Introduction
Liver cancer, divided into primary and secondary
hepatic carcinoma, is one of the most common malignant tumors and
the second leading cause of cancer-associated mortality worldwide
based on the GLOBOCAN 2008 estimates (1). It is believed that the pathogenesis of
primary liver cancer is a complicated process involving multiple
factors (2). Epidemiological
research has shown that hepatitis B virus (HBV) and hepatitis C
virus (HCV) infection, aflatoxin, alcoholism and nitrosamines are
the main risk factors that lead to the incidence of primary hepatic
carcinoma (3). Secondary hepatic
carcinoma may develop in different ways, such as with blood and
lymph metastasis and direct invasion of the liver (4). Surgical resection, including liver
transplantation, is the major liver cancer therapy and results in
survival benefits for patients with liver cancer (5); however, the overall survival and
prognosis of patients remain unsatisfactory on account of the high
relapse rate (6). Approximately 20%
of patients survive for ≥5 years after diagnosis (7). Therefore, further exploration of
crucial liver cancer molecular mechanisms is of paramount
importance to seek effective therapeutic strategies.
Long non-coding RNAs (lncRNAs), a novel type of RNAs
comprising >200 nucleotides in length, are known to be involved
in the tumorigenesis and progression of numerous human malignancies
(8). For example, lncRNA-ATB
promotes cell proliferation and migration in papillary thyroid
cancer (9), while lncRNA SOCS2-AS1
promotes the proliferation and suppresses apoptosis of prostate
cancer cells (10). lncRNA HEIH
serves as a marker of poor prognosis and facilitates colorectal
cancer tumorigenesis (11).
Moreover, the pivotal roles of lncRNAs in liver cancer progression
have been previously clarified (12–14).
Increasing research has demonstrated that lncRNA LOXL1-antisense
RNA 1 (LOXL1-AS1) serves an oncogenic role in multiple types of
cancer. For instance, LOXL1-AS1 downregulation suppresses the
proliferation and induces apoptosis of breast cancer cells
(15). Additionally, LOXL1-AS1
exerts oncogenic effects on the tumorigenesis of lung
adenocarcinoma (16). LncRNA
LOXL1-AS1 facilitates pancreatic cancer progression by promoting
pancreatic cancer cell proliferation and migration (17). Moreover, LOXL1-AS1 facilitates
gastric cancer progression by modulating the malignant phenotypes
of gastric cancer cells (18) and
facilitates the proliferation and invasion of non-small cell lung
cancer cells (19). However, its
role in liver cancer remains unclear.
The present study focused on the role of LOXL1-AS1
in liver cancer, which may provide potential novel insights for the
treatment of liver cancer.
Materials and methods
Patients and samples
Liver cancer tissues (n=38) and paired adjacent
non-tumor tissues (n=38) (≥3 cm from the tumor margin) were
collected from 24 male and 14 female patients with liver cancer
(age range, 33–71 years; mean age ± SD, 53.1±5.4 years) of the
Affiliated Hospital of Yangzhou University (Yangzhou, China)
between August 2017 and August 2019. The collected samples were
stored at −80°C. The inclusion criteria were as follows: i)
Patients diagnosed with hepatocellular carcinoma based on imaging
and pathological biopsy; ii) patients meeting the TNM staging
criteria for hepatocellular carcinoma by the American Joint
Committee on Cancer in 2009 (20);
iii) patients who had not taken part in any previous targeted tumor
research (surgery, radiotherapy or chemotherapy), understood the
study and signed an informed consent form (including their
families); and iv) patients whose expected survival time was >3
months. The exclusion criteria were as follows: Patients with other
combined tumors, renal function diseases or infection before
admission. The present study received approval from the Ethics
Committee of the Affiliated Hospital of Yangzhou University.
Cell lines
Liver cancer cell lines (SK-HEP-1, Hep3B and SNU1)
and a normal human hepatic cell line (THLE-2) were obtained from
The Cell Bank of Type Culture Collection of The Chinese Academy of
Sciences. All cells were cultured in DMEM/F-12 (cat no. 31330095;
Gibco; Thermo Fisher Scientific, Inc.) mixed with high glucose
containing 10% FBS (Invitrogen; Thermo Fisher Scientific, Inc.) and
were grown in a humid atmosphere with 5% CO2 at
37°C.
Transfection
Short hairpin RNA (shRNA) against LOXL1-AS1
(sh-LOXL1-AS1) was utilized to knock down LOXL1-AS1, and sh-NC was
used as the negative non-targeting control. To overexpress nuclear
factor I B (NFIB), the sequences of NFIB were subcloned into the
pcDNA3.1 vector to produce pcDNA3.1/NFIB. The empty pcDNA3.1 vector
served as the control. The microRNA (miRNA/miR)-377-3p mimics
vector was used to overexpress miR-377-3p, and NC mimics (cat. no.
B01001) were used as the scrambled control. All vectors were
obtained from Shanghai GenePharma Co., Ltd. SK-HEP-1 and Hep3B
cells (1×106 cells/well) were seeded in 24-well plates
and 500 µl DMEM was added to each well. When the cells reached
40–60% confluence, the aforementioned vectors were transfected into
cells at a final concentration of 50 nM using
Lipofectamine® 3000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) at 37°C with 5% CO2. Transfection was
conducted for 48 h according to the manufacturer's
instructions.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from tissues and cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). The extracted RNA was reverse transcribed into cDNA using a
PrimeScript RT reagent kit (Takara Bio, Inc.) according to the
manufacturer's protocol. qPCR was conducted using a SYBR Premix Ex
Taq II kit (Takara Bio, Inc.) with the ABI7500 system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The PCR amplification
conditions were as follows: A pre-denaturation step of 1 min at
94°C, followed by 40 cycles at 95°C for 30 sec and 60°C for 1 min.
The relative quantitation of gene expression was calculated using
the 2−ΔΔCq method (21).
GAPDH and RNU6 (U6) were used as internal controls for lncRNAs (or
mRNAs) and miRNAs, respectively. The PCR primers used are shown in
Table I.
 | Table I.Primer sequences used for reverse
transcription-quantitative PCR. |
Table I.
Primer sequences used for reverse
transcription-quantitative PCR.
| Target | Primer sequences
(5′-3′) |
|---|
| LOXL1-AS1 | F:
TTCCCATTTACCTGCCCGAAG |
|
| R:
GTCAGCAAACACATGGCAAC |
| miR-377-3p | F:
ATCACACAAAGGCAACTTTTGT |
|
| R:
ATCACACAAAGGCAACTTTTGT |
| miR-524-5p | F:
GCTGTGACCCTACAAAGGGA |
|
| R:
ACCGTAACACTCCAAAGGGA |
| miR-526b-5p | F:
ACCCTCTTGAGGGAAGCACT |
|
| R:
GACAGTAAGCCTCTAAAAGGAAGC |
| miR-3614-5p | F:
TTCAAAACACCAAGATCTGAAGGC |
|
| R:
CTTGGGCCACTTGGATCTGA |
| miR-18b-5p | F:
TGTGCAAATCCATGCAAAACTGA |
|
| R:
GTGCAGGGTCCGAGGT |
| NFIB | F:
ACAAAGTCTGGCGTCTGGAT |
|
| R:
GGCTGGACACAAAGTGCTG |
| NR3C1 | F:
GAATGAACCTGGAAGCTCG |
|
| R:
AGGTTTCTTGTGAGACTCCT |
| GRSF1 | F:
AAGCTGATGTGCACTTTGAG |
|
| R:
TATACCTATGATGAACGTGGGAC |
| PRR7 | F:
ACCACCGTGTTACGAAGAG |
|
| R:
GTGAGCACCTCGCTATAGG |
| MADD | F:
AAAGCATTTAAAGCAGGCCT |
|
| R:
TGGCCCTCATGAAATTTCTC |
| ARHGAP29 | F:
CTAACAAATTTGTGCAGCCTC |
|
| R:
GCTCCTGTTTCCAAAGCTC |
| SSFA2 | F:
ATATGGCTCAAGGACTGCC |
|
| R:
TTTCTCACAAGCACACCCT |
| KDM6B | F:
AGGAAGTCCTGTGATTGGC |
|
| R:
CTTTCACAGCCAATTCCGG |
| XIAP | F:
GGAGGGCTAACTGATTGGA |
|
| R:
TAACAGATATTTGCACCCTGGA |
| FMR1 | F:
CATGCACTTTCGGAGTCTG |
|
| R:
CAAGCTGCCTTGAACTCTC |
| NELFA | F:
ATCTTCCCATCTCAGCCTC |
|
| R:
ACTCACCCACTGTAATCCC |
| GAPDH | F:
CGGAGTCAACGGATTTGGTCGTAT |
|
| R:
AGCCTTCTCCATGGTGGTGAAGAC |
| U6 | F:
GCTTCGGCAGCACATATACTAAAAT |
|
| R:
CGCTTCACGAATTTGCGTGTCAT-3 |
MTT assay
An MTT assay was performed to examine cell
proliferation. SK-HEP-1 and Hep3B cells were plated in 96-well
plates at a density of 4×103 cells/well and cultured at
room temperature for 0, 24, 48 and 72 h at a density of 60–70%
after adherence. At every transfection time point, 20 µl of MTT
solution (5 mg/ml) was added to each well of the culture plate,
followed by another 4 h of incubation at room temperature.
Subsequently, the precipitated formazan was dissolved in dimethyl
sulfoxide. The absorbance at 490 nm was detected using an ELX-800
spectrometer reader (Bio-Tek Instruments, Inc.; Agilent
Technologies, Inc.).
Colony formation assay
In brief, the transfected liver cancer cells were
seeded in 6-well plates (1×103 cells/well) and grown in
DMEM mixed with high glucose containing 10% FBS at 37°C. The
culture medium was replaced every 3 days. After incubation for 2
weeks, the colonies were fixed with methanol at room temperature
for 10 min, followed by staining using crystal violet at room
temperature for 5 min. The visible colonies with >50 cells were
counted manually with a light microscope.
Glucose uptake assay
A 2-Deoxyglucose Glucose Uptake Assay kit (Abcam)
was used to perform the glucose uptake assay. A total of
1×106 transfected SK-HEP-1 and Hep3B cells cultured in
DMEM/F-12 mixed with glucose (0, 5 and 20 mM; cat no. 31330095;
Gibco; Thermo Fisher Scientific, Inc.) containing 10% FBS
(Invitrogen; Thermo Fisher Scientific, Inc.) were grown in 96-well
plates. After overnight incubation, the cells were incubated with
2-deoxyglucose in the dark for 30 min in a humid atmosphere with 5%
CO2 at 37°C. The detection of 2-deoxyglucose uptake was
observed under a fluorescence microplate reader.
Western blot analysis
Liver cancer cells were lysed in RIPA lysis buffer
(Beyotime Institute of Biotechnology), and the protein
concentration was measured utilizing a bicinchoninic acid assay.
Equal amounts of protein (20 µg/lane) were separated via 12%
SDS-PAGE for 2 h and transferred onto polyvinylidene fluoride
membranes (EMD Millipore). Subsequently, the membranes were blocked
with 5% skim milk for 1 h at room temperature, followed by
incubation with diluted primary antibodies, including
anti-E-cadherin (1:1,000; cat. no. ab194982; Abcam),
anti-N-cadherin (1:4,000; cat. no. ab18203; Abcam), anti-LOXL1
(1:200; cat. no. ab238152; Abcam) and anti-GAPDH (1:500; cat. no.
ab181602; Abcam) at 4°C overnight. The membranes were further
incubated with horseradish peroxidase-conjugated goat anti-rabbit
secondary antibody (cat. no. sc-2357; 1:5,000; Santa Cruz
Biotechnology, Inc.) for 1 h at 37°C. The protein bands were
visualized using an electrochemiluminescence assay (EMD Millipore)
and analyzed using the Odyssey Infrared Imaging system version 2.1
(LI-COR Biosciences).
Wound healing assay
Horizontal cell migration was examined using a wound
healing assay. Liver cancer cells were treated with 3 µM of
mitomycin C (Roche Diagnostics) for 1 h at 37°C to prevent further
cell proliferation. After transfection for 48 h, cells were
subjected to serum starvation for 4 h and incubated in 6-well
plates overnight. When cells reached 80% confluence, a wound was
created using a sterile pipette tip. Subsequently, the cells were
washed twice with PBS and cultured for 12 and 24 h at room
temperature. The wound was photographed at 0, 12 and 24 h under a
light microscope (magnification, ×100) and quantified using
Image-Pro Plus Analysis software (National Institutes of
Health).
Bioinformatics analysis
The potential downstream genes of LOX1-AS1 were
predicted using the starBase database (http://starbase.sysu.edu.cn/) with the condition of 4
cancer types. The putative downstream genes of miR-377-3p were
predicted using the starBase database (http://starbase.sysu.edu.cn/) overlapped by microT,
miRanda, miRmap, PicTar and PITA (CLIP Data: Strict stringency;
Degradome data: High stringency). The interaction between LOX1-AS1
and miR-377-3p or the interaction between NFIB and miR-377-3p was
also predicted using the starBase database.
RNA pull-down assay
For the RNA pull-down assay, Bio-LOXL1-AS1 (or
Bio-miR-377-3p) and Bio-NC (Guangzhou RiboBio Co., Ltd.) were
transcribed by the Biotin RNA Labeling Mix (Roche Diagnostics) and
T7 RNA polymerase (Promega Corporation), and liver cancer cells
were transfected with 50 nM Biotin-labeled RNAs using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) for 48 h at room temperature. Subsequently, the
cells were lysed using RIPA lysis buffer (Sangon Biotech Co., Ltd.)
containing RNase inhibitors (Invitrogen; Thermo Fisher Scientific,
Inc.). After 1 ml of cell lysates were collected, 20 µl of
biotin-labeled RNA and 40 µl of streptavidin magnetic beads were
added and incubated with cell lysates for 2 h, forming RNA
complexes. Proteinase K (Sigma-Aldrich; Merck KGaA) was added and
incubated with the supernatants overnight at 4°C to isolate the
RNA. Beads were isolated from the supernatant after centrifugation
(2,500 × g, 5 min, 4°C) and washed with washing buffer (10 mM
Tris-HCl pH 7.5, 1 mM EDTA, 2 M NaCl and 0.1% Tween-20) followed by
another centrifugation step (2,500 × g, 5 min, 4°C). The pellet was
then collected, and the RNA-RNA complexes were eluted using
Tris-EDTA buffer (Invitrogen; Thermo Fisher Scientific, Inc.) and
purified using ethanol. Finally, miR-377-3p, miR-524-5p,
miR-526b-5p, miR-3614-5p and miR-18b-5p expression was measured by
RT-qPCR, as aforementioned.
Luciferase reporter assays
The full-length LOXL1-AS1 [wild-type (Wt)/mutant
(Mut)] or NFIB 3′-untranslated region (UTR) (Wt/Mut) was subcloned
into pmirGLO vectors (Promega Corporation) to construct luciferase
reporter vectors (LOXL1-AS1-Wt/Mut and NFIB-Wt/Mut). The
LOXL1-AS1-Wt vectors or LOXL1-AS1-Mut vectors were co-transfected
with the miR-377-3p mimics or NC mimics into liver cancer cells,
respectively. In addition, NFIB-Wt vectors or NFIB-Mut vectors were
co-transfected with the miR-377-3p mimics or NC mimics into liver
cancer cells. Transfection was conducted using
Lipofectamine® 3000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) for 48 h, after which the relative luciferase
activity was detected using a luciferase reporter assay system
(E1910; Promega Corporation) in comparison with Renilla
luciferase activity.
Statistical analysis
All assays were repeated ≥3 times. Statistical data,
presented as the mean ± SD, were analyzed using SPSS 22.0 software
(IBM Corp.). For data conforming to normal distribution and
homogeneity of variance, the paired t-test was used to compare
paired samples, while the unpaired t-test was used for comparisons
between two groups, and the differences among more than two groups
were compared by one-way ANOVA followed by Tukey's post-hoc test.
Patients with liver cancer were classified into either a low
expression group or a high expression group based on the median
expression value of LOXL1-AS1 (median expression value, 1.5) in
tumor samples. Kaplan-Meier survival plots were generated using the
log-rank test. Spearman's correlation analysis was utilized to
analyze the correlations among genes in liver cancer tissues.
P<0.05 was considered to indicate a statistically significant
difference.
Results
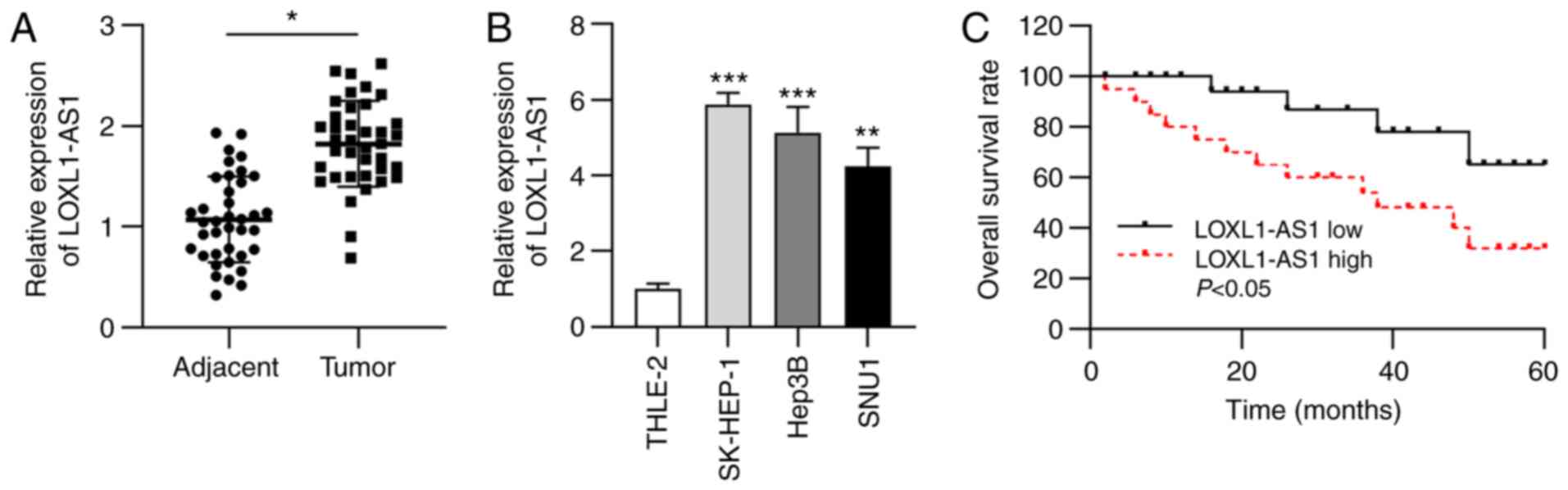
LOXL1-AS1 expression is upregulated in
liver cancer tissues and cells
Abnormal expression of LOXL1-AS1 has been previously
reported in breast cancer and lung adenocarcinoma (15,16);
therefore, the present study aimed to test the expression status of
LOXL1-AS1 in liver cancer. Initially, RT-qPCR analysis was
performed to determine LOXL1-AS1 expression in liver cancer
tissues. The data revealed that LOAL1-AS1 expression in liver
cancer tissues was significantly upregulated compared with that in
corresponding normal tissues (Fig.
1A). Furthermore, LOXL1-AS1 expression was measured in liver
cancer cell lines. RT-qPCR analysis indicated significant
upregulation of LOXL1-AS1 expression in liver cancer cell lines
(SK-HEP-1, Hep3B and SNU1) compared with that in the normal human
hepatic cell line (THLE-2) (Fig.
1B). Subsequently, liver cancer samples were grouped into a
high LOXL1-AS1 expression group and a low expression group.
Kaplan-Meier analysis indicated that higher LOXL1-AS1 expression
was associated with a lower overall survival rate in patients with
liver cancer (Fig. 1C).
LOXL1-AS1 mediates glucose metabolism,
proliferation, migration and epithelial-mesenchymal transition
(EMT) in liver cancer cells
To probe the biological function of LOXL1-AS1 in
liver cancer, liver cancer cells (SK-HEP-1 and Hep3B, which showed
higher LOXL1-AS1 expression) were transfected with sh-LOXL1-AS1 or
sh-NC. The results revealed that LOXL1-AS1 expression was
significantly downregulated after transfection with sh-LOXL1-AS1
compared with sh-NC (Fig. 2A).
Additionally, lncRNA LOXL1-AS1 is the antisense strand of LOXL1.
RT-qPCR and western blot results suggested that LOXL1 mRNA and
protein expression in liver cancer cells were not affected by
LOXL1-AS1 silencing (Fig. S1A).
Previous evidence has demonstrated that the metabolism of cancer
cells is reprogrammed to maintain cell survival and growth
(22). Therefore, the present study
investigated whether LOXL1-AS1 influenced glucose metabolism in
liver cancer cells. SK-HEP-1 and Hep3B cells were treated with
different concentrations of glucose (0, 5 and 20 mM). Through
glucose uptake assays, it was revealed that LOXL1-AS1 silencing
significantly inhibited the glucose uptake of liver cancer cells
treated with 5 and 20 mM glucose (Fig.
2B), indicating that LOXL1-AS1 participated in the progression
of glucose metabolism in liver cancer. Subsequently, the effects of
LOXL1-AS1-knockdown on liver cancer progression were investigated.
LOXL1-AS1-knockdown triggered significant decreases in cell
viability and proliferation, as demonstrated by MTT and colony
formation assays (Fig. 2C-E).
Moreover, the results from the wound healing assay indicated that
the migration of cells was significantly suppressed after LOXL1-AS1
silencing (Fig. 2F). Since EMT
serves a key role in tumor metastasis, western blot analysis was
conducted to assess the effects of LOXL1-AS1 on the expression
levels of EMT markers (E-cadherin and N-cadherin) in liver cancer
cells. The results revealed that LOXL1-AS1-knockdown significantly
increased E-cadherin expression but impaired N-cadherin expression
(Fig. 2G), suggesting the regulatory
role of LOXL1-AS1 on cell migration.
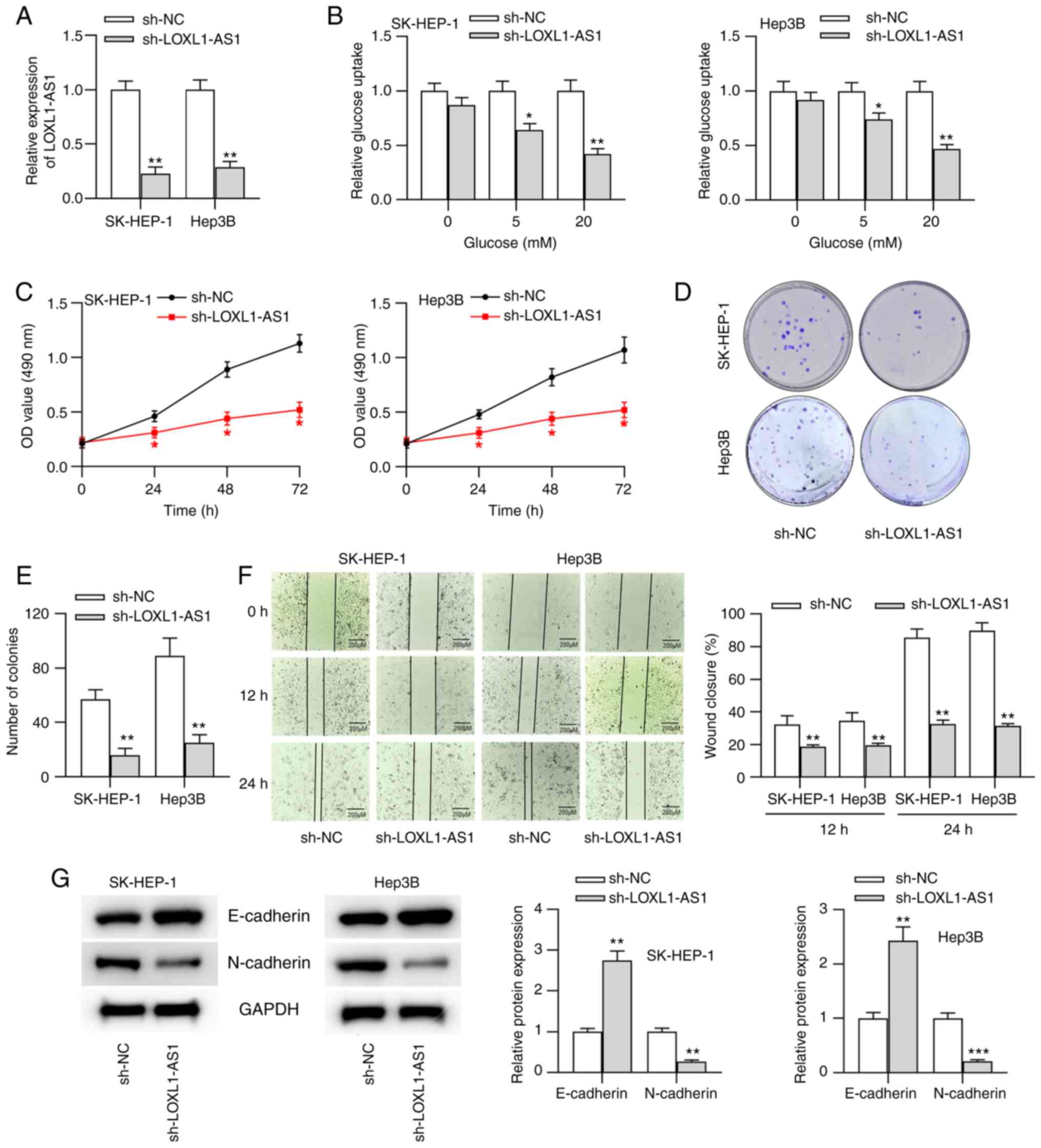 | Figure 2.LOXL1-AS1 mediates glucose
metabolism, proliferation, migration and EMT in liver cancer cells.
(A) sh-NC or sh-LOXL1-AS1 were transfected into SK-HEP-1 and Hep3B
cells, and the transfection efficacy was tested by reverse
transcription-quantitative PCR analysis. (B) Glucose uptake ability
in transfected liver cancer cells was examined by a glucose uptake
assay. Cell viability and proliferation were detected through (C)
MTT and (D and E) colony formation assays, respectively. (F) Cell
migration was evaluated using a wound healing assay (scale bar, 200
µm). (G) Protein expression levels of EMT markers (E-cadherin and
N-cadherin) were measured by western blot analysis. *P<0.05,
**P<0.01 and ***P<0.001 vs. sh-NC. sh, short hairpin RNA; NC,
negative control; LOXL1-AS1, LOXL1-antisense RNA 1; OD, optical
density. |
LOXL1-AS1 functions as a sponge of
miR-377-3p
As lncRNAs have been reported to act as competitive
endogenous RNAs (ceRNAs) to sponge miRNAs in numerous types of
cancer, such as oral squamous cell carcinoma and bladder urothelial
carcinoma (23,24), the present study aimed to explore
whether LOXL1-AS1 interacted with miRNA to affect liver cancer
progression. A total of 29 potential miRNAs were identified that
may bind to LOXL1-AS1 using StarBase under pancancer conditions (4
cancer types). RNA pull-down assays indicated that miR-377-3p was
the most significantly enriched in RNA complexes pulled down by the
Bio-LOXL1-AS1 probe among all candidate miRNAs (the 5 genes that
showed enrichment in pull-down products are shown in Fig. 3A). Thus, miR-377-3p was selected for
subsequent assays. The predicted binding site between LOXL1-AS1 and
miR-377-3p is shown in Fig. 3B.
miR-377-3p expression was significantly downregulated in liver
cancer cell lines compared with in normal liver cells (Fig. 3C). Additionally, miR-377-3p
expression in cells transfected with miR-377-3p mimics was assessed
by RT-q-PCR (Fig. 3D). To further
validate the binding capacity between miR-377-3p and LOXL1-AS1 in
liver cancer cells, a luciferase reporter assay was performed. The
results revealed that the luciferase activity of LOXL1-AS1-Wt
reporters was significantly weakened by miR-377-3p mimics in liver
cancer cells, while that of LOXL1-AS1-Mut reporters remained
unchanged (Fig. 3E), indicating that
miR-377-3p bound to LOXL1-AS1 in liver cancer cells. In addition,
LOXL1-AS1-knockdown significantly increased miR-377-3p expression
in liver cancer cells (Fig. 3F).
Furthermore, the RT-qPCR results suggested that miR-377-3p
expression in liver cancer tissues was significantly downregulated
compared with that in adjacent non-tumor tissues (Fig. 3G). Moreover, a negative correlation
between miR-377-3p and LOXL1-AS1 expression in liver cancer tissues
was observed using Spearman's correlation analysis (Fig. 3H).
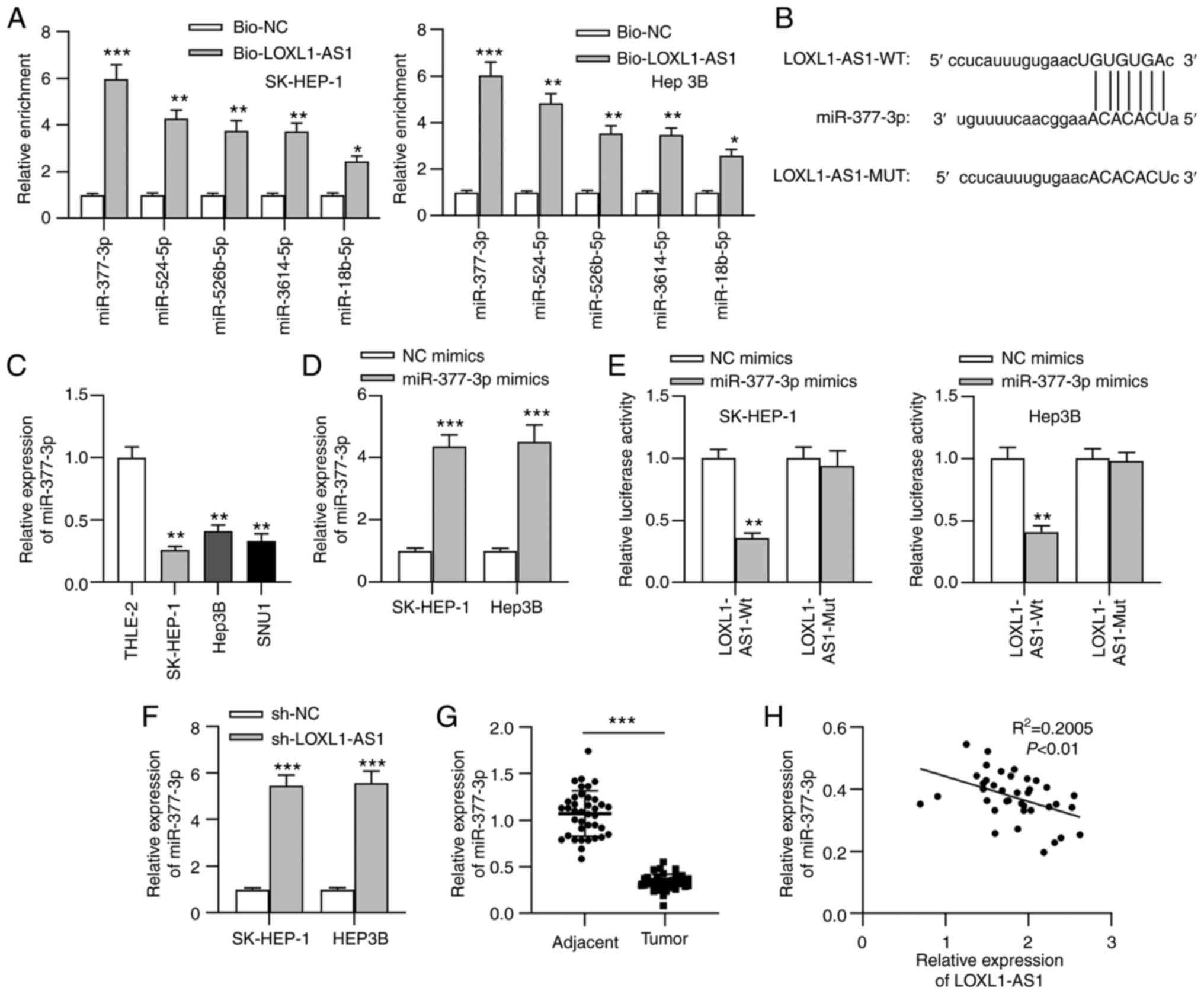 | Figure 3.LOXL1-AS1 functions as a sponge of
miR-377-3p. (A) A total of 29 miRNAs that may bind to LOXL1-AS1
were obtained from StarBase, and only 5 miRNAs showed enrichment,
with miR-377-3p exhibiting the most significant binding capacity to
LOXL1-AS1 through an RNA pull-down assay. *P<0.05, **P<0.01
and ***P<0.001 vs. Bio-NC. (B) Bioinformatics analysis predicted
the binding site between LOXL1-AS1 and miR-377-3p. (C) miR-377-3p
expression in liver cancer cell lines and a normal liver cell line
was measured by RT-qPCR. **P<0.01 vs. THLE-2. (D) SK-HEP-1 and
Hep3B cells were transfected with miR-377-3p mimics or NC mimics,
and RT-qPCR analysis was used to test the transfection efficacy.
(E) Binding ability between LOXL1-AS1 and miR-377-3p in liver
cancer cells was assessed using a luciferase reporter assay.
**P<0.01 and ***P<0.001 vs. NC mimics. (F) RT-qPCR analysis
measured miR-377-3p expression in liver cancer cells transfected
with sh-LOXL1-AS1 or sh-NC. ***P<0.001 vs. sh-NC. (G) RT-qPCR
data of miR-377-3p expression in liver cancer tissues and adjacent
non-tumor tissues. ***P<0.001. (H) Spearman's correlation
analysis assessed the correlation between LOXL1-AS1 and miR-377-3p
expression in liver cancer tissues. RT-qPCR, reverse
transcription-quantitative PCR; LOXL1-AS1, LOXL1-antisense RNA 1;
sh, short hairpin RNA; NC, negative control; miR, microRNA; WT,
wild-type; MUT, mutant; Bio, biotinylated. |
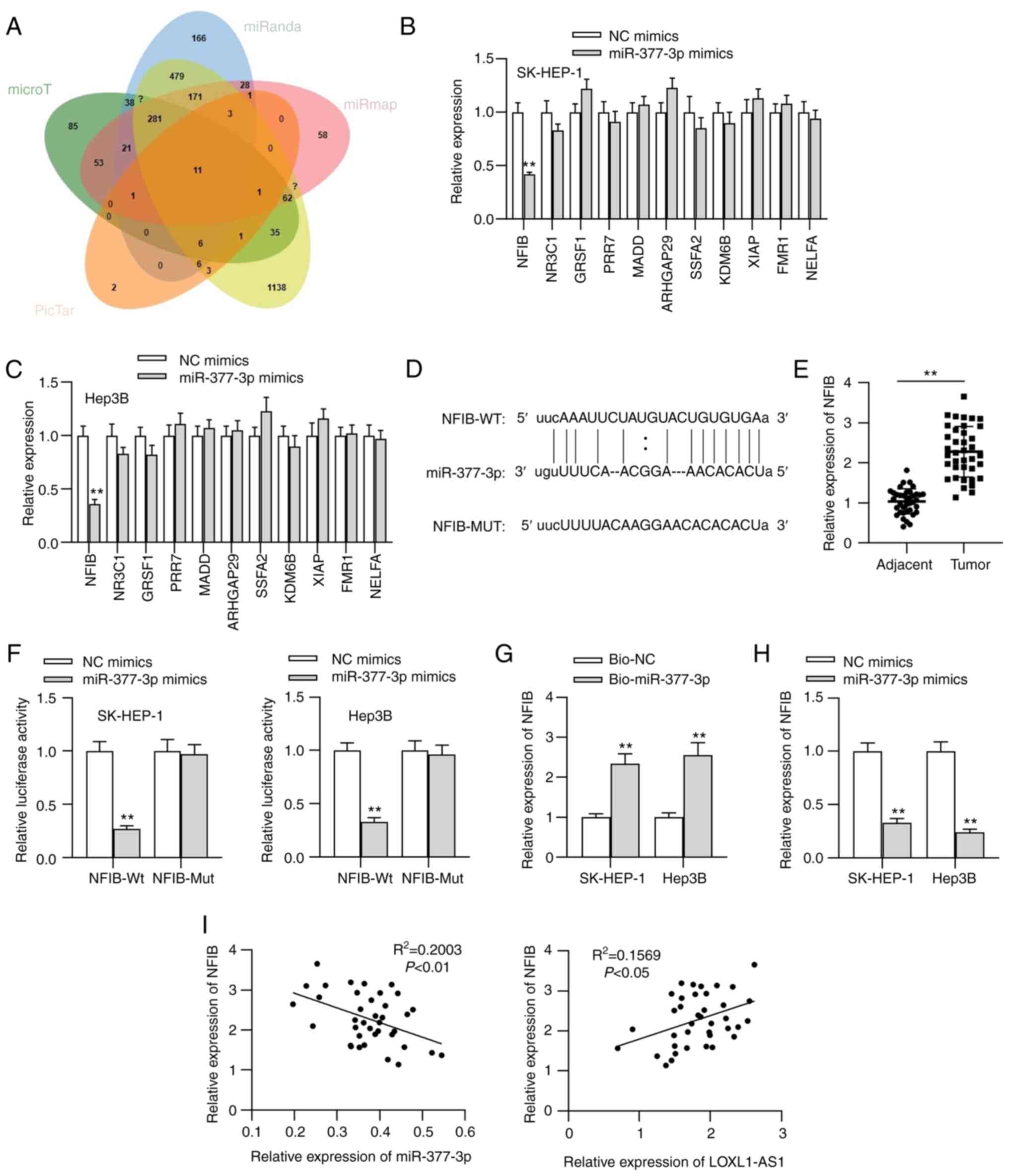
NFIB is targeted by miR-377-3p
To further investigate the regulatory mechanism of
LOXL1-AS1 in liver cancer cells, 11 underlying target genes of
miR-377-3p were identified using online tools (Fig. 4A). As shown in Fig. 4B and C, the level of NFIB presented
with a significant decrease compared with the levels of the other
mRNAs upon miR-377-3p overexpression. NFIB is a transcription
factor that has been implicated in driving a highly migratory
phenotype by enhancing cell-cell adhesion and motility (25). Therefore, NFIB was chosen as the
object of further study. The putative binding site between NFIB and
miR-377-3p was predicted using StarBase (Fig. 4D). The upregulation of NFIB
expression in liver cancer tissues was verified through RT-qPCR
(Fig. 4E). The association between
NFIB and miR-377-3p was further explored in liver cancer cells
through luciferase reporter and RNA pull-down assays. As shown in
Fig. 4F and G, the luciferase
activity of NFIB-Wt reporters was significantly decreased by
miR-377-3p overexpression, and NFIB was significantly enriched in
the RNA complexes pulled down by the Bio-miR-377-3p probe,
confirming the interaction between NFIB and miR-377-3p in liver
cancer cells. Additionally, miR-377-3p mimics significantly
decreased NFIB expression in liver cancer cells (Fig. 4H). Moreover, a negative correlation
between NFIB and miR-377-3p expression, as well as a positive
correlation between NFIB and LOXL1-AS1 expression, was observed in
clinical liver cancer tissues (Fig.
4I).
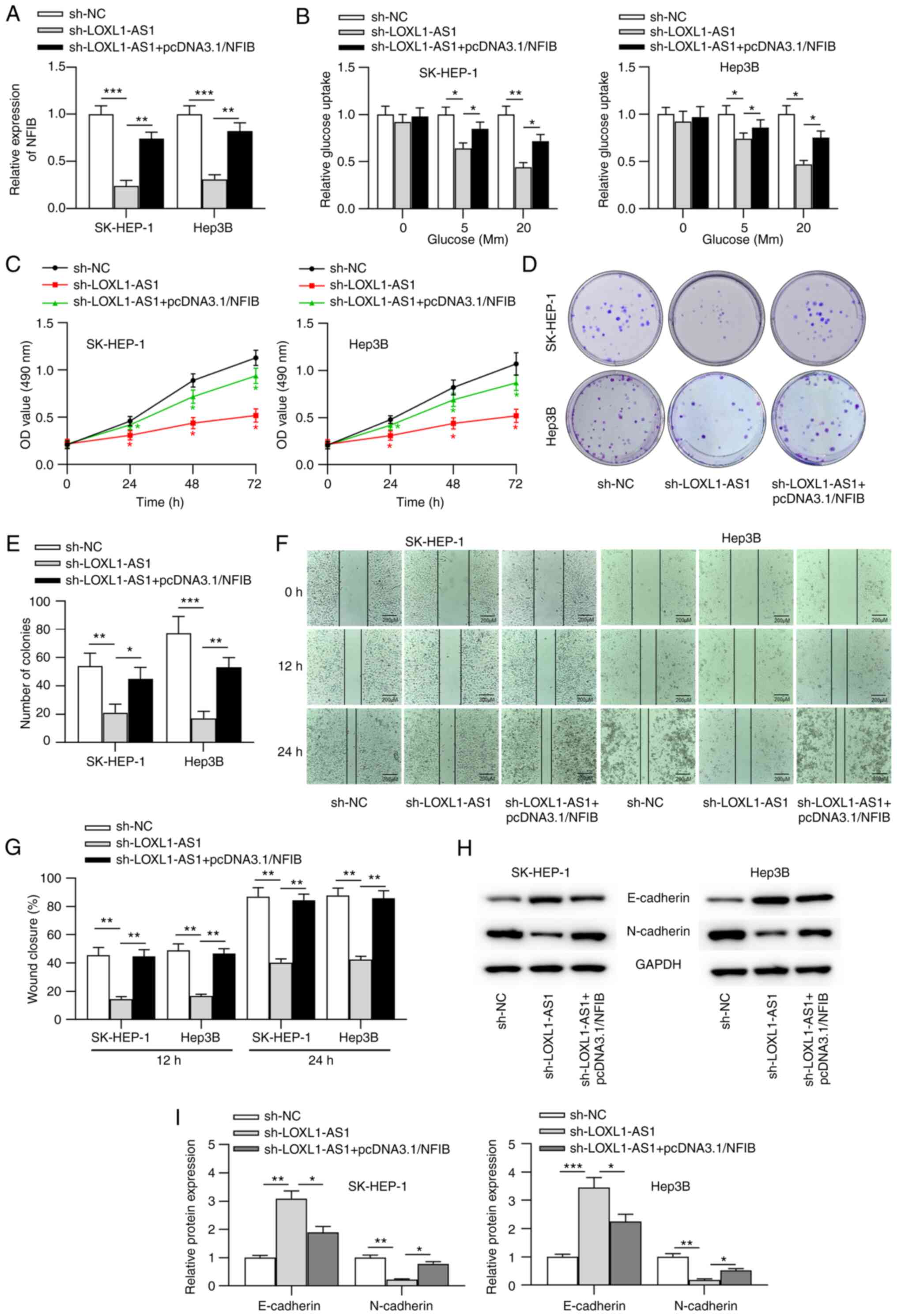
LOXL1-AS1 promotes liver cancer
cellular malignancy via the miR-377-3p/NFIB axis
To determine whether LOXL1-AS1 mediated the cellular
processes of liver cancer via the regulation of the miR-377-3p/NFIB
axis, follow-up rescue assays were performed. First, pcDNA3.1/NFIB
was transfected into liver cancer cells. RT-qPCR results indicated
that NFIB expression was significantly increased in liver cancer
cells following transfection with pcDNA3.1/NFIB (Fig. S1B). Subsequently, sh-LOXL1-AS1 alone
or sh-LOXL1-AS1 with pcDNA3.1/NFIB were transfected into liver
cancer cells. RT-qPCR analysis revealed that sh-LOXL1-AS1
significantly inhibited NFIB expression, which was counteracted by
NFIB overexpression in liver cancer cells (Fig. 5A). Moreover, the repression of
glucose metabolism caused by sh-LOXL1-AS1 was reversed by NFIB
overexpression in liver cancer cells (Fig. 5B). Moreover, overexpression of NFIB
counteracted the inhibitory influence of LOXL1-AS1-knockdown on the
viability, proliferation, and migration of liver cancer cells
(Fig. 5C-G). The western blot
analysis results suggested that NFIB overexpression abolished the
LOXL1-AS1 silencing-mediated repression of the EMT process
(Fig. 5H and I).
Discussion
Previously, Liu et al (26) revealed that LOXL1-AS1 expression was
upregulated in hepatocellular carcinoma tissues and promoted
hepatocellular carcinoma cell proliferation, migration and
invasion. In the present study, the current data confirmed the
upregulation of LOXL1-AS1 expression in liver cancer tissues and
cells, indicating that LOXL1 may act as an oncogene in liver cancer
cellular processes. Additionally, the present study suggested that
high LOXL1-AS1 expression predicts an adverse prognosis in patients
with liver cancer. LOXL1-AS1 is an antisense strand of LOXL1. The
present study revealed that LOXL1 mRNA and protein expression was
not affected by lncRNA LOXL1-AS1 in liver cancer cells. Previous
studies have demonstrated that LOXL1-AS1 promotes cancer
progression by promoting cancer cell proliferation, migration and
invasion (27,28). In the current study, LOXL1-AS1 was
revealed to promote liver cancer progression by facilitating
glucose metabolism, proliferation, migration and EMT in liver
cancer cells.
miRNAs, a class of small, single-stranded RNAs 20–22
nucleotides in length, are known to modulate the expression of
target genes at the post-transcriptional level (29). Increasing studies have demonstrated
that lncRNAs act as ceRNAs to regulate mRNAs by competitively
binding to miRNAs, thus affecting the progression of various types
of cancer, including liver cancer (23,24,30).
There is evidence suggesting that LOXL1-AS1 serves as a ceRNA to
mediate the development of cancer (31,32). In
the present study, miR-377-3p was predicted as the downstream
molecule of LOXL1-AS1 using online tools. miR-377-3p has been
reported to be a tumor suppressor in several types of human cancer.
For example, miR-377-3p inhibits the development of ovarian cancer
by repressing cell proliferation and invasion (33). A high level of miR-377-3p is
associated with longer overall survival of patients with breast
cancer (34). The present study
revealed that miR-377-3p expression was downregulated in liver
cancer tissues and cells. The current mechanistic experimental
results verified the binding capacity between miR-377-3p and
LOXL1-AS1 in liver cancer cells and the negative correlation of
miR-377-3p and LOXL1-AS1 expression in liver cancer tissues. In
addition, miR-377-3p expression was upregulated by LOXL1-AS1
silencing. The present results suggested that LOXL1-AS1 served as a
miR-377-3p sponge.
Multiple transcription factors have been reported to
participate in the metastatic cascade (35). NFIB is a transcription factor that
belongs to the NFI family, which regulates development and cellular
differentiation in several types of tissue (36). Bioinformatics analysis predicted that
NFIB was a potential target gene of miR-377-3p. Recent studies have
suggested that NFIB exerts carcinogenic effects on the progression
of diverse types of cancer. For example, NFIB-knockdown suppresses
cell proliferation, migration and EMT in gastric cancer (37). Additionally, NFIB exhibits an
oncogenic function in colorectal cancer (38). The present study revealed that the
NFIB 3′-UTR was directly targeted by miR-377-3p in liver cancer.
Furthermore, NFIB expression was negatively modulated by miR-377-3p
in liver cancer cells. A positive correlation was identified
between NFIB and LOXL1-A1 expression, as well as a negative
correlation between NFIB and miR-377-3p expression. LOXL1-AS1
upregulated NFIB by competitively binding to miR-377-3p in liver
cancer cells. Rescue assays indicated that NFIB overexpression
countervailed the inhibitory influence of LOXL1-AS1 silencing on
liver cancer cellular processes.
In conclusion, to the best of our knowledge, the
present study was the first to propose and verify that LOXL1-AS1
expression was upregulated in liver cancer tissues and cells, and
that high LOXL1-AS1 expression predicted a poor prognosis for
patients with liver cancer. The current study innovatively provided
evidence that the LOXL1-AS1/miR-377-3p/NFIB axis may aggravate the
cellular processes of liver cancer, which may provide novel
insights for the clinical treatment of liver cancer.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WY and YD participated in the literature search,
analysis and interpretation of the data, and the writing of the
manuscript. Both authors confirm the authenticity of the raw data
and have read and approved the final manuscript.
Ethics approval and consent to
participate
The present study received approval from the Ethics
Committee of the Affiliated Hospital of Yangzhou University
(approval no. 2020-004). All patients provided written informed
consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ding LJ, Li Y, Wang SD, Wang XS, Fang F,
Wang WY, Lv P, Zhao DH, Wei F and Qi L: Long noncoding RNA
lncCAMTA1 promotes proliferation and cancer stem cell-like
properties of liver cancer by inhibiting CAMTA1. Int J Mol Sci.
17:16172016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gan HY, Li N, Zhang Q and Feng ZZ:
Silencing FOXA1 gene regulates liver cancer cell apoptosis and cell
proliferation. Eur Rev Med Pharmacol Sci. 22:397–404.
2018.PubMed/NCBI
|
|
3
|
Petruzziello A: Epidemiology of hepatitis
B virus (HBV) and Hepatitis C virus (HCV) related hepatocellular
carcinoma. Open Virol J. 12:26–32. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Robert J: Biology of cancer metastasis.
Bull Cancer. 100:333–342. 2013.(In French). View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Abdel-Misih SR and Bloomston M: Liver
anatomy. Surg Clin North Am. 90:643–53. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lu WP and Dong JH: Hepatectomy for
hepatocellular carcinoma in the era of liver transplantation. World
J Gastroenterol. 20:9237–9244. 2014.PubMed/NCBI
|
|
7
|
Islami F, Miller KD, Siegel RL, Fedewa SA,
Ward EM and Jemal A: Disparities in liver cancer occurrence in the
United States by race/ethnicity and state. CA Cancer J Clin.
67:273–289. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ye Y, Shen A and Liu A: Long non-coding
RNA H19 and cancer: A competing endogenous RNA. Bull Cancer.
106:1152–1159. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fu XM, Guo W, Li N, Liu HZ, Liu J, Qiu SQ,
Zhang Q, Wang LC, Li F and Li CL: The expression and function of
long noncoding RNA lncRNA-ATB in papillary thyroid cancer. Eur Rev
Med Pharmacol Sci. 21:3239–3246. 2017.PubMed/NCBI
|
|
10
|
Misawa A, Takayama K, Urano T and Inoue S:
Androgen-induced long noncoding RNA (lncRNA) SOCS2-AS1 promotes
cell growth and inhibits apoptosis in prostate cancer cells. J Biol
Chem. 291:17861–17880. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cui C, Zhai D, Cai L, Duan Q, Xie L and Yu
J: Long noncoding RNA HEIH promotes colorectal cancer tumorigenesis
via counteracting miR-939-mediated transcriptional repression of
Bcl-xL. Cancer Res Treat. 50:992–1008. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang H, Huo X, Yang XR, He J, Cheng L,
Wang N, Deng X, Jin H, Wang N, Wang C, et al: STAT3-mediated
upregulation of lncRNA HOXD-AS1 as a ceRNA facilitates liver cancer
metastasis by regulating SOX4. Mol Cancer. 16:1362017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jiao Y, Li Y, Ji B, Cai H and Liu Y:
Clinical value of lncRNA LUCAT1 expression in liver cancer and its
potential pathways. J Gastrointestin Liver Dis. 28:439–447. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang Z, Wang S, Yang F, Meng Z and Liu Y:
LncRNA ROR1AS1 high expression and its prognostic significance in
liver cancer. Oncol Rep. 43:55–74. 2020.PubMed/NCBI
|
|
15
|
Li GH, Yu JH, Yang B, Gong FC and Zhang
KW: LncRNA LOXL1-AS1 inhibited cell proliferation, migration and
invasion as well as induced apoptosis in breast cancer via
regulating miR-143-3p. Eur Rev Med Pharmacol Sci. 23:10400–10409.
2019.PubMed/NCBI
|
|
16
|
Li W, Zhang B, Jia Y, Shi H, Wang H, Guo Q
and Li H: LncRNA LOXL1-AS1 regulates the tumorigenesis and
development of lung adenocarcinoma through sponging miR-423-5p and
targeting MYBL2. Cancer Med. 9:689–699. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu Y, Guo C, Li F and Wu L: LncRNA
LOXL1-AS1/miR-28-5p/SEMA7A axis facilitates pancreatic cancer
progression. Cell Biochem Funct. 38:58–65. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li M, Cai O and Tan S: LOXL1-AS1 drives
the progression of gastric cancer via regulating miR-142-5p/PIK3CA
Axis. Onco Targets Ther. 12:11345–11357. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xie N, Fei X, Liu S, Liao J and Li Y:
LncRNA LOXL1-AS1 promotes invasion and proliferation of
non-small-cell lung cancer through targeting miR-324-3p. Am J
Transl Res. 11:6403–6412. 2019.PubMed/NCBI
|
|
20
|
Kee KM, Wang JH, Lin CY, Wang CC, Cheng YF
and Lu SN: Validation of the 7th edition TNM staging system for
hepatocellular carcinoma: An analysis of 8,828 patients in a single
medical center. Dig Dis Sci. 58:2721–2728. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li Z and Zhang H: Reprogramming of
glucose, fatty acid and amino acid metabolism for cancer
progression. Cell Mol Life Sci. 73:377–392. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhao J, Bai X, Feng C, Shang X and Xi Y:
Long non-coding RNA HCP5 facilitates cell invasion and
epithelial-mesenchymal transition in oral squamous cell carcinoma
by miR-140-5p/SOX4 Axis. Cancer Manag Res. 11:10455–10462. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang J, Zhang C, Wu Y, He W and Gou X:
Identification and analysis of long non-coding RNA related miRNA
sponge regulatory network in bladder urothelial carcinoma. Cancer
Cell Int. 19:3272019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Denny SK, Yang D, Chuang CH, Brady JJ, Lim
JS, Grüner BM, Chiou SH, Schep AN, Baral J, Hamard C, et al: Nfib
promotes metastasis through a widespread increase in chromatin
accessibility. Cell. 166:328–342. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu J, Zhai C, Liu D and Liu J: The long
noncoding RNA LOXL1-AS1 promotes the proliferation, migration, and
invasion in hepatocellular carcinoma. Anal Cell Pathol (Amst).
2020:41820922020.PubMed/NCBI
|
|
27
|
Yang X, Xing G, Liu S, Li B, He Y and Wang
F: LncRNA LOXL1-AS1 promotes endometrial cancer progression by
sponging miR-28-5p to upregulate RAP1B expression. Biomed
Pharmacother. 125:1098392020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xue F, Xu YH, Shen CC, Qin ZL and Zhou HB:
Non-coding RNA LOXL1-AS1 exhibits oncogenic activity in ovarian
cancer via regulation of miR-18b-5p/VMA21 axis. Biomed
Pharmacother. 125:1095682020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Meng Q and Lan D: A review on
muscle-specific microRNAs as the biomarker for Duchenne muscular
dystrophy. Zhongguo Dang Dai Er Ke Za Zhi. 21:1148–1152. 2019.(In
Chinese). PubMed/NCBI
|
|
30
|
He J, Zhao H, Deng D, Wang Y, Zhang X,
Zhao H and Xu Z: Screening of significant biomarkers related with
prognosis of liver cancer by lncRNA-associated ceRNAs analysis. J
Cell Physiol. 235:2464–2477. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang B, Zhou M, Zou L, Miao J, Wang Y, Li
Y, Lu S and Yu J: Long non-coding RNA LOXL1-AS1 acts as a ceRNA for
miR-324-3p to contribute to cholangiocarcinoma progression via
modulation of ATP-binding cassette transporter A1. Biochem Biophys
Res Commun. 513:827–833. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Long B, Li N, Xu XX, Li XX, Xu XJ, Liu JY
and Wu ZH: Long noncoding RNA LOXL1-AS1 regulates prostate cancer
cell proliferation and cell cycle progression through miR-541-3p
and CCND1. Biochem Biophys Res Commun. 505:561–568. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tang L, Yang B, Cao X, Li Q, Jiang L and
Wang D: MicroRNA-377-3p inhibits growth and invasion through
sponging JAG1 in ovarian cancer. Genes Genomics. 41:919–926. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang X, Chen T, Zhang Y, Zhang N, Li C, Li
Y, Liu Y, Zhang H, Zhao W, Chen B, et al: Long noncoding RNA
Linc00339 promotes triple-negative breast cancer progression
through miR-377-3p/HOXC6 signaling pathway. J Cell Physiol.
234:13303–13317. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ell B and Kang Y: Transcriptional control
of cancer metastasis. Trends Cell Biol. 23:603–611. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wilczynska KM, Singh SK, Adams B, Bryan L,
Rao RR, Valerie K, Wright S, Griswold-Prenner I and Kordula T:
Nuclear factor I isoforms regulate gene expression during the
differentiation of human neural progenitors to astrocytes. Stem
Cells. 27:1173–1181. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wu C, Zhu X, Liu W, Ruan T, Wan W and Tao
K: NFIB promotes cell growth, aggressiveness, metastasis and EMT of
gastric cancer through the Akt/Stat3 signaling pathway. Oncol Rep.
40:1565–1573. 2018.PubMed/NCBI
|
|
38
|
Sun L, Fang Y, Wang X, Han Y, Du F, Li C,
Hu H, Liu H, Liu Q, Wang J, et al: miR-302a inhibits metastasis and
cetuximab resistance in colorectal cancer by targeting NFIB and
CD44. Theranostics. 9:8409–8425. 2019. View Article : Google Scholar : PubMed/NCBI
|