Introduction
Lung cancer is the most common malignancy worldwide
and one-quarter of all cancer-associated deaths are due to lung
cancer globally (1). As the
predominant subtype of lung cancer, non-small cell lung cancer
(NSCLC) is furtherly divided into adenocarcinoma and squamous cell
carcinoma, of which lung adenocarcinoma (LUAD) is the most common
type and comprises ~40% of all lung cancer globally in 2020
(2). Although treatments for LUAD
are plentiful, including surgery, radiotherapy, chemotherapy,
targeted therapy and immunotherapy, the prognosis of patients with
LUAD remains poor, with a 5-year overall survival rate <20%
globally in 2020 (3). Thus,
exploring novel genes functioning in the development of LUAD is
important and may provide potential targets for the diagnosis and
treatment of LUAD.
The Gene Expression Profiling Interactive Analysis
(GEPIA) website (http://gepia.cancer-pku.cn/) is an online tool for
visually analyzing gene expression in all types of cancer. The
database was explored to seek for potential oncogenes in LUAD.
FAM83A belongs to the FAM83 family of proteins, which are
characterized by a highly conserved domain of unknown function
(4). It has been demonstrated that
FAM83 proteins exhibit oncogenic properties and have significantly
elevated levels of expression in multiple human tumor types,
including breast, lung, bladder, testis and ovarian cancer
(5). Further research showed that
ablation of numerous FAM83 proteins resulted in a marked
suppression of cancer-associated signaling and loss of tumorigenic
potential (6). Therefore, it is
important to study the function and mechanism of FAM83A in
LUAD.
Long non-coding (lnc)RNAs are a type of transcript
that are >200 nucleotides in length and with limited or no
protein-coding capacity (7,8). It has been reported that lncRNAs exert
vital roles in cancer, including cell cycle procession, apoptosis,
drug resistance and stemness maintenance (9,10).
Gene-editing tools to target lncRNA have already shown the
feasibility of gene editing in treating cancer (11). FAM83A-AS1 is the anti-sense lncRNA of
FAM83A, and the association of FAM83A-AS1 with the development of
LUAD was analyzed by GEPIA.
The aim of the present study was to explore the
roles of FAM83A and FAM83A-AS1 in LUAD. Public databases were
analyzed and experiments were performed to uncover the expression
and function of FAM83A and FAM83A-AS1 in LUAD. The oncogenic role
of FAM83A and FAM83A-AS1 might provide diagnostic markers and
therapeutic targets for patients with LUAD.
Materials and methods
Bioinformatics
The GEPIA website (http://gepia.cancer-pku.cn/) was used to screen for
potential oncogenes in LUAD and to analyze the correlation between
gene expression and clinical characteristics. The differentially
expressed genes in LUAD were obtained by searching the
‘Differential Expression Analysis’ section of GEPIA website
(http://gepia2.cancer-pku.cn/#degenes)
with the following terms: Cancer name, LUAD; |log2 fold-change
(FC)|≥1; q-value ≤0.01 and differential methods, ANOVA. The top 500
differential survival genes in LUAD were obtained by searching the
‘Most Differential Survival Genes’ section of GEPIA website
(http://gepia2.cancer-pku.cn/#survival) with the
following terms: Cancer name, LUAD and group cut-off, median. The
Kaplan-Meier curves were downloaded from the ‘Survival Analysis’
section of GEPIA website (http://gepia2.cancer-pku.cn/#survival) with the
following terms: Gene, FAM83A or FAM83A-AS1; group cut-off, median
and cancer name, LUAD. The staging boxplots were downloaded from
the ‘Expression DIY-Stage Plot’ section of GEPIA website
(http://gepia2.cancer-pku.cn/#analysis) with the
following terms: Gene, FAM83A or FAM83A-AS1 and cancer name, LUAD.
The median expression (transcripts per million) of genes in normal
and LUAD tissue was obtained from ‘Differential Expression
Analysis’ section of GEPIA website (http://gepia2.cancer-pku.cn/#degenes). The
hierarchical clustering map in Fig.
1B was drafted with these data using GraphPad Prism version 6
(GraphPad Software). The Kaplan-Meier curves were downloaded from
the ‘Survival Analysis’ section of GEPIA website (http://gepia2.cancer-pku.cn/#survival)
with the following terms: Gene, FAM83A or FAM83A-AS1; group
cut-off, median and cancer name, LUAD. The violin plots in Fig. 2B and E were downloaded from the
‘Expression DIY-Stage Plot’ section of GEPIA website (http://gepia2.cancer-pku.cn/#analysis)
with the following terms: Gene, FAM83A or FAM83A-AS1 and cancer
name, LUAD. The boxplots in Figs. 3A
and 4A were downloaded from the CCLE
dataset (https://portals.broadinstitute.org/ccle) by searching
for FAM83A or FAM83A-AS1. The Ensembl genome browser (http://asia.ensembl.org/index.html) was scanned
to observe the genomic location of FAM83A and FAM83A-AS1. Gene
clusters highly correlated with FAM83A and FAM83A-AS1 (Pearson
score >0.3) were submitted to the database for Annotation,
Visualization and Integrated Discovery Bioinformatics resources
version 6.8 (http://david.abcc.ncifcrf.gov/) (12) for Gene Ontology (GO) and Kyoto
Encyclopedia of Genes and Genomes (KEGG) analysis, and GO terms and
KEGG pathways with a gene count ≥3 were included in subsequent
analysis. The mRNA expression data of FAM83A and FAM83A-AS1 in
different types of cancer cells was downloaded from the CCLE
dataset (https://portals.broadinstitute.org/ccle). The
histograms in Figs. 3B and 4B were constructed using these data and
GraphPad. The secondary structure of FAM83A-AS1 was downloaded from
the HUGO Gene Nomenclature Committee (http://www.genenames.org/) (13). The cBioPortal website (http://www.cbioportal.org/) (14) was utilized to analyze the correlation
between FAM83A and FAM83A-AS1. LncBase version 2 (http://carolina.imis.athena-innovation.gr/diana_tools/web/index.php)
(15) and StarBase version 2.0
(http://starbase.sysu.edu.cn/starbase2/browseNcRNA.php)
(16) were browsed to identify the
miRNAs that bind FAM83A and FAM83A-AS1.
Cell culture and transfection
NCIH1650 and A549 cells was purchased from the
Shanghai Institutes for Biological Science and were cultured in
RPMI1640 (Nanjing KeyGen Biotech Co., Ltd.) medium containing 10%
fetal bovine serum (Invitrogen; Thermo Fisher Scientific, Inc.) in
a humidified incubator containing 5% CO2 at 37°C. LUAD
cells were seeded in 6-well plates with a density of
5×105 cells per well. Two small interfering (si)RNAs
targeting FAM83A/FAM83A-AS1, miR-495-3p mimics and miR-495-3p
inhibitors were constructed and purchased from Guangzhou RiboBio
Co., Ltd, and si-NC or miR-NC (non-targeting) was used as a
control. Transfection of siRNA and miRNA mimics was performed in
room temperature for 6 h according to the Lipofectamine®
3000 reagent (Invitrogen; Thermo Fisher Scientific, Inc.) protocol.
For 6-well plates, 5 µl siRNAs (20 nM) or miRNAs mimics (20 nM)
were transfected in each well. Subsequent experiments were
performed after 48 h of transfection. The rescue experiments were
performed by transfecting siRNA of FAM83A/FAM83A-AS1 and miR-495-3p
inhibitor. The sequence of siRNAs and miRNA mimics were as follows:
si-NC, forward: 5′-UUCUCCGAACGUGUCACGUTT-3′and reverse:
5′-ACGUGACACGUUCGGAGAATT-3′; si1-FAM83A, forward:
5′-UUCUCCGAACGUGUCACGUTT-3′ and reverse:
5′-ACGUGACACGUUCGGAGAATT-3′; si2-FAM83A, forward:
5′-UGAACUUCUCCCGGAUUUGTT-3′ and reverse:
5′-CAAAUCCGGGAGAAGUUCATT-3′; si1-FAM83A-AS1, forward:
5′-GCUGCCACCUACAAGAUAATT-3′ and reverse:
5′-UUAUCUUGUAGGUGGCAGCTT-3′; si2-FAM83A-AS1, forward:
5′-GGCCCUGGGCUGAAUAAUUTT-3′ and reverse:
5′-AAUUAUUCAGCCCAGGGCCTT-3′; miR-NC, 5′-UUCUCCGAACGUUCACGUTT-3′;
miR-495-3p mimics, 5′-AGGAUGUCUAAAUGUUUGUUA-3′ and miR-495-3p
inhibitor, 5′-AAGAAGUGCACCAUGUUUGUUU-3′.
RNA extraction and reverse
transcription-quantitative (RT-q)PCR
TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) was used to extract the total RNAs from
cells transfected as aforementioned. cDNAs were obtained by reverse
transcription using a Reverse Transcription kit (cat. no. RR036A;
Takara Bio, Inc.) at 37°C for 15 min and 85°C for 5 min. RT-qPCR
was conducted with SYBR Select Master mix (cat. no. 4472908;
Applied Biosystems; Thermo Fisher Scientific, Inc.) and the
relative expression was calculated using the 2−ΔΔCq
method (17), with ACTB as a
reference gene. The thermocycling conditions for RT-qPCR were as
follows: Initial denaturation, 95°C for 30 sec; denaturation, 95°C
for 5 sec, 60°C for 35 sec, for a total of 40 cycles; annealing and
extension, 95°C for 15 sec, 60°C for 1 min and 95°C for 15 sec. The
primers used are shown in Table
SI.
Western blotting
For protein extraction, cells transfected as
aforementioned were lysed in RIPA lysis buffer with freshly added
protease inhibitor PMSF (1:100; Nanjing KeyGen Biotech Co., Ltd.).
Protein concentration was determined using a BCA kit (Nanjing
KeyGen Biotech Co., Ltd.) and 30 µg of lysate protein was loaded
per lane for western blotting. Briefly, protein extracts were
loaded on 10% SDS-PAGE gels for electrophoresis and then were
electrotransferred to a PVDF membrane. The membrane was blocked at
room temperature with non-fat milk for 2 h and incubated overnight
with respective primary antibodies at 4°C. The following primary
antibodies were used: Anti-β-actin (1:1,000; cat. no. 3700; Cell
Signaling Technology, Inc.) and anti-FAM83A (1:1,000; cat. no.
ab214014; Abcam). Then secondary antibodies (goat anti-rabbit
antibody, 1:10,000; cat. no. ab6721; Abcam, or goat anti-mouse
antibody, 1:10,000; cat. no. ab6789; Abcam) were used to incubate
the membrane for 2 h. Blots were visualized using ECL detection
with an ECL Substrate kit (cat. no. ab133406; Abcam) and ImageJ
(version 1.48; National Institutes of Health) software was used to
quantify protein expression.
EdU assay
An EdU Apollo® 488 In Vitro
Imaging kit (Guangzhou RiboBio Co., Ltd.) was used to detect
proliferation rate of NCIH1650 and A549 cells. After transfection
of 24 h, 8,000 cells/100 µl were plated in 96-well plates overnight
and then incubated with 100 µl 50 µM EdU solution for 2 h. The
cells were then fixed with 50 µl 4% paraformaldehyde at room
temperature for 10 min and neutralized using 50 µl 2 mg/ml glycine.
After permeabilization with 0.5% Triton X-100 for 20 min, the cells
were stained for DAPI with 100 µl 1× Apollo solution and stained
for Edu with 100 µl 1× Hoechst33342 solution, both at room
temperature for 30 min. Images were captured using fluorescence
microscopy with 40 times magnification and proliferation cells
ratios were counted from three random fields of view.
Cell Counting Kit (CCK)-8 assay
The CCK-8 assay was performed to evaluate the
relative number of NCIH1650 and A549 cells. After transfection of
24 h, 2,000 cells/100 µl were plated in 96-well plates and total of
20 µl CCK-8 reagent (Guangzhou RiboBio Co., Ltd.) was added to each
test well and cells were then incubated for 2 h at 37°C. The
absorbance at 450 nm was measured every 24 h.
Migration and invasion assay
Transwell assay inserts (8 µM PET, 24-well
Millicell) and Matrigel-coated membranes (BD Biosciences) were used
to detect the migration and invasion of cells, respectively. In
total, 200 µl serum-free medium was added to the upper chamber and
800 µl 10% DMEM serum-containing medium to the lower chamber.
Matrigel pre-coating was performed at 37°C for 30 min. The NCIH1650
and A549 cells were harvested 24 h after transfection and added to
the upper chamber (50,000 cells). After incubation at 37 C for 24 h
(migration assay) or 48 h (invasion assay), the migrated or invaded
cells were fixed with 4% polyformaldehyde and stained with crystal
violet, both at room temperature for 30 min. Images were captured
with light microscopy and cells were counted by ImageJ
software.
Luciferase reporter assay
A Dual Luciferase Reporter Assay system (Promega
Corporation) was used for luciferase assays. The binding sites of
FAM83A-AS1 with FAM83A 3′UTR were identified using StarBase as
aforementioned. A fragment of the FAM83A-AS1 or FAM83A 3′UTR
sequence containing a putative target site or mutated target site
for miR-495-3p was amplified and cloned into the pGL3 reporter
vector (Guangzhou RiboBio Co., Ltd.). The luciferase reporter
plasmids were co-transfected miR-495-3p mimics or miR control
(Guangzhou RiboBio Co., Ltd.) as aforementioned. After transfection
for 48 h, Renilla and Firefly luciferase activities were detected
in the dual luciferase reporter gene kit (Promega Corporation).
Firefly/Renilla value was used to measure relative luciferase
activity and results were normalized to the activity of the
control. The sequences of miRNA mimic/inhibitor were as follows:
miR-NC, 5′-UUCUCCGAACGUUCACGUTT-3′; miR-495-3p mimics,
5′-AGGAUGUCUAAAUGUUUGUUA-3′ and miR-495-3p inhibitor,
5′-AAGAAGUGCACCAUGUUUGUUU-3′
Statistical analysis
Generally experiments were repeated three times and
data are presented as the mean ± standard deviation (unless
otherwise shown). Unpaired student's t-tests were used to determine
statistical significance between two groups. ANOVA test and
Dunnett's post hoc were used for the comparison of multiple groups.
Spearman's test was used for the correlation analysis of two genes.
Graphs were made using the GraphPad Prism 6.0 software package.
Statistical analysis was performed using SPSS version 20.0 (IBM
Corp) and P<0.05 was considered to indicate a statistically
significant difference.
Results
FAM83A and FAM83A-AS1 are potential
oncogenes according to GEPIA analysis
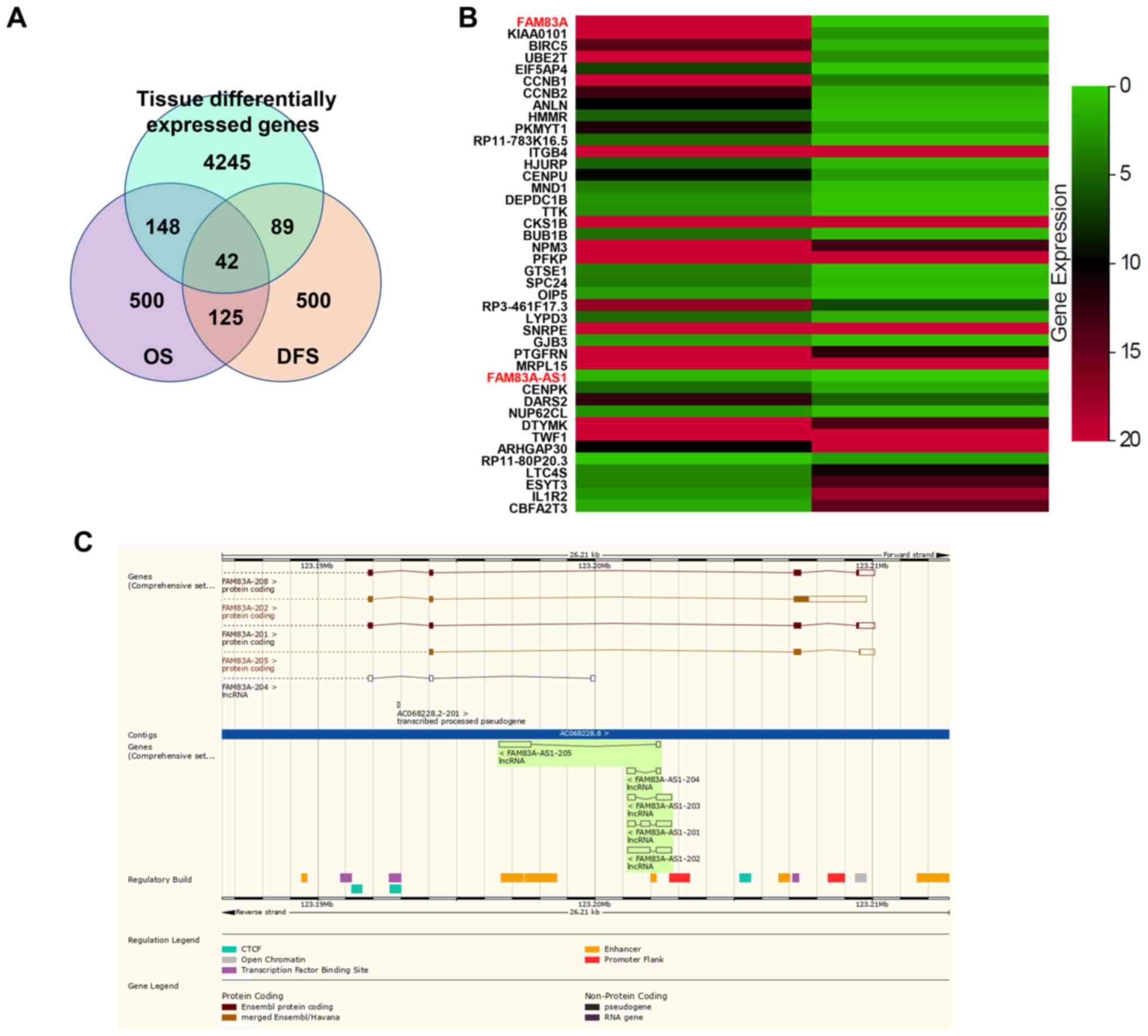
To screen for genes abnormally expressed and
associated with survival of patients with LUAD, the following
filter criterion was used: Cancer name, LUAD; gene expression,
|log2 FC|≥1, q-value ≤0.01 (tumor vs. normal); differential
survival genes with overall survival (OS) rates (top 500) and
disease-free survival (top 500) rates. Using this method, 42 genes
were obtained (Fig. 1A). To sort
these genes by differences in expression, it was demonstrated that
FAM83A was the most differentially expressed gene and its antisense
lncRNA, FAM83A-AS1, was also among the list (Fig. 1B). As shown in Ensembl genome
browser, FAM83A-AS1 is the antisense of FAM83A and both have
several different transcripts (Fig.
1C).
FAM83A and FAM83A-AS1 are correlated
with clinical characteristics of LUAD
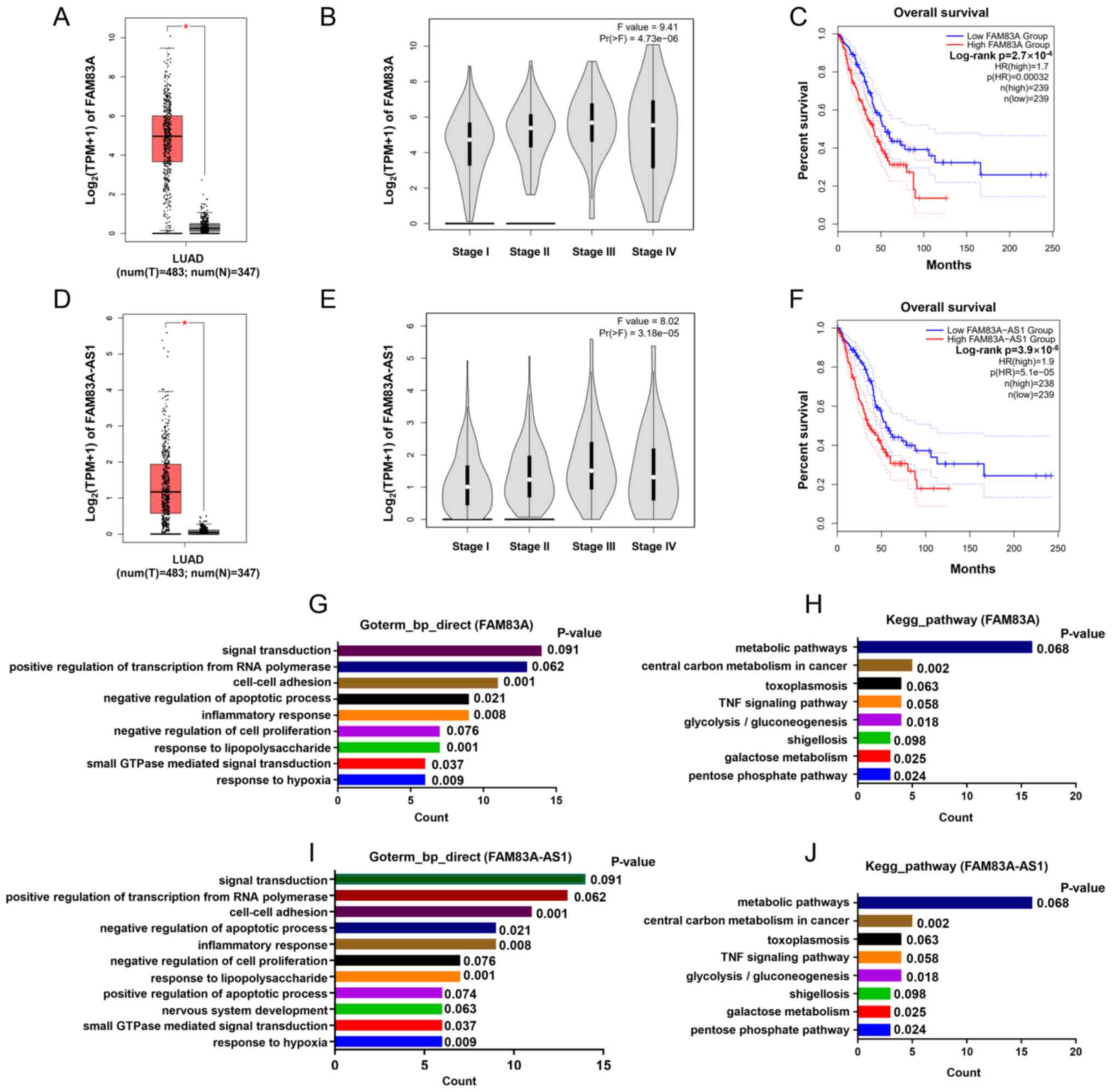
Further analysis of GEPIA indicated that FAM83A was
significantly upregulated in LUAD (tumor, n=483 vs. normal, n=347)
(Fig. 2A) and its expression was
relatively higher in patients with LUAD with more advanced stages
(Fig. 2B). Survival analysis showed
that patients of high FAM83A (n=239) had a worse OS (P<0.001)
compared paired with patients with low FAM83A expression (n=239)
(Fig. 2C). Analogously, FAM83A-AS1
was also overexpressed in LUAD and associated with tumor stage and
survival of patients (Fig. 2D-F).
Moreover, to predict the potential biological function of FAM83A
and FAM83A-AS1, genes highly co-expressed with these were submitted
for GO and KEGG pathway analysis, respectively. Results suggested
that FAM83A and FAM83A-AS1 bear strong similarities in biological
processes, mainly including ‘cell-cell adhesion’ and ‘central
carbon metabolism in cancer’ (Fig.
2G-J). FAM83A and FAM83A-AS1 both associated with clinical
characteristics of LUAD and it is important to deeply study their
roles in LUAD.
Knockdown of FAM83A inhibits
proliferation, migration and invasion of LUAD cells
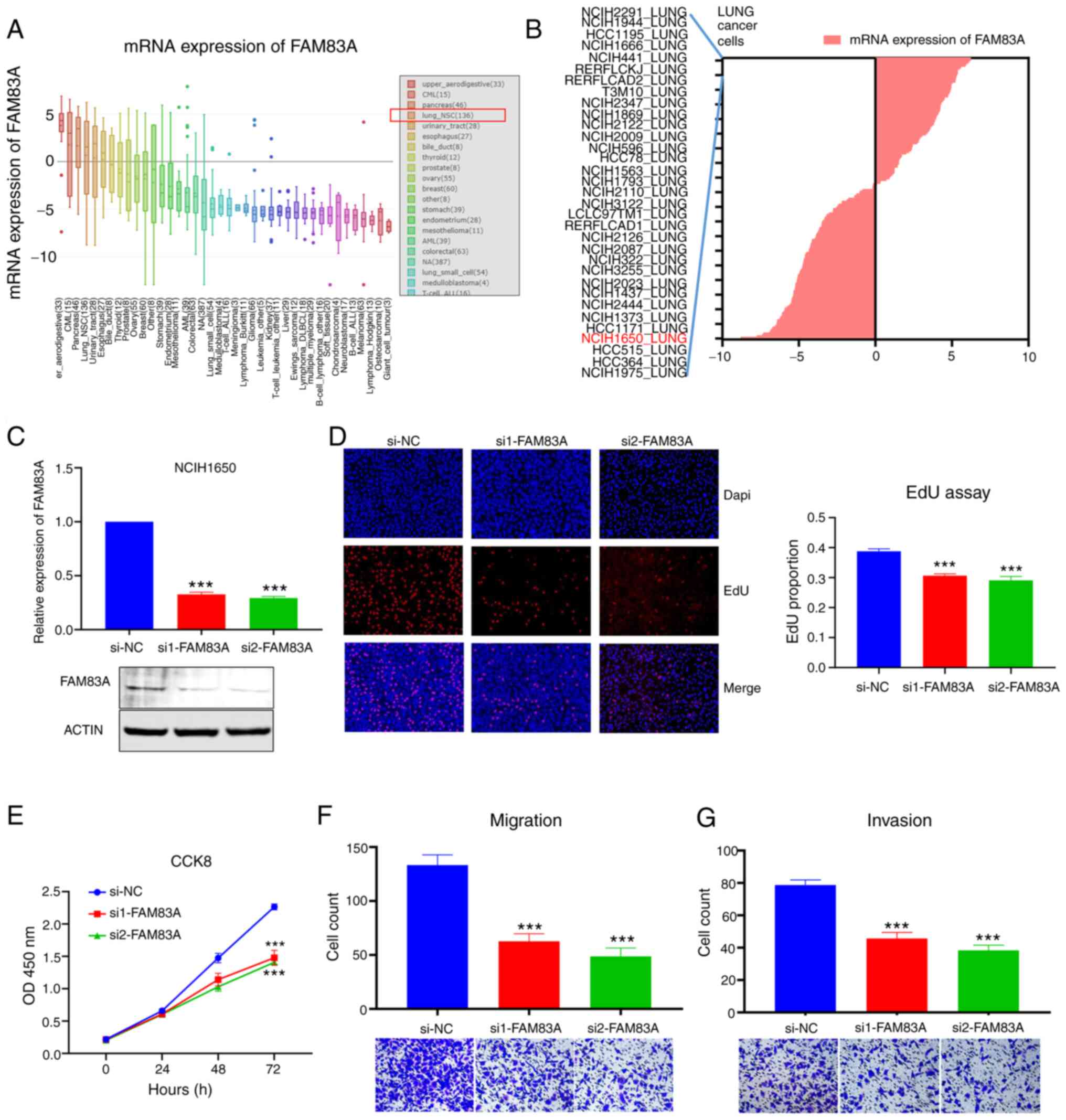
The CCLE dataset was searched to evaluate the
expression of FAM83A in different cancer cells and the results
indicated that the expression of FAM83A in NSCLC (lung_NSC) and
multiple other types of cancer cell was elevated (Fig. 3A). In the list of lung_NSC cell lines
with high expression of FAM83A, the common LUAD cell line,
NCIH1650, has been used for research in a previous study (18) and was selected for subsequent
experiments (Fig. 3B). To explore
the biological role of FAM83A, two siRNAs targeting FAM83A were
transfected into NCIH1650 and both siRNAs effectively silenced the
expression of FAM83A, confirmed by RTq-PCR and western blotting
(Fig. 3C). CCK-8 and EdU assays
validated that knockdown of FAM83A inhibited proliferation of
NCIH1650 cells (Fig. 3D and E).
Transwell and Matrigel assays revealed that silencing FAM83A
suppressed the ability of NCIH1650 cells to migrate and invade
(Fig. 3F and G). Additionally,
knockdown of FAM83A inhibited proliferation, migration and invasion
of A549 cells (Fig. S1). These
results suggested that FAM83A played a vital role in the
carcinogenesis of LUAD.
Knockdown of FAM83A-AS1 inhibits the
malignant phenotype of LUAD cells
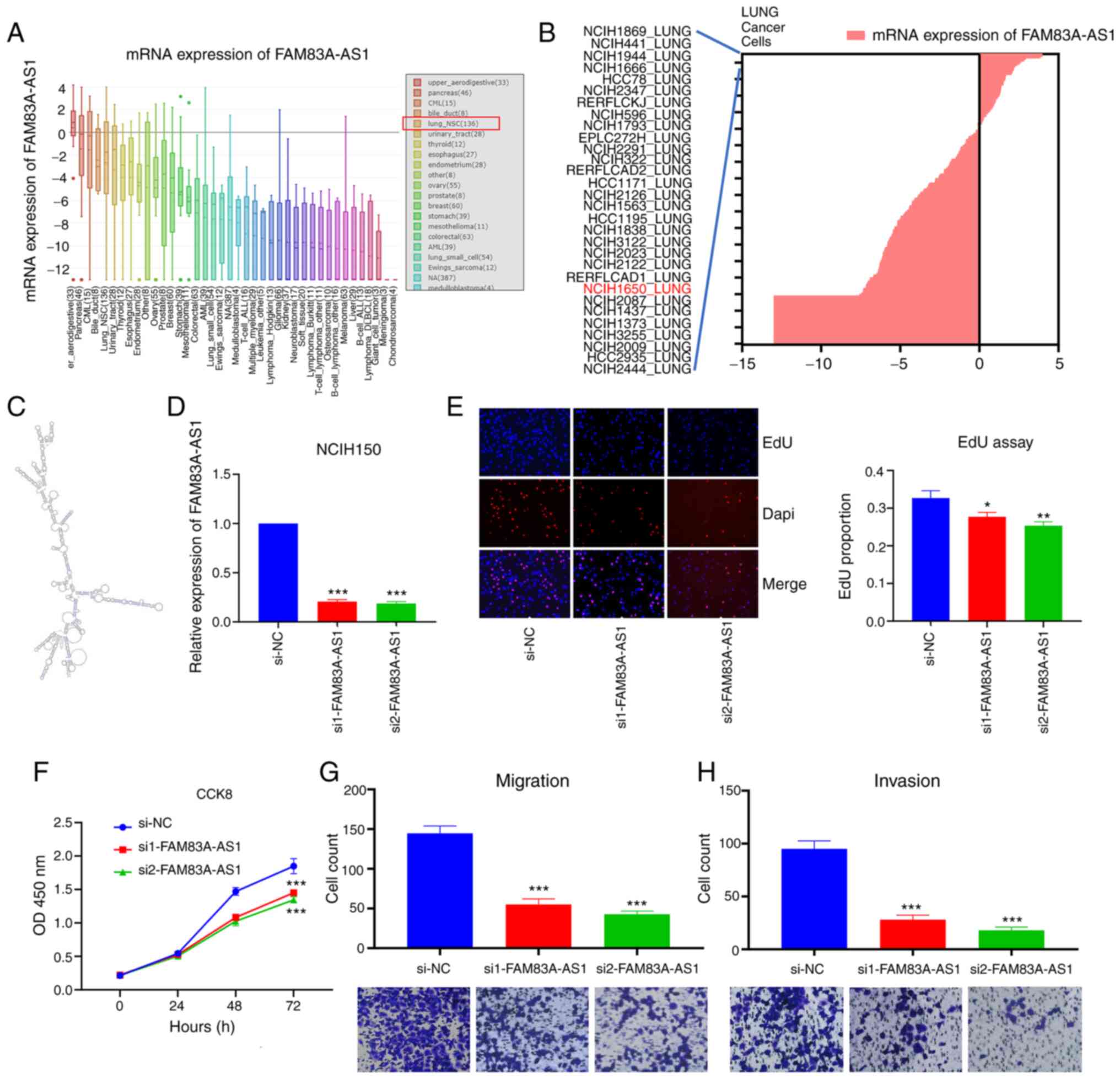
The average expression of FAM83A-AS1 in lung_NSC
cells was high, ranking 5th in all types of cancer (Fig. 4A). The expression of FAM83A-AS1 in
NCIH1650 was also elevated compared with other common LUAD cell
lines (Fig. 4B). FAM83A-AS1 is an
antisense transcript and with limited ability to encode proteins
(19,20). Two siRNAs were transfected to knock
down the expression of FAM83A-AS1 in NCIH1650 cell and to further
study its function (Fig. 4D).
Consistently, silencing FAM83A-AS1 inhibited the proliferation,
migration and invasion of NCIH1650 cells (Fig. 4E-H). Moreover, knockdown of
FAM83A-AS1 also suppressed the proliferation, migration and
invasion of A549 cells (Fig. S2).
Based on these results, it was concluded that both FAM83A and
FAM83A-AS1 played oncogenic roles in LUAD.
FAM83A-AS1 functions as a competing
endogenous (ce)RNA to regulate FAM83A by sponging miR-495-3p
Previous studies have shown that numerous antisense
lncRNAs exert biological roles via regulating its natural antisense
transcript (21–23). Thus, it was hypothesized that
FAM83A-AS1 might promote the progression of LUAD by influencing the
expression of FAM83A. Using the cBioPortal website, it was reported
that the expression of FAM83A-AS1 positively correlated with FAM83A
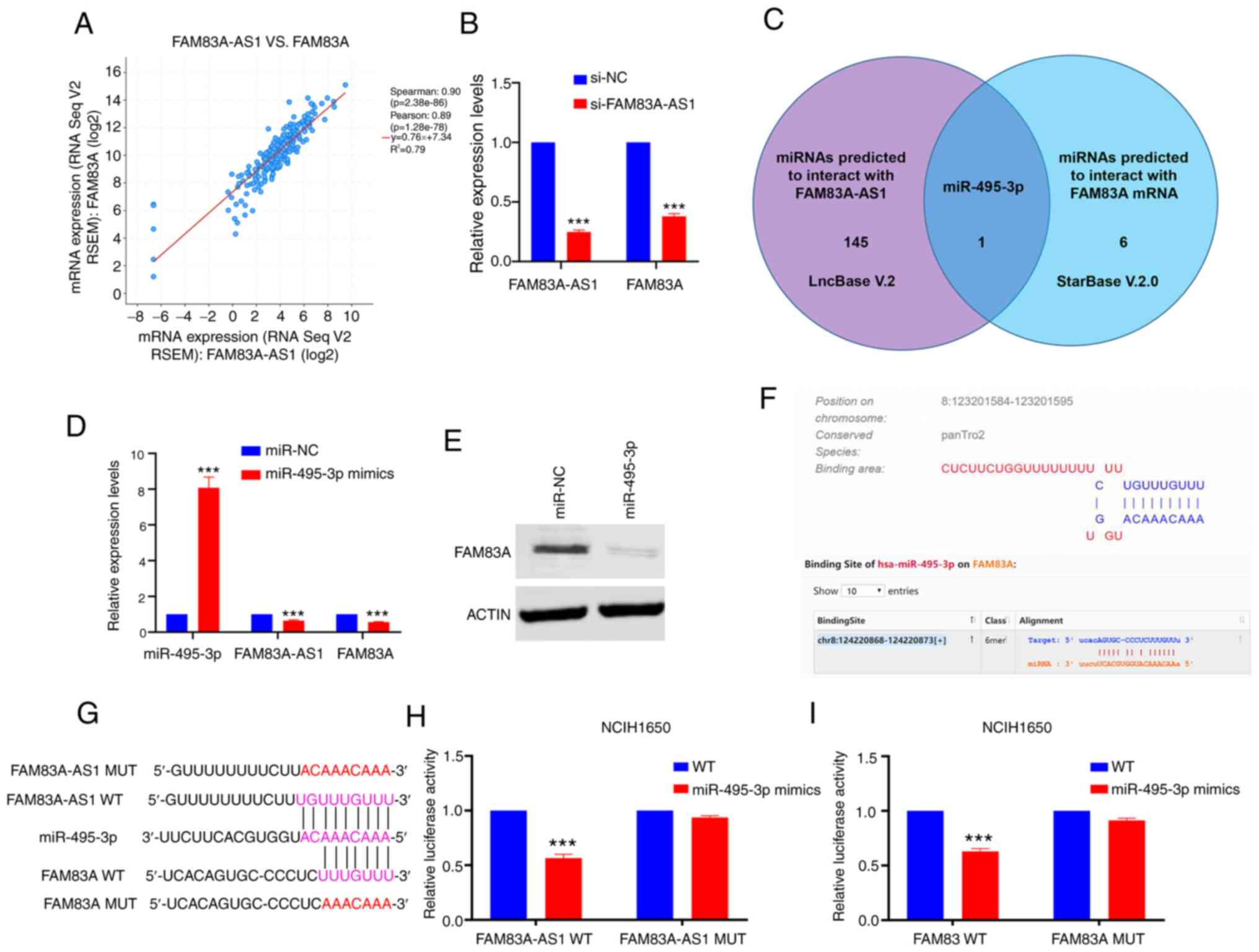
(Spearman score=0.90; Fig. 5A).
After knockdown of FAM83A-AS1, the expression of FAM83A was
decreased (Fig. 5B). Accumulating
evidence has highlighted the important roles of lncRNAs acting as
ceRNAs in cancer development (24–26), and
FAM83A-AS1 might regulate FAM83A via sponging specific miRNA. Using
LncBase and StarBase, miR-495-3p was predicted to interact with
both FAM83A-AS1 and FAM83A (Fig.
5C). Consistent with the assumption that FAM83A-AS1 might
regulate FAM83A via sponging specific miRNA, treatment of
miR-495-3p mimics significantly decreased mRNA levels of FAM83A-AS1
and FAM83A in NCIH1650 cells (P<0.001; Fig. 5D and E). To clarify their potential
interactions, luciferase vectors (wild-type and mutant type) were
constructed according to the putative binding sites (Fig. 5F and G). The luciferase reporter
assay showed that ectopic expression of miR-495-3p decreased the
luciferase activity of FAM83A/FAM83A-AS1 wild-type reporters, but
not mutant type (Fig. 5H and I).
Furthermore, the rescue experiment showed that miR-495-3p
counteracted the effect of FAM83A and FAM83A-AS1 in NCIH1650 cells.
The influence of FAM83A/FAM83A-AS1-knockdown on proliferation,
migration and invasion in LUAD cells could be restored by
miR-495-3p inhibitor (Fig. S3).
These results indicated that FAM83A-AS1 functioned as a ceRNA
through binding miR-495-3p, thereby decreasing the levels of FAM83A
mRNA transcripts.
Discussion
The poor prognosis of LUAD is largely due to that
most cases are diagnosed in advanced stages and lose the
opportunity of surgical treatment (27). Therefore, it is critical to identify
novel molecular drivers of LUAD and thus to provide diagnostic
markers and therapeutic targets for patients. The present study
identified that coding gene FAM83A and lncRNA FAM83A-AS1 were both
upregulated in LUAD tissues, and the overexpression of FAM83A and
FAM83A-AS1 was associated with more advanced clinical stages and
poorer prognosis of patients with LUAD. Further experiments
indicated that knockdown of FAM83A or FAM83A-AS1 could inhibit
proliferation, migration and invasion of LUAD. Moreover, it was
confirmed that FAM83A-AS1 regulated the expression of its natural
antisense transcript FAM83A by acting as a ceRNA through binding
miR-495-3p.
FAM83A belongs to FAM83 family of proteins, which
contain a conserved domain of unknown function, namely the DUF1669
domain (28). This family consists
of eight members (A-H), and a study demonstrated that some of the
FAM83 family proteins have oncogenic properties and have
significantly elevated expression levels in multiple human tumor
types (29), including cervical,
prostate, liver and ovarian cancer. For example, FAM83B promotes
endometrial cancer cell proliferation and invasion by inhibiting
autophagy via activating the PI3K/AKT/mTOR pathway (30). FAM83D is associated with sex,
American Joint Committee on Cancer stage, OS and DFS in
hepatocellular carcinoma (31).
FAM83H is involved in progression of osteosarcoma via stabilization
of β-catenin (32). As for FAM83A,
previous studies have indicated that FAM83A conferred EGFR-tyrosine
kinase inhibitor resistance in breast cancer cells by interacting
with and inducing phosphorylation of c-RAF and PI3K p85 (33). FAM83A expression is significantly
elevated in human and murine pancreatic cancer and is essential for
the proliferation and tumorigenesis of pancreatic cancer cells
(34). In conformity with these
studies, the present study identified the oncogenic role of FAM83A
in LUAD and deepened the cognizance of FAM83 family proteins in
cancer development.
lncRNAs are RNA molecules >200 nucleotides in
length with limited ability to encode proteins and participate in
the regulation of various biological processes, such as embryonic
development, tissue differentiation and cell proliferation
(35). As a type of lncRNA, natural
antisense transcripts (NATs) are non-coding RNA sequences that are
complementary to and overlap with either protein-coding or
non-coding transcripts (36). NATs
have diverse transcriptional and post-transcriptional regulatory
mechanisms with a wide variety of biological roles, such as
alternative splicing, chromatin remodeling and transcriptional
regulation (37). In particular,
NATs show potential for use as therapeutic targets because of their
highly locus-specific effects (38).
In the present study, FAM83A-AS1, the antisense transcript of
FAM83A, was overexpressed in LUAD and involved in the malignant
properties of LUAD cells. A previous review of studies identified
the tumor-promoting function of FAM83A-AS1 in lung tumorigenesis,
but the mechanism is still unclear (39,40). It
has been demonstrated that NATs are abundant non-coding RNAs that
regulate their corresponding sense mRNA through a variety of
molecular mechanisms (41). For
instance, receptor-type tyrosine-protein phosphatase ε (PTPRE)-AS1
binds to WD repeat-containing protein 5 and mediates H3 lysine 4
trimethylation of the PTPRE promoter region, thereby epigenetically
activating PTPRE gene expression (42). NCK1-AS1 competed with NCK1 mRNA for
miR-137 binding to boost NCK1 mRNA expression and thus to
facilitate tumorigenesis and chemo-resistance in ovarian cancer
(43). Metastasis-associated in
colon cancer protein 1 (MACC1)-AS1 promoted MACC1 mRNA stability
and expression through AMPK activation and subsequent Lin28
translocation from the nucleus to the cytoplasm (44). The most common mode of lncRNA
function is as ceRNAs to sponge certain miRNAs, hence relieving
repression of target mRNAs at a post-transcriptional level
(45), and the present study
validated that FAM83A-AS1 regulated the expression of FAM83A via a
ceRNA network by sponging miR-495-3p. miR-495-3p act as a tumor
suppressor in multiple cancer types (46–48). In
lung cancer, miR-495-3p is associated with prolonged OS time of
patients treated with nivolumab (49) and is a novel biomarker to predict
radiotherapy response clinically (50). The FAM83A-AS1/miR-495-3p/FAM83A axis
played important roles in the regulation of LUAD progression and
the current results provided new evidence for the pathogenesis of
LUAD. The limitation of the present study is the lack of an
interaction network involving FAM83A-AS1 and miR-495-3p. Mapping
the proposed network of ceRNAs will determine whether miR-495-3p is
part of a wider ceRNA network. Additional studies should be
conducted to further our understanding of the role of this ceRNA
network in lung cancer.
In conclusion, the current study demonstrated that
FAM83A and FAM83A-AS1 were both upregulated in LUAD and their
expression is associated with advanced stages and poor prognosis of
patients with LUAD. Moreover, knockdown of FAM83A or FAM83A-AS1
could inhibit proliferation, migration and invasion of LUAD and
FAM83A-AS1 regulated the expression of FAM83A via sponging
miR-495-3p. These results highlighted that FAM83A and FAM83A-AS1
are promising novel targets for the diagnosis and therapy of
patients with LUAD.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The study was funded by The National Natural Science
Foundation for Youth of China (grant no. 81902354), The Science and
Technology Program of Xu Zhou (grant no. KC18037), The Xuzhou
Clinical Technology Key Research Project (grant no. 2019GG021) and
The Excellent Talents Fund Project of Xuzhou Medical University
(grant no. XYFY2020017).
Availability of data and materials
The datasets used/and or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YS and JL designed and supervised the study. GW, XL
and YY performed most experiments and wrote the manuscript. ZJ and
HZ helped to perform parts of experiments. KX analyzed the data and
revised the article. YS and JL confirm the authenticity of all raw
data. All authors contributed to the article and approved the
submitted version. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen Z, Fillmore CM, Hammerman PS, Kim CF
and Wong KK: Non-small-cell lung cancers: A heterogeneous set of
diseases. Nat Rev Cancer. 14:535–546. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hirsch FR, Scagliotti GV, Mulshine JL,
Kwon R, Curran WJ Jr, Wu YL and Paz-Ares L: Lung cancer: Current
therapies and new targeted treatments. Lancet. 389:299–311. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cipriano R, Bryson BL, Miskimen KL, Bartel
CA, Hernandez-Sanchez W, Bruntz RC, Scott SA, Lindsley CW, Brown HA
and Jackson MW: Hyperactivation of EGFR and downstream effector
phospholipase D1 by oncogenic FAM83B. Oncogene. 33:3298–3306. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cipriano R, Miskimen KL, Bryson BL, Foy
CR, Bartel CA and Jackson MW: Conserved oncogenic behavior of the
FAM83 family regulates MAPK signaling in human cancer. Mol Cancer
Res. 12:1156–1165. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bartel CA, Parameswaran N, Cipriano R and
Jackson MW: FAM83 proteins: Fostering new interactions to drive
oncogenic signaling and therapeutic resistance. Oncotarget.
7:52597–52612. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Iyer MK, Niknafs YS, Malik R, Singhal U,
Sahu A, Hosono Y, Barrette TR, Prensner JR, Evans JR, Zhao S, et
al: The landscape of long noncoding RNAs in the human
transcriptome. Nat Genet. 47:199–208. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Blokhin I, Khorkova O, Hsiao J and
Wahlestedt C: Developments in lncRNA drug discovery: Where are we
heading? Expert Opin Drug Discov. 13:837–849. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lin C and Yang L: Long noncoding RNA in
cancer: Wiring signaling circuitry. Trends Cell Biol. 28:287–301.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jiang MC, Ni JJ, Cui WY, Wang BY and Zhuo
W: Emerging roles of lncRNA in cancer and therapeutic
opportunities. Am J Cancer Res. 9:1354–1366. 2019.PubMed/NCBI
|
|
12
|
Huang DW, Sherman BT, Tan Q, Kir J, Liu D,
Bryant D, Guo Y, Stephens R, Baseler MW, Lane HC and Lempicki RA:
DAVID bioinformatics resources: Expanded annotation database and
novel algorithms to better extract biology from large gene lists.
Nucleic Acids Res. 35:W169–W175. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Braschi B, Denny P, Gray K, Jones T, Seal
R, Tweedie S, Yates B and Bruford E: Genenames.org: The HGNC and
VGNC resources in 2019. Nucleic Acids Res. 47:D786–D792. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gao J, Aksoy BA, Dogrusoz U, Dresdner G,
Gross B, Sumer SO, Sun Y, Jacobsen A, Sinha R, Larsson E, et al:
Integrative analysis of complex cancer genomics and clinical
profiles using the cBioPortal. Sci Signal. 6:pl12013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Paraskevopoulou MD, Vlachos IS, Karagkouni
D, Georgakilas G, Kanellos I, Vergoulis T, Zagganas K, Tsanakas P,
Floros E, Dalamagas T and Hatzigeorgiou AG: DIANA-LncBase v2:
Indexing microRNA targets on non-coding transcripts. Nucleic Acids
Res. 44:D231–D238. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42:D92–D97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sun H, Liu K, Huang J, Sun Q, Shao C, Luo
J, Xu L, Shen Y and Ren B: FAM111B, a direct target of p53,
promotes the malignant process of lung adenocarcinoma. Onco Targets
Ther. 12:2829–2842. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
He J and Yu J: Long noncoding RNA
FAM83A-AS1 facilitates hepatocellular carcinoma progression by
binding with NOP58 to enhance the mRNA stability of FAM83A. Biosci
Rep. 39:BSR201925502019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Huang GM, Zang HL, Geng YX and Li YH:
LncRNA FAM83A-AS1 aggravates the malignant development of
esophageal cancer by binding to miR-495-3p. Eur Rev Med Pharmacol
Sci. 24:9408–9415. 2020.PubMed/NCBI
|
|
21
|
Wu F, Zhong Y, Lang XB, Tu YL and Sun SF:
MNX1-AS1 accelerates the epithelial-mesenchymal transition in
osteosarcoma cells by activating MNX1 as a functional oncogene. Eur
Rev Med Pharmacol Sci. 23:8194–8202. 2019.PubMed/NCBI
|
|
22
|
Sun J, Wang X, Fu C, Wang X, Zou J, Hua H
and Bi Z: Long noncoding RNA FGFR3-AS1 promotes osteosarcoma growth
through regulating its natural antisense transcript FGFR3. Mol Biol
Rep. 43:427–436. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu LM, Wu SG, Chen F, Wu Q, Wu CM, Kang
CM, He X, Zhang RY, Lu ZF, Li XH, et al: Atorvastatin inhibits
pyroptosis through the lncRNA NEXN-AS1/NEXN pathway in human
vascular endothelial cells. Atherosclerosis. 293:26–34. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bi X, Guo XH, Mo BY, Wang ML, Luo XQ, Chen
YX, Liu F, Olsen N, Pan YF and Zheng SG: LncRNA PICSAR promotes
cell proliferation, migration and invasion of fibroblast-like
synoviocytes by sponging miRNA-4701-5p in rheumatoid arthritis.
EBioMedicine. 50:408–420. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kong X, Duan Y, Sang Y, Li Y, Zhang H,
Liang Y, Liu Y, Zhang N and Yang Q: LncRNA-CDC6 promotes breast
cancer progression and function as ceRNA to target CDC6 by sponging
microRNA-215. J Cell Physiol. 234:9105–9117. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xu F, Zha G, Wu Y, Cai W and Ao J:
Overexpressing lncRNA SNHG16 inhibited HCC proliferation and
chemoresistance by functionally sponging hsa-miR-93. Onco Targets
Ther. 11:8855–8863. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hoffman PC, Mauer AM and Vokes EE: Lung
cancer. Lancet. 355:479–485. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bozatzi P and Sapkota GP: The FAM83 family
of proteins: From pseudo-PLDs to anchors for CK1 isoforms. Biochem
Soc Trans. 46:761–771. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Snijders AM, Lee SY, Hang B, Hao W and
Bissel MJ: FAM83 family oncogenes are broadly involved in human
cancers: An integrative multi-omics approach. Mol Oncol.
11:167–179. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lin Q, Chen H, Zhang M, Xiong H and Jiang
Q: Knocking down FAM83B inhibits endometrial cancer cell
proliferation and metastasis by silencing the PI3K/AKT/mTOR
pathway. Biomed Pharmacother. 115:1089392019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu X, Gao H, Zhang J and Xue D: FAM83D is
associated with gender, AJCC stage, overall survival and
disease-free survival in hepatocellular carcinoma. Biosci Rep.
39:BSR201816402019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kim KM, Hussein UK, Park SH, Park SH, Kang
MA, Moon YJ, Zhang Z, Song Y, Park HS, Bae JS, et al: FAM83H is
involved in stabilization of β-catenin and progression of
osteosarcomas. J Exp Clin Cancer Res. 38:2672019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lee SY, Meier R, Furuta S, Lenburg ME,
Kenny PA, Xu R and Bissell MJ: FAM83A confers EGFR-TKI resistance
in breast cancer cells and in mice. J Clin Invest. 122:3211–3220.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Parameswaran N, Bartel CA,
Hernandez-Sanchez W, Miskimen KL, Smigiel JM, Khalil AM and Jackson
MW: A FAM83A positive feed-back loop drives survival and
tumorigenicity of pancreatic ductal adenocarcinomas. Sci Rep.
9:133962019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Peng WX, Koirala P and Mo YY:
LncRNA-Mediated regulation of cell signaling in cancer. Oncogene.
36:5661–5667. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Balbin OA, Malik R, Dhanasekaran SM,
Prensner JR, Cao X, Wu YM, Robinson D, Wang R, Chen G, Beer DG, et
al: The landscape of antisense gene expression in human cancers.
Genome Res. 25:1068–1079. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wanowska E, Kubiak MR, Rosikiewicz W,
Makałowska I and Szcześniak MW: Natural antisense transcripts in
diseases: From modes of action to targeted therapies. Wiley
Interdiscip Rev RNA. 9:e14612018. View Article : Google Scholar
|
|
38
|
Wahlestedt C: Targeting long non-coding
RNA to therapeutically upregulate gene expression. Nat Rev Drug
Discov. 12:433–446. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shi R, Jiao Z, Yu A and Wang T: Long
noncoding antisense RNA FAM83A-AS1 promotes lung cancer cell
progression by increasing FAM83A. J Cell Biochem. 120:10505–10512.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Xiao G, Wang P, Zheng X, Liu D and Sun X:
FAM83A-AS1 promotes lung adenocarcinoma cell migration and invasion
by targeting miR-150-5p and modifying MMP14. Cell Cycle.
18:2972–2985. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Pelechano V and Steinmetz LM: Gene
regulation by antisense transcription. Nat Rev Genet. 14:880–893.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Han X, Huang S, Xue P, Fu J, Liu L, Zhang
C, Yang L, Xia L, Sun L, Huang SK and Zhou Y: LncRNA PTPRE-AS1
modulates M2 macrophage activation and inflammatory diseases by
epigenetic promotion of PTPRE. Sci Adv. 5:eaax92302019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chang H, Li B, Zhang X and Meng X:
NCK1-AS1 promotes NCK1 expression to facilitate tumorigenesis and
chemo-resistance in ovarian cancer. Biochem Biophys Res Commun.
5:292–299. 2020. View Article : Google Scholar
|
|
44
|
Zhao Y, Liu Y, Lin L, Huang Q, He W, Zhang
S, Dong S, Wen Z, Rao J, Liao W and Shi M: The lncRNA MACC1-AS1
promotes gastric cancer cell metabolic plasticity via AMPK/Lin28
mediated mRNA stability of MACC1. Mol Cancer. 17:692018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chan JJ and Tay Y: Noncoding RNA:RNA
regulatory networks in cancer. Int J Mol Sci. 19:13102018.
View Article : Google Scholar
|
|
46
|
Zhao G, Zhang L, Qian D, Sun Y and Liu W:
MiR-495-3p inhibits the cell proliferation, invasion and migration
of osteosarcoma by targeting C1q/TNF-related protein 3. Onco
Targets Ther. 12:6133–6143. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Chen S, Wu J, Jiao K, Wu Q, Ma J, Chen D,
Kang J and Zhao G, Shi Y, Fan D and Zhao G: MicroRNA-495-3p
inhibits multidrug resistance by modulating autophagy through
GRP78/mTOR axis in gastric cancer. Cell Death Dis. 9:10702018.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Eun JW, Kim HS, Shen Q, Yang HD, Kim SY,
Yoon JH, Park WS, Lee JY and Nam SW: MicroRNA-495-3p functions as a
tumor suppressor by regulating multiple epigenetic modifiers in
gastric carcinogenesis. J Pathol. 244:107–119. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Halvorsen AR, Sandhu V, Sprauten M, Flote
VG, Kure EH, Brustugun OT and Helland Å: Circulating microRNAs
associated with prolonged overall survival in lung cancer patients
treated with nivolumab. Acta Oncol. 57:1225–1231. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Chen X, Xu Y, Liao X, Liao R, Zhang L, Niu
K, Li T, Li D, Chen Z, Duan Y and Sun J: Plasma miRNAs in
predicting radiosensitivity in non-small cell lung cancer. Tumour
Biol. 37:11927–11936. 2016. View Article : Google Scholar : PubMed/NCBI
|