Introduction
Gastric cancer is one of the most common types of
malignant tumor in the gastrointestinal tract worldwide. Its
morbidity and mortality are ranked fourth and fifth, respectively,
amongst all cancer types globally (1,2). The
development of gastroscopy technology has significantly improved
the diagnostic and cure rates of patients with early gastric
cancer. Except for a small number of early gastric cancer cases
with small lesion areas, gastroscopy is suitable for endoscopic
treatment. For other cases of gastric cancer, surgery and
chemotherapy remain the primary treatment options (3,4).
Cisplatin (DDP) is a commonly used chemotherapeutic drug in the
clinic; however, the insensitivity of gastric cancer cells to DDP
limits its efficacy (5). Therefore,
further in-depth studies into the molecular mechanisms of gastric
cancer and cellular resistance are required to identify novel
therapeutic targets and implement individualized treatment
strategies for gastric cancer.
Long non-coding RNAs (lncRNA) are a type of
non-coding RNA of >200 nucleotides in length, which contain
neither a start nor stop codon (6).
LncRNAs have been revealed to play vital roles in multiple cellular
processes, such as gene expression regulation, genome imprinting
and chromatin packaging, and during various stages, including
cellular differentiation and embryonic development (7). lncRNA CDKN2B antisense RNA 1 (ANRIL) is
an antisense non-coding RNA (8), and
a 403-kb deletion in the gene was first identified in patients with
melanoma and tumors of the nervous system (9). ANRIL is expressed in a variety of
normal tissues, with the highest expression levels found in ovarian
tissue and the lowest in muscle tissue (10). In addition, previous studies have
reported that the expression levels of ANRIL were significantly
upregulated in lung cancer, liver cancer and esophageal squamous
cell carcinoma, which suggests that ANRIL may function as an
oncogene (11,12).
It is well established that lncRNAs can regulate
microRNA (miRNA/miR) expression and thereby influence tumor
progression (13). A large number of
studies have reported that miRNAs bind to target genes to regulate
the occurrence and development of numerous tumor types (14). Previous research has also shown that
the expression of miR-181a-5p was downregulated in gastric cancer
and that the overexpression of miR-181a-5p inhibited cellular
proliferation (15).
Drug resistance in gastric cancer cells limits the
efficacy of drugs in the clinic. The present study aimed to
determine the effects of ANRIL and miR-181-5p on DDP-sensitive and
-resistant gastric cancer cells, as well as the underlying
mechanisms of action. The results may provide novel targets to
overcome drug resistance in gastric cancer.
Materials and methods
Cell lines and culture
Normal gastric epithelial cells (GES1) and gastric
cancer cell lines (HGC-27, AGS, HSC-39 and FU97) were purchased
from the American Type Culture Collection. DDP (purity >99.99%)
was purchased from Merck KGaA. Cells were cultured in DMEM (Gibco;
Thermo Fisher Scientific, Inc.) supplemented with 10% FBS (Gibco;
Thermo Fisher Scientific, Inc.), and maintained at 37°C with 5%
CO2. Cells were exposed to increasing concentrations of
DDP to establish DDP-resistant gastric cancer cells. Briefly, cells
in the logarithmic growth phase were first cultured with 0.05 µg/ml
DDP. Following a week, the surviving cells were subsequently
cultured with increasing concentrations of DDP until the cells
could be stably subcultured in medium containing 0.5 µg/ml DDP.
Each time the concentration of DDP was increased by 1.2 or 1.5
times, the process experienced 9 times of concentration increase
and six months in total.
Reverse transcription-quantitative PCR
(RT-qPCR)
For mRNA expression analysis, total RNA was
extracted from cultured cells using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). Total mRNA was
reverse transcribed into cDNA using the FSQ-101 reverse
transcription system kit (Toyobo Life Science), and qPCR was
subsequently performed using the QuantiTect SYBR-Green PCR kit
(Qiagen, Inc.). For miRNA expression analysis, total RNA was
extracted from cells using the fast microRNA isolation kit (BioTeke
Corporation). Reverse transcription and qPCR were subsequently
performed using the miScript RT kit and miScript SYBR Green qPCR
kit (GeneCopoeia, Inc.), respectively, according to the
manufacturers' protocols. The qPCR thermocycling conditions were as
follows: Pre-denaturation at 95°C for 30 sec, followed by 40 cycles
of denaturation at 95°C for 5 sec, annealing at 60°C for 30 sec,
and extension at 72°C for 10 sec. The primers used for qPCR are
listed in Table I. Expression levels
of mRNA and miRNA were quantified using the 2−∆∆Cq
method (16) and normalized to that
of GADPH or U6, respectively.
 | Table I.Sequences of the primers for
qPCR. |
Table I.
Sequences of the primers for
qPCR.
| Gene | Sequence |
|---|
| CCNG1 | Forward:
5′-AATGAAGGTACAGCCCAAGCA-3′ |
|
| Reverse:
5′-GCTTTGACTTTCCAACACACC-3′ |
| ANRIL | Forward:
5′-CTCCAGACAGGGTCTCACTC-3′ |
|
| Reverse:
5′-GGAAATTCCTAGCTCCGTAA-3′ |
| miR-181a-5p | Forward:
5′-GCCGAACATTCAACGCTGTCG-3′ |
|
| Reverse:
5′-GTGCAGGGTCCGAGGT-3′ |
| GAPDH | Forward:
5′-AAATCCCATCACCATCTTCCAG-3′ |
|
| Reverse:
5′-GAGTCCTTCCACGATACCAAAGTTG-3′ |
| U6 | Forward:
5′-ATTGGAACGATACAGAGAAGATT-3′ |
|
| Reverse:
5′-GGAACGCTTCACGAATTTG-3′ |
Transfection
Cells were digested with 0.25% trypsin and seeded
into a 6-well plate at a density of 2×105 cells/well,
which was then incubated at 37°C (5% CO2) for 24 h. The
cells were subsequently transfected with 100 nM miR-181a-5p
inhibitors, inhibitor-negative control (NC), short hairpin RNA
(shRNA)-ANRIL vectors and scrambled shRNA-NC (all Shanghai
GenePharma Co., Ltd.) using Lipofectamine® 2000 reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) for 24 h at 37°C.
shRNA-ANRIL-1 and shRNA-ANRIL-2 were incorporated into the same
vector, and both sequences were provided by GenePharma Co., Ltd.
Following transfection for 24 h, the cells were utilized for
subsequent experiments.
Cell Counting Kit-8 (CCK-8) assay
Transfected cells in the logarithmic growth phase
were digested with 0.25% trypsin, seeded into a 96-well plate at a
density of 5×104 cells/well, and incubated at 37°C for
24 h. Following incubation, 10 µl CCK-8 reagent (Beyotime Institute
of Biotechnology) was added to each well and the plate was
incubated for a further hour. The optical density of each well was
measured at a wavelength of 450 nm using a microplate reader
(Thermo Fisher Scientific, Inc.).
Flow cytometry
For apoptosis analysis, cells were digested with
0.25% trypsin and washed twice with PBS. Cells (1×105
per sample) were subsequently centrifuged at 150 × g, resuspended
in binding buffer, and then incubated with Annexin V-FITC in the
dark at room temperature for 30 min. The cells were then incubated
at room temperature with propidium iodide (PI) in the dark for 5
min. For cell cycle distribution analysis, following centrifugation
at 150 × g and resuspension in PBS, the cells were incubated with
PI staining solution (50 mg/l P1, 1 g/l Triton X and 100 g/l RNase)
in the dark at 4°C for 30 min. Data were analyzed using FlowJo
(v7.6.1; FlowJo, LLC) following flow cytometry on a FACSCalibur
flow cytometer (BD Biosciences).
Western blotting
Total protein was extracted from cells using RIPA
lysis (Beyotime Institute of Biotechnology), and quantified using a
BCA assay kit (Beyotime Institute of Biotechnology). Protein
samples (25 µg) were separated via 10% SDS-PAGE, transferred onto
PVDF membranes, and then blocked with 5% BSA (Thermo Fisher
Scientific, Inc.) at room temperature for 1 h. The membranes were
subsequently incubated with the following primary antibodies at 4°C
overnight: Anti-p53 (cat. no. MA5-16387; 1:1,000), anti-Bax (cat.
no. PA5-11378; 1:2,000), anti-Bcl-2 (cat. no. PA5-27094; 1:1,000),
anti-cleaved caspase-3 (cat. no. PA5-114687; 1:1,000), anti-cyclin
G1 (CCNG1; cat. no. PA5-36050; 1:1,000), anti-cyclin D1 (CCND1;
cat. no. PA5-32373; 1:1,000), anti-CDK4 (cat. no. PA5-27827;
1:1,000) and anti-GAPDH (cat. no. MA1-16757; 1:2,000) (all Thermo
Fisher Scientific, Inc.). Following primary antibody incubation,
the membranes were incubated with an HRP-conjugated goat
anti-rabbit antibody (cat. no. G-21234; Thermo Fisher Scientific,
Inc.; 1:100,000) at room temperature for 1 h. Protein bands were
visualized using chemiluminescent reagents and densitometric
analysis was performed using ImageJ version 1.52 software (National
Institutes of Health).
Dual-luciferase reporter assay
ANRIL and CCNG1 wild-type (WT) and mutant-type (MUT)
pmiRGlo luciferase expression vectors containing miR-181-5p binding
sites were constructed by Shanghai GenePharma Co., Ltd., and
co-transfected with NC mimic or miR-181-5p mimic into cells
(5×105 cells/well) plated into 96-well plates using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). The Dual Luciferase Reporter Gene Assay kit
(cat. no. RG027; Beyotime Institute of Biotechnology) was used
according to the manufacturer's instructions. The relative
luciferase activity was measured after 48 h and normalized to the
activity of Renilla luciferase. Sequences used to regulate
endogenous miR-181a-5p were listed in Table II.
| Table II.Sequences used to regulate endogenous
miR-181a-5p. |
Table II.
Sequences used to regulate endogenous
miR-181a-5p.
| Gene | Sequence |
|---|
| miR-181a-5p
mimic- | Forward:
5′-AACAUUCAACGCUGUCGGUGAGU-3′ |
|
| Reverse:
5′-UUUUGUAAGUUGCGACAGCCACU-3′ |
| mimic NC | Forward:
5′-UUCUCCGAACGUGUCACGUTT-3′ |
|
| Reverse:
5′-TTAAGAGGCUUGCACAGUGCA-3′ |
| miR-181a-5p
inhibitor |
5′-ACUCACCGACAGCGUUGAAUGUU-3′ |
| inhibitor NC |
5′-CAGUACUUUUGUGUAGUACAA-3′ |
Bioinformatics analysis
The Encyclopedia of RNA Interactomes (ENCORI)
(17) was used to predict miRNAs
that bind to ANRIL, and TargetScan, PicTar and miRanda were used to
predict target genes that bind to miRNA. ENCORI (http://starbase.sysu.edu.cn/) is an open-source
platform for studying the interactions between various RNAs.
‘CDKN2B AS’ was entered into the webpage and the associated miRNAs
were displayed. TargetScan (18)
(http://www.targetscan.org/) predicts the
biological targets of miRNAs by searching for the presence of
conserved 8mer and 7mer sites that match the seed region of each
miRNA. PicTar (19) (https://pictar.mdc-berlin.de/) uses an algorithm to
identify the microRNA targets. miRanda (20) (http://www.microrna.org/microrna/home.do) is an online
database for miRNA target prediction and functional annotations.
miR-181a-5p was entered and the results of the three databases were
comprehensively considered.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 8.0 software (GraphPad Software, Inc.) and quantitative data
are presented as the mean ± SD. Statistical differences between two
groups were determined using unpaired Student's t-test, while
one-way ANOVA and Tukey's post hoc test were used to determine
statistical differences between multiple groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
Construction of drug-resistant cell
lines and detection of relative CCNG1, ANRIL and miR-181a-5p
expression levels
Using the ENCORI database, miR-181a-5p was predicted
to share a targeting relationship with ANRIL and CCNG1 via
TargetScan, PicTar and miRanda. In particular, the 3′-untranslated
region (UTR) region of miR-181a-5p was found possess a binding site
for ANRIL. Furthermore, the 3′-UTR region of CCNG1 was found to
share binding sites with miR-181a-5p, which suggested that CCNG1
may be a potential target gene of miR-181a-5p. Multiple
DDP-resistant gastric cancer cell lines were constructed, and the
expression levels of CCNG1 in normal and DDP-resistant cells were
analyzed using RT-qPCR. Compared with GES1 cells, CCNG1 expression
levels were dysregulated in DDP-sensitive cell lines (**P<0.01
and ***P<0.001). Notably, the expression levels of CCNG1 in
DDP-resistant cell lines were upregulated compared with those of
the GES1 cell line (***P<0.001; Fig.
1A), and CCNG1 expression was upregulated to the greatest
extent in HGC-27 and FU97 DDP-resistant cell lines. The expression
levels of ANRIL and miR-181a-5p were also analyzed using RT-qPCR,
both of which were dysregulated in DDP-sensitive cells; however,
ANRIL expression was upregulated and miR-181a-5p expression was
downregulated in drug-resistant cells (*P<0.05, **P<0.01 and
***P<0.001; Fig. 1B and C). For
HGC-27 and FU97 cell lines, the differences in expression between
drug-resistant and sensitive cell lines were more apparent;
therefore, these two cell lines were used for subsequent
experimentation.
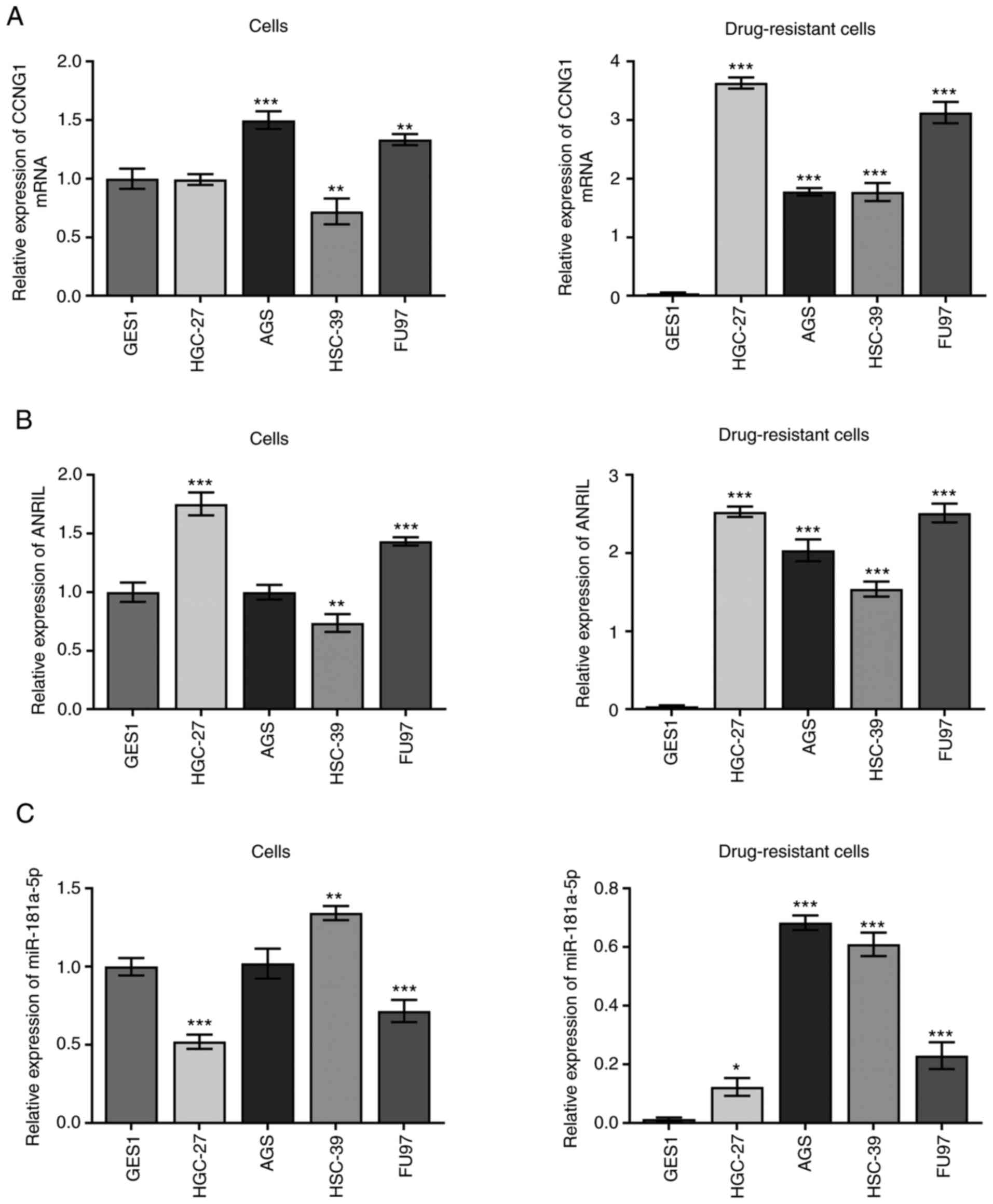 | Figure 1.Expression levels of CCNG1,
lncRNA-ANRIL and miR-181a-5p in cisplatin-sensitive and -resistant
cell lines. Expression levels of (A) CCNG1, (B) lncRNA-ANRIL and
(C) miR-181a-5p in gastric GES1 epithelial cells and HGC-27, AGS,
HSC-39 and FU97 gastric cancer cell lines, and their associated
drug-resistant lines. *P<0.05, **P<0.01 and ***P<0.001 vs.
GES1 cells; n≥3. CCNG1, cyclin G1; lncRNA, long non-coding RNA;
ANRIL, CDKN2B antisense RNA 1; miR, microRNA. |
Proliferation of gastric cancer cells
and sensitivity to DDP
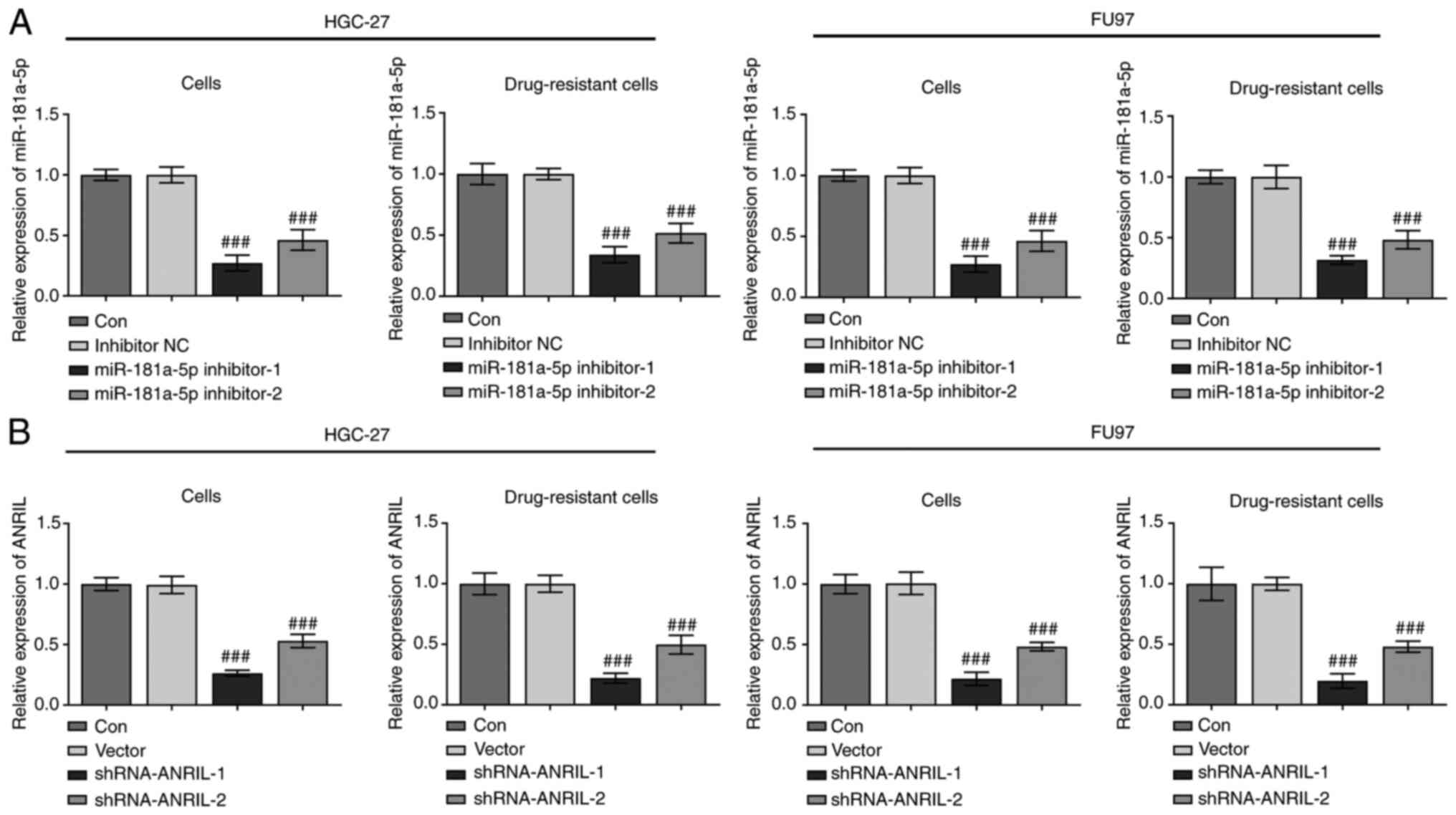
Cells were transfected with miR-181a-5p inhibitors
and shRNA-ANRIL vectors, and RT-qPCR was then used to analyze
transfection efficiency. In both the HGC-27 and FU97 sensitive and
DDP-resistant cell lines, compared with the inhibitor NC, the
expression levels of miR-181-5p were significantly downregulated in
the miR-181-5p inhibitor groups (###P<0.001), with
the greatest effect observed in cells transfected with miR-181a-5p
inhibitor-1 compared with miR-181a-5p inhibitor-2 (Fig. 2A). In addition, the decrease in ANRIL
expression was more significant when cells were transfected with
shRNA-ANRIL-1 compared with shRNA-ANRIL-2 (##P<0.001;
Fig. 2B). Therefore, miR-181a-5p
inhibitor-1 and shRNA-ANRIL-1 were selected for use in subsequent
experiments.
Cells were then divided into five groups: i)
Control; ii) shRNA-NC; iii) shRNA-ANRIL; iv) shRNA-ANRIL +
inhibitor NC; and v) shRNA-ANRIL + miR-181a-5p inhibitor. The
proliferation of sensitive and DDP-resistant cell lines was
subsequently detected using a CCK-8 assay. The proliferation of
DDP-resistant cell lines was increased compared with sensitive cell
lines in all groups. In addition, transfection with shRNA-ANRIL
decreased cellular proliferation (###P<0.001), while
upon co-transfection with the miR-181a-5p inhibitor, this decrease
in proliferation was reversed ($$$P<0.001; Fig. 3A). These results suggested that ANRIL
may promote gastric cell proliferation, while miR-181a-5p may
inhibit proliferation. The sensitivity of cells in different groups
to DDP was also detected using a CCK-8 assay. Following the
knockdown of ANRIL, cell sensitivity increased
(##P<0.01, ###P<0.001); however,
following co-transfection of the miR-181a-5p inhibitor, cell
sensitivity was decreased ($$P<0.01,
$$$P<0.001). This trend was more apparent in
drug-resistant cell lines (Fig.
3B).
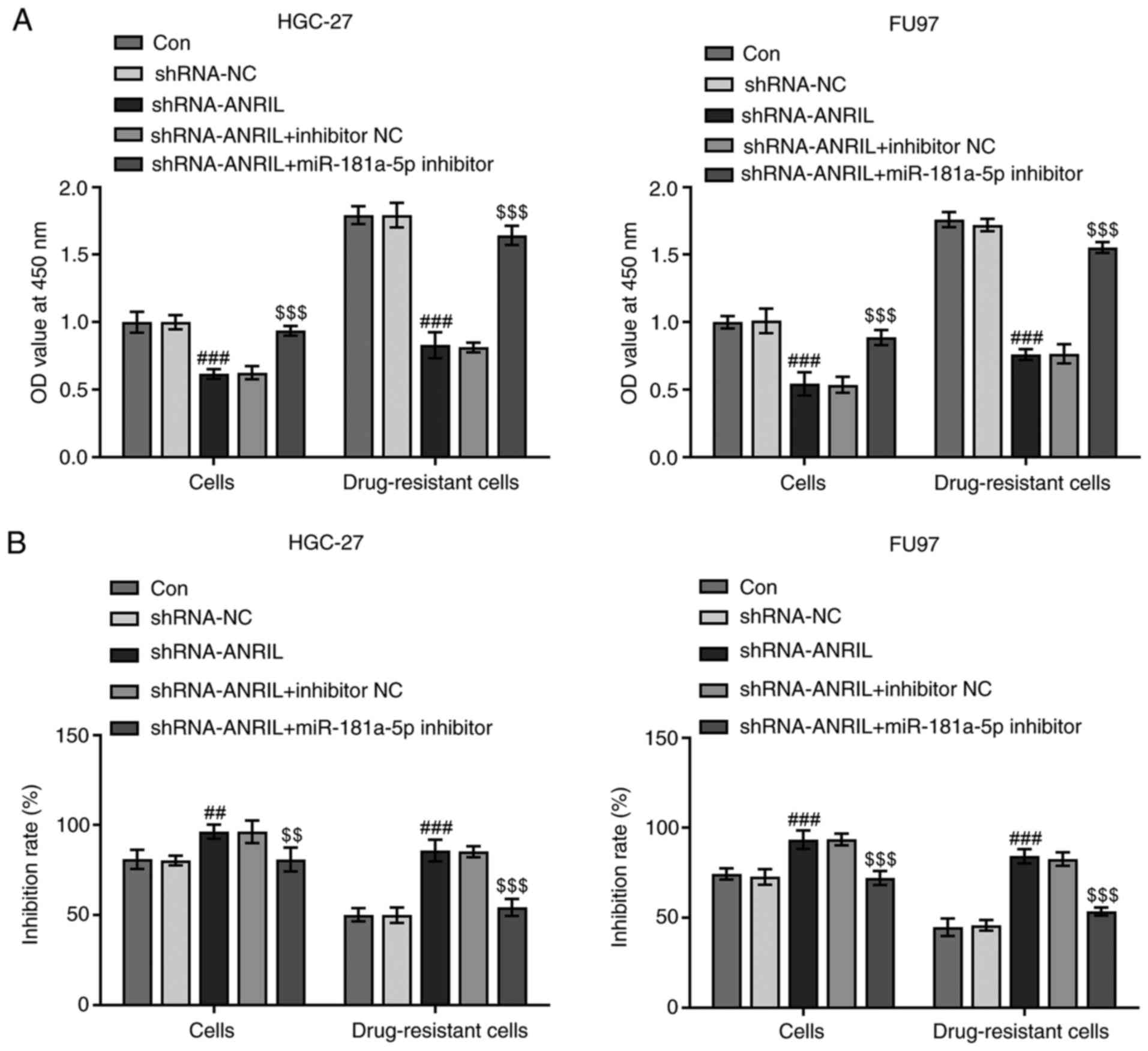 | Figure 3.Proliferation and sensitivity of
cells to DDP. (A) Cells were divided into five groups: Control,
shRNA-NC, shRNA-ANRIL, shRNA-ANRIL + inhibitor NC and shRNA-ANRIL +
miR-181a-5p inhibitor. OD value at 450 nm of HGC-27, FU97 and
drug-resistant cell lines was determined. ###P<0.001
vs. shRNA-NC; $$$P<0.001 vs. shRNA-ANRIL + inhibitor
NC; n≥3. (B) Sensitivity to DDP is shown as the inhibition rate of
HGC-27, FU97 and drug-resistant cell lines. ##P<0.01
and ###P<0.001 vs. shRNA-NC; $$P<0.01
and $$$P<0.001 vs. shRNA-ANRIL + inhibitor NC; n≥3.
DDP, cisplatin; shRNA, short hairpin RNA; Con, control; NC,
negative control; miR, microRNA; OD, optical density; ANRIL, CDKN2B
antisense RNA 1. |
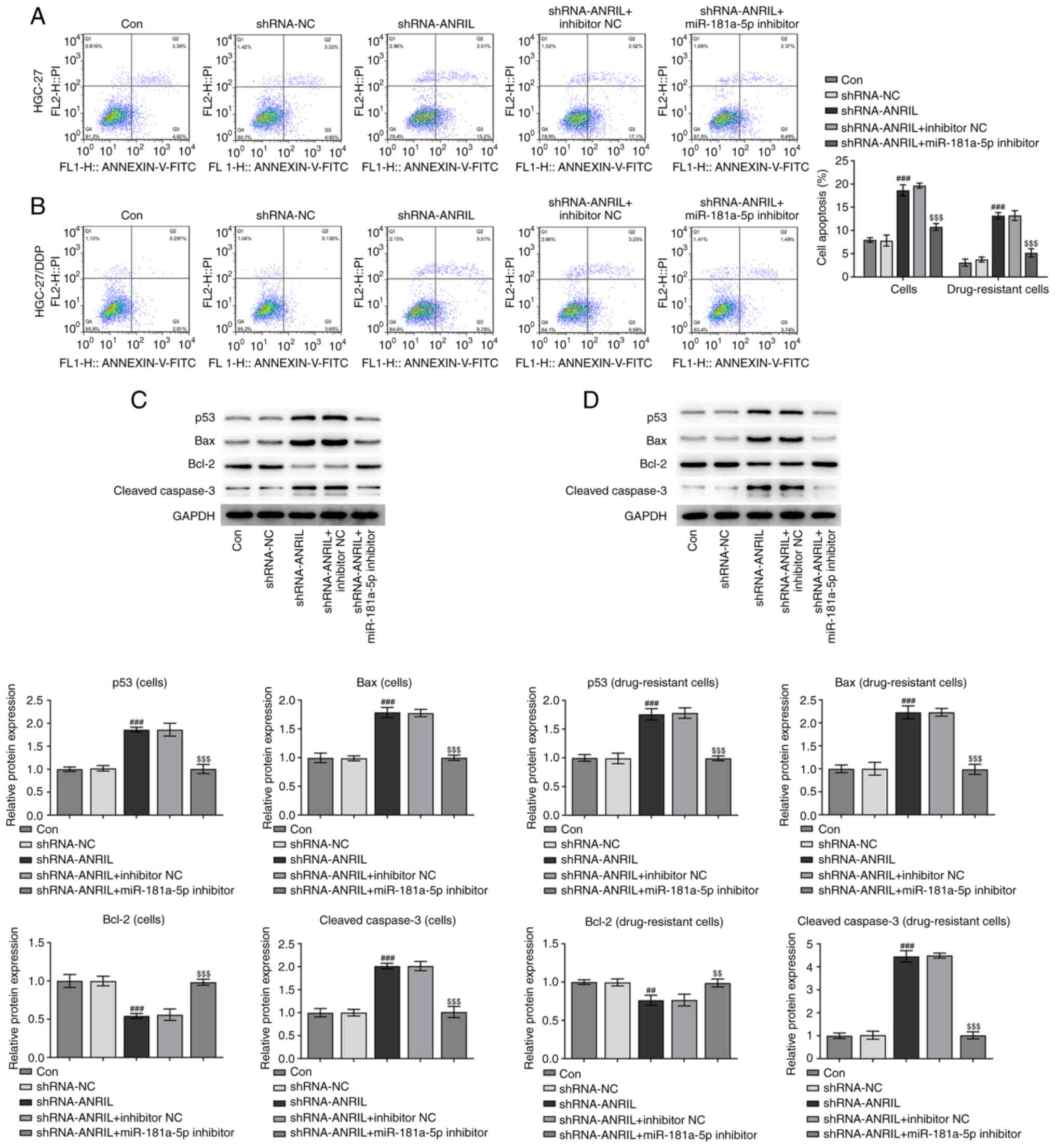
Detection of apoptosis
Apoptosis in the different groups was analyzed using
flow cytometry. The knockdown of ANRIL in DDP-sensitive or
-resistant cell lines increased the levels of apoptosis.
Conversely, the subsequent co-transfection with the miR-181a-5p
inhibitor reversed this trend (###P<0.001 and
$$$P<0.001; Fig. 4A and
B). In addition, western blotting was used to analyze the
expression levels of p53, Bax, Bcl-2 and cleaved caspase-3, which
are all associated with apoptosis. The expression levels of p53,
Bax and cleaved caspase-3 were upregulated following the
transfection of cells with shRNA-ANRIL, while the expression levels
of Bcl-2 were downregulated (###P<0.001; Fig. 4C). Notably, the expression levels of
each protein were reversed following the co-transfection with the
miR-181-5p inhibitor ($$P<0.01 and
$$$P<0.001). The trends in the expression levels of
each protein in the DDP-resistant cell groups were similar;
however, the magnitude of the changes were more notable
(##P<0.01, ###P<0.001,
$$P<0.01 and $$$P<0.001; Fig. 4D). Results from flow cytometry and
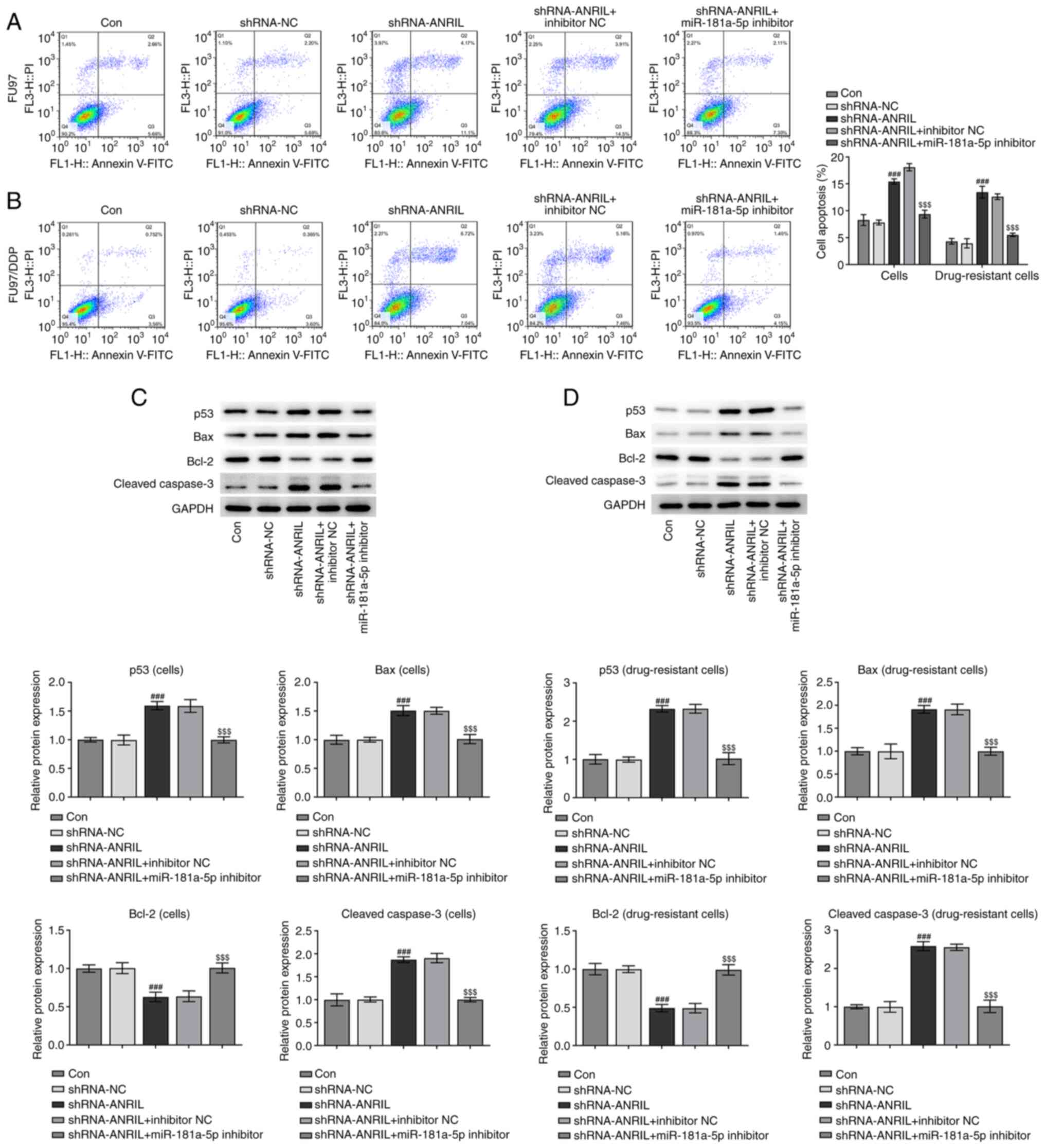
western blotting analyses in FU97 cell lines revealed a similar
overall trend to that in the HGC-27 cell line
(###P<0.001 and $$$P<0.001; Fig. 5A-D).
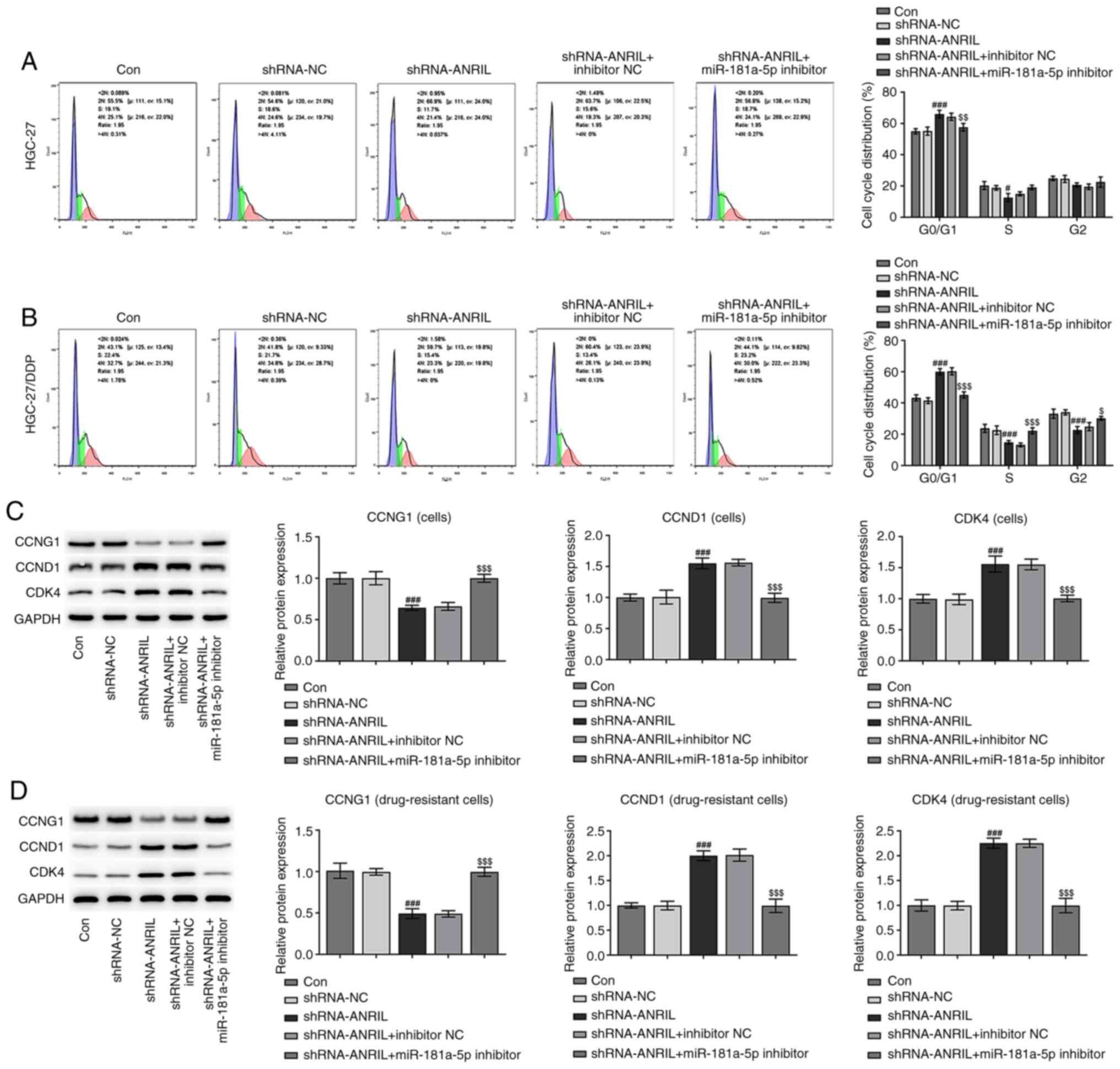
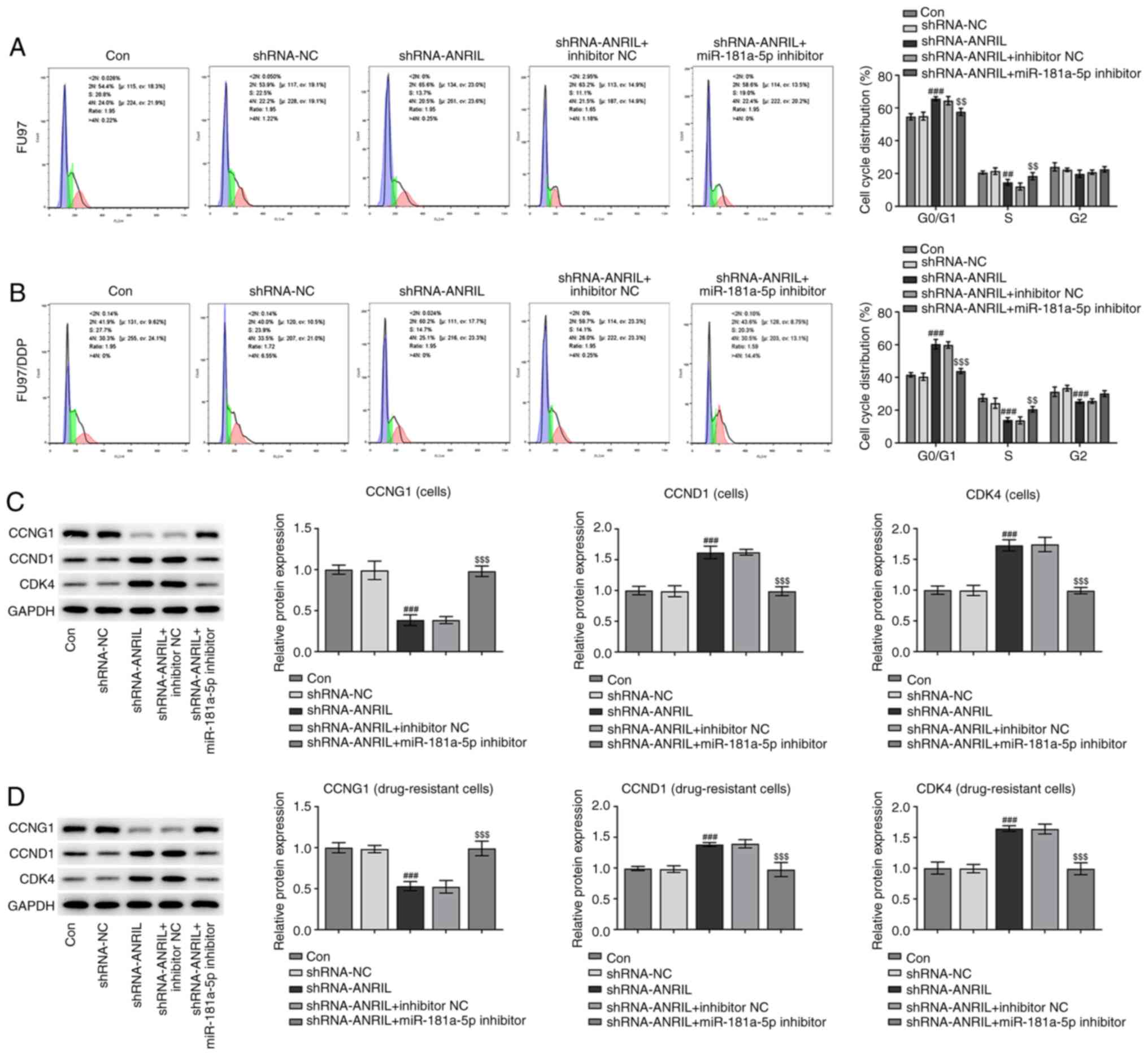
Cell cycle distribution analysis. Following the
transfection of HGC-27 cells with shRNA-ANRIL, the number of cells
in the G0/1 phase was increased; however,
following co-transfection with the miR-181a-5p inhibitor, the
number of cells in the G0/1 phase was
decreased. Notably, there were fewer DDP-resistant cells in the
G0/1 phase compared with sensitive cells
without transfection, and more cells in the S and G2/M
phases. Following the transfection of DDP-resistant cells with
shRNA-ANRIL, the number of cells in the G0/1 phase was
increased, but then decreased following co-transfection with the
miR-181a-5p inhibitor (#P<0.05,
###P<0.001, $P<0.05,
$$P<0.01 and $$$P<0.001; Fig. 6A and B). In addition, western
blotting was used to analyze the expression levels of CCNG1, CCND1
and CDK4, which are cell cycle-related proteins. The expression
levels of CCND1 and CDK4 were upregulated following ANRIL-knockdown
(###P<0.001), and were subsequently downregulated
following co-transfection with the miR181a-5p inhibitor
($$$P<0.001). The combined action of CCND1 and CDK4
promotes the progression of cells from the G1 to S
phase. These results indicated that tANRIL-knockdown may induce
cell cycle arrest in the G0/1 phase, while
the knockdown of miR-181a-5p may relieve cell cycle arrest
(Fig. 6C). This phenomenon was more
apparent in DDP-resistant cells (###P<0.001 and
$$$P<0.001; Fig.
6D).
Following ANRIL-knockdown in FU97 cells, the number
of cells arrested in the G0/1 phase was
increased, indicating that ANRIL may be involved in the progression
of cells from the G0/1 to S phase. Following
the knockdown of miR-181a-5p, the number of cells in the
G2/M phase was increased and the number of cells in the
G0/1 phase was decreased, indicating that the
knockdown of miR-181a-5p may promote progression to the
G2/M phase (##P<0.01,
###P<0.001, $$P<0.01 and
$$$P<0.001; Fig. 7A and
B). These findings are consistent with the results in HGC-27
cells. The results of western blot analysis of the expression
levels of CCNG1, CCND1 and CDK4 in FU97 cells were consistent with
the findings in the HGC-27 cell line (###P<0.001 and
$$$P<0.001; Fig. 7C and
D).
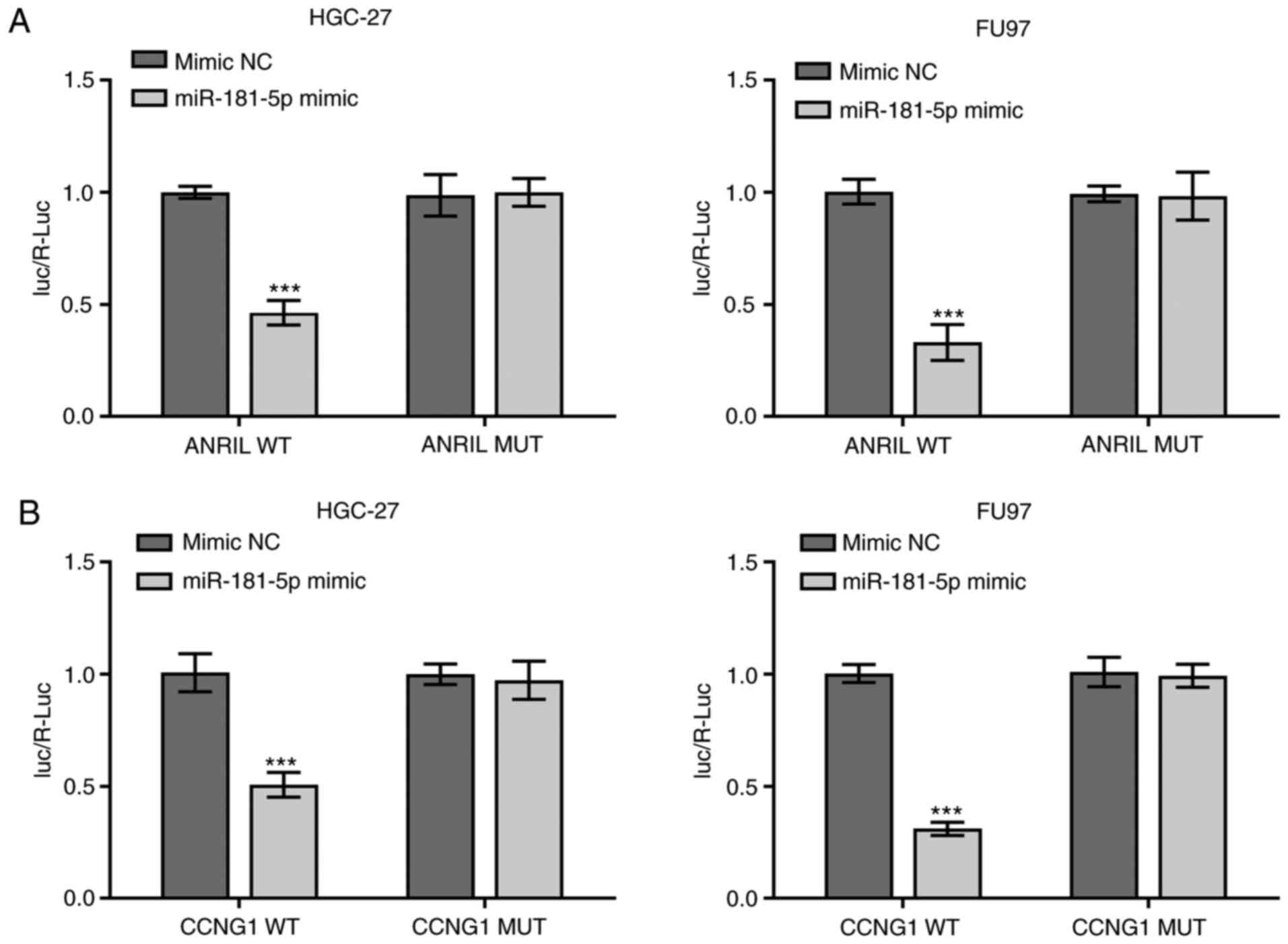
Verification of the association
between miR-181a-5p and lncRNA-ANRIL or -CCNG1
The relationship between miR-181a-5p and
lncRNA-ANRIL or -CCNG1 were verified using a dual luciferase
reporter assay. Compared with the mimic NC-transfected cells, the
relative luciferase activity of both HGC-27 and FU97 cells
co-transfected with the WT-ANRIL vector and miR-181-5p mimics were
significantly reduced (***P<0.001), while the differences in
relative luciferase activitiy between cells co-transfected with the
MUT-ANRIL vectors and miR-181-5p mimics or mimic NC were not
statistically significant (Fig. 8A).
In addition, the relative luciferase activity of cells
co-transfected with the WT-CCNG1 vector and miR-181-5p mimics was
also significantly reduced (***P<0.001), while the differences
between the relative luciferase activities of cells co-transfected
with MUT-CCNG1 and miR-181-5p mimic or mimic NC were not
statistically significant (Fig.
8B).
Discussion
Gastric cancer has a high incidence worldwide. For
early gastric cancer, direct surgical resection is the most
effective treatment option (21),
while for advanced gastric cancer, chemotherapy is required
following surgery (22). When the
tumor adheres to the surrounding tissues, chemotherapy is the main
treatment option, and it has been found to significantly improve
the survival rate of patients with advanced gastric cancer
(23). DDP is a commonly used
chemotherapeutic drug; however, the development of drug resistance
is a significant challenge that limits its clinical efficacy
(24). The mechanisms underlying
drug resistance in tumor cells are complex (25), thus the establishment of
drug-resistant cell lines in vitro is a prerequisite for
studying the biological changes of tumor cells. In the present
study, a stepwise continuous selection method was used to construct
DDP-resistant gastric cancer cell lines.
Previous studies reported that the expression levels
of CCNG1 were significantly upregulated in a number of tumor tissue
types, which promoted tumor cell proliferation (26,27). The
dysregulated expression of CCNG1 resulted in abnormal cell cycle
regulation, thereby affecting tumor occurrence and development
(28). Zhang et al (29) analyzed the association between ANRIL
and clinicopathological features and prognosis in cervical cancer
by determining the expression of ANRIL in cervical cancer tissues
and cell lines. The results revealed that patients with high ANRIL
expression had advanced FIGO stage, lymph node metastasis and poor
overall survival. In addition, the knockdown of ANRIL expression
inhibited cellular proliferation, migration and the occurrence of
cervical cancer.
In the present study, following induction with DDP,
the expression levels of CCNG1, ANRIL and miR-181a-5p determined to
confirm the successful construction of drug-resistant cells. The
results of RT-qPCR analysis indicated that the expression levels of
CCNG1 and ANRIL were significantly upregulated in drug-resistant
compared with sensitive cell lines. As for miR-181-5p, the
expression levels in sensitive cells were somewhat inconsistent,
and multiple studies have reported that the role of miR-181-5p in
gastric cancer is controversial. For example, Chen et al
(30) demonstrated that miR-181a-5p
expression levels were upregulated in gastric cancer, and that the
overexpression of miR-181a-5p promoted cellular proliferation.
Furthermore, Lu et al (31)
reported that knocking down metastasis associated lung
adenocarcinoma 1 (MALAT1) inhibited proliferation and promoted the
apoptosis of MGC-803 cells. The overexpression of miR-181a-5p
exhibited similar effects to MALAT1-knockdown. In addition, Ghaedi
et al (32) did not identify
any significant differences in miR-181a-5p expression between
patients with gastric cancer and healthy participants. However,
these findings do not affect those of the present study on
drug-resistant cell lines, because miR-181a-5p was found to be
downregulated in all constructed drug-resistant cell lines. In the
current study, HGC-27 and FU97 cell lines were selected for
subsequent experiments.
With the development of high-throughput sequencing
technology, the roles of lncRNAs have attracted the attention of
numerous research groups. Previous studies have found that lncRNAs
have a high degree of tissue specificity, and are involved in a
variety of tumor cells processes (33). ANRIL has been associated with the
development of malignant tumor types, suggesting its significant
potential in cancer (11,12). However, the current underlying
mechanism of ANRIL remains undetermined, and further investigation
is required to establish its function. To elucidate the role of
ANRIL in sensitive and drug-resistant cell lines, cells were
transfected with an miR-181a-5p inhibitor and shRNA-ANRIL vector,
and transfection efficiency was determined using RT-qPCR. A CCK-8
assay was also used to detect the proliferation and sensitivity of
non-resistant and DDP-resistant cells to DDP. Following
ANRIL-knockdown in drug-resistant cells, the current results
suggested that cellular proliferation was significantly decreased
following 24 h of culture, indicating that ANRIL may promote the
proliferation of gastric cancer cells. Compared with the control
cells, the sensitivity of drug-resistant cell lines to DDP was
markedly increased. To the best of our knowledge, only one previous
study has studied the role of ANRIL in drug-resistant gastric
cancer cells (34), and the
downstream regulation of ANRIL has not been studied. In the present
study, using bioinformatics analysis via ENCORI database,
miR-181a-5p was predicted to share a targeting relationship with
ANRIL. miR-181a-5p was also found to inhibit the proliferation of
drug-resistant cells and enhance sensitivity to DDP. To the best of
our knowledge, these findings have not been reported by others.
Based on the aforementioned findings, flow cytometry
was used to investigate the effect of ANRIL and miR-181a-5p on
apoptosis and cell cycle distribution. The results of the flow
cytometric apoptosis analysis were consistent with those of the
CCK-8 assay. Furthermore, ANRIL-knockdown was found to promote cell
cycle arrest, while following the knockdown of miR-181a-5p, cell
cycle arrest was alleviated. These findings suggested that ANRIL
and miR-181-5p may both be involved in the cell cycle.
Subsequently, western blotting was used to detect the expression
levels of proteins related to apoptosis and the cell cycle, which
verified the results of the flow cytometric experiments. Finally,
the relationships between miR-181a-5p and ANRIL or CCNG1 were
verified using dual luciferase reporter assays. The binding of
miR-181a-5p to lncRNA ANRIL and CCNG1 was investigated separately,
and binding interactions were identified between both.
In conclusion, the results of the present study
suggested that the knockdown of ANRIL expression may inhibit the
proliferation of gastric cancer cells, promote apoptosis and induce
cell cycle arrest. In addition, its downstream target, miR-181a-5p,
was found to not only reverse the effects of ANRIL (especially in
drug-resistant cells), thereby playing a tumor-suppressive role,
but also to regulate the function of the target gene, CCNG1. To the
best of our knowledge, the present study is the first to
investigate the downstream regulation of ANRIL in gastric cancer.
The discovery that miR-181a-5p improves the sensitivity of
drug-resistant cells to DDP may provide insights into novel
treatments for drug-resistant gastric cancer. However, only in
vitro experiments were conducted, and further in vivo
investigation is required to validate the present findings.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Ningbo
Natural Science Foundation (grant no. 2019A610337).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XH and TL designed the study, and performed the
experiments alongside CY and YSW. XT wrote the manuscript and
analyzed the data alongside YW. TZ conceived the study, supervised
the experiments and revised the manuscript for important
intellectual content. All authors read and approved the final
manuscript. XH and TZ confirm the authenticity of all the raw
data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Taft RJ, Pheasant M and Mattick JS: The
relationship between non-protein-coding DNA and eukaryotic
complexity. Bioessays. 29:288–299. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Iyer MK, Niknafs YS, Malik R, Singhal U,
Sahu A, Hosono Y, Barrette TR, Prensner JR, Evans JR, Zhao S, et
al: The landscape of long noncoding RNAs in the human
transcriptome. Nat Genet. 47:199–208. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fidler IJ, Yano S, Zhang RD, Fujimaki T
and Bucana CD: The seed and soil hypothesis: Vascularisation and
brain metastases. Lancet Oncol. 3:53–57. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ellis LM and Hicklin DJ: VEGF-targeted
therapy: Mechanisms of anti-tumour activity. Nat Rev Cancer.
8:579–591. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang C, Wen Z, Xie J, Zhao Y, Zhao L,
Zhang S, Liu Y, Xue Y and Shi M: MACC1 mediates chemotherapy
sensitivity of 5-FU and cisplatin via regulating MCT1 expression in
gastric cancer. Biochem Biophys Res Commun. 485:665–671. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hombach S and Kretz M: Non-coding RNAs:
Classification, biology and functioning. Adv Exp Med Biol.
937:3–17. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bhan A and Mandal SS: Long noncoding RNAs:
Emerging stars in gene regulation, epigenetics and human disease.
ChemMedChem. 9:1932–1956. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yap KL, Li S, Muñoz-Cabello AM, Raguz S,
Zeng L, Mujtaba S, Gil J, Walsh MJ and Zhou MM: Molecular interplay
of the noncoding RNA ANRIL and methylated histone H3 lysine 27 by
polycomb CBX7 in transcriptional silencing of INK4a. Mol Cell.
38:662–674. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tachibana I, Smith JS, Sato K, Hosek SM,
Kimmel DW and Jenkins RB: Investigation of germline PTEN, p53,
p16(INK4A)/p14(ARF), and CDK4 alterations in familial glioma. Am J
Med Genet. 92:136–141. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shen XH, Qi P and Du X: Long non-coding
RNAs in cancer invasion and metastasis. Mod Pathol. 28:4–13. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nie FQ, Sun M, Yang JS, Xie M, Xu TP, Xia
R, Liu YW, Liu XH, Zhang EB, Lu KH and Shu YQ: Long noncoding RNA
ANRIL promotes non-small cell lung cancer cell proliferation and
inhibits apoptosis by silencing KLF2 and P21 expression. Mol Cancer
Ther. 14:268–277. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen D, Zhang Z, Mao C, Zhou Y, Yu L, Yin
Y, Wu S, Mou X and Zhu Y: ANRIL inhibits p15(INK4b) through the
TGFβ1 signaling pathway in human esophageal squamous cell
carcinoma. Cell Immunol. 289:91–96. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Arita T, Ichikawa D, Konishi H, Komatsu S,
Shiozaki A, Shoda K, Kawaguchi T, Hirajima S, Nagata H, Kubota T,
et al: Circulating long non-coding RNAs in plasma of patients with
gastric cancer. Anticancer Res. 33:3185–3193. 2013.PubMed/NCBI
|
|
14
|
Rupaimoole R and Slack FJ: MicroRNA
therapeutics: Towards a new era for the management of cancer and
other diseases. Nat Rev Drug Discov. 16:203–222. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lin F, Li Y, Yan S, Liu S, Qian W, Shen D,
Lin Q and Mao W: MicroRNA-181a inhibits tumor proliferation,
invasiveness, and metastasis and is downregulated in gastric
cancer. Oncol Res. 22:75–84. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42((Database Issue)): D92–D97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lewis BP, Shih IH, Jones-Rhoades MW,
Bartel DP and Burge CB: Prediction of mammalian microRNA targets.
Cell. 115:787–798. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Krek A, Grün D, Poy MN, Wolf R, Rosenberg
L, Epstein EJ, MacMenamin P, da Piedade I, Gunsalus KC, Stoffel M
and Rajewsky N: Combinatorial microRNA target predictions. Nat
Genet. 37:495–500. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Enright AJ, John B, Gaul U, Tuschl T,
Sander C and Marks DS: MicroRNA targets in Drosophila.
Genome Biol. 5:R12003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Oda I, Shimazu T, Ono H, Tanabe S, Iishi
H, Kondo H and Ninomiya M: Design of Japanese multicenter
prospective cohort study of endoscopic resection for early gastric
cancer using web registry (J-WEB/EGC). Gastric Cancer. 15:451–454.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jim MA, Pinheiro PS, Carreira H, Espey DK,
Wiggins CL and Weir HK: Stomach cancer survival in the United
States by race and stage (2001–2009): Findings from the CONCORD-2
study. Cancer. 123 (Suppl 24):S4994–S5013. 2017. View Article : Google Scholar
|
|
23
|
Song Z, Wu Y, Yang J, Yang D and Fang X:
Progress in the treatment of advanced gastric cancer. Tumour Biol.
39:10104283177146262017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li Y, Lv S, Ning H, Li K, Zhou X, Xv H and
Wen H: Down-regulation of CASC2 contributes to cisplatin resistance
in gastric cancer by sponging miR-19a. Biomed Pharmacother.
108:1775–1782. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wei L, Sun J, Zhang N, Zheng Y, Wang X, Lv
L, Liu J, Xu Y, Shen Y and Yang M: Noncoding RNAs in gastric
cancer: Implications for drug resistance. Mol Cancer. 19:622020.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xu Y, Zhang Q, Miao C, Dongol S, Li Y, Jin
C, Dong R, Li Y, Yang X and Kong B: CCNG1 (Cyclin G1) regulation by
mutant-P53 via induction of Notch3 expression promotes high-grade
serous ovarian cancer (HGSOC) tumorigenesis and progression. Cancer
Med. 8:351–362. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Han H, Zhang Z, Yang X, Yang W, Xue C and
Cao X: miR-23b suppresses lung carcinoma cell proliferation through
CCNG1. Oncol Lett. 16:4317–4324. 2018.PubMed/NCBI
|
|
28
|
Ye XX, Liu CB, Chen JY, Tao BH and Zhi-Yi
C: The expression of cyclin G in nasopharyngeal carcinoma and its
significance. Clin Exp Med. 12:21–24. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang D, Sun G, Zhang H, Tian J and Li Y:
Long non-coding RNA ANRIL indicates a poor prognosis of cervical
cancer and promotes carcinogenesis via PI3K/Akt pathways. Biomed
Pharmacother. 85:511–516. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen G, Shen ZL, Wang L, Lv CY, Huang XE
and Zhou RP: hsa-miR-181a-5p expression and effects on cell
proliferation in gastric cancer. Asian Pac J Cancer Prev.
14:3871–3875. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lu Z, Luo T, Pang T, Du Z, Yin X, Cui H,
Fang G and Xue X: MALAT1 promotes gastric adenocarcinoma through
the MALAT1/miR-181a-5p/AKT3 axis. Open Biol. 9:1900952019.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ghaedi H, Mozaffari MAN, Salehi Z, Ghasemi
H, Zadian SS, Alipoor S, Hadianpour S and Alipoor B: Co-expression
profiling of plasma miRNAs and long noncoding RNAs in gastric
cancer patients. Gene. 687:135–142. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Capizzi M, Strappazzon F, Cianfanelli V,
Papaleo E and Cecconi F: MIR7-3HG, a MYC-dependent modulator
of cell proliferation, inhibits autophagy by a regulatory loop
involving AMBRA1. Autophagy. 13:554–566. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lan WG, Xu DH, Xu C, Ding CL, Ning FL,
Zhou YL, Ma LB, Liu CM and Han X: Silencing of long non-coding RNA
ANRIL inhibits the development of multidrug resistance in gastric
cancer cells. Oncol Rep. 36:263–270. 2016. View Article : Google Scholar : PubMed/NCBI
|