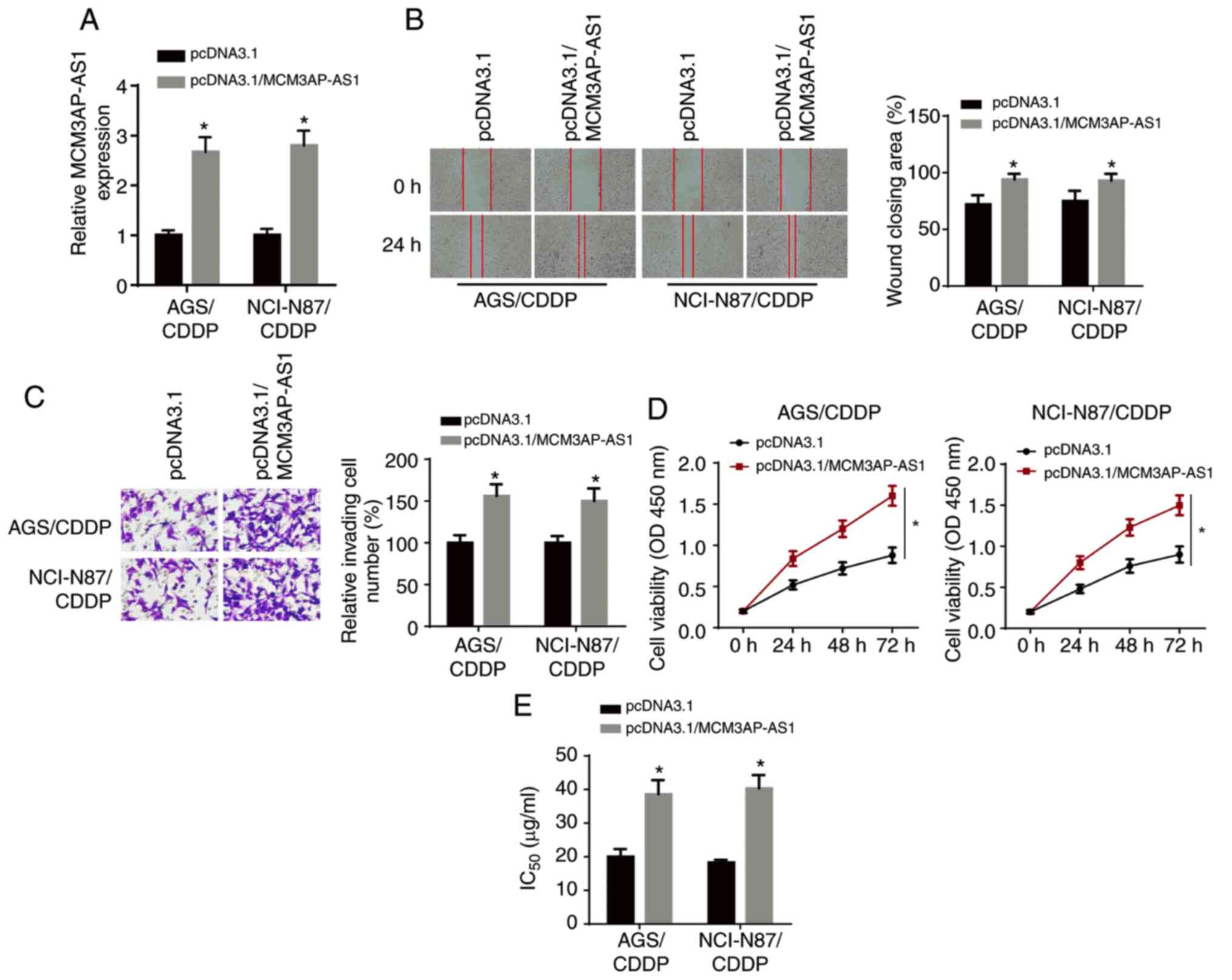
MCM3AP-AS1 overexpression enhances
CDDP resistance in GC cells
The overexpression efficiency of MCM3AP-AS1 was
verified via RT-qPCR analysis (Fig.
2A). Wound healing and Transwell assay results demonstrated
that the overexpression of MCM3AP-AS1 significantly enhanced the
migratory and invasive abilities of NCI-N87/CDDP and AGS/CDDP cells
(Fig. 2B and C). Moreover, CCK-8
assay results identified that the proliferative ability of
NCI-N87/CDDP and AGS/CDDP cells was increased after MCM3AP-AS1
overexpression (Fig. 2D). It was
found that the IC50 was significantly enhanced in
NCI-N87/CDDP and AGS/CDDP cells after MCM3AP-AS1 overexpression
(Fig. 2E). Thus, MCM3AP-AS1
overexpression decreased the CDDP sensitivity of GC cells.
MCM3AP-AS1 facilitates CDDP resistance
in GC cells via targeting miR-138
Accumulating evidence has revealed that lncRNAs
function as competitive endogenous (ce)RNAs in the regulation of
multiple types of human cancer by sponging miRNAs (21,22).
Therefore, it was hypothesized that MCM3AP-AS1 may be involved in
the regulation of CDDP resistance in GC via the ceRNA network.
First, the LncLocator program (http://www.csbio.sjtu.edu.cn/bioinf/LncLocator/)
predicted that MCM3AP-AS1 was mainly expressed in the cytoplasm
(Fig. 3A). Subsequently,
nuclear-cytoplasmic fractionation assay results suggested that the
majority of MCM3AP-AS1 was distributed in the cytoplasm of GC cells
(Fig. 3B). Using the starBase v2.0
database, the potential binding sites between MCM3AP-AS1 and
miR-138 were predicted (Fig. 3C). A
luciferase reporter assay demonstrated that the luciferase activity
of pmirGLO-MCM3AP-AS1-Wt reporters was significantly decreased in
the miR-138 mimics group, but was enhanced in the miR-138 inhibitor
group; moreover, no significant differences were identified in
pmirGLO-MCM3AP-AS1-Mut reporters (Fig.
3D).
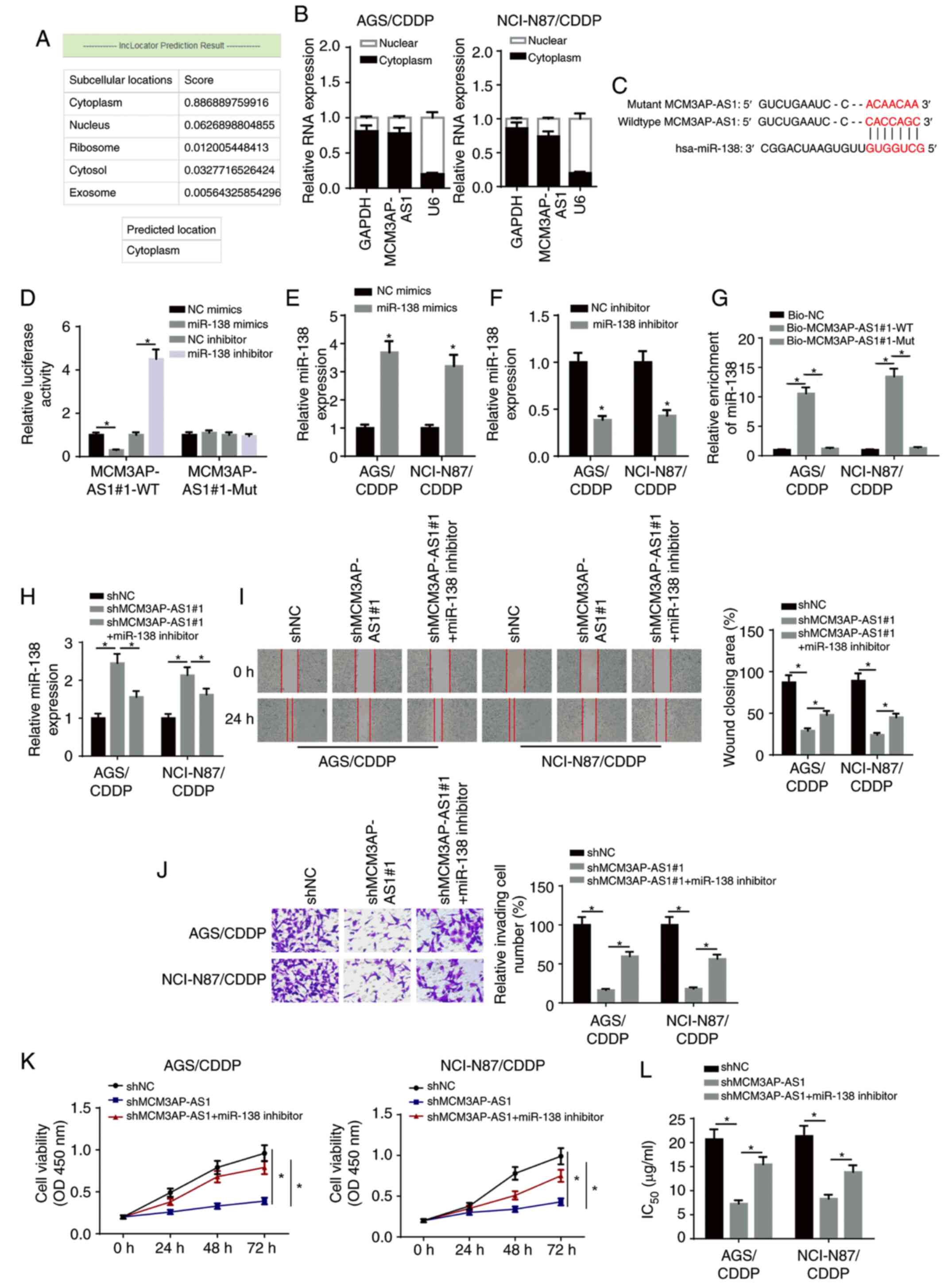 | Figure 3.MCM3AP-AS1 facilitates CDDP
resistance of GC cells by targeting miR-138. (A) LncLocator program
predicted the subcellular localization of MCM3AP-AS1. (B)
MCM3AP-AS1 expression in the cytoplasmic and nuclear fractions was
measured in NCI-N87/CDDP and AGS/CDDP cells by subcellular fraction
assay. (C) The binding site between MCM3AP-AS1 and miR-138 was
predicted using the starBase website. (D) The binding ability of
MCM3AP-AS1 for miR-138 was detected in 293T cells by luciferase
reporter assay. miR-138 expression in NCI-N87/CDDP and AGS/CDDP
cells transfected with (E) NC mimics and miR-138 mimics, and (F) NC
inhibitor and miR-138 inhibitor. (G) The binding capacity between
MCM3AP-AS1 and miR-138 was verified by RNA pull-down assay. (H)
miR-138 expression was assessed in shNC, shMCM3AP-AS1#1 and
shMCM3AP-AS1#1+miR-138 inhibitor groups by reverse
transcription-quantitative PCR. (I) Migratory (magnification, ×200)
and (J) invasive abilities (magnification, ×200) of NCI-N87/CDDP
and AGS/CDDP cells were detected in shNC, shMCM3AP-AS1#1 and
shMCM3AP-AS1#1+miR-138 inhibitor groups by wound healing and
Transwell assay, respectively. (K) Cell viability of NCI-N87/CDDP
and AGS/CDDP cells was measured in shNC, shMCM3AP-AS1#1 and
shMCM3AP-AS1#1+miR-138 inhibitor groups by Cell Counting Kit-8
assay. (L) The IC50 values of CDDP-resistant GC cells
was tested in shNC, shMCM3AP-AS1#1 and shMCM3AP-AS1#1+miR-138
inhibitor groups by drug-sensitivity assay. *P<0.05. miR,
microRNA; MCM3AP-AS1, MCM3AP antisense RNA 1; CDDP, cisplatin; GC,
gastric cancer; sh, short hairpin RNA; NC, negative control; OD,
optical density; WT, wild-type; Mut, mutant; Bio, biotinylated. |
RT-qPCR revealed that miR-138 expression was
upregulated in NCI-N87/CDDP and AGS/CDDP cells transfected with
miR-138 mimics, and downregulated in NCI-N87/CDDP and AGS/CDDP
cells transfected with miR-138 inhibitor (Fig. 3E and F). RNA-pull down assay results
indicated that biotinylated-MCM3AP-AS1 (Biotin-MCM3AP-AS1-WT) was
able to directly precipitate miR-138, while biotinylated-MCM3AP-AS1
with predicted mutant binding sites (Biotin-MCM3AP-AS1-Mut) could
not precipitate miR-138 (Fig. 3G).
In addition, miR-138 expression was significantly increased by
knocking down MCM3AP-AS1, but this effect was abrogated by the
miR-138 inhibitor (Fig. 3H).
Additionally, MCM3AP-AS1-knockdown inhibited the migration,
invasion and proliferation of CDDP-resistant GC cells, which was
then reversed by miR-138 silencing (Fig.
3I-K). Additionally, the miR-138 inhibitor partially abolished
the inhibitory effect of MCM3AP-AS1-knockdown on the
IC50 in NCI-N87/CDDP and AGS/CDDP cells (Fig. 3L). Therefore, it was suggested that
MCM3AP-AS1 increased the CDDP resistance of CDDP-resistant GC cells
via sponging miR-138.
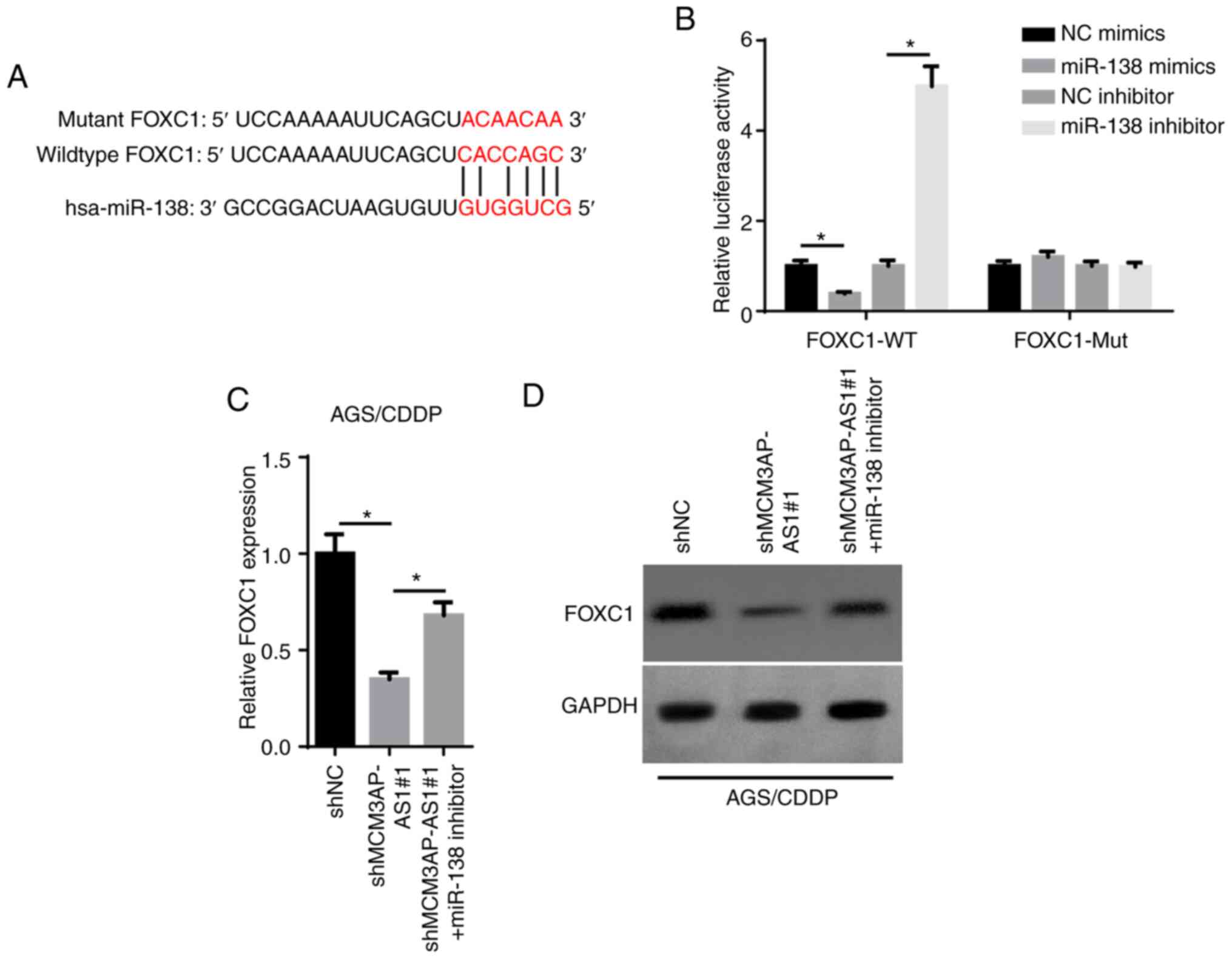
MCM3AP-AS1 serves as a ceRNA for
miR-138 to upregulate FOXC1 expression
Upregulated FOXC1 has been reported to be associated
with a poor prognosis in patients with GC (23). The starBase database was used to
predict the binding sites between miR-138 and FOXC1 (Fig. 4A). As presented in Fig. 4B, the luciferase activity of the
FOXC1-WT reporters was significantly decreased by overexpressing
miR-138, while it was enhanced by inhibiting miR-138. Additionally,
no significant change was observed in the FOXC1-Mut group (Fig. 4B). Furthermore, FOXC1 mRNA and
protein expression was decreased by MCM3AP-AS1-knockdown, but this
effect was counteracted by the miR-138 inhibitor in AGS/CDDP cells
(Fig. 4C and D). Overall, these
findings demonstrated that MCM3AP-AS1 positively regulated FOXC1
expression by directly sponging miR-138.
MCM3AP-AS1/miR-138 axis increases the
resistance of GC cells to CDDP by regulating FOXC1
RT-qPCR revealed that transfection of the FOXC1
overexpression plasmid significantly increased FOXC1 expression in
AGS/CDDP cells (Fig. 5A). As
illustrated in Fig. 5B and C, FOXC1
expression was decreased in AGS/CDDP cells transfected with
shMCM3AP-AS1#1 or miR-138 mimics, but this effect was counteracted
by overexpressing FOXC1. Rescue assays were performed to examine
whether the MCM3AP-AS1/miR-138/FOXC1 axis affected the resistance
of GC cells to CDDP. Subsequently, wound healing, Transwell and
CCK-8 assay results identified that the overexpression of FOXC1
could reverse the decrease in cellular migration, invasion and
proliferation caused by MCM3AP-AS1-knockdown or miR-138 mimics in
AGS/CDDP cells (Fig. 5D-F). In
addition, FOXC1 overexpression partially abrogated the inhibitory
effect of MCM3AP-AS1-knockdown or miR-138 overexpression on the
IC50 in AGS/CDDP cells (Fig.
5G). Therefore, it was indicated that the MCM3AP-AS1/miR-138
axis regulated CDDP resistance of GC cells via FOXC1.
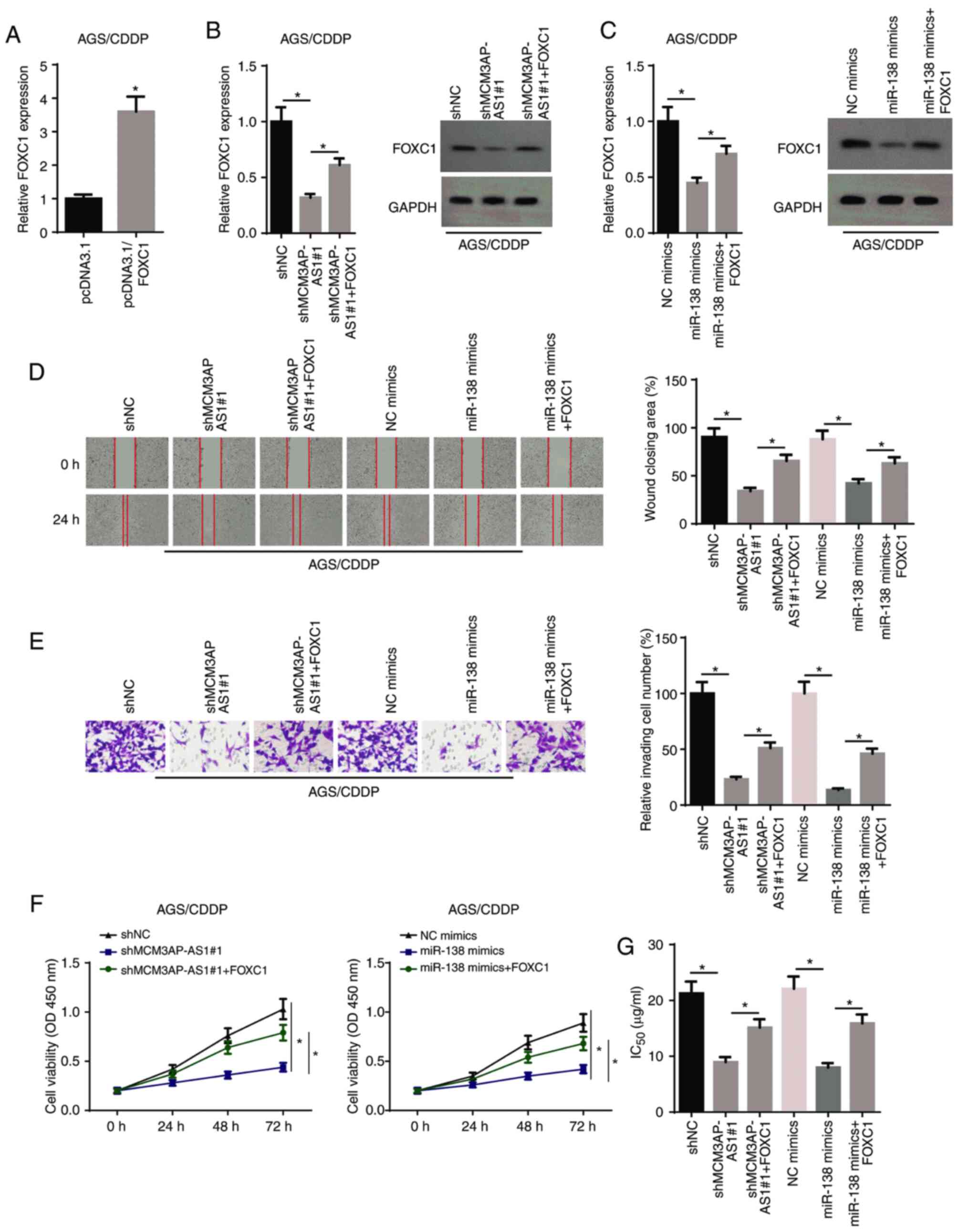 | Figure 5.MCM3AP-AS1/miR-138 increases gastric
cancer cell resistance to CDDP via regulating FOXC1. (A) FOXC1
expression in AGS/CDDP cells transfected with pcDNA3.1 and
pcDNA3.1-FOXC1. (B) FOXC1 expression was assessed in AGS/CDDP cells
transfected with shNC, shMCM3AP-AS1#1 and shMCM3AP-AS1#1+FOXC1
groups by RT-qPCR and western blot assays. (C) FOXC1 expression was
assessed in AGS/CDDP cells transfected with NC mimics, miR-138
mimics and miR-138 mimics+FOXC1 groups by RT-qPCR and western blot
assays. (D) Migratory (magnification, ×200) and (E) invasive
abilities (magnification, ×200) of AGS/CDDP cells were detected in
different groups by wound healing and Transwell assays,
respectively. (F) Cell viability of AGS/CDDP cells was measured in
shNC, shMCM3AP-AS1#1 and shMCM3AP-AS1#1+FOXC1 groups, as well as
NC, mimics, miR-138 mimics and miR-138 mimics+FOXC1 groups by Cell
Counting Kit-8 assay. (G) The IC50 of AGS/CDDP cells was
tested in shNC, shMCM3AP-AS1#1 and shMCM3AP-AS1#1+FOXC1 groups, as
well as NC, mimics, miR-138 mimics and miR-138 mimics+FOXC1 groups
by drug-sensitivity assay. *P<0.05. miR, microRNA; MCM3AP-AS1,
MCM3AP antisense RNA 1; CDDP, cisplatin; RT-qPCR, reverse
transcription-quantitative PCR; sh, short hairpin RNA; NC, negative
control; OD, optical density. |
Discussion
Chemoresistance is considered a major obstacle for
cancer therapy in the clinic (24).
lncRNAs are reported to be closely associated with chemoresistance
in human cancer. For example, lncRNA HOXD-AS1 contributes to CDDP
resistance in GC by epigenetically suppressing programmed cell
death 4 expression by recruiting enhancer of zeste homolog 2
(25). Moreover, lncRNA NEAT1
enhances paclitaxel resistance of ovarian cancer via the
miR-194/ZEB1 axis (26). Although
MCM3AP-AS1 has been reported to act as an oncogene in various types
of cancer, the biological role of lncRNA MCM3AP-AS1 in the
resistance of GC cells to CDDP remains unknown. The present study
demonstrated that MCM3AP-AS1 expression was upregulated in
CDDP-resistant GC cells. In addition, MCM3AP-AS1-knockdown
inhibited the migration, invasion and proliferation of
CDDP-resistant GC cells, suggesting that MCM3AP-AS1 enhanced CDDP
resistance in GC cells.
miRNAs are a class of endogenous non-coding RNAs
with a length of 22–25 nucleotides (27,28) and
they serve vital roles in various types of cancer, including GC.
For example, Hu et al (29)
reported that miR-4317 inhibited the proliferation of GC cells by
targeting zinc finger 322, while He and Zou (30) revealed that miR-96 promoted the
proliferation of GC cells by inhibiting FOXO3 expression. miR-138
has been shown to function as a tumor suppressor in various types
of cancer, such as breast, ovarian and non-small cell lung cancer
(31–33). In addition, several studies have
indicated that miR-138 is involved in the drug resistance of
multiple types of human cancer. For instance, Tang et al
(34) reported that miR-138
inhibited gefitinib resistance in non-small cell lung. Furthermore,
Li et al (35) observed that
LINC00174-knockdown decreased chemoresistance in glioma cells by
downregulating miR-138 expression. The present study identified
that miR-138 was a downstream target of MCM3AP-AS1, and that a
miR-138 inhibitor could abolish the inhibitory effect of
MCM3AP-AS1-knockdown on the CDDP resistance of GC cells.
Previous studies have reported that lncRNAs may act
as ceRNAs by sponging miRNAs to release downstream mRNAs (36–38).
FOXC1, a member of the FOX transcription factor family, serves a
vital role in numerous types of cancer, such as colorectal,
non-small cell lung and prostate cancer (39–41).
With regards to GC, Xu et al (23) found that FOXC1 overexpression
contributed to poor prognosis in patients with GC. The present
results indicated that MCM3AP-AS1-knockdown inhibited FOXC1
expression by targeting miR-138 in GC. In addition, it was
identified that the overexpression of FOXC1 abolished the
inhibitory effects of MCM3AP-AS1-knockdown (or miR-138 mimics) on
CDDP resistance in GC.
In conclusion, to the best of our knowledge, the
present study was the first to investigate the role and potential
mechanism of MCM3AP-AS1 in the resistance of GC cells to CDDP. The
current findings demonstrated that MCM3AP-AS1 promoted CDDP
resistance in GC by sponging miR-138 to upregulate FOXC1
expression, which may provide a novel promising therapeutic
approach for GC treatment. However, in vivo studies and
associated clinical trials are required to verify and elucidate
this molecular mechanism.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
HS, PW and WC designed the present study. PW, BZ and
XW performed all the experiments, analyzed the data and prepared
the figures. HS and WC drafted the initial manuscript. WC and PW
reviewed and revised the manuscript. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wadhwa R, Taketa T, Sudo K, Blum MA and
Ajani JA: Modern oncological approaches to gastric adenocarcinoma.
Gastroenterol Clin North Am. 42:359–369. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ashraf N, Hoffe S and Kim R: Adjuvant
treatment for gastric cancer: Chemotherapy versus radiation.
Oncologist. 18:1013–1021. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ghosh S: Cisplatin: The first metal based
anticancer drug. Bioorg Chem. 88:1029252019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Galluzzi L, Senovilla L, Vitale I, Michels
J, Martins I, Kepp O, Castedo M and Kroemer G: Molecular mechanisms
of cisplatin resistance. Oncogene. 31:1869–1883. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen LL: Linking long noncoding RNA
localization and function. Trends Biochem Sci. 41:761–772. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Prensner JR and Chinnaiyan AM: The
emergence of lncRNAs in cancer biology. Cancer Discov. 1:391–407.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wu M, Huang Y, Chen T, Wang W, Yang S, Ye
Z and Xi X: lncRNA MEG3 inhibits the progression of prostate cancer
by modulating miR-9-5p/QKI-5 axis. J Cell Mol Med. 23:29–38. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gao W, Weng T, Wang L, Shi B, Meng W, Wang
X, Wu Y, Jin L and Fei L: Long non-coding RNA NORAD promotes cell
proliferation and glycolysis in non-small cell lung cancer by
acting as a sponge for miR1365p. Mol Med Rep. 19:5397–5405.
2019.PubMed/NCBI
|
|
12
|
Xu N, Qiao L, Yin L and Li H: Long
noncoding RNA ROR1-AS1 enhances lung adenocarcinoma metastasis and
induces epithelial-mesenchymal transition by sponging miR-375. J
BUON. 24:2273–2279. 2019.PubMed/NCBI
|
|
13
|
Zhang M, Jiang X, Jiang S, Guo Z, Zhou Q
and He J: lncRNA FOXD2-AS1 regulates miR-25-3p/Sema4c axis to
promote the invasion and migration of colorectal cancer cells.
Cancer Manag Res. 11:10633–10639. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xi Z, Si J and Nan J: lncRNA MALAT1
potentiates autophagy-associated cisplatin resistance by regulating
the microRNA-30b/autophagy-related gene 5 axis in gastric cancer.
Int J Oncol. 54:239–248. 2019.PubMed/NCBI
|
|
15
|
Guo Y, Yue P, Wang Y, Chen G and Li Y:
PCAT-1 contributes to cisplatin resistance in gastric cancer
through miR-128/ZEB1 axis. Biomed Pharmacother. 118:1092552019.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu L and Wang S: Long non-coding RNA
OIP5-AS1 knockdown enhances CDDP sensitivity in osteosarcoma via
miR-377-3p/FOSL2 axis. Onco Targets Ther. 13:3853–3866. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang M, Sun S, Guo Y, Qin J and Liu G:
Long non-coding RNA MCM3AP-AS1 promotes growth and migration
through modulating FOXK1 by sponging miR-138-5p in pancreatic
cancer. Mol Med. 25:552019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang H, Luo C and Zhang G: lncRNA
MCM3AP-AS1 regulates epidermal growth factor receptor and autophagy
to promote hepatocellular carcinoma metastasis by interacting with
miR-455. DNA Cell Biol. 38:857–864. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen Q, Xu H, Zhu J, Feng K and Hu C:
lncRNA MCM3AP-AS1 promotes breast cancer progression via modulating
miR-28-5p/CENPF axis. Biomed Pharmacother. 128:1102892020.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kong X, Duan Y, Sang Y, Li Y, Zhang H,
Liang Y, Liu Y, Zhang N and Yang Q: lncRNA-CDC6 promotes breast
cancer progression and function as ceRNA to target CDC6 by sponging
microRNA-215. J Cell Physiol. 234:9105–9117. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li B, Mao R, Liu C, Zhang W, Tang Y and
Guo Z: lncRNA FAL1 promotes cell proliferation and migration by
acting as a CeRNA of miR-1236 in hepatocellular carcinoma cells.
Life Sci. 197:122–129. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xu Y, Shao QS, Yao HB, Jin Y, Ma YY and
Jia LH: Overexpression of FOXC1 correlates with poor prognosis in
gastric cancer patients. Histopathology. 64:963–970. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hu Y, Zhu QN, Deng JL, Li ZX, Wang G and
Zhu YS: Emerging role of long non-coding RNAs in cisplatin
resistance. Onco Targets Ther. 11:3185–3194. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ye Y, Yang S, Han Y, Sun J, Xv L, Wu L and
Ming L: HOXD-AS1 confers cisplatin resistance in gastric cancer
through epigenetically silencing PDCD4 via recruiting EZH2. Open
Biol. 9:1900682019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
An J, Lv W and Zhang Y: lncRNA NEAT1
contributes to paclitaxel resistance of ovarian cancer cells by
regulating ZEB1 expression via miR-194. Onco Targets Ther.
10:5377–5390. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mendell JT: MicroRNAs: Critical regulators
of development, cellular physiology and malignancy. Cell Cycle.
4:1179–1184. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
He L and Hannon GJ: MicroRNAs: Small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hu X, Zhang M, Miao J, Wang X and Huang C:
miRNA-4317 suppresses human gastric cancer cell proliferation by
targeting ZNF322. Cell Biol Int. 42:923–930. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
He X and Zou K: miRNA-96-5p contributed to
the proliferation of gastric cancer cells by targeting FOXO3. J
Biochem. 167:101–108. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yeh YM, Chuang CM, Chao KC and Wang LH:
MicroRNA-138 suppresses ovarian cancer cell invasion and metastasis
by targeting SOX4 and HIF-1α. Int J Cancer. 133:867–878. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang J, Liu D, Feng Z, Mao J, Zhang C, Lu
Y, Li J, Zhang Q, Li Q and Li L: MicroRNA-138 modulates metastasis
and EMT in breast cancer cells by targeting vimentin. Biomed
Pharmacother. 77:135–141. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhu D, Gu L, Li Z, Jin W, Lu Q and Ren T:
miR-138-5p suppresses lung adenocarcinoma cell
epithelial-mesenchymal transition, proliferation and metastasis by
targeting ZEB2. Pathol Res Pract. 215:861–872. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tang X, Jiang J, Zhu J, He N and Tan J:
HOXA4-regulated miR-138 suppresses proliferation and gefitinib
resistance in non-small cell lung cancer. Mol Genet Genomics.
294:85–93. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li B, Zhao H, Song J, Wang F and Chen M:
LINC00174 down-regulation decreases chemoresistance to temozolomide
in human glioma cells by regulating miR-138-5p/SOX9 axis. Hum Cell.
33:159–174. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chan JJ and Tay Y: Noncoding RNA:RNA
regulatory networks in cancer. Int J Mol Sci. 19:13102018.
View Article : Google Scholar
|
|
37
|
Chen M, Zhang R, Lu L, Du J, Chen C, Ding
K, Wei X, Zhang G, Huang Y and Hou J: lncRNA PVT1 accelerates
malignant phenotypes of bladder cancer cells by modulating
miR-194-5p/BCLAF1 axis as a ceRNA. Aging (Albany NY).
12:22291–22312. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang Y, Li W, Lin Z, Hu J, Wang J, Ren Y,
Wei B, Fan Y and Yang Y: The long noncoding RNA Linc01833 enhances
lung adenocarcinoma progression via miR-519e-3p/S100A4 axis. Cancer
Manag Res. 12:11157–11167. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yun SH, Han SH and Park JI: COUP-TFII
knock-down promotes proliferation and invasion in colorectal cancer
cells via activation of Akt pathway and up-regulation of FOXC1.
Anticancer Res. 40:177–190. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Gong R, Lin W, Gao A, Liu Y, Li J, Sun M,
Chen X, Han S, Men C, Sun Y and Liu J: Forkhead box C1 promotes
metastasis and invasion of non-small cell lung cancer by binding
directly to the lysyl oxidase promoter. Cancer Sci. 110:3663–3676.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Huang H, Xiong Y, Wu Z, He Y, Gao X, Zhou
Z and Wang T: MIR-138-5P inhibits the progression of prostate
cancer by targeting FOXC1. Mol Genet Genomic Med. 8:e11932020.
View Article : Google Scholar : PubMed/NCBI
|