Introduction
According to global cancer statistics in 2018,
breast cancer (BC) is the most commonly diagnosed malignancy and
the leading cause of cancer-associated mortality among women, with
~17,000,000 new cases occurring annually worldwide in 2018
(1,2), posing a serious threat to the
well-being and survival of patients. With advances in cancer
research, several signal transduction pathways, including MAPK and
PI3K-Akt, and signaling pathway-associated molecules have been
found to be abnormally activated in BC, which may serve an
important role in the occurrence and development of cancer
(3). However, due to the complexity
of eukaryotic genome expression, numerous aspects of the molecular
regulation of BC remain elusive. Therefore, it is urgent to
identify potential biological biomarkers with higher specificity
and sensitivity for BC.
Long non-coding RNAs (lncRNAs) are segments of
non-coding RNA >200 nucleotides in length (4) that are mostly present in the nucleus
and cytoplasm. They are formed by transcription of RNA polymerase
II and then matured by splicing and head-to-tail junction (5). In recent years, with the development
and maturity of gene microarray and RNA sequencing technology, the
research in the field of lncRNA has gradually advanced. It has been
demonstrated that, although lncRNAs are a class of non-coding RNAs,
they may regulate gene expression by epigenetic, transcriptional
and post-transcriptional pathways, thereby regulating cell
proliferation and metabolism, and that they participate in the
regulation of various pathological processes in organisms (6,7).
As previously reported, LINC00052 is a type of
lncRNA that serves an important role in cancer cell migration and
invasion (8). For example, Xiong
et al (8) reported that
LINC00052 could sponge microRNA (miRNA/miR)-485-3p and miR-128 to
regulate the expression levels of neurotrophic receptor tyrosine
kinase 3 (NTRK3) and promote hepatocellular carcinoma (HCC) cell
metastasis. In addition to the role of LINC00052 in HCC, it was
proved that it could promote cell proliferation and metastasis in
gastric cancer (9). Despite these
findings, the exact role and function of LINC00052 in BC remains
unclear.
miRNAs are a class of small non-coding RNAs that are
18–25 nucleotides in length and are present in plants, fungi,
animals and viruses (10). miRNAs
can bind directly to the 3′-untranslated regions (UTRs) of their
target mRNAs; each miRNA can bind and regulate several target
genes, and at the same time it can also be regulated by several
genes (11). It has been
demonstrated that an lncRNA may bind to a miRNA as a competing
endogenous RNA (ceRNA) to affect the occurrence and development of
tumors (12). However, the mechanism
through which LINC00052 binds to miRNAs in BC remains unclear.
The aim of the present study was to investigate
LINC00052 expression in BC tissues and compare it with that in
adjacent normal tissue. The effects of LINC00052 on the
proliferation and metastasis of BC cells were investigated by Cell
Counting Kit-8 (CCK-8), wound healing and Transwell assays, and the
interactions among LINC00052, miR-145-5p and TGF-β receptor II
(TGFBR2) were explored by reverse transcription-quantitative PCR
(RT-qPCR) and western blot analyses. The aim was to determine
whether LINC00052 regulated TGFBR2 expression by sponging
miR-145-5p and to elucidate the functional role of LINC00052 in BC,
in the hope of providing a new perspective on the diagnosis and
treatment of BC.
Patients and methods
Human tissue samples
A total of 45 pairs of BC and adjacent tissues
(>1 cm from the tumor) were collected from patients at the
Tongji Medical College of Huazhong Science and Technology
University (Wuhan, China) between July 2017 and December 2017. The
45 fresh BC samples were acquired from surgery and immediately
frozen in liquid nitrogen and kept at −80°C until total RNA was
extracted. All patients were female, with a median age of 55 years
old (range, 33–67 years old). None of the patients had received
preoperative chemotherapy or radiotherapy. The procedures followed
the ethical standards of the responsible committees on human
experimentation (institutional and national). Written informed
consent was obtained from each patient. The present study was
approved by the Ethics Committee of Tongji Hospital at Tongji
Medical College of Huazhong Science and Technology University.
Cell lines and culturing
The human BC cell lines (MDA-MB-231, MDA-MB-468,
T47D, SKBR3 and MCF-7) and the normal breast MCF10A cell line were
obtained from the Cell Bank of Type Culture Collection of the
Chinese Academy of Sciences. The MDA-MB-231 cells were cultured in
L15 medium, while the MDA-MB-468 and T47D cell lines were cultured
in RPMI-1640 medium, and the MCF-7 and SKBR3 cell lines were
cultured in DMEM (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Sigma-Aldrich; Merck KGaA) at 37°C with
5% CO2.
Database analysis
The present study included 1,222 samples obtained
from The Cancer Genome Atlas (TCGA; http://portal.gdc.cancer.gov) database, consisting of
1,109 tumor samples and 113 normal breast tissue samples, and 113
pairs of cancer and adjacent normal tissues. The BC RNA expression
data (level 3) of the corresponding patients were downloaded from
TCGA Data Portal (November 2017). The RNA sequencing raw reads
(lncRNA and mRNA) were post-processed and normalized using TCGA
RNASeqv2 system (TCGA; http://portal.gdc.cancer.gov). The normalized miRNA
expression data were downloaded from TCGA Data Portal and quantile
normalized before performing the analysis. TargetScan Human v7.2
(http://www.targetscan.org/mamm_31/)
was used to predict the TGFBR2 gene targeted by miR-145-5p.
DIANA-LncBase (http://www.microrna.gr/LncBase) was used to predict
the LINC00052/miR-145-5p interactions.
Cell transfection
Small interfering (si)RNAs against LINC00052 and a
scrambled control siRNA used as a negative control (si-NC) were
purchased from Guangzhou RiboBio Co., Ltd. miR-145-5p mimics
(miR-145-5p-mimics), inhibitors (miR-145-5p-inhibitor), or
corresponding scrambled controls (miR-NC) were synthesized by
Guangzhou RiboBio Co., Ltd. siRNAs, miR mimics, miR inhibitors and
their NC oligonucleotides (50 nM) were transfected using
Lipofectamine™ 3000 reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) into the MDA-MB-231 and MCF-7 cells separately in 6-well
plates of cells at 50–60 or 70–80% confluency, respectively,
according to the manufacturer's protocol. Each well was transfected
with 50 nM siRNAs. After 48 h of transfection at 37°C, the cells
were collected for subsequent experiments. The sequences of siRNAs
used are listed in Table I.
 | Table I.Sequences of siRNA, mimics,
inhibitors and the NC used for transfection. |
Table I.
Sequences of siRNA, mimics,
inhibitors and the NC used for transfection.
| Name | Sequences |
|---|
| siRNA-NC |
5′-TTCTCCGAACGTGTCACGTdTdT-3′ |
| siLINC00052-1 |
5′-UUAUUCACAUCACUGCAUGTT-3′ |
| siLINC00052-2 |
5′-UUUCAGAUAUGCCAAGCUCTT-3′ |
| NC mimic sense |
5′-UUUGUACUACACAAAAGUACUG-3′ |
| NC mimic
anti-sense |
3′-AAACAUGAUGUGUUUUCAUGAC-5′ |
| miR-145-5p mimic
sense |
5′-GUCCAGUUUUCCCAGGAAUCCCU-3′ |
| miR-145-5p mimic
anti-sense |
3′-AGGGAUUCCUGGGAAAACUGGAC-5′ |
| NC inhibitor |
5′-CAGUACUUUUGUGUAGUACAAA-3′ |
| miR-145-5p
inhibitor |
5′-AGGGAUUCCUGGGAAAACUGGAC-3′ |
Cell proliferation assay
MDA-MB-231 and MCF-7 cells (5×103
cells/well) were treated with si-NC, si-1 and si-2 separately (50
nM) for 8 h at 37°C. After transfection, cells were digested with
trypsin, and 2×103 cells/well were seeded in a 96-well
culture plate. Cell proliferation was determined using a CCK-8
assay for 10 min (Dojindo Molecular Technologies, Inc.) and the
absorbance was measured at 450 nm (Molecular Devices, LLC).
Wound healing assay
The MDA-MB-231-NC, MDA-MB-231-siLINC00052-1,
MDA-MB-231-siLINC00052-2, MCF-7-NC, MCF-7-siLINC00052-1 and
MCF-7-siLINC00052-2 cells were seeded in 6-well plates at
2×105 cells/well. When the confluence had reached 90%,
the scratch wounds were created across each well using a 200-µl
sterile micropipette plastic tip. Subsequently, the cells were
cultured for 24 h at 37°C with serum-free medium (L15 medium for
MDA-MB-231 cells or DMEM for MCF-7 cells); Thermo Fisher
Scientific, Inc.). Images of each scratch were captured for 5
fields of view in three triplicates at 0 and 24 h using a light
microscope (Carl Zeiss AG; magnification, ×200). Cells in the
marked area between the lines and the edges of the wound were
counted using Image-Pro Plus 6.0 (Media Cybernetics, Inc.). The
cell migration rate was calculated using GraphPad Prism 5.0
(GraphPad Software, Inc.).
Cell migration and invasion assay
Following MCF-7 cell transfection with NC, si-1 and
si-2, Transwell migration and Matrigel invasion assays, using a
24-well chamber with an 8-µm-pore filter (Corning, Inc.), were used
to investigate the in vitro effects of LINC00052 on cell
migration and invasion. For the Transwell migration assay, the
MDA-MB-231 cells, transfected with siRNA, were trypsinized and 200
µl cell suspension (2×104 cells/ml) was added to the
upper chamber of each insert (Corning, Inc.) containing the
uncoated membrane and resuspended in L15 medium without serum (200
µl). The lower chambers were supplemented with 30–40% FBS (500 µl).
After incubation for 24 h at 37°C, the upper surface of the
membrane was removed with a cotton tip, while the cells on the
lower surface were stained for 10–15 min with 0.1% crystal violet
at room temperature. For the invasion assay, Matrigel chambers (BD
Biosciences) were performed according to the manufacturer's
instructions. Briefly, the transfected MDA-MB-231 cells (200 µl;
5,000 cells per well) were collected, resuspended in DMEM without
serum, and then added to the upper chamber containing Matrigel (50
µl). DMEM medium (500 µl) supplemented with 10% FBS was added to
the lower chambers and incubated overnight at 37°C. The cells on
the upper surface were scraped, whereas the invasive cells on the
lower surface were fixed for 20 min at room temperature, then
stained with 0.1% crystal violet for 10–15 min at room temperature.
The migratory and invasive abilities were determined by counting
the cells that migrated to the lower surface of the membrane. All
cells were counted under a light microscope (Carl Zeiss AG) at a
magnification of ×200 in ≥5 randomly selected fields in
triplicate.
Cytoplasmic and nuclear
fractionation
Cytoplasmic and nuclear fractionation was performed
using a PARIS™ kit (Ambion; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. MDA-MB-231 or MCF-7 cells
(1×107 cells/well) were collected and resuspended in
cell fraction buffer for cytoplasmic and nuclear fractionation, and
then placed on ice for 10 min. After centrifugation with 5,000 × g
for 30 sec at 4°C according to the manufacturer's instructions, the
supernatant and nuclear pellet were separated using a cell
disruption buffer to be preserved for RNA extraction.
RT-qPCR analysis
Total RNA from the human tissue samples or the cell
lines was extracted using TRIzol® reagent (Takara
Biotechnology Co., Ltd.) according to the manufacturer's protocol.
Total RNA was reverse transcribed using HiScript® II RT
SuperMix (Vazyme Biotech Co., Ltd.) according to the manufacturer's
protocol. The sequences of primers used are listed in Table II. U6 and GAPDH were used as the
control for miR-145-5p and TGFBR2, respectively. qPCR was performed
in a Roche LightCycle480 II Real-Time PCR Detection System using
SYBR Premix Ex Taq (cat. no. RR420A; Takara Bio, Inc.). OneStep PCR
parameters for lncRNA and TGFBR2 quantification were as follows:
37°C for 60 min for RT, 10 min at 95°C and then 45 cycles of 20 sec
at 95°C and 5 min at 72°C. All the measurements were conducted in
triplicates. Quantitative mRNA data were normalized and presented
as a ratio to GAPDH calculated using the 2−ΔΔCq method
(13).
| Table II.Sequences of RT-qPCR primers. |
Table II.
Sequences of RT-qPCR primers.
| Sequences | RT-qPCR
primers |
|---|
| TGFBR2F |
5′-TGTGATGTGAGATTTTCCACCTGT-3′ |
| TGFBR2R |
5′-TGTTCTCGTCATTCTTTCTCCATAC-3′ |
| LINC00052F |
5′-GTGAACCTTCGACCTTGGACTT-3′ |
| LINC00052R |
5′-AGAGAGGGAGGGAGACTGAGATT-3′ |
| miR-145-5pF |
5′-CCTTGTCCTCACGGTCCAGT-3′ |
| miR-145-5pR |
5′-AACCATGACCTCAAGAACAGTATTT-3′ |
| GAPDHF |
5′-GGAAGCTTGTCATCAATGGAAATC-3′ |
| GAPDHR |
5′-TGATGACCCTTTTGGCTCCC-3′ |
| U6F |
5′-GCTTCGGCAGCACATATACTAAAAT-3′ |
| U6R |
5′-CGCTTCACGAATTTGCGTGTCAT-3′ |
Western blot analysis
Since the premise of the lncRNA-miRNA-mRNA ceRNA
regulatory network occurrence was in the cytoplasm, the separation
of cytoplasmic and nuclear fractions was first performed to confirm
that LINC00052 was expressed in the cytoplasm. Subsequently, the
cytoplasm and nuclear fractions were isolated, and the proteins
were extracted from the cytoplasm for further experimental
verification of the ceRNA regulatory network. Briefly, cells
transfected with miR-145-5p inhibitor and LINC00052 siRNAs, or
transfected with miR-145-5p mimic were lysed in RIPA buffer [50 mM
Tris (pH 7.4), 150 mM NaCl, 1 NP-40, 0.5% sodium deoxycholate;
Beyotime Institute of Biotechnology] containing protease inhibitor
cocktail (Abcam), followed by protein concentration detection using
the Bradford protein assay. Next, 20 µg proteins/lane were
subjected to 10% SDS-PAGE and transferred onto nitrocellulose
membranes (Merck KGaA). The membranes were blocked at 26°C in
TBS-Tween-20 (0.1%) with 5% skimmed milk for 1 h, then incubated
overnight with the TGFBR2 primary antibody (1:200; cat. no.
ab184948) at 4°C. Blots were incubated with horseradish
peroxidase-conjugated goat anti-rabbit IgG (1:3,000; cat. no.
ab150077; Abcam) and goat anti-mouse IgG (1:3,000; cat. no.
ab150113; Abcam) secondary antibodies for 1 h at 26°C, then
detected using an enhanced chemiluminescence kit (Thermo Fisher
Scientific, Inc.), imaged using a Gel Doc XR+ imaging system
(Bio-Rad Laboratories, Inc.) and analyzed using Image Lab software
v3.0 (Bio-Rad Laboratories, Inc.). GAPDH was used as an internal
reference. Mouse anti-GAPDH (1:1,000; cat. no. ab8245) and rabbit
anti-TGFBR2 were purchased from Abcam.
Statistical analysis
Statistical analysis was performed using SPSS 17.0
(SPSS, Inc.). and the results are presented as the mean ± SEM. The
Kolmogorov-Smirnov test was used to assess the normality assumption
of data distribution. A paired t-test (for adjacent vs. normal
tissues) or the Mann-Whitney U test were used to compare the
difference between two groups when applicable. Expression
differences among three groups were analyzed by ANOVA followed by
Tukey's post-hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
LINC00052 expression is upregulated in
BC
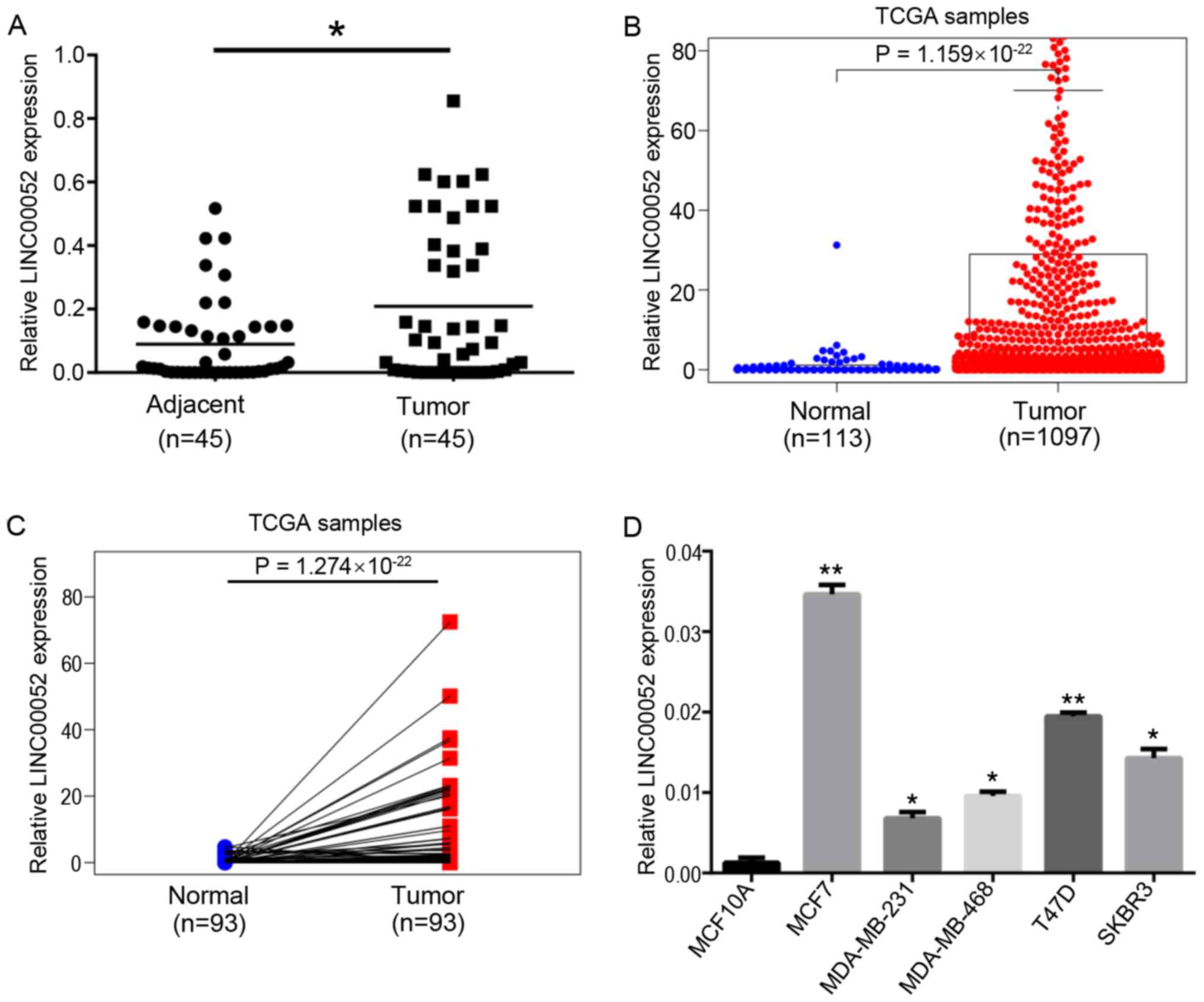
In the present study, RT-qPCR was used to detect
LINC00052 expression in 45 pairs of BC and adjacent normal breast
tissues. Compared with in normal breast tissues, LINC00052
expression in BC tissues was significantly upregulated (Fig. 1A). Subsequently, LINC00052 expression
in BC was analyzed using TCGA database. The expression levels of
LINC00052 in BC tissues were significantly higher compared with
those in normal tissues (Fig. 1B).
In addition, the results from 113 pairs of cancer samples and
adjacent tissues from TCGA demonstrated that LINC00052 expression
in tumor tissues was significantly higher compared with that in
paracancerous tissues (Fig. 1C).
Similarly, higher expression levels of LINC00052 were observed in
BC cell lines compared with that in the normal breast MCF10A cells
(Fig. 1D).
LINC00052 silencing inhibits BC cell
proliferation, migration and invasion
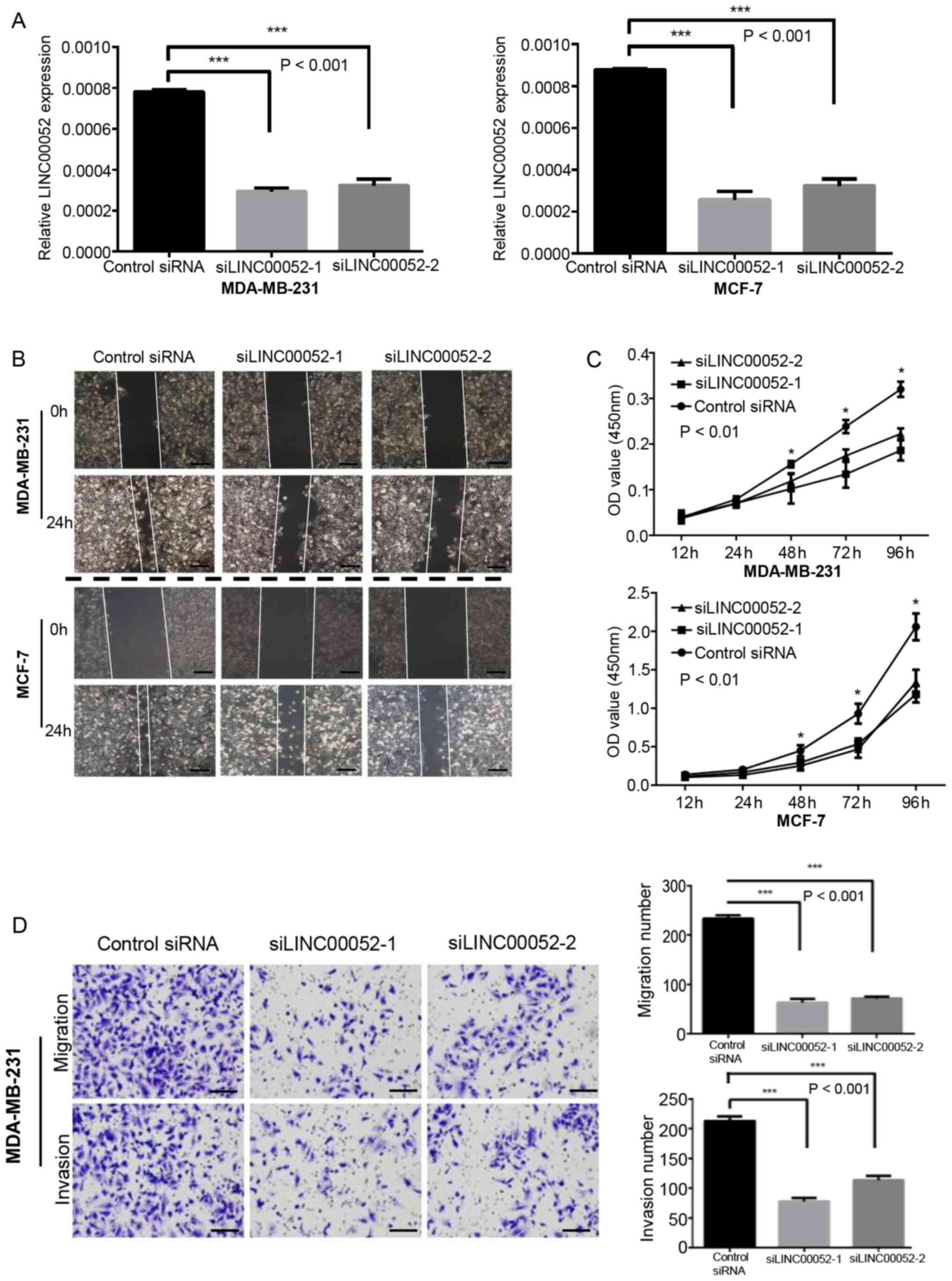
To further explore the role of LINC00052 in BC
development, two LINC00052 siRNAs, named si-1 and si-2, were used.
In the transfection experiment, si-1 and si-2 achieved good gene
interference effects in MDA-MB-231 and MCF-7 cells (Fig. 2A). The CCK-8 assay demonstrated that,
once LINC00052 was silenced, the proliferative ability of
MDA-MB-231 and MCF-7 cells was significantly decreased, indicating
that LINC00052 may serve an important role in promoting the
proliferation of BC cells (Fig. 2B).
The wound healing assay demonstrated that the migratory ability of
MDA-MB-231 and MCF-7 cells was markedly lower compared with that in
the control group following LINC00052 knockdown (Fig. 2C). The Transwell assay results
revealed that the migration and invasion of MDA-MB-231 cells
transfected with LINC00052 siRNA were significantly inhibited
(Fig. 2D). MCF-7 cells were also
used for the Transwell assay, but after 48 h of observation, no
cells had migrated to the lower chamber (data not shown). Overall,
the present results suggested that LINC00052 may promote the
proliferation, migration and invasion of BC cells.
Cellular localization and target
prediction of LINC00052
According to previous studies, lncRNAs in the
nucleus can function as scaffolds for proteins to form
ribonucleoprotein complexes and as guides to recruit
chromatin-modifying complexes to target genes (14–16). In
addition, lncRNAs in the cytoplasm may function as ceRNAs by
competitively binding to miRNAs (17,18).
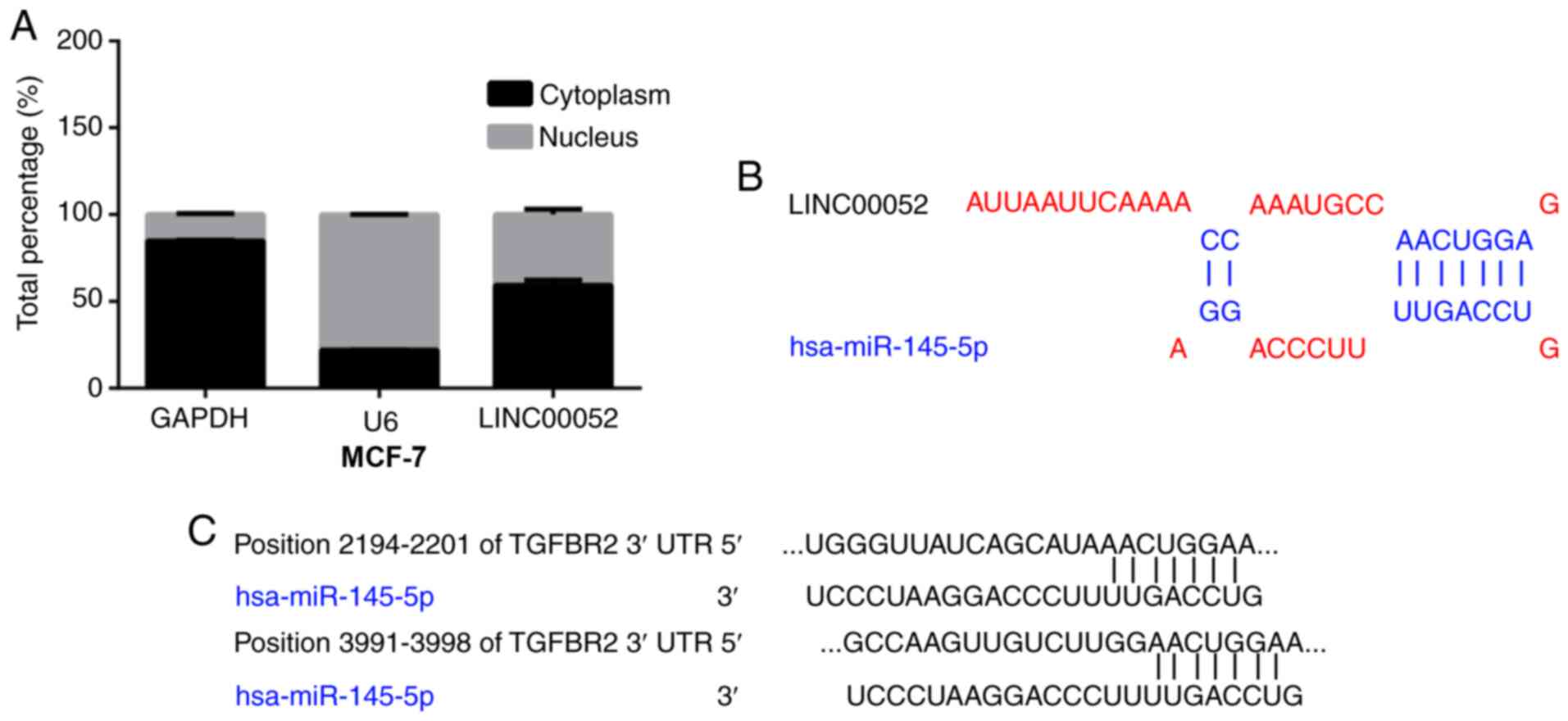
Thus, the intracellular distribution of LINC00052 was investigated
by cytoplasmic/nuclear localization experiments. GAPDH has always
been considered as an enzyme that only distributes in the cytoplasm
(19–21). Therefore, GAPDH was used as the
internal reference for cytoplasmic and nuclear fractionation. The
cytoplasmic and nuclear fractions were first isolated, and then the
mRNA expression levels were detected by RT-qPCR. The results
indicated that GAPDH RNA was relatively stable in the cytoplasm and
U6 was relatively stable in the nucleus, while LINC00052 was
expressed in both the cytoplasm and nucleus (Fig. 3A). In the present study, the
bioinformatics prediction software TargetScan, was used to predict
the target gene of miR-145-5p, and the binding sites between
miR-145-5p and TGFBR2 were successfully predicted, revealing that
there was >1 site of TGFBR2 binding to miR-145-5p (Fig. 3C). Furthermore, the binding site
between LINC00052 and miR-145-5p was obtained using the
DIANA-LncBase database (Fig. 3B).
The aforementioned results provide theoretical evidence for the
regulatory mechanism of action of LINC00052.
miR-145-5p is involved in LINC00052
upregulating TGFBR2 expression in BC
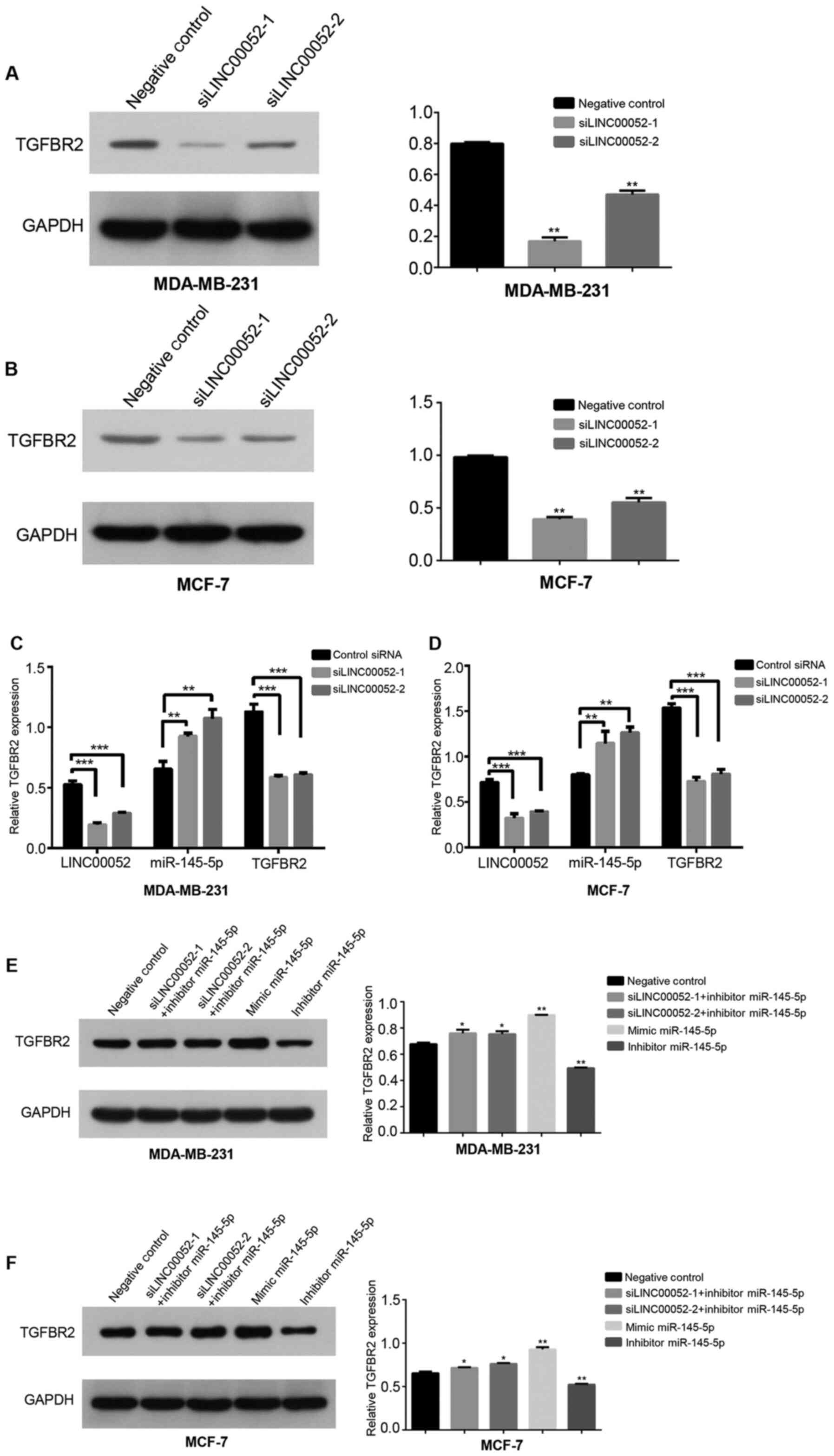
In order to elucidate whether mir-145-5p was
involved in the regulation of TGFBR2 expression mediated by
LINC00052, the mRNA and protein expression levels of TGFBR2 were
detected by RT-qPCR and western blot analysis, respectively, after
transfection. In the transfection experiment, mir-145-5p mimic and
mir-145-5p inhibitor achieved good gene interference effects in
MDA-MB-231 and MCF-7 cells (Fig. 4C and
D). The results demonstrated that knockdown of LINC00052 in BC
cells significantly decreased the mRNA (Fig. 4E and F) and protein (Fig. 4A and B) expression levels of TGFBR2,
and significantly increased the expression levels of mir-145-5p
(Fig. 4E and F). However, knockdown
of LINC00052 combined with knockdown of miR-145-5p in BC cells was
associated with lower protein expression levels of TGFBR2 compared
with those with miR-145-5p-knockdown alone (Fig. 4G and H). In addition, overexpression
of miR-145-5p significantly decreased the protein expression levels
of TGFBR2, and knockdown of miR-145-5p significantly increased the
protein expression levels of TGFBR2 (Fig. 4G and H). These results suggested that
LINC00052 may upregulate the expression levels of TGFBR2 by
interacting with miR-145-5p.
Discussion
Previous studies have established that lncRNAs serve
a key role in tumorigenesis and tumor development (22–25).
However, the potential functions and underlying mechanisms of these
lncRNAs in BC remain unclear. A previous study reported low
expression levels of LINC00052 in HCC, and that LINC00052 inhibited
the migration and invasion of HCC cells by targeting NTRK3 and
erythrocyte membrane protein band 4.1 like 3 (8). By contrast, Salameh et al
(26) reported that LINC00052
expression was upregulated in BC and promoted tumor growth through
HER3 signaling. The different mechanisms of action of LINC00052 in
BC progression warrant further investigation. It has been reported
that LINC00052 has an oncogenic function in gastric cancer,
suggesting that LINC00052 may act as either an oncogene or a tumor
suppressor, with its different functions depending on the different
biological processes (9).
In the present study, it was observed that LINC00052
expression was upregulated in BC through TCGA database analysis and
RT-qPCR analysis of 45 pairs of BC samples and normal tissues. In
addition, RT-qPCR and western blot assays were performed to confirm
the effects of LINC00052 on the mRNA and protein expression levels
of TGFBR2. The present results revealed that LINC00052 regulated
TGFBR2 expression. These results suggested that LINC00052 may be a
factor promoting cell proliferation and metastasis by regulating
TGFBR2 in BC. Additionally, the present study revealed that
LINC00052 was differentially expressed in BC samples. Unlike
previous studies (27–29), LINC00052 expression was upregulated
in BC tissue samples compared with in normal breast tissues,
possibly acting as a tumor promoter by enhancing cell proliferation
and metastasis (25). The underlying
reason may be that the overexpression or downregulation of lncRNAs
differ among different types of tumor (30). In addition, a novel
LINC00052/miR-145-5p/TGFBR2 regulatory axis in BC was identified,
resulting in accelerated tumor progression.
TGF-β is a secreted ligand that exerts its effects
through a transmembrane heteromeric receptor complex, which
consists of TGFBR1 and TGFBR2 (31).
Aberrant TGF-β signaling contributes to tumor metastasis and is a
common finding in human cancer (32). Additionally, it has been reported
that myeloid cells are recruited into mammary carcinomas with
TGFBR2 deletion and directly promote tumor metastasis (33). It has been reported that the
combination of TGFBR2 and TGF-β can activate the downstream
TGF-β/Smad signaling pathway in lung cancer, inhibit c-myc
expression, promote the expression levels of p21, p15 and other
cell cycle inhibitors, block the cell cycle and promote the
antitumor effect of the TGF-β/Smad signaling pathway (34). A previous study revealed that TGFBR2
could activate the MAPK signaling pathway to promote the
proliferation of tumor cells by binding to growth factor
receptor-bound protein 2 (Grb2). However, when the binding site of
TGFBR2 and Grb2 mutates, TGFBR2 and Grb2 cannot bind, leading to
the failure to activate the MAPK signaling pathway, which makes
TGFBR2 lose its role in promoting tumor cell proliferation
(35). The present study further
investigated whether LINC00052 could alter miRNA expression and
affect TGFBR2 expression. Using online softwares and RT-qPCR
analysis, miR-145-5p was identified as a potential regulatory gene
of TGFBR2. miR-145-5p, a putative tumor suppressor, is
downregulated in a variety of tumors, including endometrial cancer,
laryngeal carcinoma, gastric cancer, pancreatic adenocarcinoma and
ovarian cancer (36,37). A recent study has demonstrated that
decreased miR-145-5p expression caused by promoter methylation is a
prognostic factor for endometrial carcinoma (36). Restoration of miR-145-5p expression
results in decreased cell proliferation by targeting fascin
actin-bundling protein 1 in laryngeal carcinoma (38). Additionally, miR-145-5p delivered
from exosomes has been found to repress the growth and metastasis
of pancreatic adenocarcinoma and ovarian cancer cells (36,39).
Furthermore, miR-145-5p was found to act as a tumor suppressor by
targeting ZEB2 and N-cadherin in gastric cancer (40). The aforementioned studies suggest
that miR-145-5p may be a potential therapeutic target in
cancer.
Although miR-145-5p has been proven to target a
number of protein-coding genes (41,42), the
present study demonstrated that miR-145-5p targeted TGFBR2 and
LINC00052. First, the DIANA-LncBase database was used to predict
the interaction between miRNAs and LINC00052, and the binding site
between LINC00052 and miR-145-5p was found; similarly, the binding
sites for miR-145-5p and TGFBR2 were also successfully predicted by
TargetScan. In addition, inhibition of LINC00052 expression induced
upregulation of miR-145-5p expression and downregulation of TGFBR2
expression, indicating that LINC00052 may exert a negative
regulatory effect on miR-145-5p and a positive regulatory effect on
TGFBR2. miRNAs have been found to be involved in the regulation of
gene expression and serve an important role in a series of basic
cellular processes, such as differentiation, invasion, migration,
proliferation and apoptosis (43,44),
relying on the complementarities between the limited region of
sequence at the 5′-end of the miRNA (seed) and the 3′-UTR of
specific target mRNAs (44).
Bioinformatics analysis revealed that TGFBR2 was targeted by
miR-145-5p. Overexpression of miR-145-5p decreased TGFBR2
expression, whereas the inhibition of miR-145-5p increased TGFBR2
expression. Furthermore, whether LINC00052 can promote the
proliferation and metastasis of BC cells was investigated. The
present results indicated that LINC00052-knockdown significantly
inhibited the proliferation and migration of BC cells. Therefore,
the results of the present study highlight the importance of the
interaction between miRNAs and lncRNAs in BC. In order to elucidate
whether miR-145-5p was involved in LINC00052-mediated regulation of
TGFBR2 expression, transfection combinations were performed. The
results revealed that LINC00052-knockdown or overexpression of
miR-145-5p significantly decreased the expression levels of TGFBR2.
However, when LINC00052-knockdown was combined with miR-145-5p
inhibition, the decrease of TGFBR2 expression caused by
LINC00052-knockdown was resuced, suggesting that miR-145-5p may be
involved in LINC00052-mediated TGFBR2 expression regulation. The
present results may provide new insights into the molecular
mechanisms interconnecting LINC00052, TGFBR2 and miR-145-5p.
The present study suggested that LINC00052 may
promote cancer progression in BC by regulating miR-145-5p and
TGFBR2. To the best of our knowledge, this regulatory pathway has
not been reported in BC, which provides a new direction to find
novel therapeutic targets for BC. However, the present study
presents some limitations. The study was not able to verify the
effect of LINC00052 inhibition on the migration and invasion of
MCF-7 cells using a Transwell assay, as the MCF-7 cells did not
have invasion ability, and the study also lacked in vivo
xenograft tumor experiments and dual-luciferase reporter assays,
which are required in future experiments.
In conclusion, the preliminary results of the
present study confirmed that LINC00052 expression was upregulated
in BC and that it may competitively bind to miR-145-5p to
upregulate TGFBR2 expression, thereby serving a key role in
promoting BC.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant on. 81802676) and the
Wuhan Youth Cadre Project (grant nos. 2017zqnlxr01 and
2017zqnlxr02).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MD conceived the study. MD and HL designed the
study. MD and TX performed the experiments. MD analyzed the data
and wrote the manuscript. HL and TX confirm the authenticity of all
the raw data. XL made substantial contributions to the conception
of the study, secured funding and supervised the study. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
All procedures involving human participants in the
present study were approved by the Ethics Committee of Tongji
Hospital at Tongji Medical College of Huazhong Science and
Technology University (Wuhan, China), and written informed consent
from each participant was obtained. The procedures followed the
ethical standards of the responsible committee on human
experimentation (institutional and national).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fan L, Zheng Y, Yu KD, Liu GY, Wu J, Lu
JS, Shen KW, Shen ZZ and Shao ZM: Breast cancer in a transitional
society over 18 years: Trends and present status in Shanghai,
China. Breast Cancer Res Treat. 117:409–416. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Creighton CJ: The molecular profile of
luminal B breast cancer. Biologics. 6:289–297. 2012.PubMed/NCBI
|
|
4
|
Atianand MK, Caffrey DR and Fitzgerald KA:
Immunobiology of long noncoding RNAs. Annu Rev Immunol. 35:177–198.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Caley DP, Pink RC, Trujillano D and Carter
DR: Long noncoding RNAs, chromatin, and development.
ScientificWorldJournal. 10:90–102. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Novikova IV, Hennelly SP and Sanbonmatsu
KY: Sizing up long non-coding RNAs: Do lncRNAs have secondary and
tertiary structure? Bioarchitecture. 2:189–199. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Knauss JL and Sun T: Regulatory mechanisms
of long noncoding RNAs in vertebrate central nervous system
development and function. Neuroscience. 235:200–214. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xiong D, Sheng Y, Ding S, Chen J, Tan X,
Zeng T, Qin D, Zhu L, Huang A and Tang H: LINC00052 regulates the
expression of NTRK3 by miR-128 and miR-485-3p to strengthen HCC
cells invasion and migration. Oncotarget. 7:47593–47608. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shan Y, Ying R, Jia Z, Kong W, Wu Y, Zheng
S and Jin H: LINC00052 Promotes gastric cancer cell proliferation
and metastasis via activating the Wnt/β-catenin signaling pathway.
Oncol Res. 25:1589–1599. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wong IO, Cowling BJ, Schooling CM and
Leung GM: Age-period-cohort projections of breast cancer incidence
in a rapidly transitioning Chinese population. Int J Cancer.
121:1556–1563. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ahmadzada T, Reid G and McKenzie DR:
Fundamentals of siRNA and miRNA therapeutics and a review of
targeted nanoparticle delivery systems in breast cancer. Biophys
Rev. 10:69–86. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tam C, Wong JH, Tsui SKW, Zuo T, Chan TF
and Ng TB: LncRNAs with miRNAs in regulation of gastric, liver, and
colorectal cancers: Updates in recent years. Appl Microbiol
Biotechnol. 103:4649–4677. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Engreitz JM, Pandya-Jones A, McDonel P,
Shishkin A, Sirokman K, Surka C, Kadri S, Xing J, Goren A, Lander
ES, et al: The Xist lncRNA exploits three-dimensional genome
architecture to spread across the X chromosome. Science.
341:12379732013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huarte M, Guttman M, Feldser D, Garber M,
Koziol MJ, Kenzelmann-Broz D, Khalil AM, Zuk O, Amit I, Rabani M,
et al: A large intergenic noncoding RNA induced by p53 mediates
global gene repression in the p53 response. Cell. 142:409–419.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Simon MD, Pinter SF, Fang R, Sarma K,
Rutenberg-Schoenberg M, Bowman SK, Kesner BA, Maier VK, Kingston RE
and Lee JT: High-resolution Xist binding maps reveal two-step
spreading during X-chromosome inactivation. Nature. 504:465–469.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Poliseno L, Salmena L, Zhang J, Carver B,
Haveman WJ and Pandolfi PP: A coding-independent function of gene
and pseudogene mRNAs regulates tumour biology. Nature.
465:1033–1038. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Salmena L, Poliseno L, Tay Y, Kats L and
Pandolfi PP: A ceRNA hypothesis: The Rosetta stone of a hidden RNA
language? Cell. 146:353–358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Guan Z, Wang Y, Wang Y, Liu X, Wang Y,
Zhang W, Chi X, Dong Y, Liu X, Shao S and Zhan Q: Long non-coding
RNA LOC100133669 promotes cell proliferation in oesophageal
squamous cell carcinoma. Cell Prolif. 53:e127502020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fang Y and Long F: Circular RNA
circ_0000337 contributes to osteosarcoma via the miR-4458/BACH1
pathway. Cancer Biomark. 28:411–419. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhou C, Chen Z, Peng C, Chen C and Li H:
Long noncoding RNA TRIM52-AS1 sponges miR-514a-5p to facilitate
hepatocellular carcinoma progression through increasing MRPS18A.
Cancer Biother Radiopharm. May 8–2020.doi: 10.1089/cbr.2019.3271
(Epub ahead of print). View Article : Google Scholar
|
|
22
|
Wang J, Su Z, Lu S, Fu W, Liu Z, Jiang X
and Tai S: LncRNA HOXA-AS2 and its molecular mechanisms in human
cancer. Clin Chim Acta. 485:229–233. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Huang Y: The novel regulatory role of
lncRNA-miRNA-mRNA axis in cardiovascular diseases. J Cell Mol Med.
22:5768–5775. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jarroux J, Morillon A and Pinskaya M:
History, discovery, and classification of lncRNAs. Adv Exp Med
Biol. 1008:1–46. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bhan A, Soleimani M and Mandal SS: Long
noncoding RNA and Cancer: A new paradigm. Cancer Res. 77:3965–3981.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Salameh A, Fan X, Choi BK, Zhang S, Zhang
N and An Z: HER3 and LINC00052 interplay promotes tumor growth in
breast cancer. Oncotarget. 8:6526–6539. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhu L, Yang N, Chen J, Zeng T, Yan S, Liu
Y, Yu G, Chen Q, Du G, Pan W, et al: LINC00052 upregulates EPB41L3
to inhibit migration and invasion of hepatocellular carcinoma by
binding miR-452-5p. Oncotarget. 8:63724–63737. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lin J, Nong LL, Li MQ, Yang FC, Wang SH
and Liu MJ: LINC00052 inhibits tumor growth, invasion and
metastasis by repressing STAT3 in cervical carcinoma. Eur Rev Med
Pharmacol Sci. 23:4673–4679. 2019.PubMed/NCBI
|
|
29
|
Yan S, Shan X, Chen K, Liu Y, Yu G, Chen
Q, Zeng T, Zhu L, Dang H, Chen F, et al: LINC00052/miR-101-3p axis
inhibits cell proliferation and metastasis by targeting SOX9 in
hepatocellular carcinoma. Gene. 679:138–149. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rossi MN and Antonangeli F: LncRNAs: New
players in apoptosis control. Int J Cell Biol. 2014:4738572014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Muñoz NM, Upton M, Rojas A, Washington MK,
Lin L, Chytil A, Sozmen EG, Madison BB, Pozzi A, Moon RT, et al:
Transforming growth factor beta receptor type II inactivation
induces the malignant transformation of intestinal neoplasms
initiated by Apc mutation. Cancer Res. 66:9837–9844. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu CA, Chang CY, Hsueh KW, Su HL, Chiou
TW, Lin SZ and Harn HJ: Migration/invasion of malignant gliomas and
implications for therapeutic treatment. Int J Mol Sci. 19:11152018.
View Article : Google Scholar
|
|
33
|
Yang L, Huang J, Ren X, Gorska AE, Chytil
A, Aakre M, Carbone DP, Matrisian LM, Richmond A, Lin PC and Moses
HL: Abrogation of TGF beta signaling in mammary carcinomas recruits
Gr-1+CD11b+ myeloid cells that promote metastasis. Cancer Cell.
13:23–35. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang Y, Tan X, Tang Y, Zhang C, Xu J, Zhou
J, Cheng X, Hou N, Liu W, Yang G, et al: Dysregulated
Tgfbr2/ERK-Smad4/SOX2 signaling promotes lung squamous cell
carcinoma formation. Cancer Res. 79:4466–4479. 2019.PubMed/NCBI
|
|
35
|
Galliher-Beckley AJ and Schiemann WP: Grb2
binding to Tyr284 in TbetaR-II is essential for mammary tumor
growth and metastasis stimulated by TGF-beta. Carcinogenesis.
29:244–251. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wu X, Han Y, Liu F and Ruan L:
Downregulations of miR-449a and miR-145-5p act as prognostic
biomarkers for endometrial cancer. J Comput Biol. 27:834–844. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gao W, Zhang C, Li W, Li H, Sang J, Zhao
Q, Bo Y, Luo H, Zheng X, Lu Y, et al: Promoter
methylation-regulated miR-145-5p inhibits laryngeal squamous cell
carcinoma progression by targeting FSCN1. Mol Ther. 27:365–379.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jiang SB, He XJ, Xia YJ, Hu WJ, Luo JG,
Zhang J and Tao HQ: MicroRNA-145-5p inhibits gastric cancer
invasiveness through targeting N-cadherin and ZEB2 to suppress
epithelial-mesenchymal transition. Onco Targets Ther. 9:2305–2315.
2016.PubMed/NCBI
|
|
39
|
Ding Y, Cao F, Sun H, Wang Y, Liu S, Wu Y,
Cui Q, Mei W and Li F: Exosomes derived from human umbilical cord
mesenchymal stromal cells deliver exogenous miR-145-5p to inhibit
pancreatic ductal adenocarcinoma progression. Cancer Lett.
442:351–361. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hang W, Feng Y, Sang Z, Yang Y, Zhu Y,
Huang Q and Xi X: Downregulation of miR-145-5p in cancer cells and
their derived exosomes may contribute to the development of ovarian
cancer by targeting CT. Int J Mol Med. 43:256–266. 2019.PubMed/NCBI
|
|
41
|
Shen W, Wang Y, Wang D, Zhou H, Zhang H
and Li L: miR-145-5p attenuates hypertrophic scar via reducing
Smad2/Smad3 expression. Biochem Biophys Res Commun. 521:1042–1048.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang H, Jiang M, Liu Q, Han Z, Zhao Y and
Ji S: miR-145-5p inhibits the proliferation and migration of
bladder cancer cells by targeting TAGLN2. Oncol Lett. 16:6355–6360.
2018.PubMed/NCBI
|
|
43
|
Lagos-Quintana M, Rauhut R, Lendeckel W
and Tuschl T: Identification of novel genes coding for small
expressed RNAs. Science. 294:853–858. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|