Introduction
Endometrial cancer is one of the most common
gynecological tumors that has seen a dramatic increase in incidence
worldwide (1,2). This increase has a number of causes,
including an aging population, increased calorie intake and the
concomitant rise of obesity, and reproductive factors, such as
early age menarche, nulliparity, late-onset menopause, and use of
unopposed estrogens (3). These
reproductive factors are associated with a strong risk for
endometrial cancer. Postmenopausal women have the greatest
incidence, but 14% of endometrial cancers are diagnosed in
premenopausal women, including 5% of cases younger than 40 years
old. The increased incidence and population shift to younger women
are proposed to continue into the future (3,4).
Although patients diagnosed with early stage disease are associated
with a good prognosis of endometrial cancer, patients with
recurrent or metastatic disease have limited treatment options and
shorter overall survival (5). Thus,
understanding the pathophysiology of endometrial cancer is key to
improving its diagnosis and treatment.
No targeted therapies have been approved or
available for endometrial cancer. Several completed clinical trials
targeted deregulated pathways, such as the PI3K/AKT/mammalian
target of the rapamycin (mTOR), but to date results have been poor
(3). Therefore, other potential
targets need to be investigated. Krüppel-like factors (KLFs)
comprise a family of zinc finger transcription factor proteins that
control cell proliferation, apoptosis, migration, and
differentiation (6). Multiple KLFs
function as oncogenes or tumor suppressors depending on the
specific tissues and cellular environments (7,8). KLF5 is
upregulated and functions as an oncogene in colorectal (9), cervical (10), and breast cancer (11) and downregulated and acts as a tumor
suppressor in lung (12), and
prostate cancer (13). Although KLF5
is expressed in non-tumor endometrial epithelial (14) and stromal cells (15), the role of KLF5 in endometrial cancer
remains to be explored. Therefore, this study aims at investigating
the expression and function of KLF5 in endometrial cancer.
Materials and methods
Human subjects
This study was approved by the Ethics Committee of
Tokyo Medical University (approval no. SH3298). A total of 30
patients were enrolled after obtaining written informed consent at
the Department of Obstetrics and Gynecology, Tokyo Medical
University between March 2017 and May 2018. This included 12
patients with endometrial adenocarcinoma who underwent total
hysterectomy or pelvic and para-aortic lymphadenectomy
simultaneously. Cancer tissue samples were collected during the
operation. Table I lists the
characteristics of cancer patients. Control endometrial specimens
were collected from 18 patients with benign gynecological diseases
who underwent hysterectomy. For the control endometrium, only
patients with proliferative phase of the menstrual cycle (also
recognized as the follicular phase in the ovarian cycle) were
enrolled since stromal cells differentially express KLF5 depending
on decidualization (15), resulting
in large variations in gene expression. There were no statistical
differences in age or body mass index between the cancer patients
and controls. After surgical removal, tissues were trimmed and
immediately stored at −80°C until further use for mRNA and protein
extraction. For morphological analyses, a part of the samples were
fixed in 4% paraformaldehyde and embedded in paraffin.
 | Table I.Characteristics of patients with
endometrial cancer. |
Table I.
Characteristics of patients with
endometrial cancer.
| Patient | Age, years | Body mass index | Histology | Stage (pT) | Lymph node metastasis
(pN) | Histological
grade |
|---|
| Case 1 | 40 | 17.4 | Endometrioid
adenocarcinoma with squamous differentiation | pT1a | N0 | G1 |
| Case 2 | 76 | 23.8 | Mixed endometrioid
undifferentiated carcinoma | pT1b | N0 | G3 |
| Case 3 | 58 | 25.0 | Endometrioid
adenocarcinoma | pT1a | N0 | G1 |
| Case 4 | 71 | 27.9 | Endometrioid
adenocarcinoma | pT1a | N0 | G1 |
| Case 5 | 53 | 34.9 | Endometrioid
adenocarcinoma with undifferentiated carcinoma | pT1a | N0 | G3 |
| Case 6 | 82 | 16.9 | Endometrioid
adenocarcinoma with undifferentiated carcinoma | pT1a | N0 | G1 |
| Case 7 | 52 | 19.5 | Endometrioid
adenocarcinoma | pT1a | N0 | G1 |
| Case 8 | 52 | 32.1 | Endometrioid
adenocarcinoma | pT1a | N1 | G1 |
| Case 9 | 66 | 26.1 | Endometrioid
adenocarcinoma | pT1a | N1 | G1 |
| Case 10 | 52 | 24.2 | Endometrioid
adenocarcinoma | pT2 | N1 | G1 |
| Case 11 | 57 | 25.6 | Endometrioid
adenocarcinoma | pT1a | NX | G1 |
| Case 12 | 58 | 23.0 | Endometrioid
adenocarcinoma | pT1b | N1 | G1 |
Cell culture
Human endometrial cancer cell lines, Ishikawa and
HEC1, were kindly provided by Dr Masato Nishida from the Department
of Obstetrics and Gynecology, National Kasumigaura Hospital
(Tsuchiura, Japan) and procured from the National Institute of
Health Sciences (Tokyo, Japan), respectively. Cells were cultured
in Dulbecco's modified Eagles medium (Thermo Fisher Scientific,
Inc.) containing 10% fetal bovine serum (Thermo Fisher Scientific,
Inc.), 100 U/ml penicillin, and 100 µg/ml streptomycin (Thermo
Fisher Scientific, Inc.) at 37°C in a 5% CO2 and 95% air
humidified incubator. Cells were periodically checked for
mycoplasma infection using MycoAlert Mycoplasma Detection Kit
(Lonza). They were cultured with primocin (Invivogen; Thermo Fisher
Scientific, Inc.) to inhibit mycoplasma infection during the
regular maintenance, but not during the experiments.
KLF5 knockdown
Table II shows the
sequence of two different shRNA sequences used to silence
KLF5. The pLKO.1 construct (Addgene, Inc.) with the shRNA
sequence and lentiviral packaging plasmids (Addgene, Inc.) were
used to produce lentiviral particles for the transduction of
shRNA-mediated knockdown system as previously reported (16). The pLKO.1-shSCR, containing scrambled
shRNA which is not expected to target any mRNA, was used as the
control (Addgene, Inc.). Cells were exposed to the lentiviral
particles and transduced cells were selected using 3 µg/ml
puromycin dihydrochloride (Nacalai Tesque) for two days. Samples
for mRNA and protein analyses were collected after an additional
day of culture without puromycin.
| Table II.Specific shRNA and primer
sequences. |
Table II.
Specific shRNA and primer
sequences.
| Target gene | Sequence
(5′-3′) |
|---|
| shRNAs |
| SCR
(control) |
CCTAAGGTTAAGTCGCCCTCG |
| KLF5
sh1 |
GCTGTAATGTATATGGCTTTA |
| KLF5
sh2 |
CCTATAATTCCAGAGCATAAA |
| Primers |
| 18S
rRNA-F |
GCAATTATTCCCCATGAACG |
| 18S
rRNA-R |
GGCCTCACTAAACCATCCAA |
|
KLF5-F |
AACGACGCATCCACTACTGC |
|
KLF5-R |
CAGCCTTCCCAGGTACACTT |
|
CCNA2-F |
CACCATTCATGTGGATGAAGC |
|
CCNA2-R |
ACACTCACTGGCTTTTCATCTT |
|
CCNB1-F |
GCAGCAGGAGCTTTTTGCTT |
|
CCNB1-R |
CCAGGTGCTGCATAACTGGA |
|
CCND1-F |
CAATGACCCCGCACGATTTC |
|
CCND1-R |
AAGTTGTTGGGGCTCCTCAG |
|
CCNE1-F |
CAGCCCCATCATGCCGAG |
|
CCNE1-R |
TTGTCAGGTGTGGGGATCAG |
mRNA expression analysis
Total RNA was extracted using ISOGEN (#311-02501;
Nippon Gene) and reverse-transcribed using Superscript III Reverse
Transcriptase (#1808044; Thermo Fisher Scientific, Inc.) according
to the manufacturer's instructions. Reverse
transcription-quantitative PCR (RT-qPCR) was performed using the
Power SYBR-Green Master Mix (#4368577; Thermo Fisher Scientific,
Inc.) and Applied Biosystems StepOnePlus™ Real-Time PCR system
(Thermo Fisher Scientific, Inc.). Data were normalized to the
endogenous control, 18S rRNA, and they were analyzed using the
2−∆∆Cq method (17).
Table II lists the primer sequences
used in RT-qPCR.
Western blotting
Proteins were extracted by Pierce RIPA buffer
(Thermo Fisher Scientific, Inc.) and the concentrations were
measured using Pierce BCA protein assay kit (Thermo Fisher
Scientific, Inc.). Thirty microgram each of proteins were, then,
denatured by NuPAGE™ LDS Sample Buffer, separated by
NuPAGE® Bis-Tris gel electrophoresis (4-12% gradient;
Thermo Fisher Scientific, Inc.), and transferred to polyvinylidene
fluoride membranes using the iBlot Dry Blotting System (Thermo
Fisher Scientific, Inc.). After blocking with 5% skim milk in TBST
for 1 h at room temperature, the membrane was incubated with
anti-KLF5 antibody (1:1,000 dilution, #ab137676; Abcam) at 4°C
overnight followed by horseradish peroxidase-conjugated secondary
antibody (#7074; Cell Signaling Technology) at room temperature for
2 h. Signal from the protein bands were visualized using the ECL
western blotting detection system (Bio-Rad Laboratories). β-actin
(1:10,000 dilution, #MB1051R; Merck) was used as the loading
control.
Immunohistochemistry
Paraffin-embedded samples were sectioned,
deparaffinized, antigens retrieved using heat-induced retrieval,
and incubated with the anti-KLF5 antibody (1:100 dilution). The
signal from the protein was visualized using the diaminobenzidine
staining kit (Nichirei Bioscience, Inc.). Hematoxylin was used to
stain the nuclei.
Cell proliferation and apoptosis
assays
Cell proliferation was measured by the MTT assay
using CellTiter 96® Non-Radioactive Cell Proliferation
Assay kit (Promega), and cell apoptosis was analyzed by TUNEL assay
using in situ cell death detection kit (Roche Diagnostics)
following the manufacturer's instructions. Cells, treated with
DNase I (100 U/ml; Sigma-Aldrich; Merck KGaA) to artificially
induce the DNA fragmentation, was used as a positive control for
TUNEL assay.
Statistical analysis
Data were represented as mean ± standard error of
the mean. Differences between groups were analyzed using the
unpaired Student's t-test and differences among three groups were
evaluated with ANOVA following Tukey's post hoc test by using SPSS
software. P<0.05 was considered statistically significant.
Results
KLF5 is upregulated in endometrial
cancer
Thirty patients, including 12 patients with
endometrial cancer and 18 with benign gynecological diseases
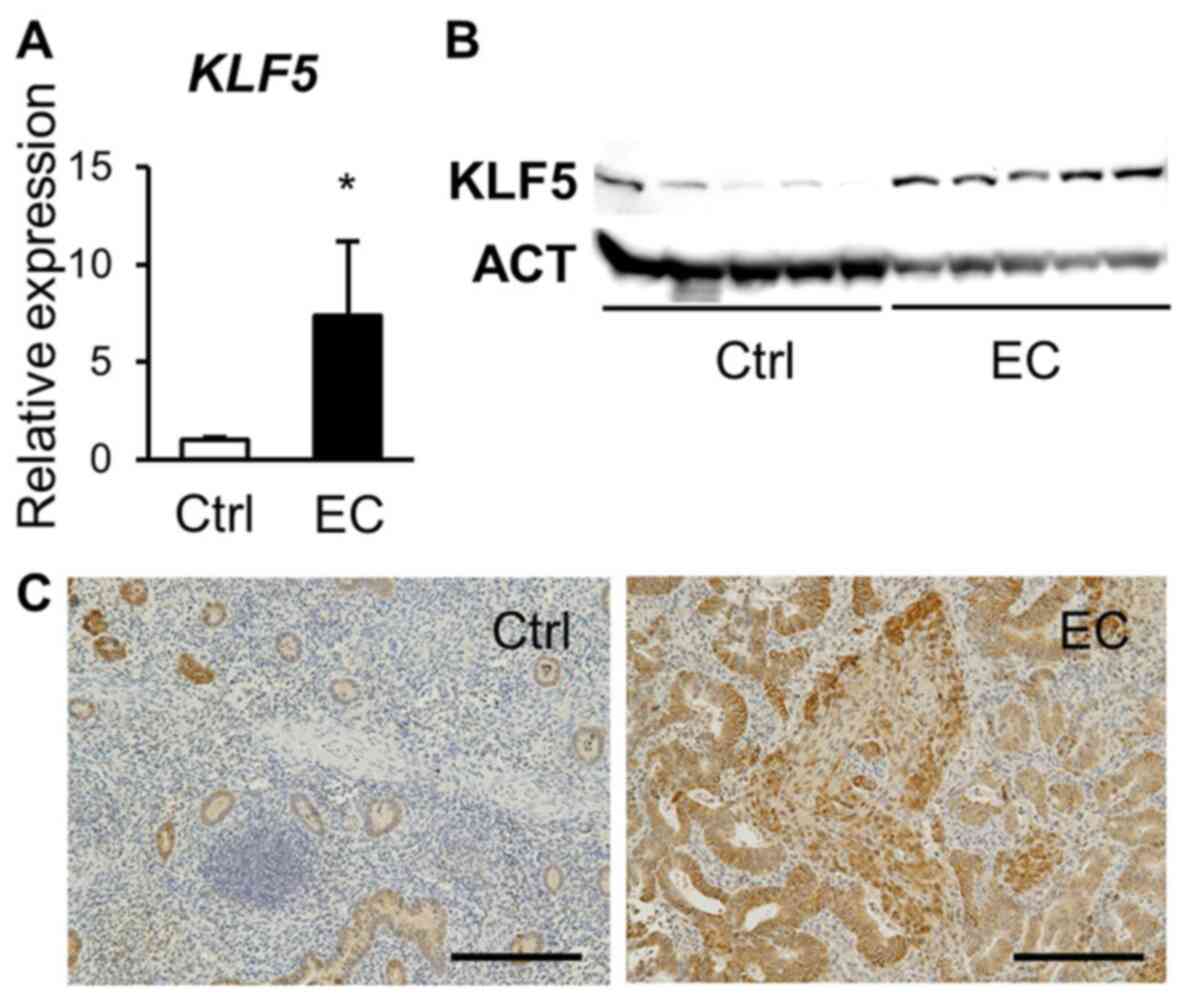
(controls), were enrolled. We used RT-qPCR and western blotting to
demonstrate increased mRNA and protein levels of KLF5,
respectively, in the endometrial cancer samples than those in the
control endometrium (Fig. 1A and B).
Immunohistochemistry showed moderate and faint expression in
glandular epithelial and stromal cells, respectively, in the
control endometrium, while a strong signal was observed in tumor
cells in the endometrial cancer specimens (Fig. 1C).
Silencing of KLF5 inhibits endometrial
cancer cell proliferation
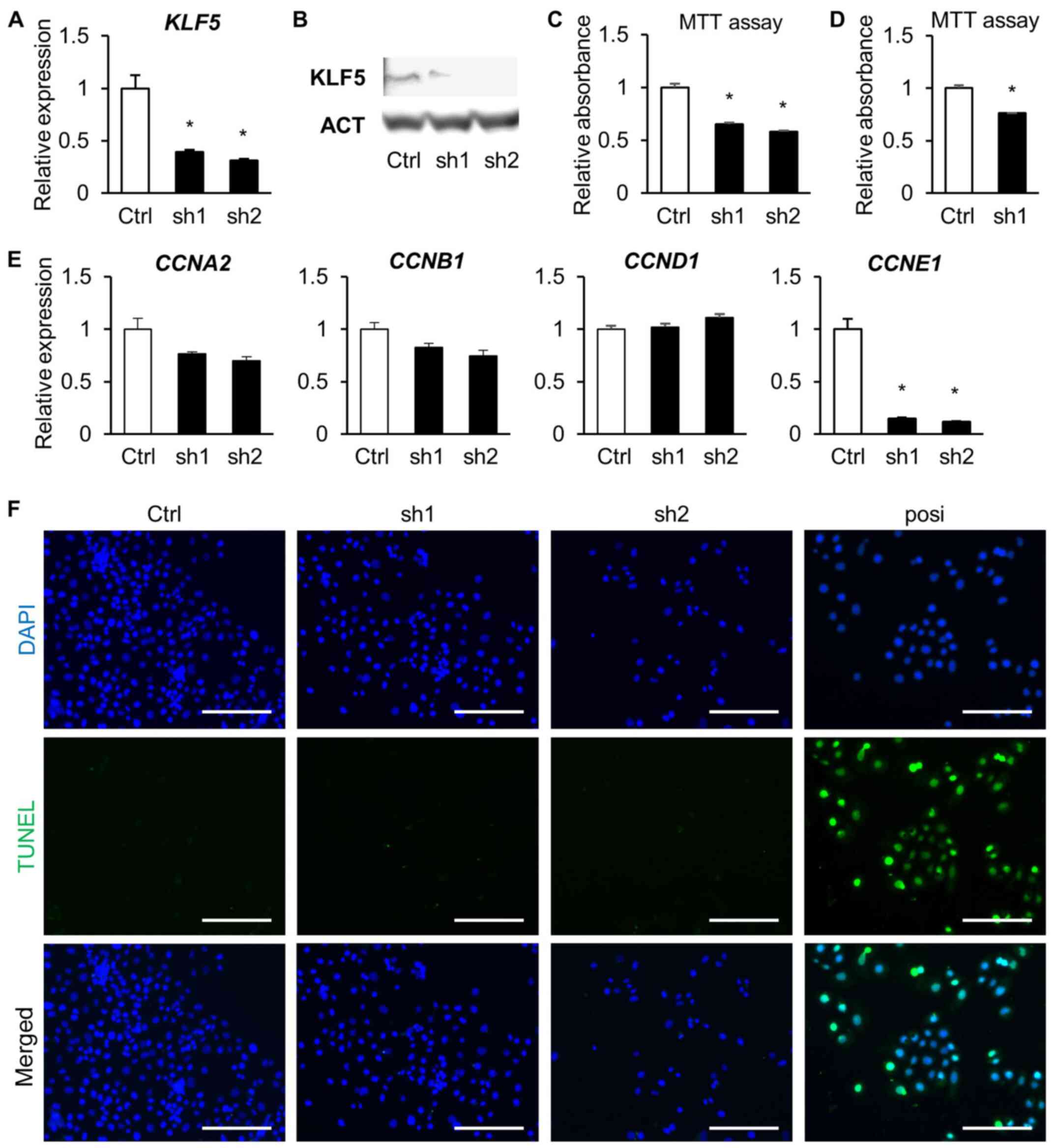
To investigate the role of KLF5 in endometrial
cancer, Ishikawa cells were transduced with the lentiviral
constructs bearing two different shRNAs targeting KLF5 and
subjected to loss-of-function assays. KLF5 knockdown efficiency was
successfully validated in the transduced Ishikawa cells (Fig. 2A and B). KLF5 expression is
upregulated in the cancer cells, and it possibly supports the
oncogenic processes, such as proliferation and apoptosis. The
results from the MTT assays revealed that depletion of KLF5 reduced
cell proliferation (Fig. 2C). To
confirm the role of KLF5 in cell proliferation, HEC1 cells were
transduced with the shRNA targeting KLF5 and subjected to MTT
assays. As observed in Ishikawa cells, KLF5 knockdown also
suppressed the proliferation of HEC1 cells (Fig. 2D).
Transactivation of the cyclins' expression by KLF5
is believed to enhance the cell proliferation (8). Among the cyclins (CCN), the expression
of CCNE1 was significantly downregulated in the KLF5-depleted cells
(Fig. 2E). To further validate the
role of KLF5 in tumorigenesis, we analyzed the cell survival
potential. Cell apoptosis was absent in the KLF5-depleted cells, as
seen from the TUNEL assays, indicating that the reduction in the
number of cells was not due to cell death (Fig. 2F).
Discussion
Endometrial cancer is the most common cancer of the
reproductive system of females in developed countries whose
incidence has been increasing worldwide (1,2). The
treatment options currently available for endometrial cancer are
limited: Surgical removal is preferentially recommended. However,
in cases of the patients with physiological complications or those
who are very old, radiation might be the next choice. The
pathogenesis and pathophysiology of endometrial cancer remains to
be understood fully. Exploring the molecular mechanism(s)
underlying tumorigenesis and metastasis of endometrial cancer will
help identify new therapeutic strategies for patients. Recent
studies have reported that KLF5 regulates cancer phenotypes,
including cellular proliferation, apoptosis, differentiation, and
motility (6–8). KLF5 has a dual role in being a tumor
suppressor or oncogene, depending on the cellular and genetic
environments (8). Thus, the role of
KLF5 differs and needs to be studied in each cancer type.
The molecular mechanism underlying the oncogenic
role of KLF5 has been clearly elucidated in colorectal cancer. More
than 50% of colon cancers exhibit activating mutations in the
oncogene K-RAS; this upregulates KLF5 and promotes cell
proliferation (9). KLF5 is also a
critical mediator of the oncogene H-RAS (18,19) and
tumor suppressor APC (20,21). Increases the expression of KLF5 is
associated with poor prognosis (22). In vitro knockdown experiments
have shown that KLF5 interacts with b-catenin and localizes to the
nucleus where the complex transactivates target genes, such as
CCND1 and c-MYC (20,22). Genome-wide association studies
identified the regulatory region of KLF5 as a risk locus for
endometrial cancer (23). However,
the expression and function of KLF5 in endometrial cancer remains
to be explored. This study showed the upregulation of KLF5 in
endometrial cancer, and the shRNA-mediated depletion of KLF5
inhibited Ishikawa and HEC1 cell proliferation. In addition, we
have demonstrated that the depletion of KLF5 downregulated the
expression of CCNE1, but not that of CCND1. It has been reported
that KLF5 binds to the first intron of CCNE1 and it regulates
transcription and cellular proliferation in bladder cancer
(24). Therefore, KLF5 promotes cell
proliferation by trans-activating a variety of cyclins depending on
the cancer type and cellular microenvironments. CCNE1 expression is
associated with the proliferative and aggressive potential, and it
is an indicator in the prognosis for endometrial cancer (25,26). The
role of CCNE1 within KLF5-mediated cell proliferation is not
apparent in this study; however, KLF5 regulates cell proliferation,
at least in part, through the action of CCNE1. Attempting to rescue
the KLF5 compromised cell proliferation with CCNE1 could be
informative. The exact mechanism of KLF5 regulation of target genes
remains to be explored. SUMOyration regulates KLF5 nuclear
localization, and phosphorylation enhances KLF5 transactivation
(27,28). The analyses of nuclear localization,
protein activation, and/or promoter activity could further confirm
the role of KLF5 in transcriptional regulation.
KLF5 has also been reported to modulate cell
survival by inhibiting apoptosis. KLF5 binds to the core promoter
region and induces the expression of the anti-apoptotic protein
survivin in leukemia (29). In colon
cancer HCT116 cells, KLF5 induces the expression of the survival
kinase Pim1 that phosphorylates and inactivates the pro-apoptotic
protein BAD (30). In this study,
however, silencing KLF5 decreased cell numbers but did not induce
apoptosis in endometrial cancer cells, suggesting that KLF5 acts as
an oncogene in endometrial cancer by directly promoting cell
proliferation rather than inducing cell survival.
This study has some limitations. First, we used a
small sample size. This led to a large variation in the expression
of KLF5 in the endometrial cancer samples. This could also be
attributed to the stage-dependent change in KLF5 expression in
non-tumor cells. KLF5 is expressed in normal endometrial epithelial
cells (14) and stromal cells,
especially after decidualization (15). Therefore, the proportion of tumor
cells against normal cells generates a difference in gene
expression. Nevertheless, we found a statistical difference in KLF5
expression, especially protein levels, in the endometrial cancer
specimens. KLF5 undergoes regulation at the transcriptional,
post-transcriptional, and post-translational stages, including its
protein stability induced by differential proteasomal degradation
(31–34). This could have resulted in the clear
and strong signals of KLF5 protein in the endometrial cancer
specimens as analyzed by western blotting and immunohistochemistry.
However, a large-scale study of KLF5 expression in the endometrial
cancer, and its correlation with the treatment outcomes and
long-time prognosis could generate valuable information. The second
limitation is that only early phase cancer patients were enrolled
in this study. KLF5 expression might vary across the different
stages and grades of cancer progression. We have focused on the
proliferative role of KLF5 and, therefore, investigated the early
phase tumor. The KLF5 expression in aggressive tumors as well as
its function in tumor metastasis is of interest for the next
studies.
In conclusion, we have demonstrated the upregulation
of KLF5 in endometrial cancer. Furthermore, KLF5 regulates cell
proliferation, at least in part, through the action of CCNE1.
Future experiments should also focus on the role of KLF5 in
epithelial-mesenchymal transition and/or metastasis in endometrial
cancer. Large-scale and randomized clinical studies will further
help validate the potential of KLF5 as a therapeutic target for
endometrial cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Japan Society
for the Promotion of Science (grant no. 19K09835) and International
Joint Usage/Research Center, Institute of Medical Science,
University of Tokyo.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
TM, JK and KK wrote the manuscript, analyzed the
data and performed the experiments. TM and JK reviewed the raw data
and confirmed its authenticity. TM, JK, FT, KI and HN collected the
samples. KK, FT, KI and HN designed the study. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Tokyo Medical University (Tokyo, Japan). Written
informed consent was obtained from the patients or guardians.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Felix AS and Brinton LA: Cancer progress
and priorities: Uterine cancer. Cancer Epidemiol Biomarkers Prev.
27:985–994. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Morice P, Leary A, Creutzberg C,
Abu-Rustum N and Darai E: Endometrial cancer. Lancet.
387:1094–1108. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sheikh MA, Althouse AD, Freese KE, Soisson
S, Edwards RP, Welburn S, Sukumvanich P, Comerci J, Kelley J,
LaPorte RE and Linkov F: USA endometrial cancer projections to
2030: Should we be concerned? Future Oncol. 10:2561–2568. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
McAlpine JN, Temkin SM and Mackay HJ:
Endometrial cancer: Not your grandmother's cancer. Cancer.
122:2787–2798. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Simmen RC, Pabona JM, Velarde MC, Simmons
C, Rahal O and Simmen FA: The emerging role of Krüppel-like factors
in endocrine-responsive cancers of female reproductive tissues. J
Endocrinol. 204:223–231. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Simmen RCM, Heard ME, Simmen AM, Montales
MT, Marji M, Scanlon S and Pabona JM: The Krüppel-like factors in
female reproductive system pathologies. J Mol Endocrinol.
54:R89–R101. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Diakiw SM, D'Andrea RJ and Brown AL: The
double life of KLF5: Opposing roles in regulation of
gene-expression, cellular function, and transformation. IUBMB Life.
65:999–1011. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nandan MO, McConnell BB, Ghaleb AM,
Bialkowska AB, Sheng H, Shao J, Babbin BA, Robine S and Yang VW:
Krüppel-like factor 5 mediates cellular transformation during
oncogenic KRAS-induced intestinal tumorigenesis. Gastroenterology.
134:120–130. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Marrero-Rodríguez D, Taniguchi-Ponciano K,
Jimenez-Vega F, Romero-Morelos P, Mendoza-Rodríguez M, Mantilla A,
Rodriguez-Esquivel M, Hernandez D, Hernandez A, Gomez-Gutierrez G,
et al: Krüppel-like factor 5 as potential molecular marker in
cervical cancer and the KLF family profile expression. Tumor Biol.
35:11399–11407. 2014. View Article : Google Scholar
|
|
11
|
Zheng HQ, Zhou Z, Huang J, Chaudhury L,
Dong JT and Chen C: Krüppel-like factor 5 promotes breast cell
proliferation partially through upregulating the transcription of
fibroblast growth factor binding protein 1. Oncogene. 28:3702–3713.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Meyer SE, Hasenstein JR, Baktula A, Velu
CS, Xu Y, Wan H, Whitsett JA, Gilks CB and Grimes HL: Kruppel-like
factor 5 is not required for K-RasG12D lung tumorigenesis, but
represses ABCG2 expression and is associated with better
disease-specific survival. Am J Pathol. 177:1503–1513. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen C, Bhalala HV, Vessella RL and Dong
JT: KLF5 is frequently deleted and down-regulated but rarely
mutated in prostate cancer. Prostate. 55:81–88. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shi H, Zhang Z, Wang X, Liu S and Teng CT:
Isolation and characterization of a gene encoding human
Kruppel-like factor 5 (IKLF): Binding to the CAAT/GT box of the
mouse lactoferrin gene promoter. Nucleic Acids Res. 27:4807–4815.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sun X, Zhang L, Xie H, Wan H, Magella B,
Whitsett JA and Dey SK: Kruppel-like factor 5 (KLF5) is critical
for conferring uterine receptivity to implantation. Proc Natl Acad
Sci USA. 109:1145–1150. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Stewart SA, Dykxhoorn DM, Palliser D,
Mizuno H, Yu EY, An DS, Sabatini DM, Chen ISY, Hahn WC, Sharp AP,
et al: Lentivirus-delivered stable gene silencing by RNAi in
primary cells. RNA. 9:493–501. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nandan MO, Chanchevalap S, Dalton WB and
Yang VW: Krüppel-like factor 5 promotes mitosis by activating the
cyclin B1/Cdc2 complex during oncogenic Ras-mediated
transformation. FEBS Lett. 579:4757–4762. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Nandan MO, Yoon HS, Zhao W, Ouko LA,
Chanchevalap S and Yang VW: Krüppel-like factor 5 mediates the
transforming activity of oncogenic H-Ras. Oncogene. 23:3404–3413.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
McConnell BB, Bialkowska AB, Nandan MO,
Ghaleb AM, Gordon FJ and Yang VW: Haploinsufficiency of
Krüppel-like factor 5 rescues the tumor-initiating effect of the
Apc(Min) mutation in the intestine. Cancer Res. 69:4125–4133. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nandan MO, Ghaleb AM, McConnell BB, Patel
NV, Robine S and Yang VW: Krüppel-like factor 5 is a crucial
mediator of intestinal tumorigenesis in mice harboring combined
ApcMin and KRASV12 mutations. Mol Cancer. 9:632010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Takagi Y, Sakai N, Yoshitomi H, Furukawa
K, Takayashiki T, Kuboki S, Takano S, Suzuki D, Kagawa S, Mishimi
T, et al: High expression of Krüppel-like factor 5 is associated
with poor prognosis in patients with colorectal cancer. Cancer Sci.
111:2078–2092. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cheng TH, Thompson DJ, O'Mara TA, Painter
JN, Glubb DM, Flach S, Lewis A, French JD, Freeman-Mills L, Church
D, et al: Five endometrial cancer risk loci identified through
genome-wide association analysis. Nat Genet. 48:667–674. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pattison JM, Posternak V and Cole MD:
Transcription factor KLF5 binds a cyclin E1 polymorphic intronic
enhancer to confer increased bladder cancer risk. Mol Cancer Res.
14:1078–1086. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nakayama K, Rahman MT, Rahman M, Nakamura
K, Ishikawa M, Katagiri H, Sato E, Ishibashi T, Iida K, Ishikawa N
and Kyo S: CCNE1 amplification is associated with aggressive
potential in endometrioid endometrial carcinomas. Int J Oncol.
48:506–516. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Santala S, Talvensaari-Mattila A, Soini Y
and Santala M: Cyclin E expression correlates with cancer-specific
survival in endometrial endometrioid adenocarcinoma. Anticancer
Res. 35:3393–3397. 2015.PubMed/NCBI
|
|
27
|
Du JX, Bialkowska AB, McConnell BB and
Yang VW: SUMOylation regulates nuclear localization of Krüppel-like
factor 5. J Biol Chem. 283:31991–32002. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang Z and Teng CT: Phosphorylation of
Kruppel-like factor 5 (KLF5/IKLF) at the CBP interaction region
enhances its transactivation function. Nucleic Acids Res.
31:2196–2208. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhu N, Gu L, Findley HW, Chen C, Dong JT,
Yang L and Zhou M: KLF5 interacts with p53 in regulating survivin
expression in acute lymphoblastic leukemia. J Biol Chem.
281:14711–14718. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhao Y, Hamza MS, Leong HS, Lim CB, Pan
YF, Cheung E, Soo KC and Iyer NG: Kruppel-like factor 5 modulates
p53-independent apoptosis through Pim1 survival kinase in cancer
cells. Oncogene. 27:1–8. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen C, Zhou Z, Guo P and Dong JT:
Proteasomal degradation of the KLF5 transcription factor through a
ubiquitin-independent pathway. FEBS Lett. 581:1124–1130. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu N, Li H, Li S, Shen M, Xiao N, Chen Y,
Wang Y, Wang W, Wang R, Wang Q, et al: The Fbw7/human CDC4 tumor
suppressor targets proproliferative factor KLF5 for ubiquitination
and degradation through multiple phosphodegron motifs. J Biol Chem.
285:18858–18867. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhao D, Zheng HQ, Zhou Z and Chen C: The
Fbw7 tumor suppressor targets KLF5 for ubiquitin-mediated
degradation and suppresses breast cell proliferation. Cancer Res.
70:4728–4738. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhao D, Zhi X, Zhou Z and Chen C: TAZ
antagonizes the WWP1-mediated KLF5 degradation and promotes breast
cell proliferation and tumorigenesis. Carcinogenesis. 33:59–67.
2012. View Article : Google Scholar : PubMed/NCBI
|