Introduction
Glioma, in particular glioblastoma (GBM), is the
most immunosuppressive and lethal malignant brain tumor in adults,
and accounts for >30% of intracranial tumors current standard
treatment options for malignant gliomas are multimodal and include
surgical resection, postoperative radiotherapy and concomitant
chemotherapy with temozolomide (1).
Despite advancements in molecular understanding and therapies, the
clinical benefits of glioma treatments remain limited (2–4), and the
life expectancy of patients with GBM is only extended to ~15 months
(5).
It has been reported that glioma-associated
mesenchymal stem cells (gb-MSCs) express classical MSC surface
markers, including CD105, CD73, CD90 and CD44, and lack the
expression of CD14, CD34 and CD45. Furthermore, gb-MSCs can adhere
to plastic and have the capacity to differentiate into osteoblasts,
adipocytes and chondroblasts in vitro (6,7). In
addition, the percentage of gb-MSCs in high-grade glioma samples is
closely related to survival of patients with GBM (8).
Vascular tube formation plays a crucial role in
tumorigenesis (9). Human MSCs
derived from bone marrow play a critical role in GBM-induced
neovascularization (10). Previous
studies from our laboratory demonstrated that gb-MSCs are
considered as integral components involved in the pericyte
transition and tumor vascular formation in the glioma
microenvironment (7,11). However, the involvement of
glioma-secreted growth factors, such as transforming growth factor
β1 (TGF-β1), platelet-derived growth factor-BB (PDGF-BB) and
fibroblast growth factor 2 (FGF-2), in gb-MSC neovascularization
and how they relate to vessel stabilization and contribute to
malignancy, remain unclear. The present study aimed therefore to
understand the role of growth factors, including PDGF-BB and
TGF-β1, in gb-MSC neovascularization. gb-MSCs may thus be
considered as an anti-angiogenic treatment strategy in patients
with GBM.
A better understanding of the molecular mechanism of
gb-MSC angiogenesis is necessary for the development of a clinical
tumor targeting strategy for glioma gene therapy. Certain
cytokines, including vascular endothelial cell growth factor
(VEGF), interleukin-6, interleukin-8, endothelial growth factors,
TGF-β1, FGF-2 and PDGF-BB, are released from glioma cells and have
been reported to mediate tumor angiogenesis (12,13).
Subsequently, the present study investigated the angiogenic
capacity of TGF-β1 and PDGF-BB on gb-MSCs.
Materials and methods
Isolation and culture of gb-MSCs
gb-MSCs were isolated from glioma samples that were
collected from patients with GBM treated at the Neurosurgery Center
in Wuhan Union Hospital and were processed within 1 h. The age and
sex of patients are not limited and the grade was III–IV. Briefly
(7), glioma samples were washed
three times using PBS to remove blood and impurities and were cut
into 1–2 mm pieces. Tissue samples were placed into culture dishes
and were digested using collagenase (Biyuntian Biotechnology Co.,
Ltd.) for 20 min at room temperature. Samples were filtered using a
70 µm nylon mesh (Pall Life Sciences) and centrifuged at 350 × g
for 10 min at room temperature. The mononuclear cells were
collected following sample centrifugation through a Ficoll (2:1;
Genview Corp.) density gradient at 350 × g for 20 min at 4°C
Finally, cells were washed with PBS three times, cultured in DMEM
(HyClone) supplemented with 10% FBS (Biological Industries), 100
µ/ml penicillin and 100 µ/ml streptomycin (Gibco; Thermo Fisher
Scientific, Inc.) and placed in a humidified atmosphere at 37°C
with 5% CO2.
Collection of conditioned medium
The human U87 glioblastoma cell line of unknown
origin was purchased from the American Type Culture Collection
(cat. no. HTB-14) and was identified by STR profiling. U87 and
gb-MSCs were cultured in DMEM (HyClone; GE Healthcare Life
Sciences) supplemented with 10% FBS (Biological Industries) and 100
µ/ml penicillin and 100 µ/ml streptomycin (Gibco; Thermo Fisher
Scientific, Inc.) in 25 cm2 culture flasks and placed in
a humidified atmosphere at 37°C with 5% CO2. When cells
reached 50–60% confluence, they were washed three times with PBS
and cultured in serum-free medium (0% DMEM) for 3 days.
Subsequently, the conditioned medium (0% gb-CM, MSC-CM) was
collected and centrifuged at 1,000 × g for 10 min at room
temperature to remove cell debris. Aliquots of conditioned medium
were stored at −20°C until they further use (tube formation and
ELISA assays).
Differentiation protocols
gb-MSCs were differentiated into osteocytes,
adipocytes and chondrocytes following treatment with specific
osteogenic, adipogenic or chondrogenic induction agents and
maintenance media (all from StemCell Technologies, Inc.). These
experiments were performed as previously described (7). Oil red O staining was used to examine
adipogenic differentiation, alizarin red staining was used to
investigate osteogenic differentiation and alcian blue staining was
used to detect chondrogenic differentiation (all from
Sigma-Aldrich; Merck KGaA). The standard medium was used as the
control group.
Flow cytometric analysis
In order to characterize gb-MSCs, cells were
digested using collagenase (Biyuntian Biotechnology Co., Ltd.) at
37°C, neutralized using complete media and finally centrifuged at
1,000 × g for 5 min at room temperature. Supernatant was removed
and the pellets were resuspended in fluorescent-activated cell
sorting (FACS) buffer (Miltenyi Biotec GmbH). These single-cell
suspensions were incubated in the dark at 4°C for 30 min with
FITC-, PE-Cy7-, APC-Cy7-, Percp- and APC-conjugated antibodies
against human CD105, CD44, CD14, CD34 and CD31 (all from
eBioscience; Thermo Fisher Scientific, Inc.). Subsequently, cells
were centrifuged at 1,000 × g for 5 min at room temperature,
resuspended in PBS and analyzed using a FACS flow cytometer (BD
Biosciences). The data were collected and analyzed using FlowJo V10
software (Tree Star, Inc.).
High-density micromass cultures of
gb-MSCs
To investigate the chondrogenic differentiation of
gb-MSCs in high-density pellet cultures (P3), complete medium
containing a total of 2×105 gb-MSCs was placed in a 15
ml polypropylene tube (Corning Inc.) and were centrifuged at 300 ×
g for 10 min. The cell pellets were collected and cultured in
chondrogenic medium (Stemcell Technologies, Inc.) for 21 days, and
half of the medium was changed during differentiation every three
days. Cartilage nodules were formed by these cell pellets and they
were subjected to standard paraffin embedding methods. The sections
were then cut into 6 µm slices and were subjected to
hematoxylin-eosin staining (H&E) staining, The slides were
rinsed in PBS, incubated overnight at 4°C with anti-SOX-9 (1:100;
cat. no. BS-4177R) and anti-collagen II (1:100; cat. no.
15943-1-AP) antibodies (Wuhan Sanying Biotechnology), and then
incubated with an HRP-conjugated secondary antibody (1:1; cat. no.
Ab7090; Wuhan Boster Biological Technology, Ltd.). Binding was
detected using a DAB solution (Wuhan Boster Biological Technology,
Ltd.). The tissues were counterstained using haematoxylin for 1 min
at room temperature (Wuhan Boster Biological Technology, Ltd.).
Images of the stained tissue samples were obtained using an Olympus
light microscope (Olympus Corporation). For immunofluorescence,
non-specific staining was blocked by pre-incubation with 5% goat
serum (cat. no. AR1009) diluted in PBS for 30 min at room
temperature. The primary antibodies used were as follows: goat
anti-SOX-9 polyclonal antibody (1:200; Abcam; cat. no. ab185966)
and rabbit anti-collagen II monoclonal antibody (1:100; Wuhan
Boster Biological Technology, Ltd.; cat. no. ab3092). After
incubation with the primary antibody overnight at 4°C, sections
were rinsed several times with PBS and incubated with the
appropriate secondary antibodies at room temperature for 1 h. The
secondary antibodies used were as follows: Cy3-conjugated goat
anti-rabbit (1:100; cat. no. BA1032) and FITC-conjugated goat
anti-goat antibodies (1:100; Wuhan Boster Biological Technology,
Ltd.; cat. no. BA1101). After washing with PBS, the sections were
counterstained with DAPI (Beyotime Institute of Biotechnology) and
mounted with anti-fade mounting medium. Immunofluorescence
microscopy was performed with an Olympus light microscope (Olympus
Corporation).
ELISA
ELISA kits (Neobioscience; cat. nos. EHC107b.96.10
and EHC181.96) were used to measure the levels of TGF-β1 and
PDGF-BB in the supernatants of gb-MSCs. All procedures were
performed according to the manufacturers' instructions. The
absorbance was measured at 450 nm using a microplate reader. Each
sample was assessed in triplicate.
Tube formation assay
Angiogenesis assays were performed according to
consensus guidelines (14). Matrigel
(BD Biosciences) was added to each well of flat-bottomed prechilled
96-well plates (Corning Inc.). After incubation at 37°C in 5%
CO2 for 40 min, gb-MSCs were seeded (1.5×105
cells/well) into the wells with serum-free glioma conditioned
medium (0% gb-CM), serum-free medium (0% DMEM), serum-free medium
containing rhTGF-β1 or serum-free medium containing rhPDGF-BB, and
plates were further incubated at 37°C in 5% CO2. Each
medium condition was assessed in triplicate. After 6 h, tube
formation was imaged using a light microscope (Olympus
Corporation). Capillary-like tube formation was analyzed in three
random fields of view per well using ImageJ software (National
Institutes of Health).
Immunofluorescence
gb-MSCs (5,000 cells/well) were cultured in an
eight-well chamber slide (Ibidi GmbH) at 37°C. Once cells reached
70% confluence, they were washed with PBS three times, fixed with
4% paraformaldehyde for 15 min at room temperature, permeabilized
with 0.5% Triton-X-100 for 20 min at room temperature and blocked
with a solution containing donkey serum (Antgene) for 1 h at room
temperature. Subsequently, cells were incubated with primary
antibodies against PDGFR and TGFβ1-R (R&D Systems, Inc.)
overnight at 4°C. After being washed three times with PBS, cells
were incubated with secondary antibodies labeled with rhodamine
(Antgene) for 1 h at room temperature. In each experiment,
non-specific staining by secondary antibodies was excluded by
incubating a well without primary antibodies. Cells were also
stained with DAPI for 5 min at room temperature (Antgene). Images
were acquired using a fluorescent microscope (Olympus Corporation)
and data were analyzed using Image-Pro Plus v6.0 (Media
Cybernetics, Inc.).
Statistical analysis
Data were expressed as the means ± standard
deviation. Three independent experiments were performed.
Statistical analyses were performed using Graph-Pad Prism software
6 (GraphPad Software, Inc.). Kruskal-Wallis one-way ANOVA was used
to compare variables followed by Dunnett's post hoc test if
required. P<0.05 was considered to indicate a statistically
significant difference.
Results
gb-MSC stemness assessment by surface
marker expression
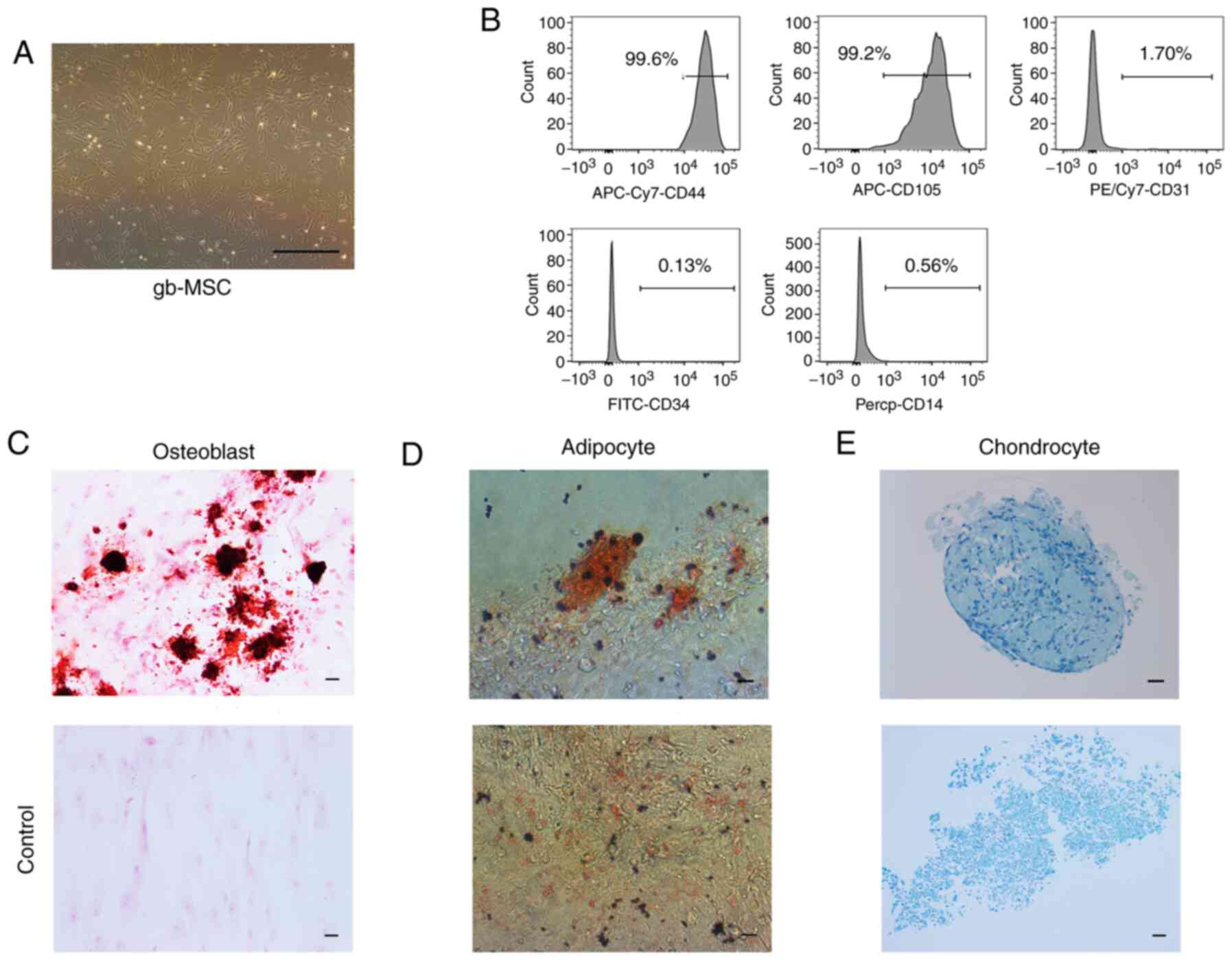
gb-MSCs displayed a fibroblastic morphology
consistent with that of MSCs (Fig.
1A). gb-MSCs were adherent in flasks containing complete
medium. To confirm the stemness of gb-MSCs, flow cytometry was used
to evaluate the expression of certain molecular markers from
passage 3 to passage 6. The results demonstrated that gb-MSCs
expressed CD44 and CD105, but did not express CD34, CD31 and CD14
(Fig. 1B).
Differentiation capacities of
gb-MSCs
In vitro, gb-MSCs can differentiate into
adipocytes, osteoblasts and chondrocytes under specific conditions,
further promoting adipogenesis, osteogenesis and chondrogenesis
(7), respectively. In the present
study, the differentiation of the gb-MSCs into three linages was
observed according to oil red O staining, alizarin red staining and
alcian blue staining (Fig.
1C-E).
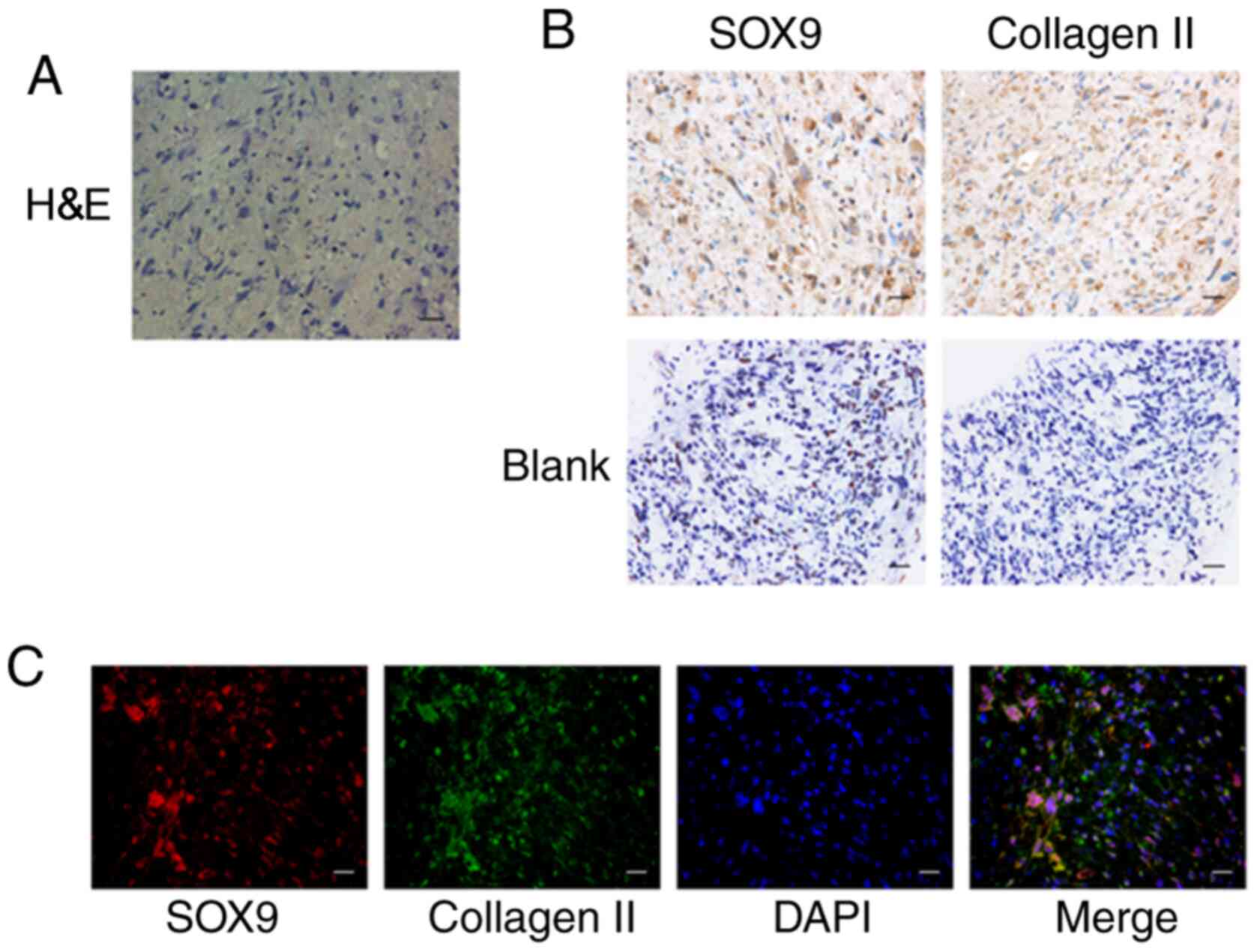
gb-MSCs can form cartilage structures
in micromass culture
The chondrogenic differentiation ability of gb-MSCs
was examined by high-density micromass cultures. This method
determined whether cells purified from human glioma specimens could
be used for cartilage tissue engineering applications. The results
demonstrated that gb-MSC micromass cultures could promote the
formation of proteoglycan (Figs. 1E
and 2A). Results from
immunohistochemistry staining demonstrated that collagen II
(extracellular) and SOX-9 (nuclear) were highly expressed in
micromass cultures compared with control (Fig. 2B). In addition, their coexpression in
micromass cultures was confirmed (Fig.
2C).
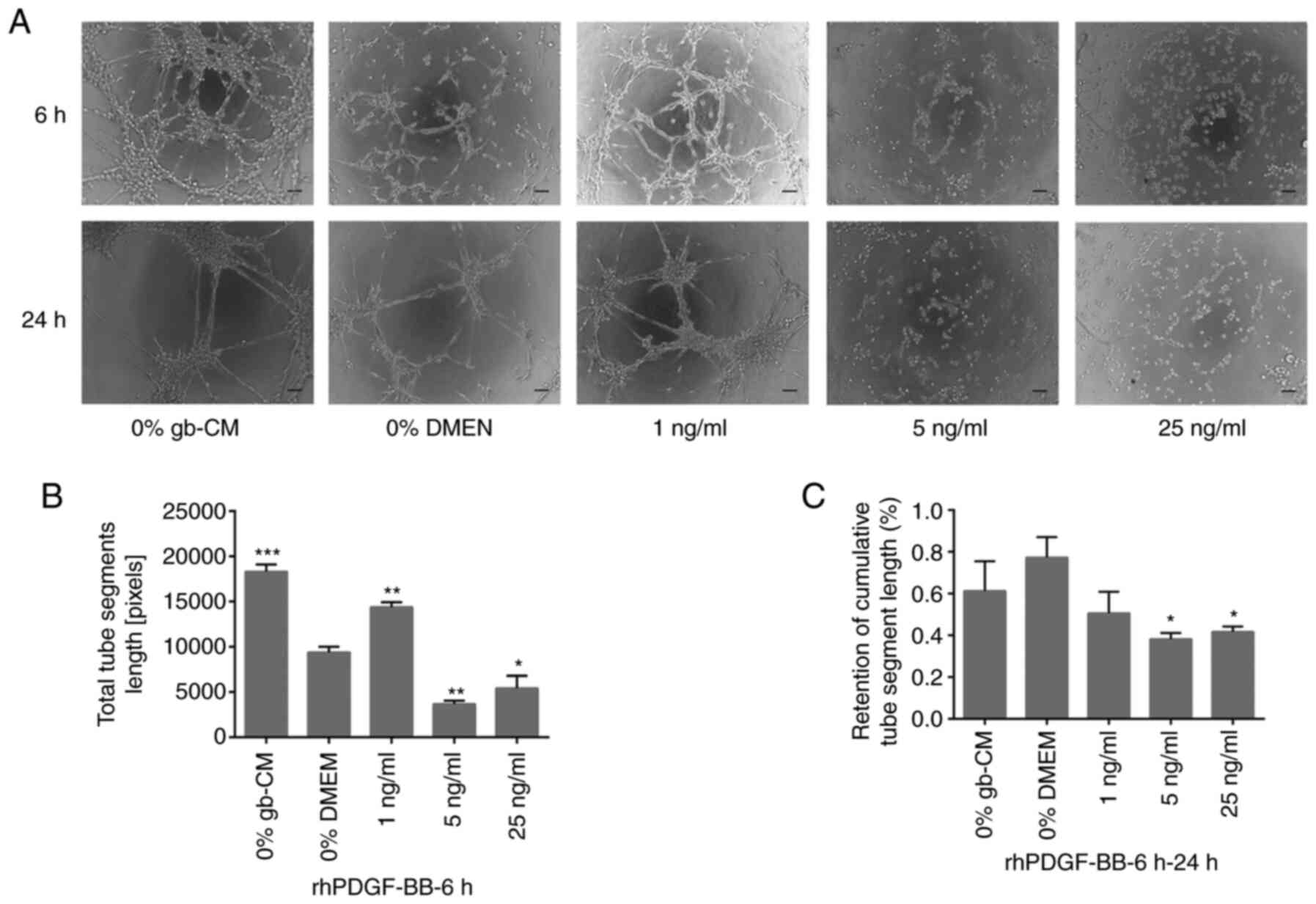
PDGF-BB and TGF-β1 significantly
improve the angiogenic capacity of gb-MSCs but decrease the
stabilization of newly formed tubes
To examine whether TGF-β1 and PDGF-BB could serve
important roles in vascularization, an in vitro tube
formation assay was performed using rhPDGF-BB and rhTGF-β1. The
results demonstrated a significant increase in gb-MSC
vascularization when cells were treated with 1 ng/ml rhPDGF-BB
after 6 h compared with 0% DMEM group, whereas higher concentration
of rhPDGF-BB (5 and 25 ng/ml) did not allow gb-MSC vascularization
(Fig. 3A and B). Furthermore, the
results demonstrated that more tubes remained intact in the 0%
gb-CM and % DMEM treated chambers compared with those in the
PDGF-BB (Fig. 3A and C).
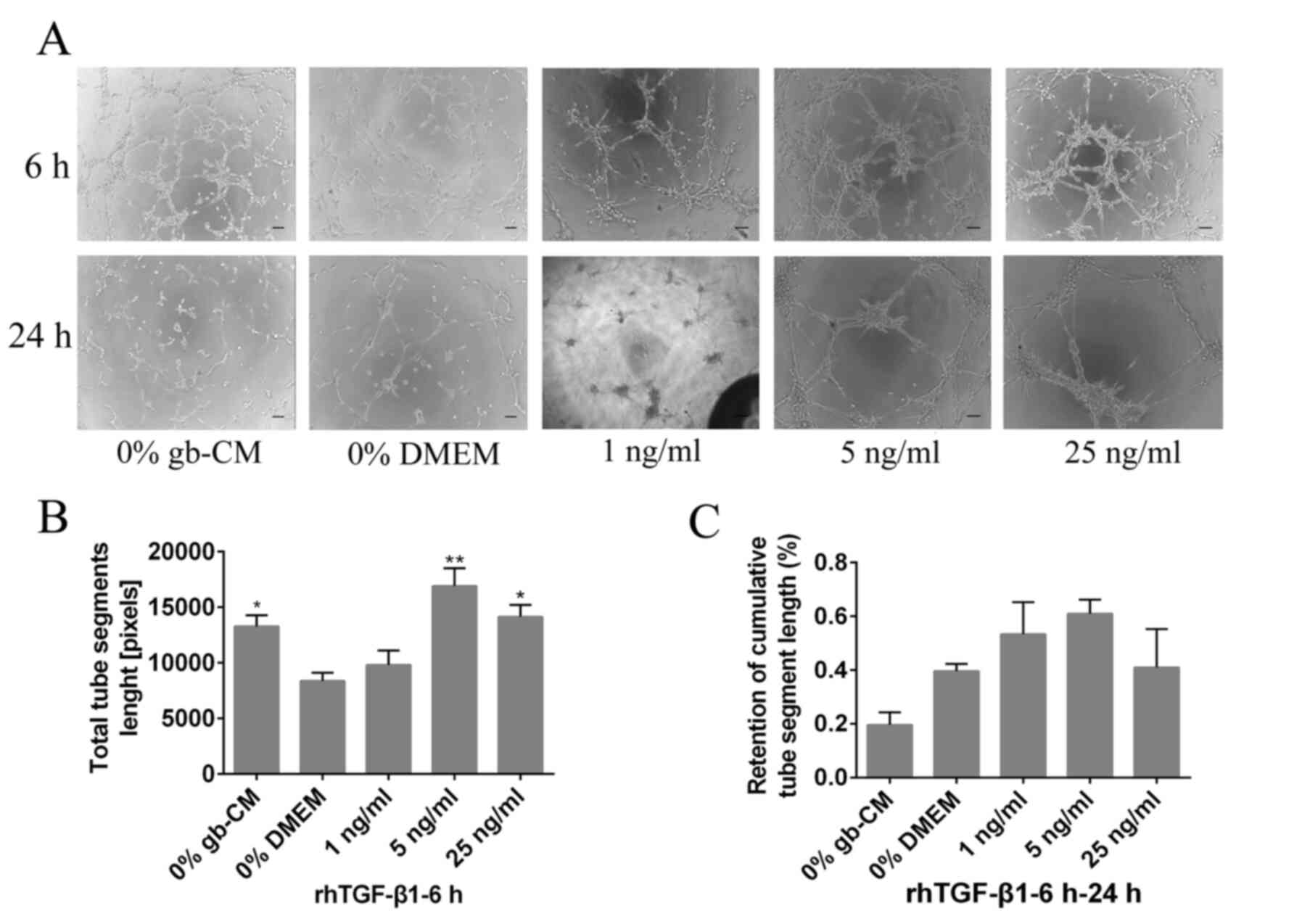
Furthermore, the results demonstrated that rhTGF-β1
could not increase significant tube formation at a lower
concentration (1 ng/ml) after 6 h, and the maximum vascularization
of gb-MSCs was observed with a 5 ng/ml rhTGF-β1 treatment (Fig. 4A and B). However, rhPDGF-BB (5 and 25
ng/ml) was able to significantly decrease the stabilization of
newly formed tubes compared with 0% DMEM group from 6 to 24 h
(Fig. 3A and C), and the tubes
formed in cells treated with the lower concentration of rhPDGF-BB
(1 or 5 ng/ml) remained intact more efficiently than those in the
0% DMEM-treated wells from 6 to 24 h (Fig. 4A and C).
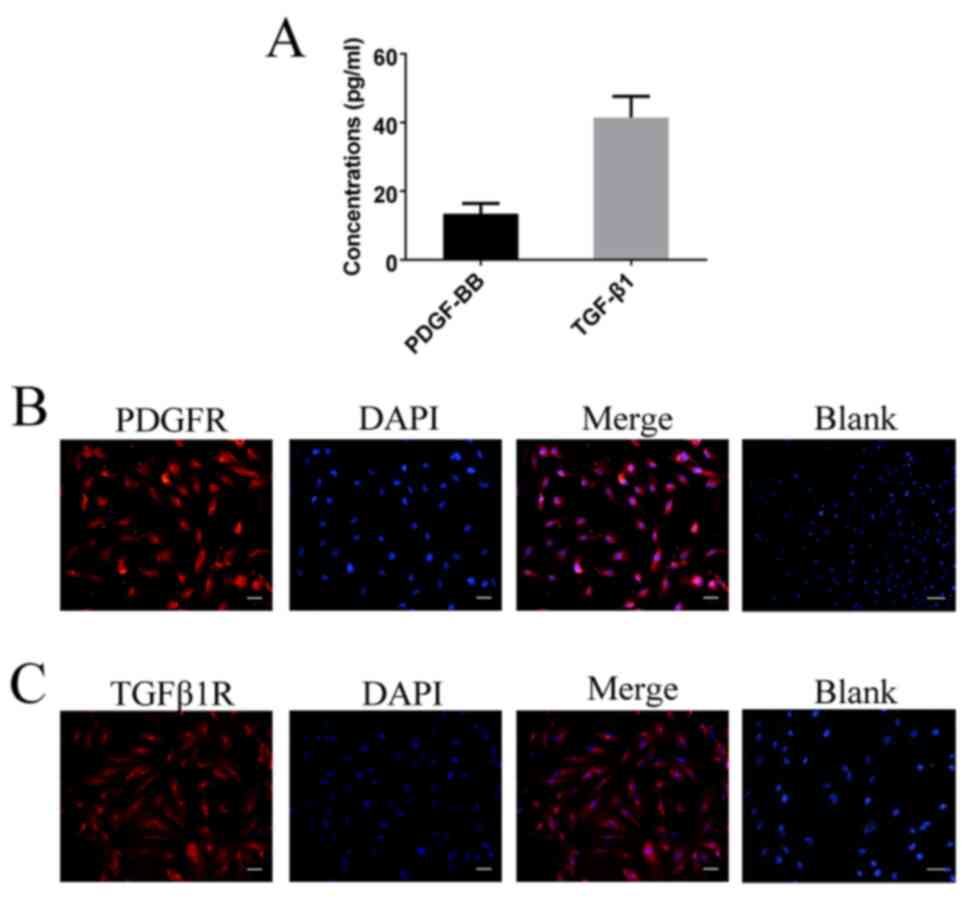
gb-MSCs express PDGF-BB, TGF-β1, PDGFR
and TGF-β1R
To determine the expression levels of PDGF-BB and
TGF-β1 in gb-MSCs, cell supernatant was collected and ELISA
experiments were performed. The results demonstrated that gb-MSCs
could secrete the growth factors PDGF-BB and TGF-β1 (Fig. 5A). Furthermore, the expression of
PDGFR and TGFβ1R was examined in gb-MSCs using immunofluorescence
(Fig. 5B and C).
Discussion
Previous studies have demonstrated that there are
two subpopulations of gb-MSCs that serve different roles in glioma
progression (7,11). The robust angiogenic capacity of
gb-MSCs could drive intracranial glioma development (7). A better understanding of the underlying
mechanisms of gb-MSC angiogenesis is therefore crucial for the
development of clinical treatments that target MSCs. The present
study demonstrated that specific growth factors, TGF-β1 and
PDGF-BB, could mediate the angiogenesis of gb-MSCs in vitro.
Previous studies from our laboratory reported some differences in
the expression of long non-coding RNAs and miRNAs in gb-MSCs
treated with different conditioned media. Furthermore, predicted
target genes were enriched for genes involved in angiogenesis
(7,11). This may suggest that the related
angiogenesis genes were upregulated to enhance the expression of
PDGF-BB and TGF-β1, further promoting angiogenic capacity by
specific signaling pathways.
TGF-β1, a member of the cytokine family that is
important for mediating the malignant phenotype of human brain
gliomas, is secreted by malignant gliomas in vitro and in
vivo (15,16). TGF-β1 expression is positively
correlated with higher-grade gliomas and with the induction of
angiogenesis (17). In addition,
TGF-β1 plays a significant role in glioma angiogenesis in
vivo (18,19). A previous study reported that TGF-β1
might be considered as a vital cytokine in MSC angiogenesis
(11). The results from the present
study demonstrated that TGFβ1R is expressed in gb-MSCs and that
gb-MSC angiogenesis was increased in response to TGF-β1. Therefore,
TGF-β1 may serve a crucial role in gb-MSC angiogenic capacity.
These findings were consistent with data from a recent study
demonstrated that TGF-β1 secreted by primary malignant glioma can
stimulate angiogenesis of MSCs (11).
The cytokine PDGF-BB, which binds to its receptor
PDGFR, plays a unique role in the regulation of angiogenesis
(20,21). Previous studies have demonstrated
that PDGF-BB contributes to angiogenesis in tumor tissues (21,22). Our
laboratory reported that PDGF-BB is secreted by glioma cells
(10) and serves a crucial role in
the regulation of MSC angiogenic capacity. It has also been
demonstrated that PDGF-BB/PDGFβR interaction plays a functional
role in tumor angiogenesis (20). In
the present study, the expression of PDGFR in gb-MSCs was assessed,
and gb-MSC angiogenic capacity in response to PDGF-BB was
determined. The results demonstrated that PDGF-BB played an
important role in stimulating the angiogenic capacity of gb-MSCs.
These findings were consistent with data from a previous study
demonstrating that TGF-β1 can stimulate MSC angiogenesis (10). Taken together, these results
suggested that TGF-β1 and PDGF-BB may contribute to the modulation
of gb-MSC angiogenesis.
In conclusion, the present study successfully
isolated gb-MSCs from human glioma tissues and demonstrated that
their fibroblast-like morphology and surface markers were similar
to those of classic MSCs. In addition, the results demonstrated
that TGFβ1 and PDGF-BB could improve the angiogenic capacity of
gb-MSCs and promote the expression of the growth factor receptors
TGFβ-R1 and PDGFβR. Further investigation of the underlying
mechanisms of gb-MSC angiogenesis may provide novel insights into
targeting vessel formation to treat patients with glioma.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81572488).
Availability of data and material
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
QZ and WX designed the study, analyzed the data and
drafted the manuscript. BX, DY and HZ collected and sorted the data
and analyzed the related literature PF, managed the project,
generated the outline of the manuscript and revised the language.
All authors read and approved the final version.
Ethics approval and consent to
participate
The present study was approved by The Ethical
Committee of Tongji Medical College of Huazhong University of
Science and Technology (Wuhan, China; approval no. S207). Written
informed consent was obtained from all patients and the experiments
were conducted in accordance with the Declaration of Helsinki.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
gb-MSCs
|
glioma-associated mesenchymal stem
cells
|
|
MSCs
|
mesenchymal stem cells
|
|
GBM
|
glioblastoma
|
|
TGF-β1
|
transforming growth factor β1
|
|
PDGF-BB
|
platelet-derived growth factor-BB
|
|
FGF
|
fibroblast growth factor
|
|
VEGF
|
vascular endothelial cell growth
factor
|
References
|
1
|
Stupp R, Mason WP, van den Bent MJ, Weller
M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn
U, et al: Radiotherapy plus concomitant and adjuvant temozolomide
for glioblastoma. N Engl J Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fu P, He YS, Huang Q, Ding T, Cen YC, Zhao
HY and Wei X: Bevacizumab treatment for newly diagnosed
glioblastoma: Systematic review and meta-analysis of clinical
trials. Mol Clin Oncol. 4:833–838. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Holland EC: Glioblastoma multiforme: The
terminator. Proc Natl Acad Sci USA. 97:6242–6244. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Surawicz TS, Davis F, Freels S, Laws ER Jr
and Menck HR: Brain tumor survival: Results from the national
cancer data base. J Neurooncol. 40:151–160. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
DeAngelis LM: Brain tumors. N Engl J Med.
344:114–123. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hossain A, Gumin J, Gao F, Figueroa J,
Shinojima N, Takezaki T, Priebe W, Villarreal D, Kang SG, Joyce C,
et al: Mesenchymal stem cells isolated from human gliomas increase
proliferation and maintain stemness of glioma stem cells through
the IL-6/gp130/STAT3 pathway. Stem Cells. 33:2400–2415. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang Q, Yi DY, Xue BZ, Wen WW, Lu YP,
Abdelmaksou A, Sun MX, Yuan DT, Zhao HY, Xiong NX, et al: CD90
determined two subpopulations of glioma-associated mesenchymal stem
cells with different roles in tumour progression. Cell Death Dis.
9:11012018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shahar T, Rozovski U, Hess KR, Hossain A,
Gumin J, Gao F, Fuller GN, Goodman L, Sulman EP and Lang FF:
Percentage of mesenchymal stem cells in high-grade glioma tumor
samples correlates with patient survival. Neuro Oncol. 19:660–668.
2017.PubMed/NCBI
|
|
9
|
Birnbaum T, Roider J, Schankin CJ, Padovan
CS, Schichor C, Goldbrunner R and Straube A: Malignant gliomas
actively recruit bone marrow stromal cells by secreting angiogenic
cytokines. J Neurooncol. 83:241–247. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Birnbaum T, Hildebrandt J, Nuebling G,
Sostak P and Straube A: Glioblastoma-dependent differentiation and
angiogenic potential of human mesenchymal stem cells in vitro. J
Neurooncol. 105:57–65. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yi D, Xiang W, Zhang Q, Cen Y, Su Q, Zhang
F, Lu Y, Zhao H and Fu P: Human glioblastoma-derived mesenchymal
stem cell to pericytes transition and angiogenic capacity in
glioblastoma microenvironment. Cell Physiol Biochem. 46:279–290.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kargiotis O, Rao JS and Kyritsis AP:
Mechanisms of angiogenesis in gliomas. J Neurooncol. 78:281–293.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Keerl S, Gehmert S, Gehmert S, Song YH and
Alt E: PDGF and bFGF modulate tube formation in adipose
tissue-derived stem cells. Ann Plast Surg. 64:487–490. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Frei K, Gramatzki D, Tritschler I,
Schroeder JJ, Espinoza L, Rushing EJ and Weller M: Transforming
growth factor-β pathway activity in glioblastoma. Oncotarget.
6:5963–5977. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liubich LD, Kovalevska L, Lisyany MI,
Semenova VM, Malysheva TA, Stayno LP and Vaslovych VV: TGF-β1
expression by glioma C6 cells in vitro. Exp Oncol. 39:258–263.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ferrari G, Cook BD, Terushkin V, Pintucci
G and Mignatti P: Transforming growth factor-beta 1 (TGF-beta1)
induces angiogenesis through vascular endothelial growth factor
(VEGF)-mediated apoptosis. J Cell Physiol. 219:449–458. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kaminska B, Kocyk M and Kijewska M: TGF
beta signaling and its role in glioma pathogenesis. Adv Exp Med
Biol. 986:171–187. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang XJ, Chen GL, Yu SC, Xu C, Xin YH, Li
TT, Shi Y, Gu A, Duan JJ, Qian C, et al: TGF-beta1 enhances
tumor-induced angiogenesis via JNK pathway and macrophage
infiltration in an improved zebrafish embryo/xenograft glioma
model. Int Immunopharmacol. 15:191–198. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Battegay EJ, Rupp J, Iruela-Arispe L, Sage
EH and Pech M: PDGF-BB modulates endothelial proliferation and
angiogenesis in vitro via PDGF beta-receptors. J Cell Biol.
125:917–928. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cumpanas AA, Cimpean AM, Ferician O,
Ceausu RA, Sarb S, Barbos V, Dema A and Raica M: The involvement of
PDGF-B/PDGFRβ axis in the resistance to antiangiogenic and
antivascular therapy in renal cancer. Anticancer Res. 36:2291–2295.
2016.PubMed/NCBI
|
|
21
|
Xiong B, Gong LL, Zhang F, Hu MB and Yuan
HY: TGF beta1 expression and angiogenesis in colorectal cancer
tissue. World J Gastroenterol. 8:496–498. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xue Y, Lim S, Yang Y, Wang Z, Jensen LD,
Hedlund EM, Andersson P, Sasahara M, Larsson O, Galter D, et al:
PDGF-BB modulates hematopoiesis and tumor angiogenesis by inducing
erythropoietin production in stromal cells. Nat Med. 18:100–110.
2011. View
Article : Google Scholar : PubMed/NCBI
|