Introduction
Lung cancer (LC), the most common type of cancer in
the world, is a global burden and a public health issue due to its
high fatality rates. In the USA in 2018, it constituted ~13 and 14%
of newly detected cancer cases in men and women, respectively
(1). In China, the age-standardized
incidence rate of LC was 36.71 per 100,000 individuals in 2014,
with men exhibiting a higher incidence than women (2). Non-small cell LC (NSCLC) accounts for
85% of all LC cases, with an overall 5-year survival rate of 16%.
Treatment mainly includes surgical resection, targeted therapy,
chemotherapy, radiotherapy and immunotherapy, although use of
personalized medicine has been speculated upon (3). It is estimated that 30–55% of patients
with NSCLC experience relapse in spite of curative resection,
leading to subsequent mortality (4).
Cancer chemotherapy resistance, be it innate or
acquired, can be a hindrance during the treatment phase; it occurs
due to a variety of reasons, including amplified drug target
molecules, decreased drug accumulation and enhanced drug export,
alterations in drug metabolism by modifying signalling transduction
molecules, DNA damage and evasion of apoptosis (5). Avoidance of apoptosis by upregulation
of anti-apoptotic proteins, such as Bcl-2, and inactivation of
pro-apoptotic proteins, such as Bax, is commonly seen in cancer
cells, leading to a lack of response to chemotherapy (6). Overexpression of P-glycoprotein 1
(P-gp), leading to the transport of anticancer drugs out of the
cancer cells, has been shown to confer resistance to chemotherapy
in NSCLC (7). While conventional
radiotherapy and stereotactic body radiation therapy are used for
inoperable cases, iodine-125 seed brachytherapy has been proven to
be a safe and effective technique, with improved median overall
survival time (16 vs. 10 months) and quality of life (8–10). It
has been shown that the combination of iodine-125 brachytherapy and
chemotherapy is better than chemotherapy alone, with improved
therapeutic efficacy for NSCLC (11,12).
Although ionizing radiation is commonly used for LC
treatment, reports suggest that it might have secondary undesirable
effects such as promoting cancer malignancy and increasing the
incidence of cardiac events, among others (13,14).
Therefore, it is desirable to find a safe radionuclide sensitizer
to reduce the side effects of radiotherapy without influencing the
antitumour efficacy in clinical practice.
Gambogic acid (GA), extracted from the gamboge resin
of Garcinia hanburyi, has been used in traditional Chinese
medicine to treat infections and tumours; it exerts antitumour,
antiangiogenic and antimetastatic activities in several types of
cancer, including LC (15–17). In addition, conventional doses do not
influence the functions of other cells in the body (18,19). GA
exerts its anticancer activities via numerous targets and
signalling pathways, promoting apoptosis, inducing cell cycle
arrest, inhibiting angiogenesis and activating lymphocytes
(20). GA in low doses causes
autophagy in mutant p53 (mtp53) degradation in cancer cells
(21) and protein levels of mtp53
are decreased following GA exposure (22).
Our previous results showed that a combination of
low-dose GA and NaI131 was able to significantly inhibit
cell proliferation, as well as induce cell apoptosis, in A549/DDP
cells (23). Therefore, further
study was required on the plausible cellular mechanisms related to
the effect of the combination therapy on two drug-resistant LC cell
lines. Additionally, exploration to see if the combination therapy
conferred inhibition to therapy resistance in NSCLC, so as to
achieve better therapeutic results, was required. The current in
vitro study aimed to elucidate whether low-dose GA could
sensitize NaI131 to enhance its influence on cell cycle
regulation, apoptosis and drug resistance in two NSCLC cell lines,
namely A549/Taxol and A549/cisplatin (DDP) cells, that are
resistant to Taxol and DDP, respectively.
Materials and methods
Cell culture
The human NSCLC A549 cell line was a gift from the
Laboratory of Pathology at the School of Medicine of Southeast
University (Nanjing, China). The DDP-resistant A549/DDP cell line
and the Taxol-resistant cell line A549/Taxol were procured from
Nanjing KeyGen Biotech Co., Ltd.. Cells were cultured in RPMI-1640
medium supplemented with 10% FBS (Shanghai ExCell Biology, Inc.)
and incubated in a humidified 5% CO2 incubator at 37°C.
When the cells reached a confluency of 80–90%, the original culture
medium was discarded, the cells were treated with trypsin (0.25%)
for 1–2 min, and the digestion was terminated by adding an equal
volume of serum-containing culture medium. Cells were dispersed by
gentle pipetting, and the cell suspension was transferred to a
15-ml centrifuge tube, after which the suspension was centrifuged
at 200 × g at 4°C for 5 min. The supernatant was discarded, and 1–2
ml of culture medium was added to resuspend the cells. The cells
were then transferred to a culture flask for continuous
culture.
Experimental groups and drug
intervention
A549, A549/DDP and A549/Taxol cells were harvested
in the logarithmic phase of growth. A total of 5×104
cells per well were seeded in 6-well plates and cultured at 37°C,
with 5% CO2, for 24 h and then divided into the
NaI131, GA, combination and control groups. The
NaI131 group was treated with 10.3 MBq
NaI131, the GA group with 2.9 µg/ml GA, and the
combination group with 10.3 MBq NaI131 and 0.3 µg/ml GA,
as described previously (23). The
control group was treated with PBS (2 ml per well). The cells were
harvested after 48 h of drug treatment.
Cell cycle assay
Cells were fixed in 70% alcohol for 2 h. RNase A
(100 µl) was added to the cells for 30 min, followed by the
addition of 400 µl propidium iodide stain (Nanjing KeyGen Biotech
Co., Ltd.) and incubation at 4°C in the dark for 30 min. Cellular
DNA content was measured using a flow cytometer with
fluorescence-activated cell sorting (FACS) (FACSCalibur; Becton,
Dickinson and Company). The cell cycle profiles were analysed using
FACSDiva software (version 7.0; Becton, Dickinson and Company).
Cell apoptosis assay
An Annexin V-APC/7-AAD cell apoptosis detection kit
(cat. no. KGA1024; Nanjing KeyGen Biotech Co., Ltd.) was used for
the detection of apoptosis according to the manufacturer's
instructions. After drug treatment, cells were mixed with 500 µl
binding buffer, 5 µl Annexin V-APC and 5 µl 7-AAD, and incubated at
room temperature in the dark for 5–15 min. Cell apoptosis was
measured using a FACSCalibur flow cytometer (Becton, Dickinson and
Company). Data were analysed using FACSDiva software (version 7.0;
Becton, Dickinson and Company).
Immunofluorescence (IF)
Cells were air-dried on a glass slide and fixed in
4% paraformaldehyde at room temperature for 30 min. Slides were
rinsed three times in PBS, followed by blocking with 50–100 µl goat
serum (1:10; cat. no. C0265; Beyotime Institute of Biotechnology)
and incubation at room temperature for 20 min. Next, 50–100 µl
primary antibodies, namely, anti-mtp53 (1:200; cat. no. ab32509;
Abcam) or anti-heat shock protein 90 (HSP90) (1:200; sc-69703;
Santa Cruz Biotechnology, Inc.), was added and incubated at 37°C
for 2 h under humidified conditions. Slides were rinsed three times
in PBS, followed by addition of 50–100 µl horseradish peroxidase
(HRP)-conjugated anti-mouse or anti-rabbit secondary antibody
(1:200; cat. no. KGAA35 or KGAA37; Nanjing KeyGen Biotech Co.,
Ltd.) and incubated at 37°C for 1 h at room temperature. After
rinsing the slides three times in PBS, 50–100 µl DAPI staining
solution (Nanjing KeyGen Biotech Co., Ltd.) was added and
incubation occurred at room temperature for 5 min in the dark.
Sections were mounted using anti-fade mounting medium (Sangon
Biotech, Co., Ltd.). Protein expression in cells was observed under
a fluorescence microscope (BX61; Olympus Corporation). Images of
five areas that exhibited strong signals were captured and then
analysed using ImageJ software (version 1.52; National Institutes
of Health).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). cDNA synthesis was performed using a RevertAid First Strand
cDNA Synthesis kit (cat. no. K1622; Thermo Fisher Scientific, Inc.)
following the manufacturer's instructions. Specific mRNA
quantification was performed by PCR using the Thunderbird
SYBR® qPCR mix (cat. no. QPS-201; Toyobo Life Science)
in an ABI Step One Plus Real Time-PCR system (Applied Biosystems;
Thermo Fisher Scientific, Inc.) using the following reaction
conditions: Holding stage at 95°C for 5 min; cycling stage at 95°C
for l5 sec, 6°C for 20 sec and 72°C for 40 sec, for a total of 40
cycles; and melting curve stage at 95°C for 15 sec, 60°C for 1 min
and 95°C for 15 sec. The 2−∆∆Cq method (24) was used to calculate the relative mRNA
expression of the target genes. The primers used were as follows:
p53 forward, 5′-CCGCAGTCAGATCCTAGCG-3′ and reverse,
5′AATCATCCATTGCTTGGGACG-3′; HSP90 forward,
5′-CATAACGATGATGAGCAGTACGC-3′ and reverse,
5′-GACCCATAGGTTCACCTGTGT-3′; and GAPDH forward,
5′-CATCTTCTTTTGCGTCGCCA-3′ and reverse,
5′-TTAAAAGCAGCCCTGGTGACC-3′. The experiment was repeated 3
times.
Western blot analysis
Total cellular protein was extracted using RIPA
lysis buffer (cat. no. P0013B; Beyotime Institute of
Biotechnology), and the protein concentration was determined using
the bicinchoninic acid method. Proteins (20 µg per lane) separated
on a 12% gel by SDS-PAGE were transferred onto a nitrocellulose
membrane and blocked with 5% skimmed milk in Tris-buffered saline
containing 0.1% Tween-20 (TBST) for 2 h at room temperature,
followed by addition of primary antibodies as follows: Anti-CDK1
(1:300; cat. no. ab131450), anti-cyclin B (1:300; cat. no.
ab172317), anti-Bcl-2 (1:300; cat. no. ab692), anti-Bax (1:300;
cat. no. ab77566) (all Abcam) or anti-P-gp (1:300; cat. no. PB0162;
Wuhan Boster Biological Technology, Ltd.). Incubation occurred
overnight at 4°C. The following day, the membrane was washed three
times with 5% TBST for 10 min each. HRP-labelled goat anti-mouse
IgG secondary antibody (1:5,000; cat. no. KGAA35; Nanjing KeyGen
Biotech Co., Ltd.) was added, and the membrane was incubated at
room temperature for 30 min. After the membrane was washed 3 times
with 5% TBST for 10 min, protein bands were visualized using the
ECL Chemiluminescence kit (cat. no. KGP116; Nanjing KeyGen Biotech
Co., Ltd.). Gel-Pro32 software (Media Cybernetics) was used to
analyse the band intensities.
Statistical analysis
Statistical analyses of the data were performed
using SPSS software (version 22.0; IBM Corp.). All data are
presented as the mean ± SD. Comparisons among multiple groups were
performed by one-way analysis of variance, followed by Bonferroni's
post hoc test. P<0.05 was used to indicate a statistically
significant difference.
Results
Analysis of stages in the cell
cycle
First, the combination treatment was assessed with
regard to any effect it may have on the cell cycle, since our
previous study showed that treatment of A549/DDP cells with
low-dose GA together with NaI131 was able to
significantly inhibit cell proliferation (23). NaI131, GA and a
combination of both were used to treat A549, A549/DDP and
A549/Taxol cells, while the control group was treated with PBS. The
cell cycle conditions in all groups were observed after 48 h
(Fig. 1A and B). The data showed
that in the A549, A549/DDP and A549/Taxol cells, the percentage of
cells in the G2/M phase was considerably increased in
the NaI131, GA and combination groups compared with that
in the control group. The increase in the combination group was
significantly higher than that in the NaI131 and GA
groups (P<0.001). Moreover, the highest percentage of cells in
the G2/M phase was found in A549/Taxol cells.
Additionally, the percentage of cells among these groups in the
G0/G1 and S phases was not significantly different.
These results suggested that G2/M arrest occurred in the
cells in all the groups, but was most pronounced in the combination
group.
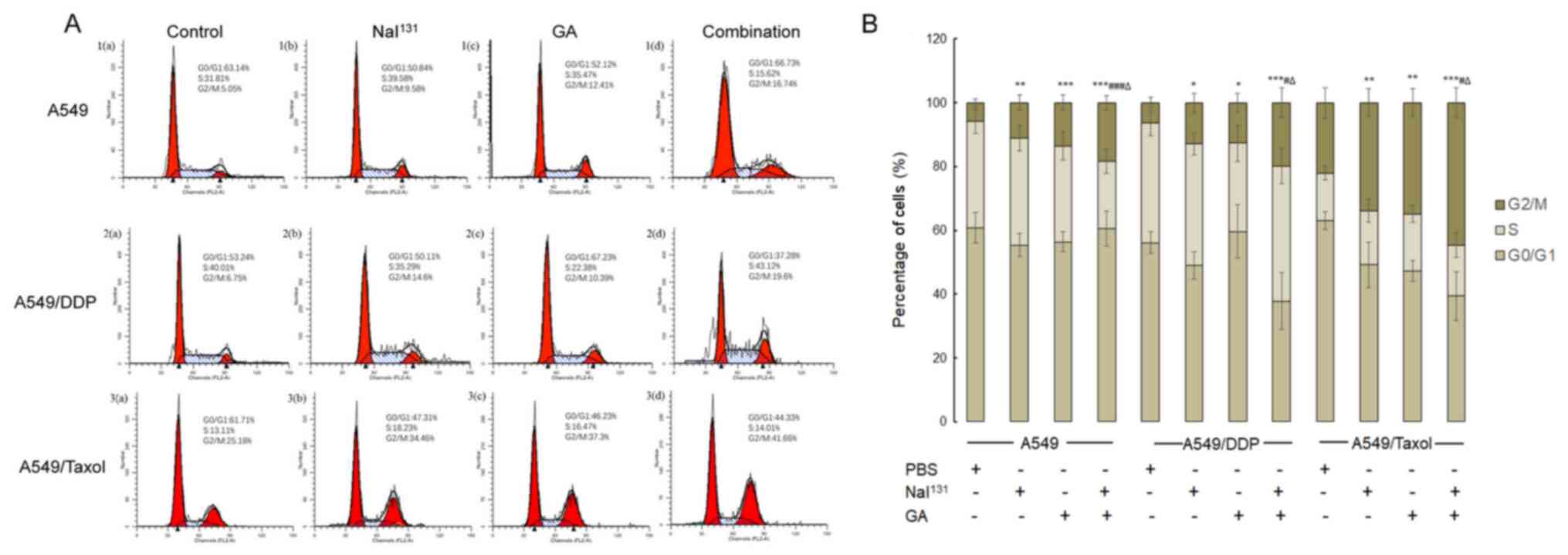 | Figure 1.Analysis of stages in the cell cycle.
(A) A549, A549/DDP and A549/Taxol cells were treated with PBS
(control), NaI131, GA or the combination of both, for 48
h, and the stages of the cell cycle in each group were detected:
The first peak represents the G0/G1 phase,
the second peak represents the G2/M phase and the
plateau represents the S phase. (B) Graphical representation of the
results. * refers to comparisons with the control group;
# refers to comparisons with the NaI131 group
and ∆ refers to comparisons with the GA group.
*,#,∆P<0.05; **P<0.01; and
***,###P<0.001. DDP, cisplatin; GA, gambogic acid;
PBS, phosphate-buffered saline. |
Analysis of cell apoptosis
An Annexin V-APC/7-AAD assay was used to revalidate
the effect of low-dose GA combined with NaI131 on cell
apoptosis. NaI131 and GA both induced apoptosis in the
A549, A549/DDP and A549/Taxol cells. The results after 48 h of
incubation are shown in Fig. 2A and
B. Compared with that in the control group, the percentage of
total apoptotic cells in the NaI131, GA and combination
groups increased significantly, while the percentage of apoptotic
cells was almost identical in the NaI131 and GA groups,
with 40–50% cell death. Cell apoptosis was most evident in the
combination group (P<0.001); late-stage apoptotic activity was
highest in this group.
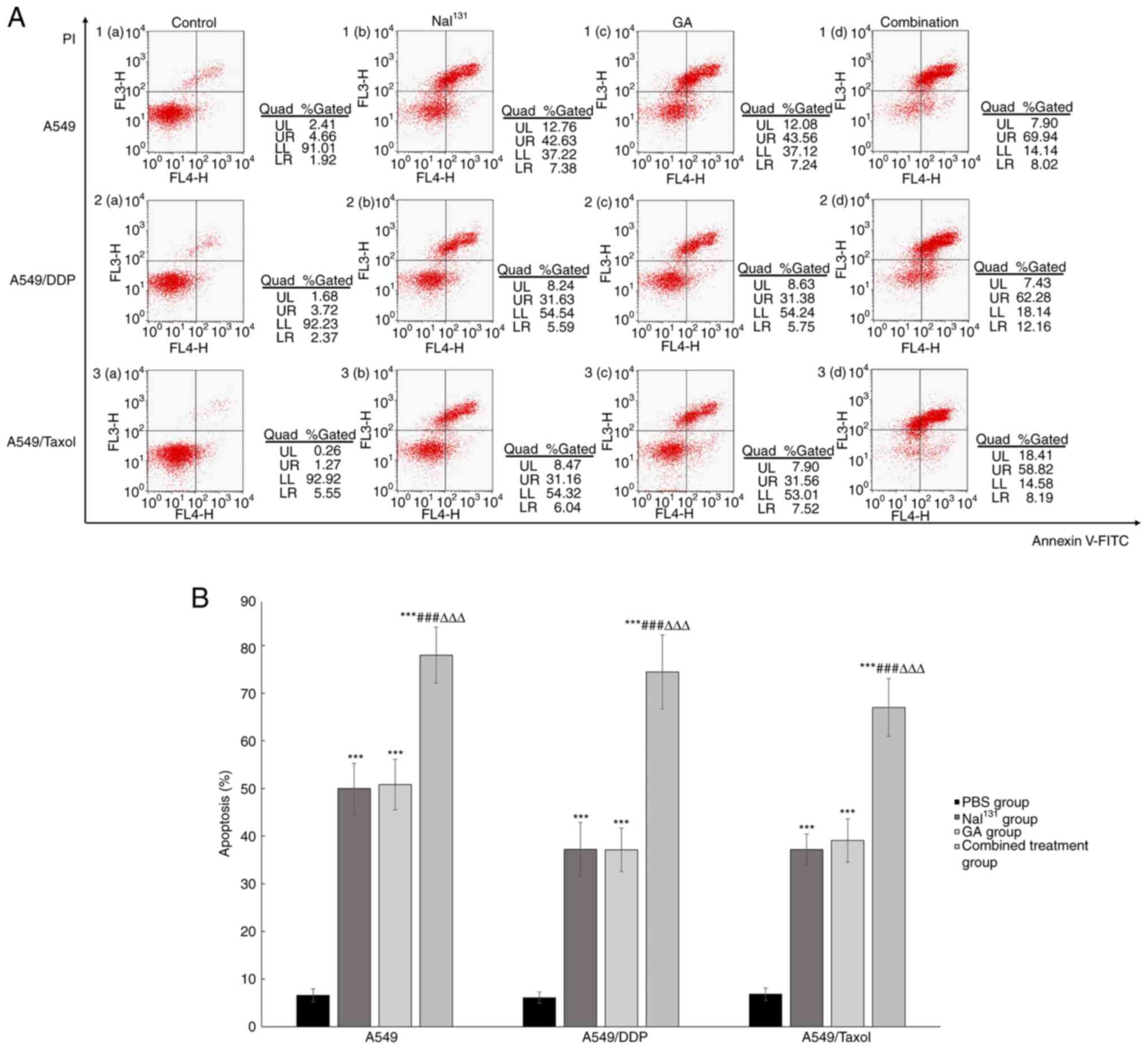 | Figure 2.Analysis of cell apoptosis. (A) A549,
A549/DDP and A549/Taxol cells were incubated with PBS (control),
NaI131, GA or the combination of both, for 48 h, and the
percentages of cells at all stages of apoptosis were detected by
flow cytometry. The upper right quadrant suggests the number of
apoptotic cells in the late stage, while the lower right quadrant
suggests the number of apoptotic cells in the early stage. (B)
Graphical representation of the results. * refers to comparisons
with the control group; # refers to comparisons with the
NaI131 group and ∆ refers to comparisons with
the GA group. ***,###,∆∆∆P<0.001. DDP, cisplatin; GA,
gambogic acid; PBS, phosphate-buffered saline. |
Expression levels of P-gp and proteins
related to cell cycle control and apoptosis
To better understand the effect of combination
treatment on drug-resistant NSCLC cells, levels of proteins related
to the cell cycle, apoptosis and drug resistance, namely CDK1,
cyclin B, Bcl-2, Bax and P-gp, were measured using western blotting
(Fig. 3A). The expression levels of
CDK1, cyclin B, Bcl-2 and P-gp were decreased in the
NaI131, GA and combination groups compared with those in
the control group. The expression levels were the lowest in the
combination group in comparison to all other groups (P<0.001)
(Fig. 3B-D and F). By contrast, the
expression levels of Bax were elevated in all the groups compared
with that in the control, but the increase was most evident in the
combination group (P<0.001) (Fig.
3E).
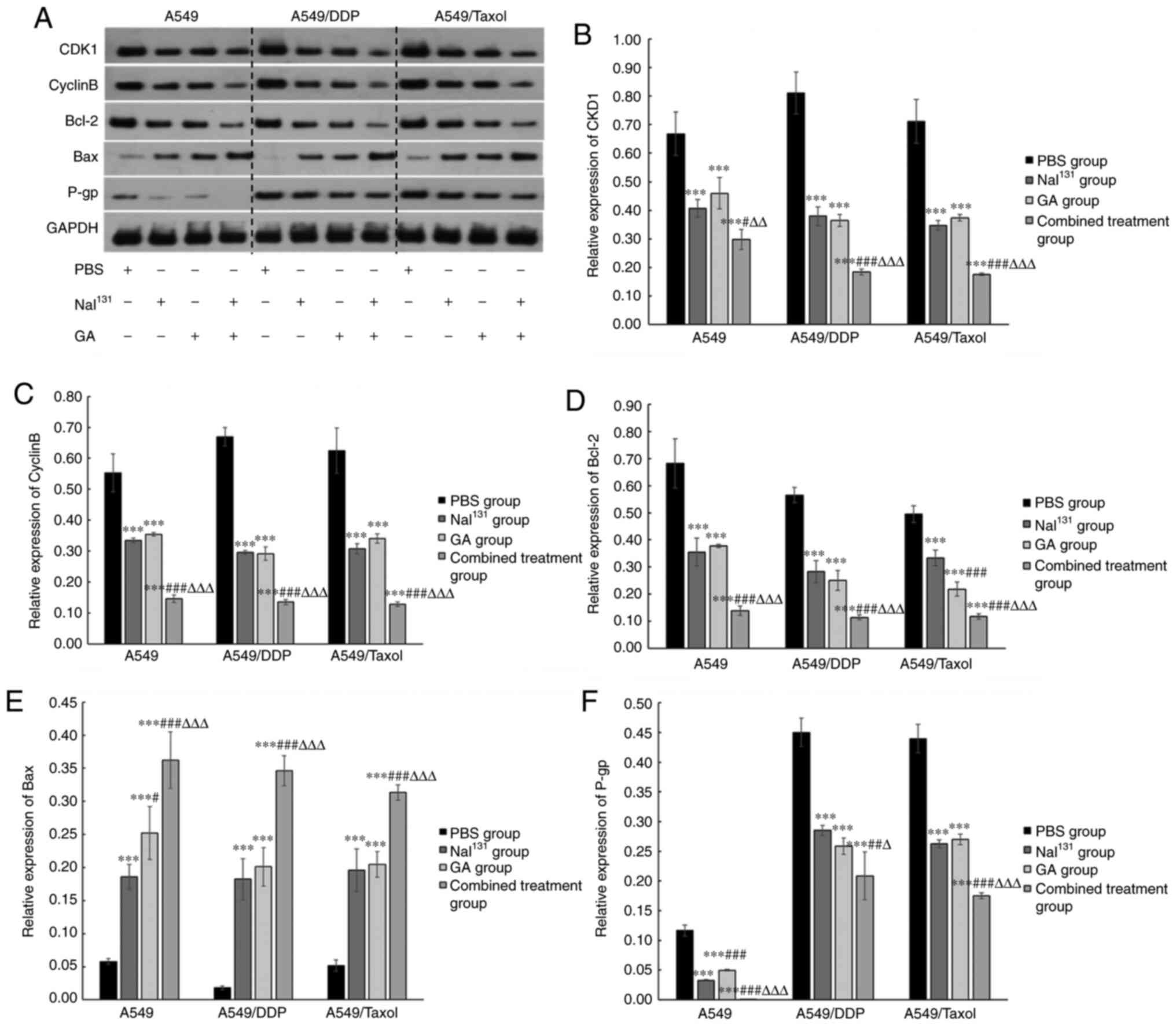 | Figure 3.Protein expression levels of P-gp and
associations with cell cycle control and apoptosis. After treatment
of A549, A549/DDP and A549/Taxol cells with NaI131, GA
or the combination of both, for 48 h, the expression of
intracellular CDK1, cyclin B, Bcl-2, Bax and P-gp protein in all
groups was detected. (A) Western blots showing expression levels of
CDK1, cyclin B, Bcl-2, Bax and P-gp protein following treatment.
Graphical representation of the results for (B) CDK1, (C) cyclin B,
(D) Bcl-2, (E) Bax and (F) P-gp. * refers to comparisons with the
control group; # refers to comparisons with the
NaI131 group and ∆ refers to comparisons with
the GA group. #,∆P<0.05; ##,∆∆P<0.01;
and ***,###,∆∆∆P<0.001. DDP, cisplatin; GA, gambogic
acid; PBS, phosphate-buffered saline; P-gp, P-glycoprotein. |
Expression of mRNA and protein levels
of p53 and HSP90
The present study observed the expression and
distribution of mtp53 protein in cells by IF, and indirectly
indicated whether mtp53 is consistent with its protein expression
by detecting the level of total p53 mRNA. The study also assessed
whether the changes in the cell cycle and apoptosis brought about
by the combination treatment on drug-resistant NSCLC cells was
influenced by p53. Results of RT-qPCR and IF demonstrated that the
expression levels of p53 decreased in all the treatment groups
compared with those in the control (Fig.
4A-C). The lowest signal was detected in the combination group
(P<0.05). While the protein expression level of mtp53 was
similar in A549 and A549/DDP cells treated with NaI131
or GA, it decreased slightly in A549/Taxol cells treated with GA
compared with that in control and NaI131-treated cells
(P<0.05; Fig. 4C).
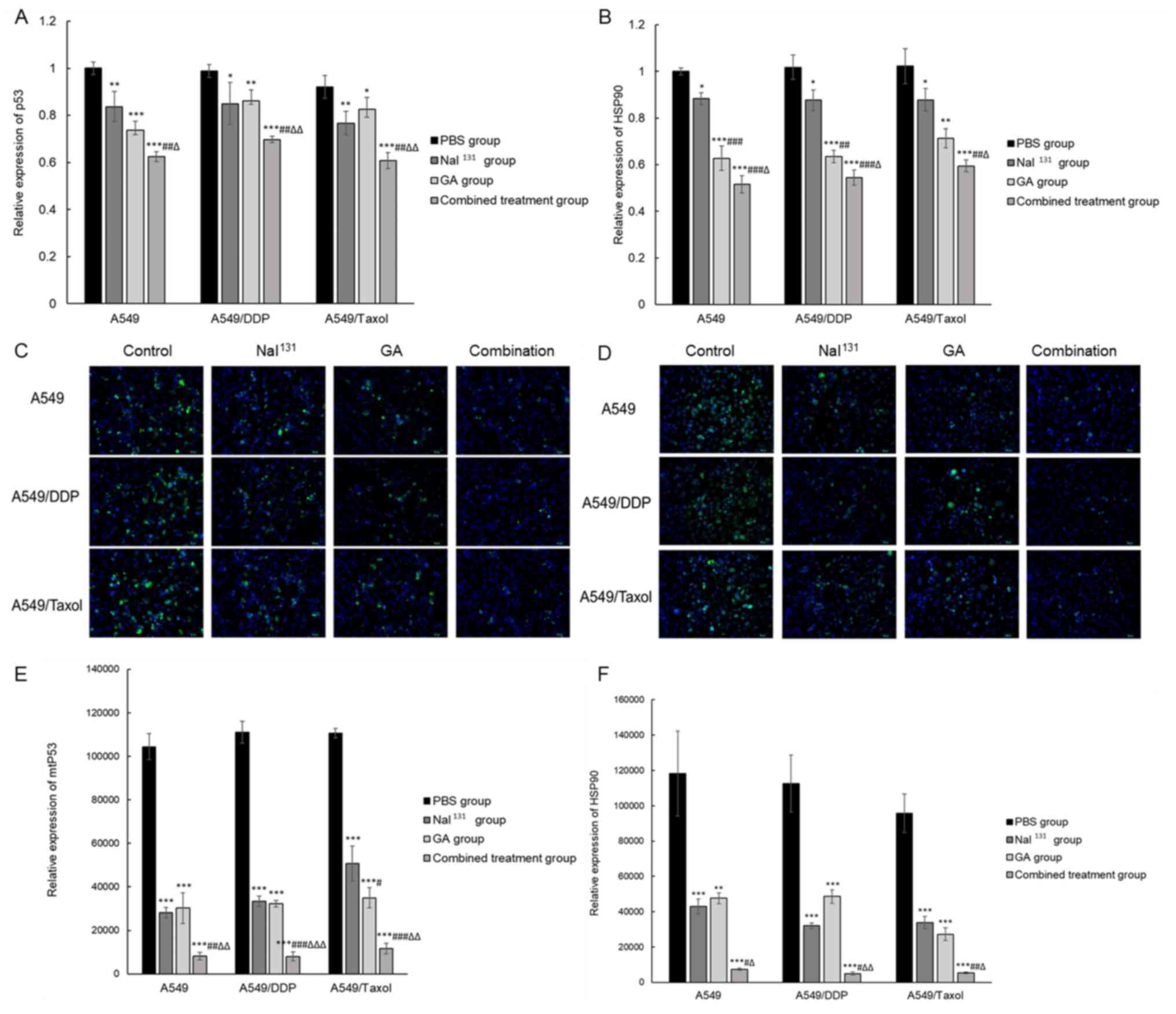 | Figure 4.Expression of mRNA and protein levels
of p53 and HSP90. Relative mRNA levels of (A) p53 and (B) HSP90 in
all the groups were detected by reverse transcription-quantitative
PCR after treatment of A549, A549/DDP and A549/Taxol cells with
NaI131, GA, or a combination of both. Protein expression
of (C) mtp53 and (D) HSP90 in all groups was detected using IF
after treatment of A549, A549/DDP and A549/Taxol cells with
NaI131, GA or a combination of both (×200
magnification). Graphical representation of the results of IF for
(E) mtp53 and (F) HSP90. * refers to comparisons with the control
group; # refers to comparisons with the
NaI131 group and ∆ refers to comparisons with
the GA group. *,#,∆P<0.05;
**,##,∆∆P<0.01; and ***,###,∆∆∆P<0.001.
DDP, cisplatin; GA, gambogic acid; PBS, phosphate-buffered saline;
mtp53, mutant p53; HSP90, heat shock protein 90; IF,
immunofluorescence. |
Additionally, the expression of HSP90 was assessed
in drug-resistant NSCLC treated with the combination of drugs. mRNA
expression of HSP90 (Fig. 4D) was
significantly decreased in the combination group compared with that
in the individual treatment groups with either NaI131 or
GA (P<0.05). IF results were in line with the RT-qPCR results,
in that the protein expression levels of HSP90 (Fig. 4E and F) were decreased in all the
groups compared with that in the control. The lowest signal was
detected in the combination group (P<0.05). The relative
expression levels of HSP90 remained very similar in A549, A549/DDP
and A549/Taxol cells treated with NaI131 and GA. These
results indicated that the effect of NaI131 combined
with GA on drug-resistant NSCLC cells may be associated with p53
and HSP90.
Discussion
The present study investigated the efficacy of using
GA as a radionuclide sensitizer to NaI131 in an in
vitro system. As an extension to our previous study (23) showing that GA below the conventional
dose can enhance the apoptosis-promoting function of
NaI131 in A549/DDP cells, the current study revealed
that compared with the control, in the combination group of all
three NSCLC lines (A549, A549/DDP and A549/Taxol) there was a
significant increase in the percentage of cells in the
G2/M phase and an increase in the number of apoptotic
cells, as well as a decrease in the levels of protein of CDK1,
cyclin B, Bcl-2, P-gp, mtp53 and HSP90, and a significant increase
in the protein levels of Bax. Studies have indicated that GA may
induce cell cycle arrest at the G1/S or G2/M
phases via various methods in different tumour cells. For example,
Wang and Yuan (25), and Yang et
al (26) reported that GA
functioned in the upregulation of endogenous reactive oxygen
species (ROS) levels in SKOV3 and RPMI-8226 cells. ROS leads to
ATM-Chk2-mediated G2/M arrest. ROS also promote cell
cycle arrest by direct actions on the Cdc25 family of protein
phosphatases (27).
A crucial function of the p53 gene is induction of
cell cycle arrest and promotion of cell apoptosis and DNA repair
(28). The cell cycle-related
regulatory proteins CDK1 and cyclin B are the major regulatory
proteins expressed in cells in the S phase and during entry into
the G2/M phase (29). p53
can cause G2/M phase arrest through downregulation of
cyclin B/CDK1 expression (30).
mtp53 can drive mitosis in tumour cells and promote tumour cell
passage through the M phase via various pathways to cause tumour
cell proliferation (31). Since
γ-radiation can be persistently released after radioactive seed
implantation in the tumour body, these cells actively proliferating
in the G2 and M phases are more sensitive to this
radiation, causing cyclin B1-mediated G2/M arrest and an
elevated Bax/Bcl2 ratio (32). GA
acting on tumour cells can arrest cancer cells at the
G1/S (33) or G2/M
(34) phase. It has been shown that
p53 has a crucial role in the induction of cell cycle arrest and
the promotion of cell apoptosis and DNA repair (28). GA in low doses was reported to cause
autophagy in mtp53 degradation in cancer cells (21). The present experimental results
showed that, in A549, A549/DDP and A549/Taxol cells, GA could
decrease the levels of cyclin B, CDK1 and mtp53 in all the groups,
and increase the number of cells in the G2/M phase,
indicating that GA could cause G2/M arrest in these
cells. In the combination group, the decrease in CDK1 and cyclin B
levels was the highest, suggesting that low-dose GA may contribute
to an increase in the number of cells in the radiation-sensitive
stage by influencing cell cycle proteins and mtp53 to increase the
lethal effect of NaI131 on tumour cells.
The mtp53 protein is upregulated in a number of
drug-resistant tumour cell lines, such as the human COLO 320DM
(homozygous R248W p53) and MIA PaCa-2 cell lines, and is thus
involved in tumour drug resistance (35,36). The
mtp53 protein can selectively upregulate the expression of multiple
drug resistance gene 1 (mdr1) (37).
P-gp encoded by mdr1 can promote cellular excretion of
chemotherapeutic drugs to produce drug resistance (6). The present study results showed that,
in the combination group, levels of mtp53 and P-gp in all the cell
types decreased, indicating that the two may have common
drug-resistance pathways. Moreover, GA has been shown to
downregulate mtp53 at the post-transcription level (22). The present study results concur with
our previous study results (23) and
demonstrate that the combinatorial effect of GA and
NaI131 may possibly assist in overcoming drug
resistance, leading to a better response to chemotherapy. In normal
cells, wild-type p53 protein is subject to ubiquitination and
degradation mediated by mouse double minute 2 (MDM2) (30). However, mtp53 protein cannot directly
activate the MDM2 ubiquitination degradation pathway, but can bind
to HSP90. HSP90 and its associated chaperones can stabilize mtp53
protein to prevent its degradation, whereas inhibition of the
function of HSP90 decreases mtp53 protein levels (38,39). GA
has been shown to prevent Hsp90/mtp53 complex formation (22). A previous study showed that Hsp90
inhibitors can enhance radiosensitivity by inhibiting the binding
of Hsp90 with client proteins (40).
However, the effect of radionuclide on mtP53 in tumour cells has
not been reported. The present study showed that, after
intervention by NaI131 and GA, the expression of HSP90
and p53 at the mRNA and protein levels decreased in all the cell
types, suggesting that these two types of intervention could
decrease the cellular expression of HSP90 and mtp53. In addition,
the decrease was more evident in the combination group, indicating
that low-dose GA combined with NaI131 could
significantly decrease HSP90 and mtp53 levels in drug-resistant
cells, likely due to the ability of GA to destabilize and degrade
mtp53 (22).
In conclusion, the present study illustrates that GA
could be a favourable radionuclide sensitizer and that a
combination of GA along with NaI131 may have advantages
in enhancing the effects of NaI131 on tumour cells by
way of governing cell cycle stages, promoting apoptosis and
reducing drug-resistance, all of which are necessary cellular
factors for cancer treatment in clinical settings. Follow-up animal
experiments should be performed to verify this conclusion.
Acknowledgements
Not applicable.
Funding
This study was supported by the Jiangsu Science and
Technology Development Project (grant no. BE2017745) and the Basic
Research Project of Southeast University (grant no.
3290005413).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JH and MD confirm the authenticity of all the raw
data. JH and MD were reponsible for conception and design. The
collection and assembly of data was performed by YW, YX and SHH.
The data analysis and interpretation was completed by XLZ, PSC and
SFY. All authors have read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
NSCLC
|
non-small cell lung cancer
|
|
GA
|
gambogic acid
|
|
P-gp
|
P-glycoprotein
|
|
mtp53
|
mutant p53
|
|
IF
|
immunofluorescence
|
|
HRP
|
horseradish peroxidase
|
|
MDM2
|
mouse double minute 2
|
References
|
1
|
de Groot PM, Wu CC, Carter BW and Munden
RF: The epidemiology of lung cancer. Transl Lung Cancer Res.
7:220–233. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cao M and Chen W: Epidemiology of lung
cancer in China. Thorac Cancer. 10:3–7. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mascaux C, Tomasini P, Greillier L and
Barlesi F: Personalised medicine for nonsmall cell lung cancer. Eur
Respir Rev. 26:1700662017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Uramoto H and Tanaka F: Recurrence after
surgery in patients with NSCLC. Transl Lung Cancer Res. 3:242–249.
2014.PubMed/NCBI
|
|
5
|
Zahreddine H and Borden KL: Mechanisms and
insights into drug resistance in cancer. Front Pharmacol. 4:282013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pan ST, Li ZL, He ZX, Qiu JX and Zhou SF:
Molecular mechanisms for tumour resistance to chemotherapy. Clin
Exp Pharmacol Physiol. 43:723–737. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Katayama R, Sakashita T, Yanagitani N,
Ninomiya H, Horiike A, Friboulet L, Gainor JF, Motoi N, Dobashi A,
Sakata S, et al: P-glycoprotein mediates ceritinib resistance in
anaplastic lymphoma kinase-rearranged non-small cell lung cancer.
EBioMedicine. 3:54–66. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li W, Guan J, Yang L, Zheng X, Yu Y and
Jiang J: Iodine-125 brachytherapy improved overall survival of
patients with inoperable stage III/IV non-small cell lung cancer
versus the conventional radiotherapy. Med Oncol. 32:3952015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang W, Li J, Li R, Zhang Y, Han M and Ma
W: Efficacy and safety of iodine-125 radioactive seeds
brachytherapy for advanced non-small cell lung cancer-A
meta-analysis. Brachytherapy. 17:439–448. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li R, Zhang Y, Yuan Y, Lin Q, Dai J, Xu R,
Hu X and Han M: Dosimetric comparison of CT-guided iodine-125 seed
stereotactic brachytherapy and stereotactic body radiation therapy
in the treatment of NSCLC. PLoS One. 12:e01873902017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yu X, Li J, Zhong X and He J: Combination
of Iodine-125 brachytherapy and chemotherapy for locally recurrent
stage III non-small cell lung cancer after concurrent
chemoradiotherapy. BMC Cancer. 15:6562015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Song J, Fan X, Zhao Z, Chen M, Chen W, Wu
F, Zhang D, Chen L, Tu J and Ji J: 125I brachytherapy of
locally advanced non-small-cell lung cancer after one cycle of
first-line chemotherapy: A comparison with best supportive care.
Onco Targets Ther. 10:1345–1352. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cui YH, Suh Y, Lee HJ, Yoo KC, Uddin N,
Jeong YJ, Lee JS, Hwang SG, Nam SY, Kim MJ and Lee SJ: Radiation
promotes invasiveness of non-small-cell lung cancer cells through
granulocyte-colony-stimulating factor. Oncogene. 34:5372–5382.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dess RT, Sun Y, Matuszak MM, Sun G, Soni
PD, Bazzi L, Murthy VL, Hearn JWD, Kong FM, Kalemkerian GP, et al:
Cardiac events after radiation therapy: Combined analysis of
prospective multicenter trials for locally advanced non-small-cell
lung cancer. J Clin Oncol. 35:1395–1402. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Banik K, Harsha C, Bordoloi D, Lalduhsaki
Sailo B, Sethi G, Leong HC, Arfuso F, Mishra S, Wang L, Kumar AP
and Kunnumakkara AB: Therapeutic potential of gambogic acid, a
caged xanthone, to target cancer. Cancer Lett. 416:75–86. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang H, Zhao Z, Lei S, Li S, Xiang Z, Wang
X and Huang X, Xia G and Huang X: Gambogic acid induces autophagy
and combines synergistically with chloroquine to suppress
pancreatic cancer by increasing the accumulation of reactive oxygen
species. Cancer Cell Int. 19:72019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Qi Q, Lu N, Li C, Zhao J, Liu W, You Q and
Guo Q: Involvement of RECK in gambogic acid induced anti-invasive
effect in A549 human lung carcinoma cells. Mol Carcinog. 54 (Suppl
1):E13–E25. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Qi Q, You Q, Gu H, Zhao L, Liu W, Lu N and
Guo Q: Studies on the toxicity of gambogic acid in rats. J
Ethnopharmacol. 117:433–438. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang Y, Yang L, You QD, Nie FF, Gu HY,
Zhao L, Wang XT and Guo QL: Differential apoptotic induction of
gambogic acid, a novel anticancer natural product, on hepatoma
cells and normal hepatocytes. Cancer Lett. 256:259–266. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kashyap D, Mondal R, Tuli HS, Kumar G and
Sharma AK: Molecular targets of gambogic acid in cancer: Recent
trends and advancements. Tumour Biol. 37:12915–12925. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Foggetti G, Ottaggio L, Russo D, Monti P,
Degan P, Fronza G and Menichini P: Gambogic acid counteracts mutant
p53 stability by inducing autophagy. Biochim Biophys Acta Mol Cell
Res. 1864:382–392. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang J, Zhao Q, Qi Q, Gu HY, Rong JJ, Mu
R, Zou MJ, Tao L, You QD and Guo QL: Gambogic acid-induced
degradation of mutant p53 is mediated by proteasome and related to
CHIP. J Cell Biochem. 112:509–519. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Huang J, Zhu X, Wang H, Han S, Liu L, Xie
Y, Chen D, Zhang Q, Zhang L and Hu Y: Role of gambogic acid and
NaI131 in A549/DDP cells. Oncol Lett. 13:37–44. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang J and Yuan Z: Gambogic acid
sensitizes ovarian cancer cells to doxorubicin through ROS-mediated
apoptosis. Cell Biochem Biophys. 67:199–206. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang LJ, Chen Y, He J, Yi S, Wen L, Zhao S
and Cui GH: Effects of gambogic acid on the activation of caspase-3
and downregulation of SIRT1 in RPMI-8226 multiple myeloma cells via
the accumulation of ROS. Oncol Lett. 3:1159–1165. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Srinivas US, Tan BWQ, Vellayappan BA and
Jeyasekharan AD: ROS and the DNA damage response in cancer. Redox
Biol. 25:1010842019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kastenhuber ER and Lowe SW: Putting p53 in
Context. Cell. 170:1062–1078. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Huang Y, Sramkoski RM and Jacobberger JW:
The kinetics of G2 and M transitions regulated by B cyclins. PLoS
One. 8:e808612013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ryan KM, Phillips AC and Vousden KH:
Regulation and function of the p53 tumor suppressor protein. Curr
Opin Cell Biol. 13:332–337. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Blandino G, Valenti F, Sacconi A and Di
Agostino S: Wild type- and mutant p53 proteins in mitochondrial
dysfunction: Emerging insights in cancer disease. Semin Cell Dev
Biol. 98:105–117. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Qu A, Wang H, Li J, Wang J, Liu J, Hou Y,
Huang L and Zhao Y: Biological effects of (125)i seeds radiation on
A549 lung cancer cells: G2/M arrest and enhanced cell death. Cancer
Invest. 32:209–217. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li R, Chen Y, Zeng LL, Shu WX, Zhao F, Wen
L and Liu Y: Gambogic acid induces G0/G1 arrest and apoptosis
involving inhibition of SRC-3 and inactivation of Akt pathway in
K562 leukemia cells. Toxicology. 262:98–105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yu J, Guo QL, You QD, Zhao L, Gu HY, Yang
Y, Zhang HW, Tan Z and Wang X: Gambogic acid-induced G2/M phase
cell-cycle arrest via disturbing CDK7-mediated phosphorylation of
CDC2/p34 in human gastric carcinoma BGC-823 cells. Carcinogenesis.
28:632–638. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hosain SB, Khiste SK, Uddin MB, Vorubindi
V, Ingram C, Zhang S, Hill RA, Gu X and Liu YY: Inhibition of
glucosylceramide synthase eliminates the oncogenic function of p53
R273H mutant in the epithelial-mesenchymal transition and induced
pluripotency of colon cancer cells. Oncotarget. 7:60575–60592.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Do PM, Varanasi L, Fan S, Li C, Kubacka I,
Newman V, Chauhan K, Daniels SR, Boccetta M, Garrett MR, et al:
Mutant p53 cooperates with ETS2 to promote etoposide resistance.
Genes Dev. 26:830–845. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nguyen KT, Liu B, Ueda K, Gottesman MM,
Pastan I and Chin KV: Transactivation of the human multidrug
resistance (MDR1) gene promoter by p53 mutants. Oncol Res. 6:71–77.
1994.PubMed/NCBI
|
|
38
|
Nagata Y, Anan T, Yoshida T, Mizukami T,
Taya Y, Fujiwara T, Kato H, Saya H and Nakao M: The stabilization
mechanism of mutant-type p53 by impaired ubiquitination: The loss
of wild-type p53 function and the hsp90 association. Oncogene.
18:6037–6049. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Patel HJ, Modi S, Chiosis G and Taldone T:
Advances in the discovery and development of heat-shock protein 90
inhibitors for cancer treatment. Expert Opin Drug Discov.
6:559–587. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Spiegelberg D, Dascalu A, Mortensen AC,
Abramenkovs A, Kuku G, Nestor M and Stenerlöw B: The novel HSP90
inhibitor AT13387 potentiates radiation effects in squamous cell
carcinoma and adenocarcinoma cells. Oncotarget. 6:35652–35666.
2015. View Article : Google Scholar : PubMed/NCBI
|














