Introduction
Colorectal cancer (CRC) is the third most frequent
malignant tumor, with 1.85 million new cases per year, and the
second primary cause of death worldwide, with 881,000 deaths
estimated in 2018 (1).
5-fluorouracil (5-FU) has been widely used as a chemotherapeutic
agent for CRC (2). However, the
therapeutic effects of 5-FU on patients with CRC are very limited
due to the development of drug resistance (3). It is imperative to investigate the
potential mechanisms of CRC chemoresistance, which may help to
develop novel treatment strategies to improve patient outcomes.
MicroRNAs (miRNAs/miRs), a class of short endogenous
non-coding RNAs, serve key roles in gene regulation after
transcription via targeting the 3′-untranslated region (UTR) of
mRNAs (4). A previous study has
revealed the antitumor effects of miRNAs in numerous types of
cancer, such as chronic lymphocytic leukemia, prostate cancer and
gastrointestinal stromal tumors (5).
Dysregulation of miRNAs has been reported to exert a crucial
function in drug chemoresistance (6). For example, miR-20a overexpression
sensitizes breast cancer cells to chemotherapy drugs and inhibits
their proliferation (7). Wang et
al (8) demonstrated that miR-26b
was involved in 5-FU-induced apoptosis in CRC. Recent studies have
suggested that miR-9 behaves as a tumor suppressor in different
types of cancer, such as hepatocellular carcinoma (9), gastric cancer (10), acute myeloid leukemia (11) and CRC (12). Cekaite et al (13) observed low miR-9-5p expression in CRC
cells and found that miR-9-5p overexpression inhibits cell survival
and promotes apoptosis. Additionally, miR-9-5p represses CRC cell
migration and invasion via downregulating transmembrane-4-L6 family
1 expression (14). However, the
impact of miR-9-5p in CRC chemoresistance remains unclear.
The high mobility group A2 (HMGA2) gene is located
on human chromosome 12q14 (15).
Accumulating evidence has revealed that HMGA2 is highly expressed
in a number of malignant tumors, including lung cancer (16), breast cancer (17) and CRC (18). Previous studies have demonstrated
that high HMGA2 expression is associated with a poor prognosis and
a lower survival rate in patients with CRC (18,19).
Furthermore, Xu et al (20)
revealed that HMGA2 contributes to 5-FU chemoresistance in CRC via
the dishevelled segment polarity protein 2/Wnt signaling pathway.
Based on the aforementioned studies, the present study speculated
that miR-9-5p may modulate 5-FU resistance in CRC by regulating
HMGA2 expression.
In the current study, the function and underlying
mechanism of miR-9-5p in 5-FU resistance in two CRC cell lines,
including HCT-116 and HT-29 cells, were investigated.
Materials and methods
Cell culture
According to previous studies (21,22),
human CRC HCT-116 and HT-29 cell lines were chosen to evaluate the
effect of miR-9-5p in CRC chemoresistance. HCT-116 and HT-29 cells
were obtained from Procell Life Science & Technology Co., Ltd.,
and maintained in McCoy's 5A medium (Procell Life Science &
Technology Co., Ltd.) containing 10% fetal bovine serum (FBS). 293T
cells (Shanghai Zhongqiao Xinzhou Biotechnology Co., Ltd.) were
incubated in Dulbecco's modified Eagle's medium (DMEM;
Sigma-Aldrich; Merck KGaA) containing 10% FBS. Cells were cultured
at 37°C with 5% CO2.
Cell transfection
Human miR-9-5p agomir (sense,
5′-UCUUUGGUUAUCUAGCUGUAUGA-3′ and antisense,
5′-AUACAGCUAGAUAACCAAAGAUU-3′) and its scrambled negative control
(NC; sense, 5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense,
5′-ACGUGACACGUUCGGAGAATT-3′), miR-9-5p antagomir
(5′-UCAUACAGCUAGAUAACCAAAGA-3′) and its scrambled NC
(5′-CAGUACUUUUGUGUAGUACAA-3′) were bought from Shanghai GenePharma
Co., Ltd. HCT-116 and HT-29 cells were seeded in a 12-well plate
(4×104 cells/well) and incubated overnight at 37°C with
5% CO2. At 70% confluency, HT-29 cells were transfected
with 100 pmol miR-9-5p agomir or agomir NC at room temperature for
48 h using Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) following the manufacturer's protocol.
Similarly, HCT-116 cells were transfected with 100 pmol miR-9-5p
antagomir or antagomir NC. An HMGA2-overexpression pcDNA3.1 plasmid
(over-HMGA2) was obtained from GenScript, whereas an empty pcDNA3.1
vector (GenScript) was applied as a NC (vector). A small
interference (si)RNA sequence of HMGA2 (si-HMGA2; sense,
5′-AGAGGCAGACCUAGGAAAUTT-3′ and antisense,
5′-ATTTCCTAGGTCTGCCTCTTT-3′) was bought from JTS Scientific, Ltd. A
scrambled siRNA (JTS Scientific, Ltd.) was used as a NC (siRNA NC;
sense, 5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense,
5′-ACGUGACACGUUCGGAGAATT-3′). A total of 1 µg of over-HMGA2 was
transfected alone or co-transfected with 50 pmol miR-9-5p agomir
into HT-29 cells at room temperature for 48 h using Lipofectamine
2000 following the manufacturer's protocol. HCT-116 cells were
transfected with 50 pmol si-HMGA2 or co-transfected with 50 pmol
miR-9-5p antagomir and si-HMGA2. Transfected cells were collected
48 h after incubation for subsequent experiments.
Hoechst staining
After fixation with 4% paraformaldehyde for 20 min
at room temperature, two CRC cell lines (HCT-116 and HT-29 cells)
were used to observe nuclear changes and apoptotic body formation
using the Hoechst staining kit (Nanjing KeyGen Biotech Co., Ltd.)
according to the manufacturer's protocol. Cells were visualized
under a fluorescence microscope (IX53; Olympus Corporation;
magnification, ×400) and counted by a professional researcher, who
was blind to the grouping. The percentage of apoptotic cells was
calculated as follows: Apoptotic cells (%)=(the number of apoptotic
cells/the number of total cells) ×100%.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA from HCT-116 and HT-29 cells was isolated
using the TRIpure lysis buffer (BioTeke Corporation). Subsequently,
RT to cDNA was performed using M-MLV Reverse Transcriptase kit
(cat. no. PR6502; BioTeke Corporation) according to the
manufacturer's protocol. qPCR reactions were performed using the
SYBR Green PCR kit (Sigma-Aldrich; Merck KGaA) according to the
manufacturer's protocol. The PCR reaction was run for an initial
denaturation for 5 min at 94°C, followed by 40 cycles of
denaturation at 94°C for 15 sec, annealing at 60°C for 25 sec and
extension at 72°C for 30 sec. Subsequently, the 2−ΔΔCq
comparative method was applied to determine the relative expression
levels of mRNA and miRNA (23). U6
was used as the internal reference for miR-9-5p, while β-actin
acted as the internal reference for HMGA2. The primer sequences
used are listed in Table I.
 | Table I.Primers used for reverse
transcription-quantitative PCR. |
Table I.
Primers used for reverse
transcription-quantitative PCR.
| Gene | Sequence
(5′-3′) |
|---|
| HMGA2 | F:
CTTCAGCCCAGGGACAAC |
|
| R:
TCCAGTGGCTTCTGCTTTC |
| β-actin | F:
CTTAGTTGCGTTACACCCTTTCTTG |
|
| R:
CTGTCACCTTCACCGTTCCAGTTT |
| U6 | F:
GCTTCGGCAGCACATATACT |
|
| R:
GTGCAGGGTCCGAGGTATTC |
| hsa-miR-9-5p | F:
GCAGCCTCTTTGGTTATCTAGC |
|
| R:
GTGCAGGGTCCGAGGTATTC |
Dual-luciferase reporter assay
TargetScan (http://www.targetscan.org) was used to predict the
potential target genes of miR-9-5p. For the miR-9-5p-target
analysis, a fragment of the HMGA2 3′-UTR containing either the
wild-type (WT) or mutated (Mut) binding sites for miR-9-5p was
cloned in a pmirGLO luciferase reporter vector (Promega
Corporation) following the manufacturer's instructions. 293T cells
were plated into 12-well plates and co-transfected using
Lipofectamine 2000 with luciferase reporter plasmid and miR-9-5p
agomir and agomir NC. After 4 h, cells were incubated in new DMEM
for 48 h at 37°C. Subsequently, luciferase activities were detected
using the Dual-Luciferase Reporter Assay kit (Promega Corporation)
following the manufacturer's protocol. Luciferase activity was
normalized to the Renilla luciferase activity.
Western blotting
Cells were lysed using the RIPA lysis buffer
(Beyotime Institute of Biotechnology) and the supernatants were
collected via centrifugation (10,000 × g for 5 min at 4°C). Protein
concentration was measured using the BCA Protein Concentration
Determination kit (Beyotime Institute of Biotechnology) following
the manufacturer's protocol. Protein separation (40 µg/lane) was
performed via 12 or 15% SDS-PAGE (Beyotime Institute of
Biotechnology). Proteins were transferred onto polyvinylidene
fluoride membranes (Thermo Fisher Scientific, Inc.) and blocked
with 5% (m/v) skimmed milk, followed by incubation with the
following primary antibodies at 4°C overnight: Polyclonal rabbit
anti-HMGA2 (1:1,000; cat. no. 20795-1-AP; ProteinTech Group, Inc.)
and monoclonal mouse anti-β-actin (1:1,000; cat. no. sc-47778;
Santa Cruz Biotechnology, Inc.). After washing with TBS-Tween (0.1%
Tween-20), proteins were incubated with HRP-conjugated goat
anti-rabbit IgG (1:5,000; cat. no. A0208; Beyotime Institute of
Biotechnology) and goat anti-mouse IgG (1:5,000; cat. no. A0216;
Beyotime Institute of Biotechnology) secondary antibodies for 40
min at 37°C. The protein bands were visualized using an enhanced
chemiluminescence kit (Beyotime Institute of Biotechnology)
according to the manufacturer's protocol, and quantified using the
Gel-Pro-Analyzer 4.0 software (Media Cybernetics, Inc.).
Cell counting Kit-8 (CCK-8) assay
Briefly, 4×103 cells/well were plated in
a 96-well plate. Subsequently, CRC cells were subjected to
distilled water (vehicle control group) or different concentrations
of 5-FU (0.625, 1.25, 2.5, 5, 10, 20, 40 and 80 µM; Shanghai
Aladdin Biochemical Technology Co., Ltd.). After 48 h at 37°C, the
CCK-8 assay kit (Beyotime Institute of Biotechnology) was used to
measure CRC cell viability at 37°C for 1 h following the
manufacturer's protocol. Optical density values at 450 nm were
detected using an ELX-800 microplate reader (BioTek Instruments,
Inc.; Agilent Technologies, Inc.).
Apoptosis assay
The Annexin V-FITC Apoptosis Detection kit (Nanjing
KeyGen Biotech Co., Ltd.) was used to measure apoptosis. After
washing two times with PBS, CRC cells were double-stained with
Annexin V-FITC and PI in the dark for 10 min at room temperature.
Transfected cells were subjected to 2.5 or 10 µM 5-FU. After 48 h,
apoptosis was measured via flow cytometry (NovoCyte; ACEA
Biosciences, Inc.; Agilent Technologies, Inc.) and analyzed using
NovoExpress 1.2.5 (ACEA Biosciences, Inc.; Agilent Technologies,
Inc.).
Caspase activity assay
Cells were lysed in cold lysis buffer and maintained
on ice for 15 min. After centrifugation at 16,000 × g for 15 min at
4°C, protein concentration in the cell supernatant was detected
using the Bradford Protein Concentration Determination kit
(Beyotime Institute of Biotechnology). Subsequently, caspase-3 and
activities of the CRC cells were measured using the Caspase 3
Activity Assay kit (cat. no. C1116; Beyotime Institute of
Biotechnology) and caspase-9 activities were detected using the
Caspase 9 Activity Assay kit (cat. no. BC3890; Beijing Solarbio
Science & Technology Co., Ltd.), following the manufacturers'
protocol.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism (v8.0; GraphPad Software, Inc.) and data were presented as
the mean ± SD (n=3). IC50 values were measured using
GraphPad Prism. The IC50 value represents the drug
concentration at which tumor cells are inhibited by half. Unpaired
two-tailed Student's t-test was used for comparisons between two
groups, while one-way or two-way ANOVA with Tukey's post-hoc test
was used for multiple comparisons to analyze statistically
significant differences. P<0.05 was considered to indicate a
statistically significant difference.
Results
5-FU upregulates miR-9-5p expression
in CRC cells
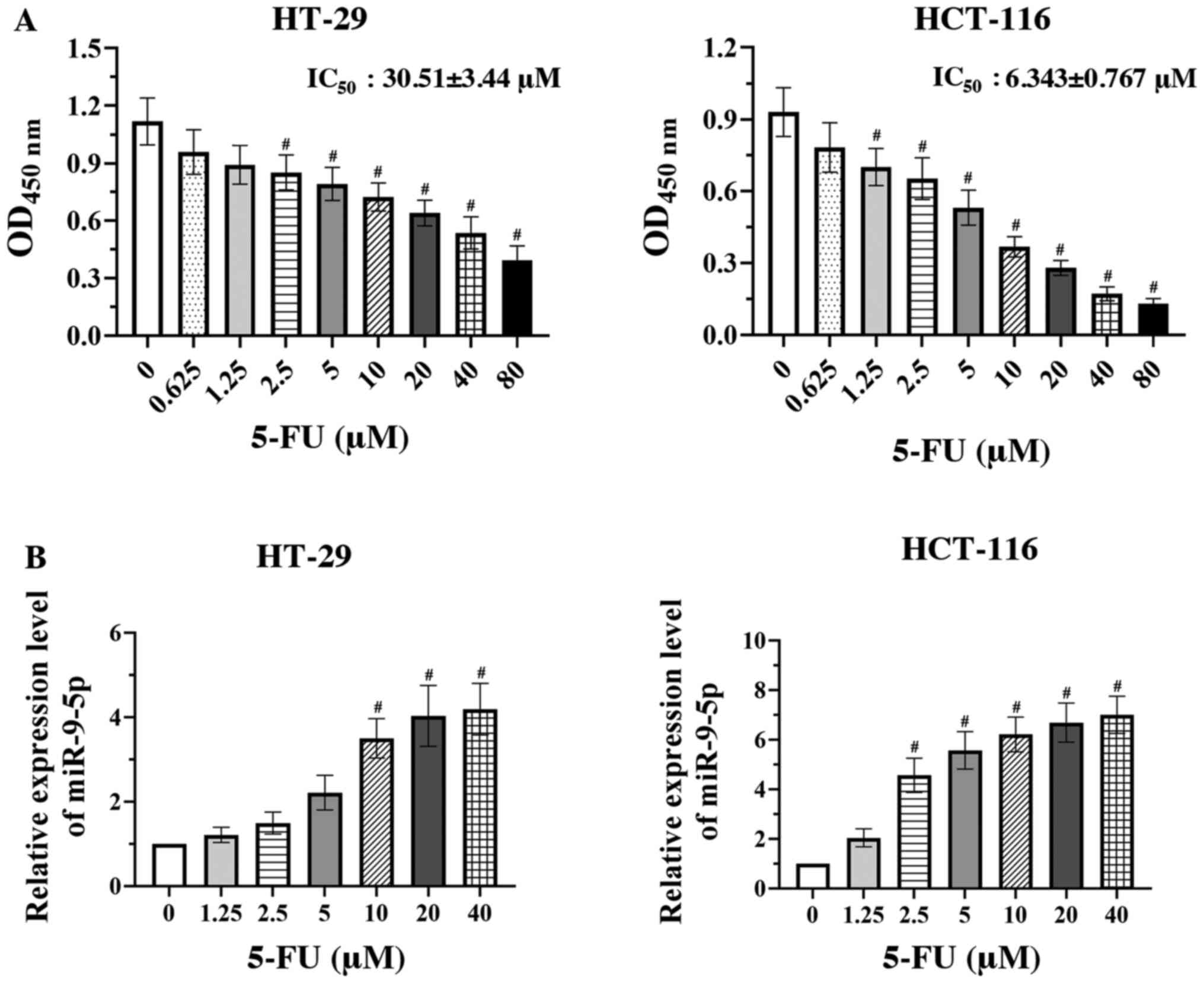
The effect of 5-FU on two CRC cell lines, including
HT-29 and HCT-116 cells, was assessed via CCK-8 assay. The cell
viability of both CRC cell lines was significantly decreased in a
concentration-dependent (0.65–80 µM) manner compared with the
vehicle group (0 µM 5-FU) (Fig. 1A;
P<0.05). HCT-116 cells were more sensitive to 5-FU than HT-29
cells (IC50 6.343±0.767 and 30.510±3.440 µM,
respectively). A previous study demonstrated that miR-9-5p
expression was markedly lower in CRC cells compared with that in
normal colorectal epithelial cells (13); thus, the present study further
assessed miR-9-5p expression in 5-FU-treated CRC cells. The RT-qPCR
results revealed that treatment with 5-FU significantly upregulated
miR-9-5p expression in both CRC cells in a dose-dependent manner
compared with the vehicle group (Fig.
1B; P<0.05).
miR-9-5p overexpression enhances CRC
cell sensitivity to 5-FU
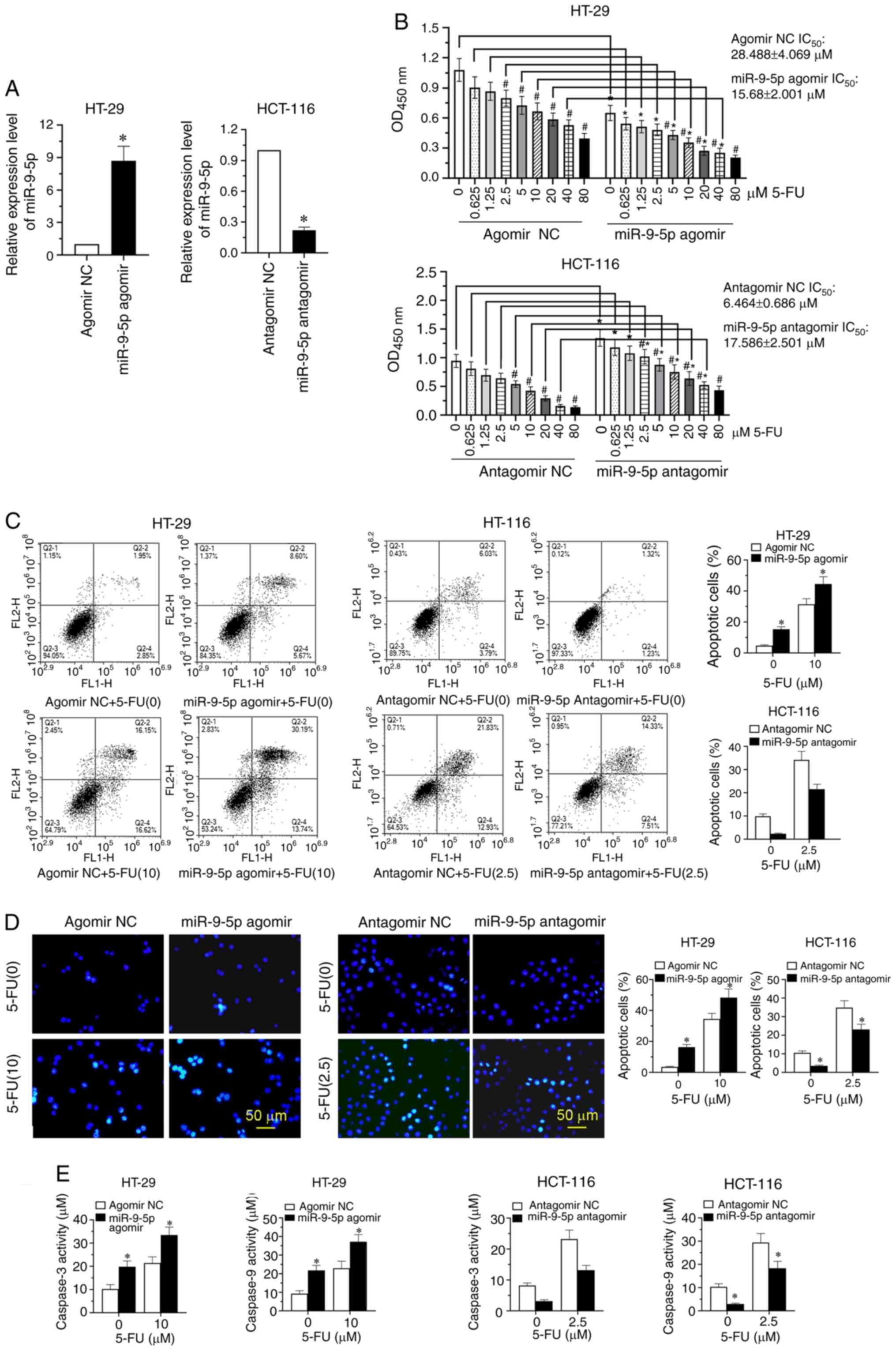
To explore the effect of miR-9-5p in overcoming 5-FU
resistance, HT-29 cells with a high IC50 value were
transfected with miR-9-5p agomir or agomir NC, and HCT-116 cells
with a low IC50 value were transfected with miR-9-5p
antagomir or antagomir NC. The results revealed that miR-9-5p
overexpression significantly increased miR-9-5p expression in HT-29
cells, while miR-9-5p expression was significantly downregulated in
HCT-116 transfected with antagomiR-9-5p (Fig. 2A; P<0.05). The CCK-8 analysis
indicated that after transfection with miR-9-5p agomirs and
antagomirs, treatment with 5-FU significantly decreased CRC cell
viability in a dose-dependent manner (Fig. 2B; P<0.05). Upregulation of
miR-9-5p significantly enhanced HT-29 cell sensitivity to 5-FU,
with the IC50 value decreasing from 28.488±4.069 µM in
the agomir NC group to 15.680±2.001 µM in the miR-9-5p agomir
group, whereas miR-9-5p-knockdown decreased HCT-116 cell
sensitivity to 5-FU, with the IC50 value increasing from
6.464±0.686 to 17.586±2.501 µM in the antagomir NC and miR-9-5p
antagomir groups, respectively (Fig.
2B; P<0.05). Additionally, based on the IC50
value of the two CRC cell lines (HT-29 and HCT-116), one-third of
the IC50 value (2.5 µM 5-FU for HCT-116 cells and 10 µM
5-FU for HT-29 cells) was chosen to explore the impact of miR-9-5p
on the sensitivity of CRC cells to 5-FU. The results revealed that
miR-9-5p overexpression significantly induced HT-29 apoptosis,
whereas knockdown of miR-9-5p significantly repressed the apoptosis
of HCT-116 cells (Fig. 2C;
P<0.05). Hoechst staining revealed that the nuclei of apoptotic
cells were densely stained after transfection with the miR-9-5p
agomir compared with the agomir NC, while treatment with miR-9-5p
antagomir decreased the staining number of apoptotic cells compared
with the antagomir NC (Fig. 2D).
Corresponding quantitative analysis revealed that the percentage of
apoptotic cells was significantly increased by miR-9-5p agomir and
significantly decreased by miR-9-5p antagomir compared with the
corresponding NCs (Fig. 2D;
P<0.05). Furthermore, the expression levels of
apoptosis-associated markers, including caspase-3 and caspase-9,
were significantly upregulated after miR-9-5p overexpression and
downregulated after miR-9-5p-knockdown (Fig. 2E; P<0.05).
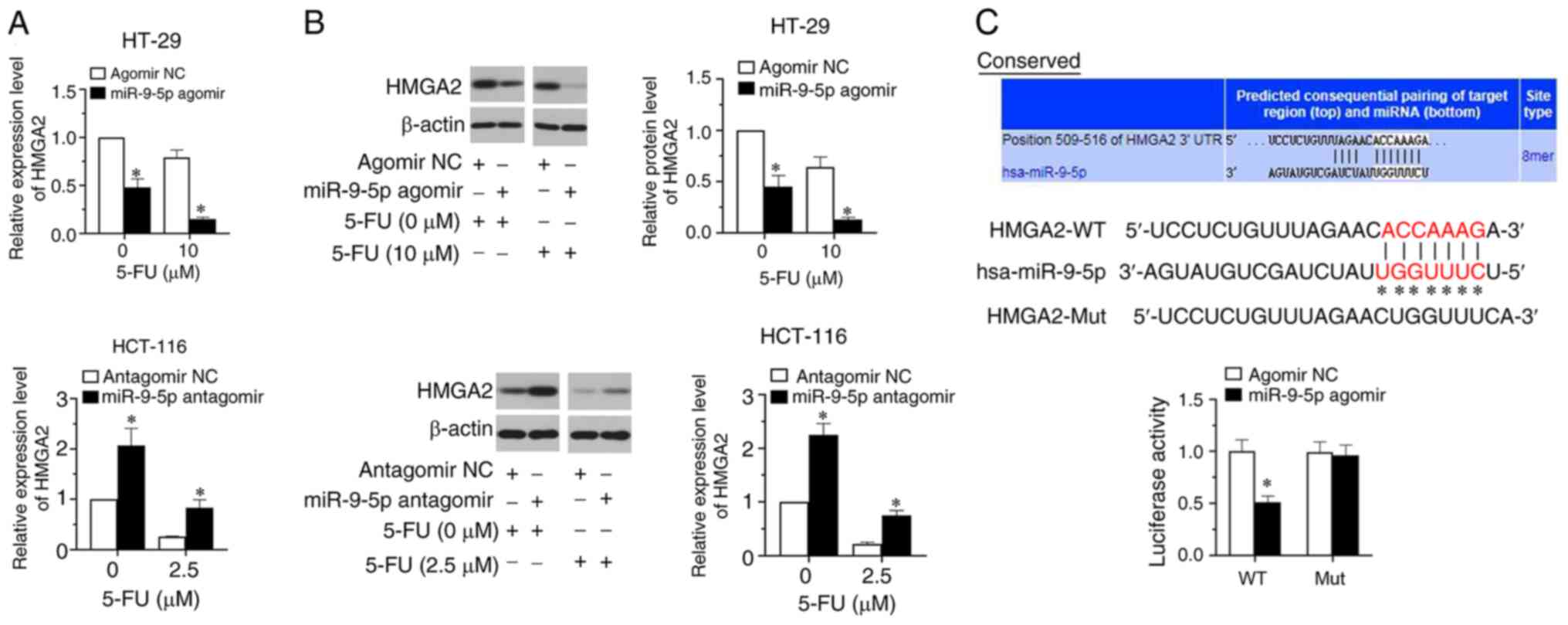
miR-9-5p directly targets HMGA2
To clarify the functional effect of miR-9-5p in CRC
cell sensitivity to 5-FU, bioinformatics analysis was applied to
predict its target genes. The results revealed that HMGA2 was one
of target genes of miR-9-5p. Therefore, HMGA2 expression was
determined by RT-qPCR and western blot assays. The results
indicated that compared with the vehicle group (0 µM 5-FU),
treatment with 5-FU decreased HMGA2 expression in the two CRC cell
lines (Fig. 3A and B). Additionally,
miR-9-5p overexpression significantly downregulated HMGA2
expression in HT-29 cells, whereas miR-9-5p silencing significantly
upregulated HMGA2 expression in HCT-116 cells compared with their
respective controls (Fig. 3A and B;
P<0.05). TargetScan predicted that miR-9-5p potentially bound to
HMGA2 mRNA. There was a targeted binding association between
miR-9-5p and HMGA2 (Fig. 3C).
Results of the luciferase reporter assay verified that miR-9-5p
overexpression significantly repressed the luciferase activity of
the reporter via binding to the WT but not the MUT 3′-UTR of HMGA2
(Fig. 3C; P<0.05).
Overexpression of HMGA2 reverses
miR-9-5p-induced HT-29 apoptosis
To confirm whether HMGA2 was involved in
miR-9-5p-mediated CRC drug resistance response,
HMGA2-overexpression plasmid was co-transfected with miR-9-5p
agomir into HT-29 cells. Additionally, a HMGA2 siRNA was
co-transfected with miR-9-5p antagomir into HCT-116 cells. Fig. 4A shows that HMGA2 expression was
significantly upregulated in HT-29 cells after transfection with
over-HMGA2 and significantly downregulated in si-HMGA2-transfected
HCT-116 cells (P<0.05). After co-transfection with miR-9-5p
agomir + vector/over-HMGA2 into HT-29 cells, and miR-9-5p antagomir
+ siRNA NC/si-HMGA2 into HCT-116 cells, CCK-8 assays revealed that
overexpression of HMGA2 significantly increased the viability of
miR-9-5p agomir-transfected HT-29 cells, whereas the viability of
miR-9-5p antagomir-transfected HCT-116 cells was significantly
inhibited by HMGA2-knockdown (Fig.
4B; P<0.05). Results of flow cytometry analysis exhibited
that the apoptotic rates in miR-9-5p agomir-transfected HT-29 cells
were significantly decreased after HMGA2 overexpression, while
increased apoptosis was observed in miR-9-5p antagomir-transfected
HCT-116 cells following downregulation of HMGA2 expression
(Fig. 4C; P<0.05). A similar
trend was observed in the caspase activity assays. As shown in
Fig. 4D, HMGA2 overexpression
significantly decreased the levels of caspase-3 and caspase-9 in
miR-9-5p agomir-transfected HT-29 cells, while HMGA2-knockdown
significantly increased the levels of caspase-3 and caspase-9 in
miR-9-5p antagomir-transfected HCT-116 cells (P<0.05). The
current findings indicated that HMGA2 overexpression reversed
miR-9-5p-induced apoptosis of HT-29 cells.
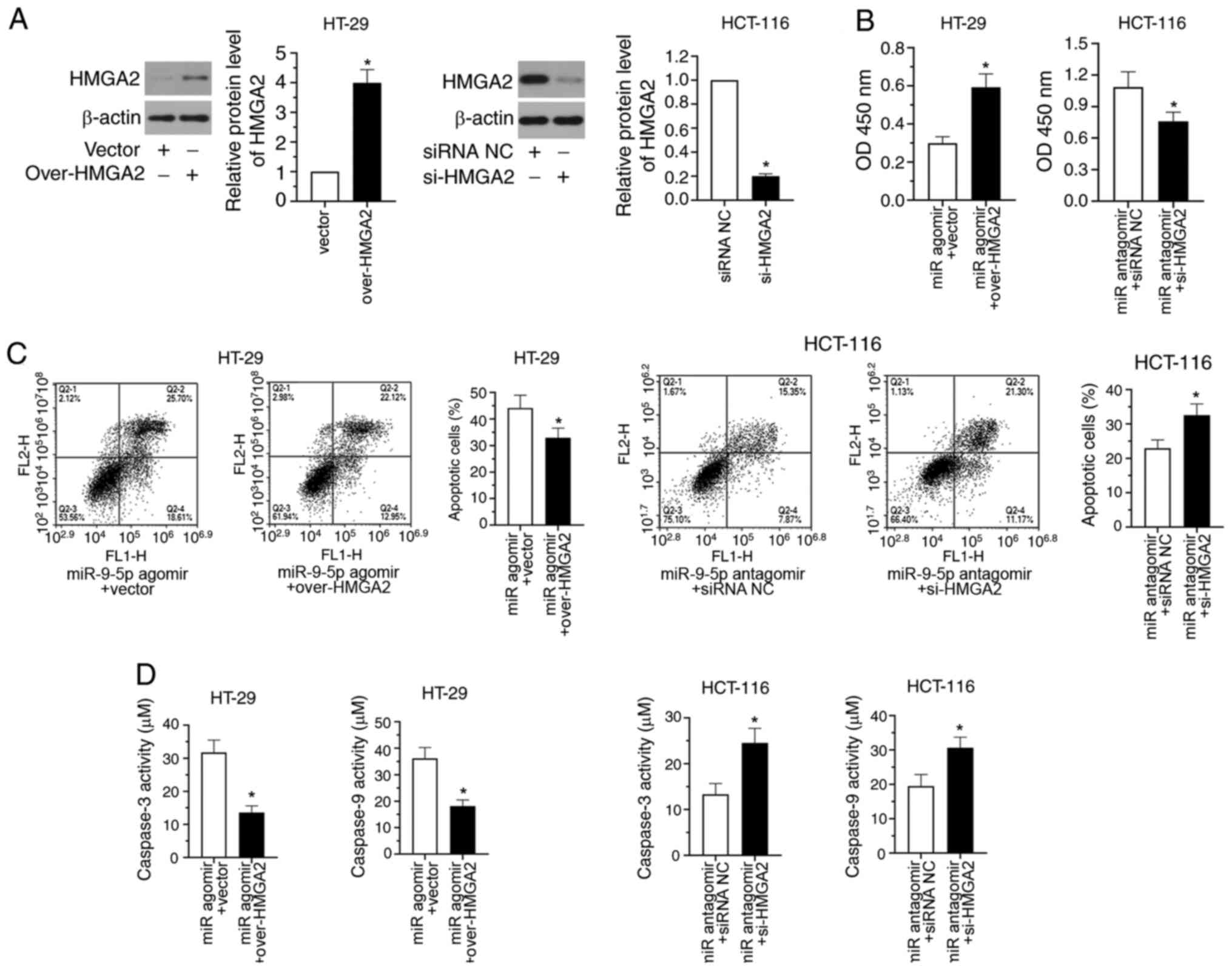 | Figure 4.HMGA2 overexpression reverses
miR-9-5p-induced HT-29 apoptosis. (A) HT-29 cells were transfected
with the HMGA2-overexpression plasmid, while HCT-116 cells were
transfected with HMGA2 siRNA. After 48 h, the protein levels of
HMGA2 were detected by western blotting. Subsequently,
HMGA2-overexpression plasmid was co-transfected with miR-9-5p
agomir into HT-29 cells, while HMGA2 siRNA was co-transfected with
miR-9-5p antagomir into HCT-116 cells. Transfected cells were
subjected to 2.5 or 10 µM 5-FU. After 48 h of transfection, (B)
cell viability was analysed using the Cell Counting Kit-8 assay,
(C) apoptosis was examined by flow cytometry and (D) caspase-3 and
caspase-9 activities were determined. *P<0.05. Data are shown as
the mean ± SD (n=3). OD, optical density; 5-FU, 5-fluorouracil;
miR, microRNA; NC, negative control; HMGA2, high mobility group A2;
siRNA, small interfering RNA; over, overexpression. |
Discussion
Various types of treatment plans have been developed
for a more effective treatment of CRC; 5-FU is one of the most
important anticancer drugs for the therapy of CRC (2). However, resistance to anticancer drugs
frequently drives tumor progression (2,3).
Accumulating studies have demonstrated that miRNAs serve vital
roles in drug resistance for cancer chemotherapy based on their
functions and targets (24,25). A previous study revealed that
upregulation of miR-9-5p increased the sensitivity of
hepatocellular carcinoma cells to cisplatin via modulating
epithelial-mesenchymal transition by eukaryotic translation
initiation factor 5A2 signaling (9).
However, whether miR-9-5p is involved in 5-FU resistance of CRC is
unknown. A previous study suggested that miR-9-5p expression was
lower in CRC cells, including HCT-116 and HT-29 cells, compared
with that in normal colorectal epithelial cells (13). In the current study, the functional
effect of miR-9-5p in CRC chemoresistance was further investigated
and the targeted signaling that mediated CRC escaping from drug
toxicity was explored. The present study revealed that the cell
viability of HT-29 cells (p53 wild-type) and HCT-116 cells (TP53
allele homozygous mutant) (26) was
decreased in a concentration-dependent manner after 5-FU treatment.
HCT-116 cells seemed more sensitive to 5-FU than HT-29 cells. van
Boxtel et al (27) have
reported that homozygous mutant rats lacking TP53 exhibit decreased
survival due to spontaneous tumor development compared with
heterozygous rats, which can explain the aforementioned finding.
Additionally, RT-qPCR was performed to examine miR-9-5p expression,
and the results suggested that 5-FU significantly upregulated
miR-9-5p expression in CRC cells. Notably, overexpression of
miR-9-5p suppressed cell viability and induced apoptosis in
5-FU-treated HT-29 cells, whereas these responses were mitigated by
miR-9-5p-knockdown. Application of miR-9-5p agomir alleviated 5-FU
chemoresistance in HT-29 cells. The present study confirmed the
crucial role of miR-9-5p in 5-FU chemoresistance.
Numerous studies have demonstrated that miR-9 acts
as a tumor suppressor by binding to various targets in CRC, such as
C-X-C motif chemokine receptor 4 (12), ubiquitin-like with plant homeodomain
and ring finger domains 1 (28) and
P21-activated kinase 4 (29).
However, the association between miR-9-5p and its target gene in
CRC cells to 5-FU resistance has not been defined. The chromatin
structuring protein HMGA2 has been found to affect the
chemosensitivity of CRC to irinotecan and the stability of human
subtelomere (30). A previous study
revealed that HMGA2 promotes gemcitabine resistance in pancreatic
cancer cells in vitro by increasing histone acetylation
(31). Additionally, high HMGA2
expression protects CRC cells from DNA breaks caused by the drug
irinotecan, which is a clinically important inhibitor of DNA
topoisomerases I (TOP-I) (30).
During the shift of replication forks, the transient accumulation
of DNA supercoil disrupts gene stability and is manipulated by DNA
TOP (30). The protective role of
HMGA2 may be due to inhibition of the formation of TOP-I/DNA
complexes (32). On the other hand,
low HMGA2 expression contributes to the formation of TOP-I/DNA
complexes induced by irinotecan (30). Tsavaris et al (33) have confirmed that TOP-I is highly
expressed in CRC following 5-FU treatment. In the current study,
miR-9-5p overexpression significantly decreased HMGA2 expression by
directly targeting the 3′-UTR of HMGA2. Overexpression of HMGA2
restored miR-9-5p-repressed 5-FU chemoresistance in vitro.
The present findings indicated an association between miR-9-5p and
5-FU resistance in CRC. Further exploration is required on whether
TOP-I may be a miR-9-5p-inhibited 5-FU sensitivity determinant.
In summary, the present study confirmed that the
inhibition of miR-9-5p in 5-FU resistance was mediated by
downregulating HMGA2 expression. miR-9-5p enhanced the CRC cell
sensitivity to 5-FU and induced apoptosis. The current data
provides evidence supporting the association of miR-9-5p with 5-FU
resistance in CRC.
Acknowledgements
Not applicable.
Funding
The present study was supported by a grant from the
Fundamental Research Business Expense of Universities in
Heilongjiang Province (grant no. 2018-KYYWFMY-0056).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HZ and BY designed the study. HZ and QW performed
the research and analysed the data. HZ wrote the first draft of the
manuscript. JZ performed the supplementary experiments and revised
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Pisano A, Griñan-Lison C, Farace C,
Fiorito G, Fenu G, Jiménez G, Scognamillo F, Peña-Martin J,
Naccarati A, Pröll J, et al: The inhibitory role of miR-486-5p on
CSC phenotype has diagnostic and prognostic potential in colorectal
cancer. Cancers (Basel). 12:34322020. View Article : Google Scholar
|
|
2
|
Giuliani J and Bonetti A: The
pharmacological costs of first-line therapies in unselected
patients with advanced colorectal cancer: A review of published
phase III trials. Clin Colorectal Cancer. 15:277–284. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yao J, Huang A, Zheng X, Liu T, Lin Z,
Zhang S, Yang Q, Zhang T and Ma H: 53BP1 loss induces
chemoresistance of colorectal cancer cells to 5-fluorouracil by
inhibiting the ATM-CHK2-P53 pathway. J Cancer Res Clin Oncol.
143:419–431. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wu C, Bardes EE, Jegga AG and Aronow BJ:
ToppMiR: Ranking microRNAs and their mRNA targets based on
biological functions and context. Nucleic Acids Res. 42((Web Server
Issue)): W107–W113. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Berindan-Neagoe I, Monroig Pdel C,
Pasculli B and Calin GA: MicroRNAome genome: A treasure for cancer
diagnosis and therapy. CA Cancer J Clin. 64:311–336. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhao L, Shan Y, Liu B, Li Y and Jia L:
Retraction Note: Functional screen analysis reveals miR-3142 as
central regulator in chemoresistance and proliferation through
activation of the PTEN-AKT pathway in CML. Cell Death Dis.
11:1212020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Si W, Shen J, Du C, Chen D, Gu X, Li C,
Yao M, Pan J, Cheng J, Jiang D, et al: A miR-20a/MAPK1/c-Myc
regulatory feedback loop regulates breast carcinogenesis and
chemoresistance. Cell Death Differ. 25:406–420. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang B, Lu FY, Shi RH, Feng YD, Zhao XD,
Lu ZP, Xiao L, Zhou GQ, Qiu JM and Cheng CE: miR-26b regulates
5-FU-resistance in human colorectal cancer via down-regulation of
Pgp. Am J Cancer Res. 8:2518–2527. 2018.PubMed/NCBI
|
|
9
|
Bao Y, Zhang Y, Lu Y, Guo H, Dong Z, Chen
Q, Zhang X, Shen W, Chen W and Wang X: Overexpression of microRNA-9
enhances cisplatin sensitivity in hepatocellular carcinoma by
regulating EIF5A2-mediated epithelial-mesenchymal transition. Int J
Biol Sci. 16:827–837. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Meng Q, Xiang L, Fu J, Chu X, Wang C and
Yan B: Transcriptome profiling reveals miR-9-3p as a novel tumor
suppressor in gastric cancer. Oncotarget. 8:37321–37331. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhu B, Xi X, Liu Q, Cheng Y and Yang H:
miR-9 functions as a tumor suppressor in acute myeloid leukemia by
targeting CX chemokine receptor 4. Am J Transl Res. 11:3384–3397.
2019.PubMed/NCBI
|
|
12
|
Xiong WC, Han N, Ping GF, Zheng PF, Feng
HL, Qin L and He P: microRNA-9 functions as a tumor suppressor in
colorectal cancer by targeting CXCR4. Int J Clin Exp Pathol.
11:526–536. 2018.PubMed/NCBI
|
|
13
|
Cekaite L, Rantala JK, Bruun J, Guriby M,
Agesen TH, Danielsen SA, Lind GE, Nesbakken A, Kallioniemi O, Lothe
RA and Skotheim RI: miR-9, −31, and −182 deregulation promote
proliferation and tumor cell survival in colon cancer. Neoplasia.
14:868–879. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Park YR, Lee ST, Kim SL, Liu YC, Lee MR,
Shin JH, Seo SY, Kim SH, Kim IH, Lee SO and Kim SW: MicroRNA-9
suppresses cell migration and invasion through downregulation of
TM4SF1 in colorectal cancer. Int J Oncol. 48:2135–2143. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Borrmann L, Schwanbeck R, Heyduk T,
Seebeck B, Rogalla P, Bullerdiek J and Wiśniewski JR: High mobility
group A2 protein and its derivatives bind a specific region of the
promoter of DNA repair gene ERCC1 and modulate its activity.
Nucleic Acids Res. 31:6841–6851. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gao X, Dai M, Li Q, Wang Z, Lu Y and Song
Z: HMGA2 regulates lung cancer proliferation and metastasis. Thorac
Cancer. 8:501–510. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mansoori B, Mohammadi A, Asadzadeh Z,
Shirjang S, Minouei M, Abedi Gaballu F, Shajari N, Kazemi T,
Gjerstorff MF, Duijf PHG and Baradaran B: HMGA2 and Bach-1
cooperate to promote breast cancer cell malignancy. J Cell Physiol.
234:17714–17726. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang X, Liu X, Li AY, Chen L, Lai L, Lin
HH, Hu S, Yao L, Peng J, Loera S, et al: Overexpression of HMGA2
promotes metastasis and impacts survival of colorectal cancers.
Clin Cancer Res. 17:2570–2580. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li Y, Zhao Z, Xu C, Zhou Z, Zhu Z and You
T: HMGA2 induces transcription factor Slug expression to promote
epithelial-to-mesenchymal transition and contributes to colon
cancer progression. Cancer Lett. 355:130–140. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xu X, Wang Y, Deng H, Liu C, Wu J and Lai
M: HMGA2 enhances 5-fluorouracil chemoresistance in colorectal
cancer via the Dvl2/Wnt pathway. Oncotarget. 9:9963–9974. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang YN, Zeng ZL, Lu J, Wang Y, Liu ZX, He
MM, Zhao Q, Wang ZX, Li T, Lu YX, et al: CPT1A-mediated fatty acid
oxidation promotes colorectal cancer cell metastasis by inhibiting
anoikis. Oncogene. 37:6025–6040. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lesuffleur T, Violette S, Vasile-Pandrea
I, Dussaulx E, Barbat A, Muleris M and Zweibaum A: Resistance to
high concentrations of methotrexate and 5-fluorouracil of
differentiated HT-29 colon-cancer cells is restricted to cells of
enterocytic phenotype. Int J Cancer. 76:383–392. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Longley DB and Johnston PG: Molecular
mechanisms of drug resistance. J Pathol. 205:275–292. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xia H, Ooi LL and Hui KM:
MicroRNA-216a/217-induced epithelial-mesenchymal transition targets
PTEN and SMAD7 to promote drug resistance and recurrence of liver
cancer. Hepatology. 58:629–641. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ahmed D, Eide PW, Eilertsen IA, Danielsen
SA, Eknæs M, Hektoen M, Lind GE and Lothe RA: Epigenetic and
genetic features of 24 colon cancer cell lines. Oncogenesis.
2:e712013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
van Boxtel R, Kuiper RV, Toonen PW, van
Heesch S, Hermsen R, de Bruin A and Cuppen E: Homozygous and
heterozygous p53 knockout rats develop metastasizing sarcomas with
high frequency. Am J Pathol. 179:1616–1622. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhu M, Xu Y, Ge M, Gui Z and Yan F:
Regulation of UHRF1 by microRNA-9 modulates colorectal cancer cell
proliferation and apoptosis. Cancer Sci. 106:833–839. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang M, Gao Q, Chen Y, Li Z, Yue L and Cao
Y: PAK4, a target of miR-9-5p, promotes cell proliferation and
inhibits apoptosis in colorectal cancer. Cell Mol Biol Lett.
24:582019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ahmed SM, Ramani PD, Wong SQR, Zhao X,
Ivanyi-Nagy R, Leong TC, Chua C, Li Z, Hentze H, Tan IB, et al: The
chromatin structuring protein HMGA2 influences human subtelomere
stability and cancer chemosensitivity. PLoS One. 14:e02156962019.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dangi-Garimella S, Sahai V, Ebine K, Kumar
K and Munshi HG: Three-dimensional collagen I promotes gemcitabine
resistance in vitro in pancreatic cancer cells through
HMGA2-dependent histone acetyltransferase expression. PLoS One.
8:e645662013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhao X, Peter S, Dröge P and Yan J:
Oncofetal HMGA2 effectively curbs unconstrained (+) and (−) DNA
supercoiling. Sci Rep. 7:84402017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tsavaris N, Lazaris A, Kosmas C, Gouveris
P, Kavantzas N, Kopterides P, Papathomas T, Agrogiannis G, Zorzos
H, Kyriakou V and Patsouris E: Topoisomerase I and IIalpha protein
expression in primary colorectal cancer and recurrences following
5-fluorouracil-based adjuvant chemotherapy. Cancer Chemother
Pharmacol. 64:391–398. 2009. View Article : Google Scholar : PubMed/NCBI
|