Introduction
Breast cancer is undoubtedly one of the most severe
heterogeneous diseases affecting women worldwide, resulting in
>500,000 deaths every year (1).
Various efforts are being devoted to overcoming this critical
issue, culminating in prevention, diagnosis, and treatment. More
specifically, breast cancer can bes treated by surgical,
chemotherapeutic or radiotherapeutic methods that differ in their
actions (2,3). Nevertheless, these approaches are
effective only at the earlier stages, and because of their
disadvantages, including high toxicity, side effects and
inefficiencies for long-term use, they have failed to realize the
cancer therapy (4).
Meanwhile, natural drugs have emerged as alternative
methods to treat breast cancer because of their low side effects,
high selectivity, safety and effectiveness. Some of the most
promising natural anticancer drugs are phenolic compounds (5). The antitumor efficiency of natural
phenolics has been associated with their strong anti-inflammatory
and antioxidant activities and their ability to moderate molecular
targets as well as signaling pathways, which are associated with
cell differentiation (6). Rosmanol
is a phenolic diterpene obtained from various plants, including
Salvia officinalis, Hyptisincana and Rosmarinus. They
have been used as potent antioxidants and anti-inflammatories due
to their superoxide anion production inhibition capabilities as
well as lipid peroxidation and free radical scavenging activities
(7). Moreover, Rosmanol inhibited
iNOS and COX-2 gene and protein expression as well as their
downstream products, NO and PGE2, respectively (8), which causes decreased translocation of
nuclear factor-κB (NF-κB) subunits (9). As previously reported, Rosmanol also
inhibits the proliferation of human colon and neuroblastoma cancer
(10).
In total, ~80% of breast cancers express the
estrogen receptor α (ERα) and the progesterone receptor (PR). ER is
a steroid hormone receptor, comprised of ERα and ERβ, in which the
ERα content in cells is higher than that in normal breast tissue in
estrogen-dependent breast cancers (11). Overexpression of ERβ and inhibition
of ERα have been shown to have anti-tumor effects against breast
cancer in cells (12).
Moreover, the PI3K/AKT pathway serves an important
role in breast cancer development (13). Estrogen receptor α-positive breast
cancers adapt to hormone deprivation and acquire resistance to
antiestrogen therapies. NF-κB is a downstream component of the
PI3K/Akt pathway and is activated via phosphorylation of IκB kinase
(IKK) (14).
Certain researchers have attempted to assess the
function of JAK2 knockout in the acceleration of cell apoptosis,
autophagy and inhibition of proliferation, and this is an important
research direction for cancer therapy in the future (15–17).
Notably, cytokines and growth factors unanimously use Janus kinase
(JAK) as the signal transducer and further activate the [JAK/signal
transducer and activator of transcription (STAT3)] pathway
transcription (18). Additionally,
STAT3 serves a key role in activating cell proliferation and growth
in cancer cells, such as breast cancer cells (19). In addition to regulating STAT3, JAK2
signaling also regulates p38 and ERK signaling (20). The above are some important
proliferation signals for breast cancer, which promote cell
proliferation and inhibit cell apoptosis.
Despite its intense biological response, to the best
of our knowledge the effects of Rosmanol on breast cancer have not
been previously investigated. Accordingly, the present research
aimed to investigate the effects of Rosmanol on proliferation and
apoptosis in breast cancer, and to further reveal the associated
mechanism by detecting the effects of Rosmanol on the p38,
PI3K/Akt/ERK, and STAT3/JAK2 pathways.
Materials and methods
Cell culture
MCF-10A, MCF-7 and MDA-MB 231 cells were purchased
from the American Type Culture Collection. Normal human breast
MCF-10A cells were cultured in DMEM/F12 (1:1) media supplemented
with 5% horse serum (Gibco; Thermo Fisher Scientific, Inc.), 1.2
mg/ml NaHCO3 (Sigma-Aldrich; Merck KGaA) 500 ng/ml
hydrocortisone (Sangon Biotech Co., Ltd.), 10 µg/ml human insulin
(Gibco; Thermo Fisher Scientific, Inc.), 10 ng/ml hunan recombinant
epidermal growth factor (Sigma-Aldrich; Merck KGaA) and 100 ng/ml
cholera toxin (Sigma-Aldrich; Merck KGaA). The two human breast
cancer cell lines, MCF-7 and MDA-MB 231, were cultured and
maintained in DMEM containing 10% fetal bovine serum (FBS; Gibco,
Thermo Fisher Scientific, Inc). The cells were cultured at 37°C in
a humidified atmosphere of 5% CO2 and allowed to grow to
a confluence of 70–80%.
Cell proliferation assay
MCF-10A, MCF-7 and MDA-MB 231 cells were harvested
and seeded in a 96-well dish to a final concentration of
5×103 cells/well and incubated in DMEM containing 1% FBS
for 24 h. Cells were treated with the indicated concentrations of
Rosmanol (Baoji Herbest Bio-tech Co., Ltd.), then incubated for 48
h. Next, 20 µl MTT (Sigma-Aldrich; Merck KGaA) solution (5 mg/ml)
was added to each well and incubated for 4 h at 37°C. Finally, the
medium was discarded, and 150 µl DMSO was added to each well. The
plates were read at a wavelength of 570 nm using a Varioskan Flash
Multimode Reader (Thermo Fisher Scientific, Inc.). Six duplicate
wells were used for each treatment. The absorbance was measured at
570 nm, results were expressed as a percentage, relative to
solvent-control incubations, and the IC50 values were
determined using non-linear regression analysis (percentage
survival versus concentration).
Annexin V/PI assay for apoptosis
Apoptotic MCF-7 and MDA-MB 231 cells were
investigated using the Annexin V/propidium iodide (PI) apoptosis
detection kit (Beyotime Institute of Biotechnology) followed by
flow cytometry, according to the manufacturer's protocol. MCF-7 and
MDA-MB 231 cells (5×103 cells/well) were cultured in
6-well plates and treated with different concentrations (15, 30 and
60 µM) of Rosmanol for 48 h at 37°C. Cells were harvested by
trypsinization with no EDTA and washed twice with PBS and then
stained with 5 µl Annexin V-FITC (Beyotime Institute of
Biotechnology) and 10 µl PI in 500 µl binding buffer for 15 min at
room temperature (RT) in the dark. Apoptotic cells were determined
by using Cytomics FC 500 flow cytometer (Beckman Coulter Inc.) and
the data were analysed using Cellquest analysis software v.3.0 (BD
Biosciences).
Determination of MCF-7 and MDA-MB 231
cell apoptosis by immunofluorescence
MCF-7 and MDA-MB 231 apoptotic cell death was
observed by DAPI staining. Briefly, cells were treated with 15, 30
and 60 µM Rosmanol for 48 h at 37°C, then fixed with 4%
paraformaldehyde for 20 min at 4°C. Subsequently, cells were washed
with precooled PBS (4°C), twice with TBST (0.05% Tween20), and
cells were stained with DAPI (1 µg/ml) at room temperature in the
dark for 3 min, and washed with PBS. Apoptotic nuclei cells were
observed under a fluorescent microscope (model IX71; Olympus
Corporation).
Determination of MCF-7 and MDA-MB 231
cell cycle distribution
The cell cycle distributions in different phases
after exposure to Rosmanol were analyzed by flow cytometry. In
brief, MCF-7 and MDA-MB 231 cells (5×105 cells/well)
were seeded into 6-well plates and exposed to 15, 30 and 60 µM
Rosmanol for 48 h at 37°C. Further, the cells were harvested,
washed twice with PBS, and fixed with 70% ethanol for 2 h at 4°C.
After that, cells were centrifuged at 3,000 × g for 5 min at 4°C
and washed with PBS, resuspended in 500 µl of buffer containing 0.5
mg/ml RNase and 25 mg/ml propidium iodide (PI, Beyotime Institute
of Biotechnology), and incubated in the dark at 37°C for 15 min.
The distribution of the cell cycle was determined by using Cytomics
FC 500 flow cytometer (Beckman Coulter Inc.), and the data were
analyzed using Cellquest analysis software v.3.0 (BD
Biosciences).
Determination of reactive oxygen
species (ROS) expression in MCF-7 and MDA-MB 231 cells
The generation of ROS was determined with
2,7-dichlorofluorescein diacetate (DCFH-DA) according to the
manufacturer's protocol. Briefly, MCF-7 and MDA-MB 231 cells were
cultured (5×103 cells/well) and then incubated with or
without N-acetyl-cysteine (NAC) for 1 h at 37°C. After the culture
was treated with 15, 30 and 60 µM of Rosmanol for 48 h at 37°C. The
cells were collected, centrifuged at 600 × g for 4 min at room
temperature, washed with PBS, resuspended in PBS containing 10 µM
DCFH-DA (Beyotime Institute of Biotechnology) and incubated in the
dark at RT for 15 min. Subsequently, the cells were washed with PBS
and measured immediately using flow cytometry (Cytomics FC 500,
Beckman Coulter, Inc.) to monitor the formation of the
fluorescent-oxidized derivative of DCFH-DA at an emission
wavelength of 525 nm and an excitation wavelength of 488 nm.
Determination of mitochondrial
membrane expression (MMP) in MCF-7 and MDA-MB 231 cells
The changes induced by Rosmanol in the mitochondrial
membrane were determined using Rhodamine 123 staining (Beyotime
Institute of Biotechnology) according to the manufacturer's
protocol. Briefly, MCF-7 and MDA-MB 231 cells (5×103
cells/well) were seeded in 6-well plates and then treated with or
without NAC and incubated at 37°C for 1 h. The cells were treated
with or without (15, 30 and 60 µM) Rosmanol for 48 h at 37°C and
stained with Rhodamine 123 for 15 min at 37°C. Mitochondrial
membrane potential was detected by Cytomics FC 500 flow cytometer
(Beckman Coulter Inc.) and the data were analyzed using Cellquest
analysis software v.3.0 (BD Biosciences).
Western blot analysis of MCF-7 and
MDA-MB 231 cells
Protein expression regulation by Rosmanol was
analyzed via western blotting and the data were analysed using
Image J software v.1.48 (National Institutes of Health) following a
protocol described previously (21)
with a small modification. In brief, MCF-7 and MDA-MB 231 cells
were seeded and incubated at 37°C for 24 h and were separated into
2 groups. Cells in one group were treated with 15, 30 and 60 µM
Rosmanol for 48 h at 37°C and MCF-7 and MDA-MB 231 cells in the
other group were pre-incubation with the caspase inhibitor
Z-VAD-FMK (50 mM; Sigma-Aldrich; Merck KGaA) at 37°C for 1 h and
then treated with Rosmanol (15, 30 and 60 µM) at 37°C for 48 h. The
cells were then harvested and lysed with RIPA buffer. Afterward,
the insoluble protein lysate was removed by centrifugation at
12,000 × g for 15 min at 4°C. Protein concentrations were
determined using a NanoDrop 1000 (Thermo Fisher Scientific, Inc.).
Proteins (40 µg/lane) were loaded on SDS-PAGE electrophoresis gel
(10 or 12% according to the protein size), and the gel was
transferred onto polyvinylidene fluoride membrane (PVDF). After the
necessary transfer time, the membrane was blocked in 5% (w/v)
non-fat milk and incubated for 1 h at room temperature. The
membranes were incubated with appropriate primary antibodies,
Caspase-3 (cat. no. 9662s), Caspase-8 (cat. no. 4927s), Caspase-9
(cat. no. 9502s), p-PI3k (cat. no. 4228), p-Akt (cat. no. 9271),
Akt (cat. no. 9272), JAK2 (cat. no. 3230), p-JAK2 (cat. no. 3771),
p-ERK (cat. no. 9101), p-p38 (cat. no. 9211), STAT3 (cat. no.
9139), p-STAT3 (cat. no. 9145), Bcl-2 (cat. no. 15071), Bax (cat.
no. 2774), NF-KB (cat. no. 8242), Cox-2 (cat. no. 12282) (all
1:1,000; all Cell Signaling Technology, Inc.) and Cyclin A (cat.
no. sc-271682), Cyclin B1 (cat. no. sc-245), Cyclin D1 (cat. no.
sc-8396), Cyclin E1 (cat. no. sc-377100), ER-α (cat. no. sc8002)
and ER-β (cat. no. sc-373853) (all 1:2,000; all Santa Cruz
Biotechnology) at 4°C overnight and washed three times with a
Tris-buffered saline-Tween solution (TBST) containing 0.05%
Tween20. Finally, the blots were incubated with horseradish
peroxidase-conjugated goat anti-mouse IgG (1:1,000; cat. no.
sc-2005; Santa Cruz Biotechnology, Inc.) and horseradish
peroxidase-conjugated goat anti-rabbit IgG (1:1,000; cat. no.
sc-2004; Santa Cruz Biotechnology, Inc.) for 1 h at RT, washed with
TBST for 30 min, and signals were detected using an ECL plus
chemiluminescence kit on X-ray film (EMD Millipore) in a dark room
or use Tanon 5500 high definition low illumination CCD system
(Tanon Science & Technology Co., Ltd.) to detect blot signals
and TanonImage software v.1.00 (Tanon Science & Technology Co.,
Ltd.) to capture and merge images. Protein bands were quantified by
Image J software v.1.48 (National Institutes of Health).
Transient transfection and luciferase
reporter as say of MCF-10A, MCF-7 and MDA-MB 231 cells
Luciferase assays were performed as previously
described (22). Briefly, transient
transfections were performed using Lipofectamine 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. Cells were seeded into 48-well plates (0.5×103
cells/well) at 37°C for 16 h before being transfected with 100 ng
STAT3-Luc promoter in the presence of 25 ng Renilla
Luciferase control pREP7 vector and incubated at 37°C for 24 h. The
cells were then treated with Rosmanol at concentrations of 0, 15,
30 and 60 µM for 24 h at 37°C. Firefly luciferase activities were
calculated using the Dual-Luciferase reporter assay system (Promega
Corporation) and the ratio of firefly luciferase activity to
Renilla luciferase activity was defined as relative
luciferase activity.
Statistical analysis
All statistical analyses were performed using Origin
Lab software version 8.0 (OriginLab), and statistically significant
differences between groups were determined by one-way ANOVA with
Bonferroni's post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Cytotoxicity of Rosmanol on breast
cancer and normal cell lines
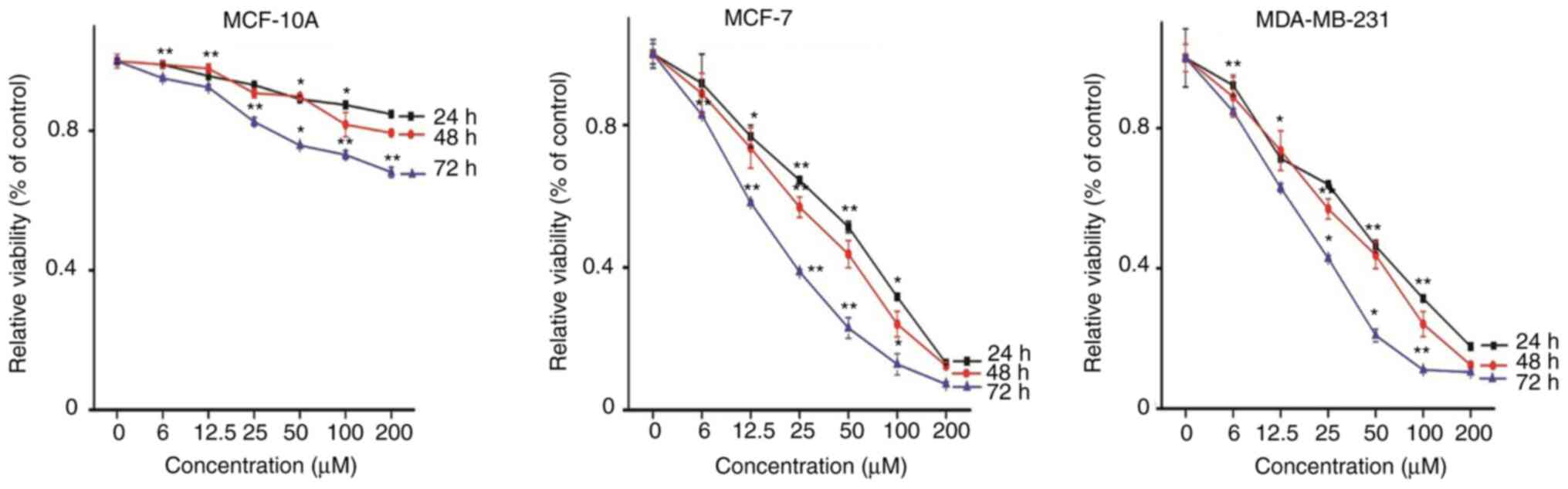
First, the effect of Rosmanol on cell viability was
investigated using the MTT assay. The cells were incubated with 0,
6, 12.5, 25, 50, 100 and 200 µM Rosmanol. As shown in Fig. 1, Rosmanol was able to inhibit the
proliferation of MCF-7 and MDA-MB 231 cells, suggesting that
Rosmanol induced cytotoxicity against human breast cancer cells. It
was revealed that Rosmanol induced cytotoxicity in MCF-7 and MDA-MB
231 cells not only in dose-dependent manner, but also in a
time-dependent manner, with their half maximal inhibitory
concentration IC50 (51, 26 and 19) and (42, 28 and 16)
µM, respectively, for 24, 48 and 72 h. However, Rosmanol did not
exhibit a significant effect on MCF-10A normal breast cells. Thus,
based on its effects on both MCF-7 and MDA-MB 231 cells, incubation
at concentrations of 15, 30 and 60 µM for 48 h was chosen for
subsequent studies.
Rosmanol induces apoptosis in MCF-7
and MDA-MB 231 cells
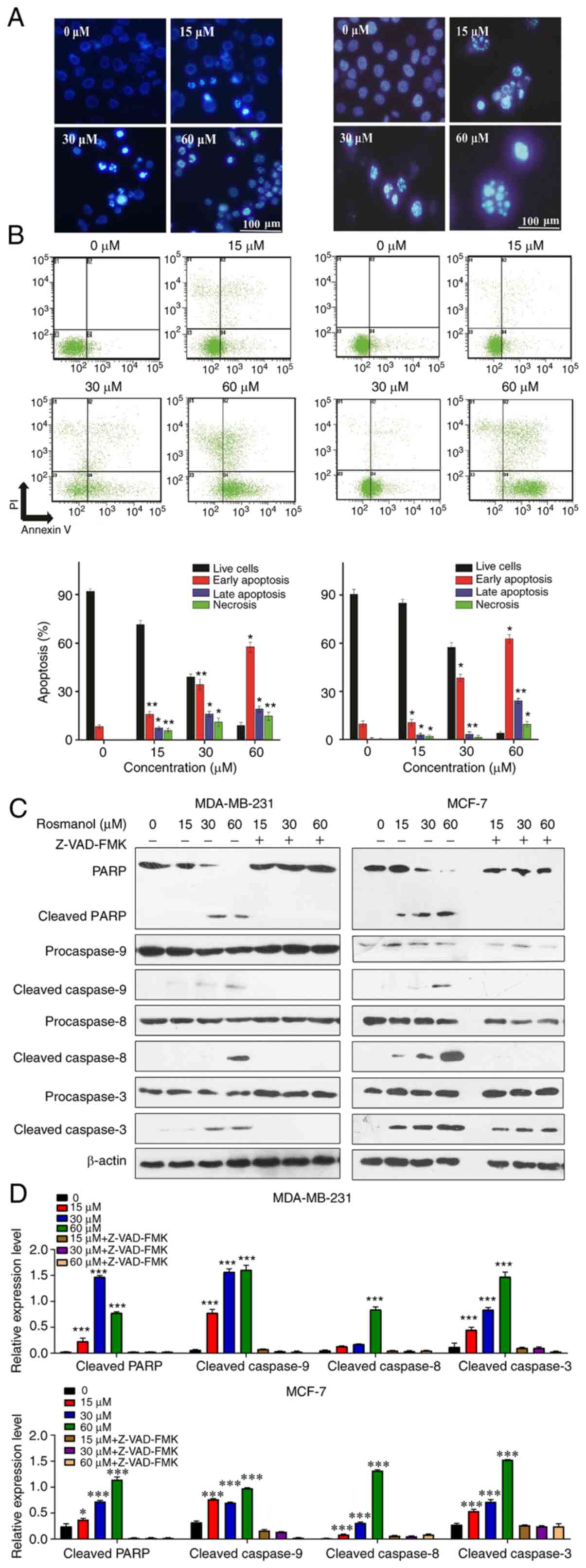
To determine whether the cytotoxicity exerted by
Rosmanol was due to apoptosis induction, morphological observations
were first performed under a microscope. The exposure of MCF-7 and
MDA-MB 231 cells to Rosmanol (15, 30 and 60 µM) for 48 h induced
cell membrane changes along with a shift in their nuclear
morphology, as well as DNA damage compared with untreated cells, as
shown by DAPI staining. Within a certain range, the degree of
apoptosis was more obvious as the Rosmanol concentration increased
(Fig. 2A).
To verify the apoptosis induction by Rosmanol, a
further measurement of apoptosis was determined by annexin V-FITC
and PI double-staining flow cytometry, which can quantitatively
assess early apoptosis (B4), late apoptosis (B2), and necrosis
(B1). As shown in Fig. 2B, the B4
values increased with an increase in the Rosmanol concentration in
both MCF-7 and MDA-MB 231 cells.
To further understand the molecular mechanism by
which Rosmanol mediated apoptosis in MCF-7 and MDA-MB 231 cells,
western blotting was conducted to detect caspases, the critical
regulators of apoptosis (23,24). As
illustrated in Fig. 2C and D,
Rosmanol stimulated the activation of caspase-8, as well as
caspase-9 which led to activated caspase-3 and consequently cleaved
PARP, the major indicator enzyme of apoptosis. Furthermore,
pre-incubation of MCF-7 and MDA-MB 231 cells with the caspase
inhibitor Z-VAD-FMK (50 mM; Sigma-Aldrich; Merck KGaA) at 37°C for
1 h and then treated with Rosmanol (15, 30 and 60 µM) at 37°C for
48 h promoted the survival of the cells. These findings
demonstrated that Rosmanol induced cancer cell apoptosis through
extrinsic mitochondrial pathways.
Cytotoxicity mediated by Rosmanol is
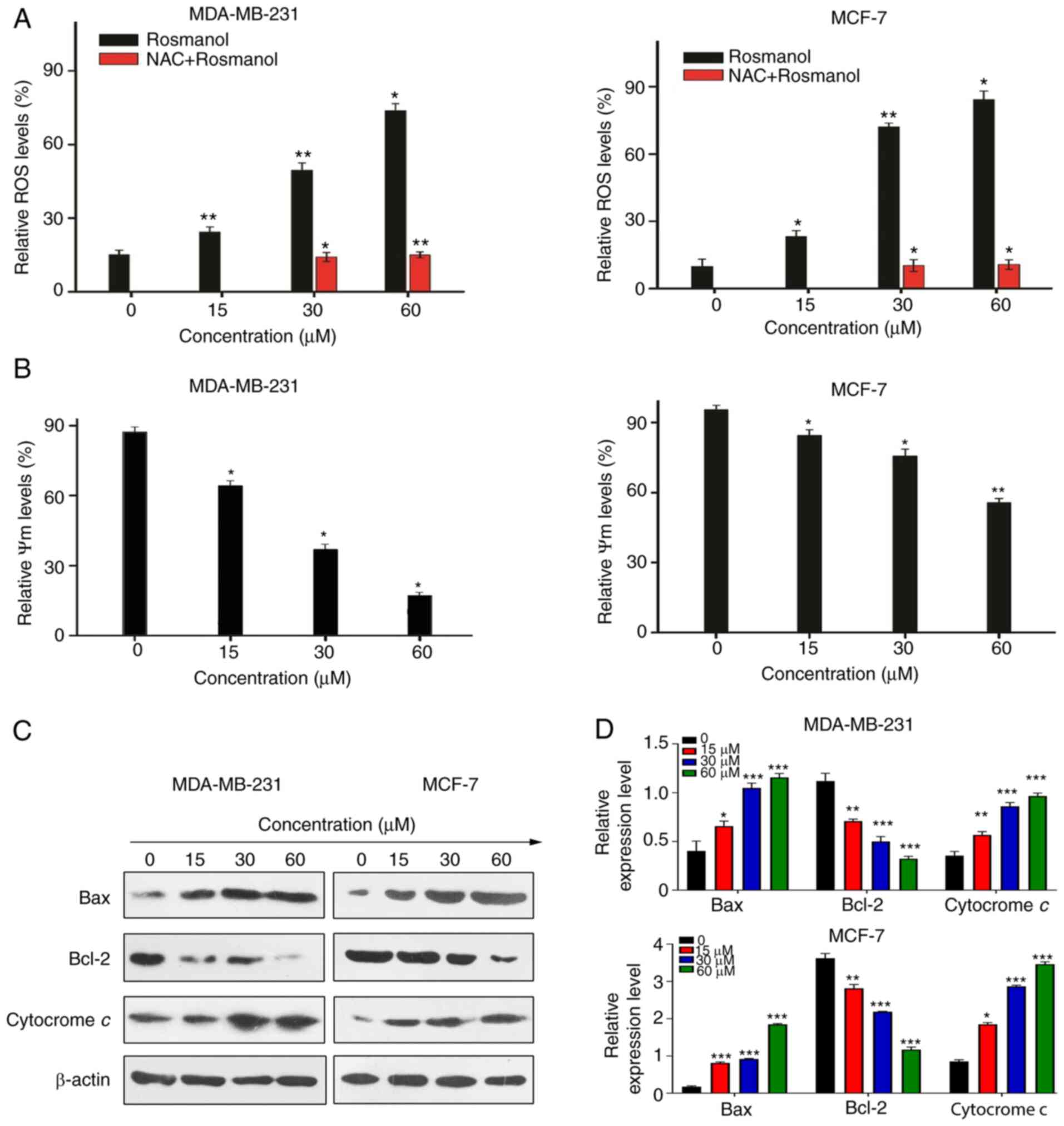
associated with generation of ROS
Immunostaining using DAPI revealed that Rosmanol
induced DNA damage in MCF-7 and MDA-MB 213 cells. Previous studies
reported that DNA damage induced oxidative stress in MCF-7 cells
(25–27), which further resulted in the
production of ROS (28). To evaluate
whether the cytotoxicity mediated by Rosmanol was associated with
the generation of ROS, MCF-7 and MDA-MB 231 cells were treated with
Rosmanol, and then probed with the fluorescent DCF-DA for the
detection and assessment of H2O2 by flow
cytometry in the presence and absence of NAC for 48 h. The results
indicated that the generation of intracellular
H2O2 increased considerably with the
treatment of Rosmanol (0, 15, 30 and 60 µM). In the meantime,
supplementation with NAC rendered ROS production almost unaffected
by Rosmanol (30 and 60 µM), which indicated a role of Rosmanol in
the generation of H2O2, as illustrated in
Fig. 3A. These results demonstrated
that Rosmanol increases ROS in MCF-7 and MDA-MB 231 cells, and that
cytotoxicity is associated with the generation of ROS.
Rosmanol stimulates apoptosis via the
mitochondrial pathway
To validate our hypotheses, the effect of Rosmanol
on mitochondrial depolarization was investigated in MCF-7 and
MDA-MB 231 cells through the employment of Rhodamine 123 staining
flow cytometry. Targeting mitochondria is a novel approach for
cancer therapy because of their ability for stimulating apoptosis.
In in this regard, mitochondrial-mediated apoptosis can be highly
regulated by counterbalancing the expression of pro-apoptotic, as
well as anti-apoptotic, Bcl-2 proteins in which a loss in the
mitochondrial transmembrane potential (Δψm) ensues in the case of
the disruption of these counterbalances (29). As shown in Fig. 3B, Rosmanol treatment of MCF-7 and
MDA-MB 231 cells induced the loss of mitochondrial membrane
potential. This decrease in mitochondrial membrane potential by
Rosmanol was particularly caused by the capability of Rosmanol to
release mitochondrial Cytochrome c, a major factor in the formation
of apoptosomes (9), and in addition,
to induce Bax and inhibit Bcl-2, as shown in Fig. 3C and D. These findings indicate the
role of Rosmanol in stimulating apoptosis via mitochondrial
pathways.
Rosmanol induces activation of
p38/MAPK and prohibition of ERK and PI3K/Akt in MCF-7 and MDA-MB
231 cells
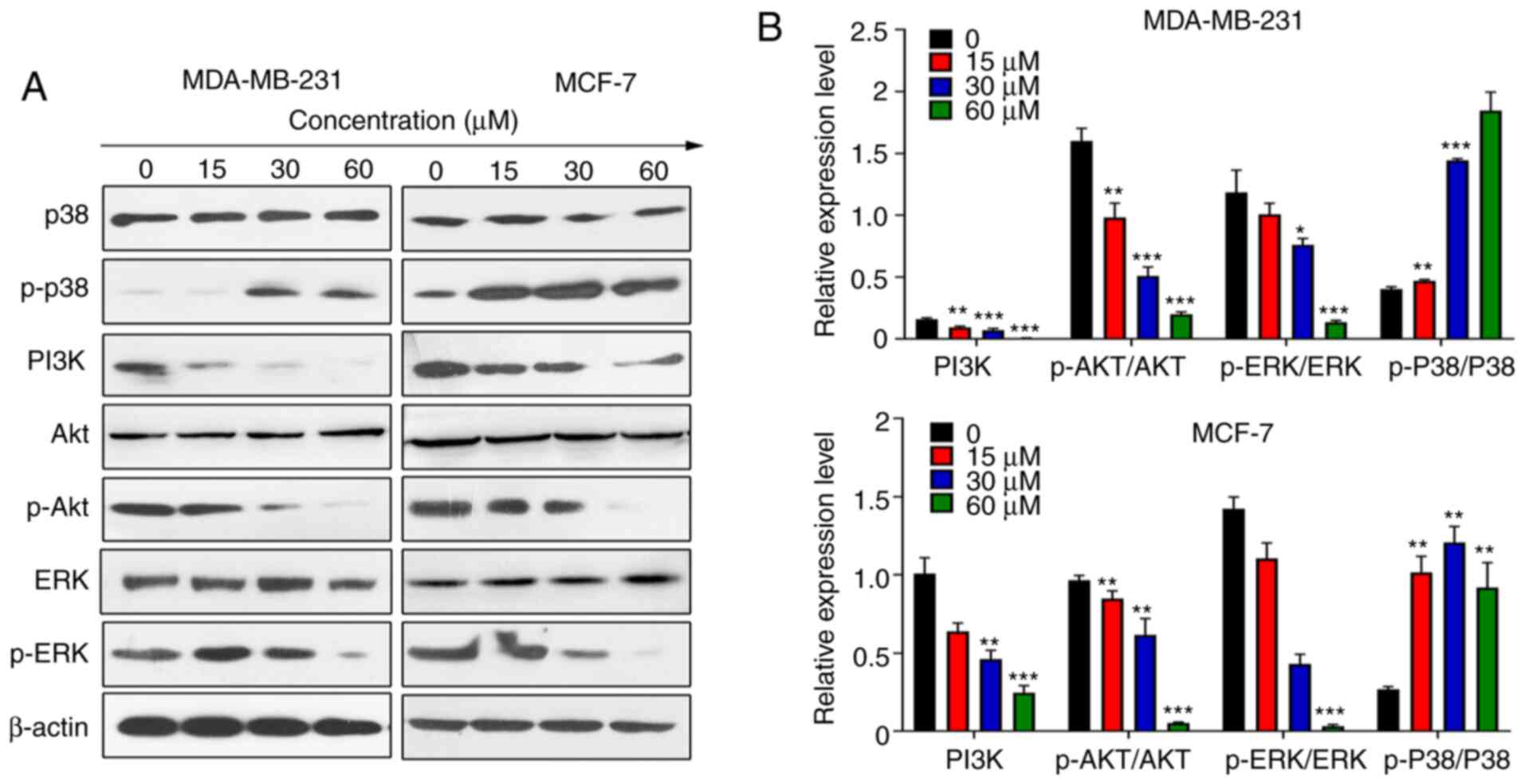
ER, NF-κB and ERK signals are significant in breast
cancer (30). First, the possibility
of ER signals being involved in Rosmanol-induced apoptosis was
investigated, and the expression of ERα and Erβ in MCF-7
ER-positive and MDA-MB231 ER-negative cells were analyzed via
western blotting. It was revealed that Rosmanol did not induce ER
expression in ER-negative MDA-MB231 cells, the data of which are
not shown.
ERK1/2 is an important pathway regulating the
signaling of multiple biological processes, such as cell
proliferation and cell growth (31).
The most classic signals in the ERK pathway are RAS and MAPK, and
ERK is also activated by the AKT signal (32). p38/MAPK signaling is ubiquitous in
normal and malignant cells and is activated in various cancer types
(33). Certain studies have shown
that the p38/MAPK pathway functions as a tumor suppressor by
regulating tumor cell proliferation and transformation (34–38). The
effect of Rosmanol on ERK signaling was investigated via western
blotting, and it was revealed that the expression of p-ERK
decreased significantly in MCF-7 and MDA-MB231 cells at varying
concentrations (Fig. 4A and B).
Concurrently, the p-p38 and PI3K/AKT signals were detected, in
particular the dramatically increased expression of p-p38 and
reduced expression of p-Akt and p-PI3K (Fig. 4A and B). Accordingly, it was
demonstrated that the upregulation of p-p38 and the downregulation
of PI3K/AKT regulate ERK signaling to participate in the apoptosis
caused by Rosmanol.
Rosmanol inhibits the JAK2/ STAT3
pathway in MCF-7 and MDA-MB 231cells
In tumors, STAT3 signaling is abnormally activated,
which to drives the proliferation, survival, invasiveness, and
metastasis of tumor cells, while strongly suppressing the apoptotic
response (39). Subsequently, the
effects of Rosmanol on the STAT3 pathway were investigated.
Transient transfection and luciferase tests were conducted in MCF-7
and MDA-MB 231 cells to test the effects of Rosmanol on STAT3
transcription. Notably, Rosmanol significantly inhibited
IL-6-induced STAT3 transcription in MCF-7 and MDA-MB 231 cells
(Fig. 5A).
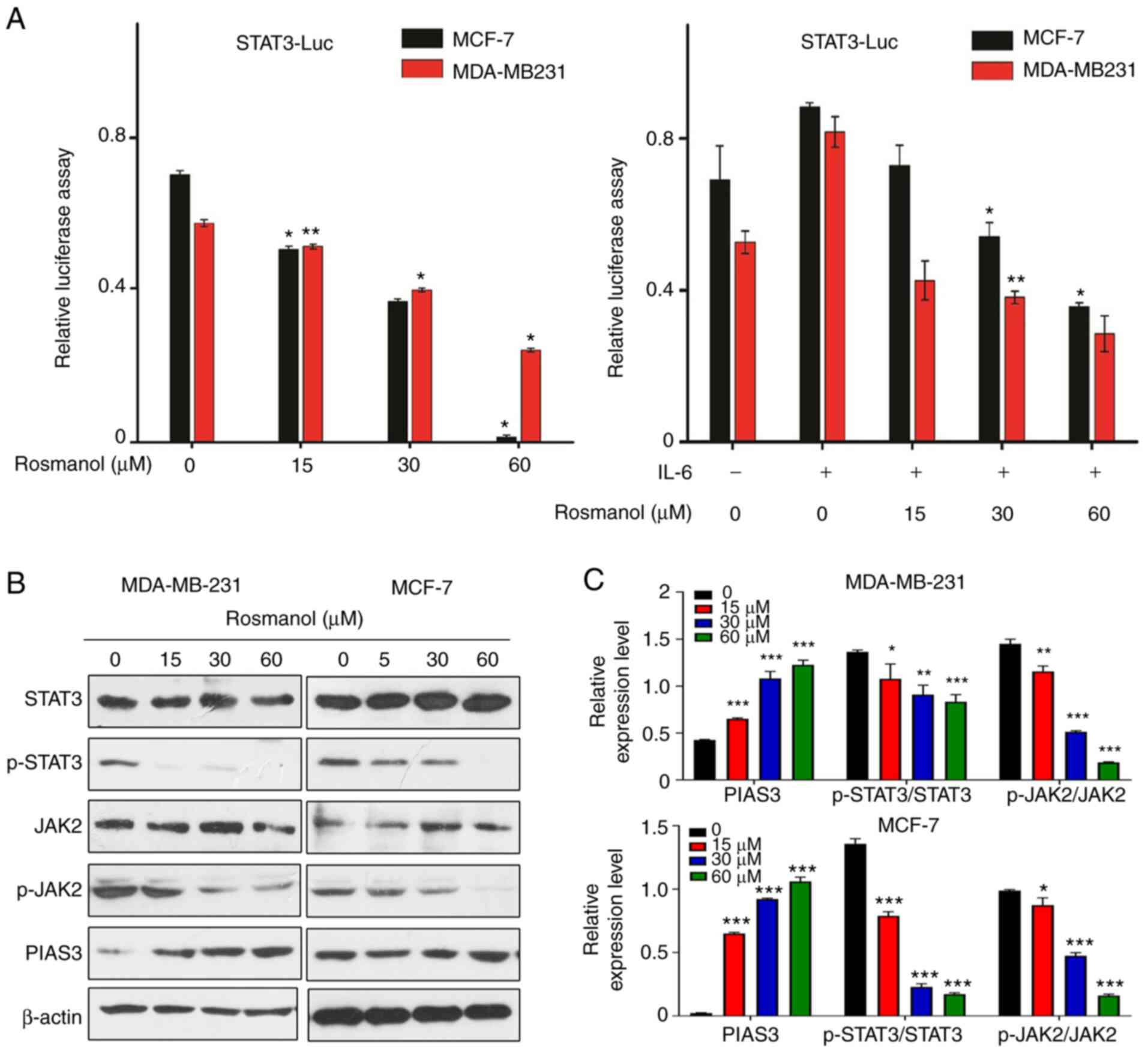 | Figure 5.Rosmanol induces MCF-7 and MDA-MB 231
cell apoptosis through STA3/JAK2 prohibition and PIAS3 induction.
(A) Transient transfection and luciferase tests of STAT3 were
conducted in MCF-7 and MDA-MB 231 cells. Each bar represents the
mean ± standard deviation of three experiments. *P<0.05,
**P<0.01 vs. control. (B) MCF-7 and MDA-MB 231 cells were
treated with different concentrations of Rosmanol, and then the
level of STAT-3, p-STAT-3, JAK2, p-JAK2, and PIAS3 were detected by
western blotting. (C) These protein bands were quantified and
statistically analyzed (β-actin as an internal control). The data
are presented as mean ± SD of three independent experiments.
*P<0.05, **P<0.01 and ***P<0.001 vs. untreated group. Luc,
luciferase; p, phosphorylated; PIAS3, protein inhibitor of
activated STAT3. |
Protein inhibitor of activated STAT3 (PIAS3) was
previously demonstrated to inhibit STAT3 transcriptional activity
and promote growth inhibition in vitro (40). The regulation of PIAS3 expression by
Rosmanol was then investigated in MCF-7 and MDA-MB 231 cells and
found that Rosmanol induced increased PIAS3 expression in a
dose-dependent manner (Fig. 5B and
C). Rosmanol also decreased the level of phosphorylated STAT3
(Fig. 5B and C). The effect of
Rosmanol on JAK2, the upstream signal of STAT3, was then
investigated and it was revealed that it was significantly reduced
(Fig. 5B and C). Therefore, it was
demonstrated that Rosmanol inhibited the phosphorylation and
transcription activity of STAT3 by augmenting PIAS3 or inhibiting
the phosphorylation of JAK2.
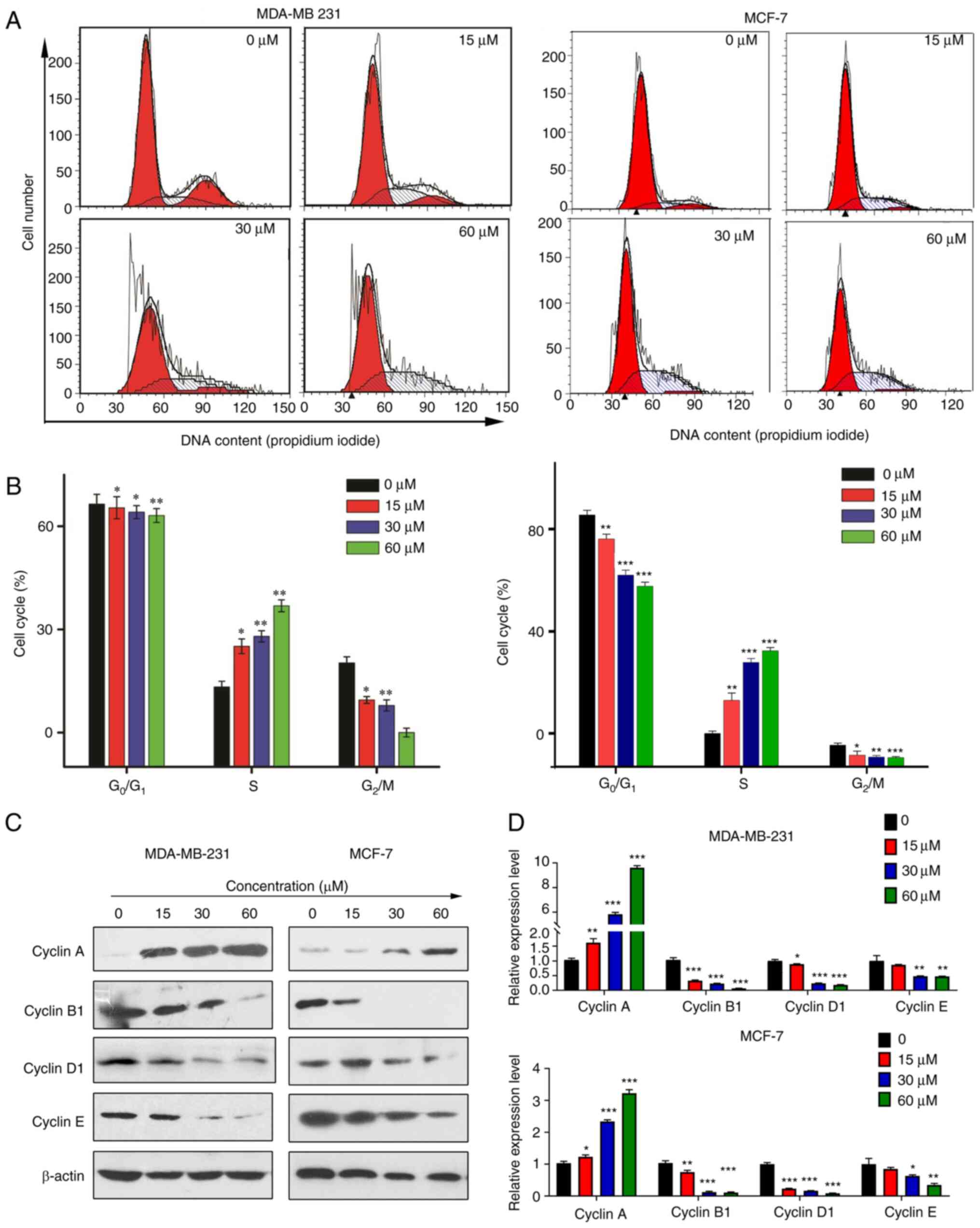
Rosmanol induces S phase cell cycle
arrest in MCF-7 and MDA-MB 231 cells
It was demonstrated that Rosmanol inhibits several
cell proliferation signals and at the same time promotes the
apoptosis of mitochondrial signals. In this situation, whether the
cytotoxicity of Rosmanol has an effect on the cell cycle was
investigated. Flow cytometry was conducted to check the cycle
arrest in MCf-7 and MDA-MB 231 cells stimulated by Rosmanol
(Fig. 6A and B), and the S phase
showed a dose-dependent increase in Rosmanol, while the
G0/G1 and G2/M percentages
decreased, respectively, for the same treatment concentration. This
proves that Rosmanol promotes S phase arrest.
Cyclins have been demonstrated as activators of
specific serine/threonine protein kinases, and are vital cell cycle
regulatory proteins that promote the cell cycle process (41,42).
Cyclin A is a marker in the S phase; it can activate two different
cyclin-dependent kinases (CDKs) and is expressed primarily in the S
phase (43). To further elucidate
the molecular mechanism underlying S phase arrest in MCf-7 and
MDA-MB 231 cells, western blotting was performed to describe the
cyclin proteins regulating the cell cycle. As shown in Fig. 6C and D, Rosmanol increased Cyclin A
expression, yet suppressed the expression of Cyclin E, Cyclin D1
and Cyclin B1 (Fig. 6C and D). These
results demonstrate that Rosmanol induces S phase arrest in MCf-7
and MDA-MB 231 cells.
Discussion
Rosmanol, an abundant natural product in the Salvia
species, was demonstrated in the present study to promote breast
cancer apoptosis in the mitochondrial pathway, simultaneously
accelerating apoptosis and inhibiting cell proliferation via ERK
and STAT3 signaling. It was also demonstrated that Rosmanol
triggers oxidative stress in MCF-7 and MDA-MB231 cells and exerts
anti-proliferation and mitochondrial apoptosis effects, depending
on ROS-induced DNA damage. Moreover, Rosmanol accelerated the
process of apoptosis through S-phase cell arrest. Moreover, it was
found that Rosmanol induced apoptosis progression associated with
the inhibition of STAT3 and ERK signaling, thereby preventing the
proliferation of breast cancer MDA-MB 231 cells.
Rosmanol was demonstrated to inhibit the
proliferation of MCF-7 and MDA-MB 231 cells and increased the
apoptosis rate of MCF-7 and MDA-MB 231 cells. Rosmanol induced
MCF-7 and MDA-MB 231 cell cytotoxicity with an IC50 (51,
26 and 19 µM) and (42, 28 and 16 µM), respectively, for 24, 48 and
72 h. However, Rosmanol had no significant effect on MCF-10A normal
breast cells. Notably, Rosmanol promoted the apoptotic process, by
activating cysteine protease or poly (ADP-ribose) polymerase
proteolysis in mitochondria, which further confirms that this
process may be attenuated by the cysteine protease inhibitor
Z-VAD-FMK. Rosmanol also stimulated apoptosis in human breast
cancer MCF-7 and MDA-MB 231 cells. Flow cytometry revealed that
Rosmanol increased ROS production which was attenuated by the
manifestation of NAC. Furthermore, the generation of ROS is
associated with improved cell apoptosis (44). These results affirmed the
cytotoxicity of Rosmanol via the induction of ROS, an important
apoptosis inducer, against human breast cancer MCF-7 and MDA-MB 231
cells.
Cell proliferation, autoregulated by the cell cycle,
is regulated via a myriad of complex cyclin proteins (45), while cyclin B is required for cells
to enter mitosis and G2/M transition (46). In addition, the S phase relies on a
pathway to regulate Cyclin A synthesis (47). Here, it was demonstrated that the
cytotoxicity of Rosmanol significantly inhibited the proliferation
of cancer cells and that the channel of cells in the
G0/G1 to S phase was synchronized by cyclins.
It was demonstrated that Rosmanol can stimulate MDA-MB 231 cell
cycle arrest at the S phase by upregulating the expression of
cyclin A and consequently downregulating the protein expression of
Cyclin B1, Cyclin D1 and Cyclin E.
In tumor cells, ERK is often activated by Akt, and
both of these Akt feedback signaling molecules serve a vital role
in the proliferation and evasion of apoptosis in cells (48). A series of clinical trials are
underway investigating the combined application of inhibitors that
target the PI3K and MEK/ERK pathways, since single reagents have
limited efficacy (49). The present
study also demonstrated that Rosmanol functions as a potential
natural antineoplastic alternative to the combination of Akt and
ERK inhibitors, significantly inhibiting the Akt and ERK pathways,
and considerably enhancing the antiproliferative effects of MCF-7
and MDA-MB 231 cells. NF-κB and Akt/ERK are two major cell-survival
pathways that are often constitutively activated and communicate
within cancer cells (50). p-p38
exerts a tumor suppressor effect. Here it was demonstrated that
Rosmanol increases the expression of p-p38.
STAT proteins are critically involved in
tumorigenesis in various cancers (51). PIAS3, the main cellular inhibitor of
STAT3, has been described as a modulator of DNA-binding
transcription factors (52).
Activated JAKs phosphorylate STAT proteins, leading to their
dimerization and translocation into the nucleus. In the nucleus,
STATs act as transcription factors with pleiotropic downstream
effects (53). In cancer cells,
STAT3 activation leads to increased expression of downstream target
genes, leading to increased cell proliferation (54). The present results showed that
Rosmanol significantly inhibited IL-6-induced STAT3 transcription
in MCF-7 and MDA-MB 231 cells, that the Rosmanol-induced increase
in PIAS3 expression was dose-dependent and that Rosmanol inhibits
JAK2 and STAT3 phosphorylation. It was demonstrated that Rosmanol
augments PIAS3 expression and inhibits the JAK2/STA3 pathway.
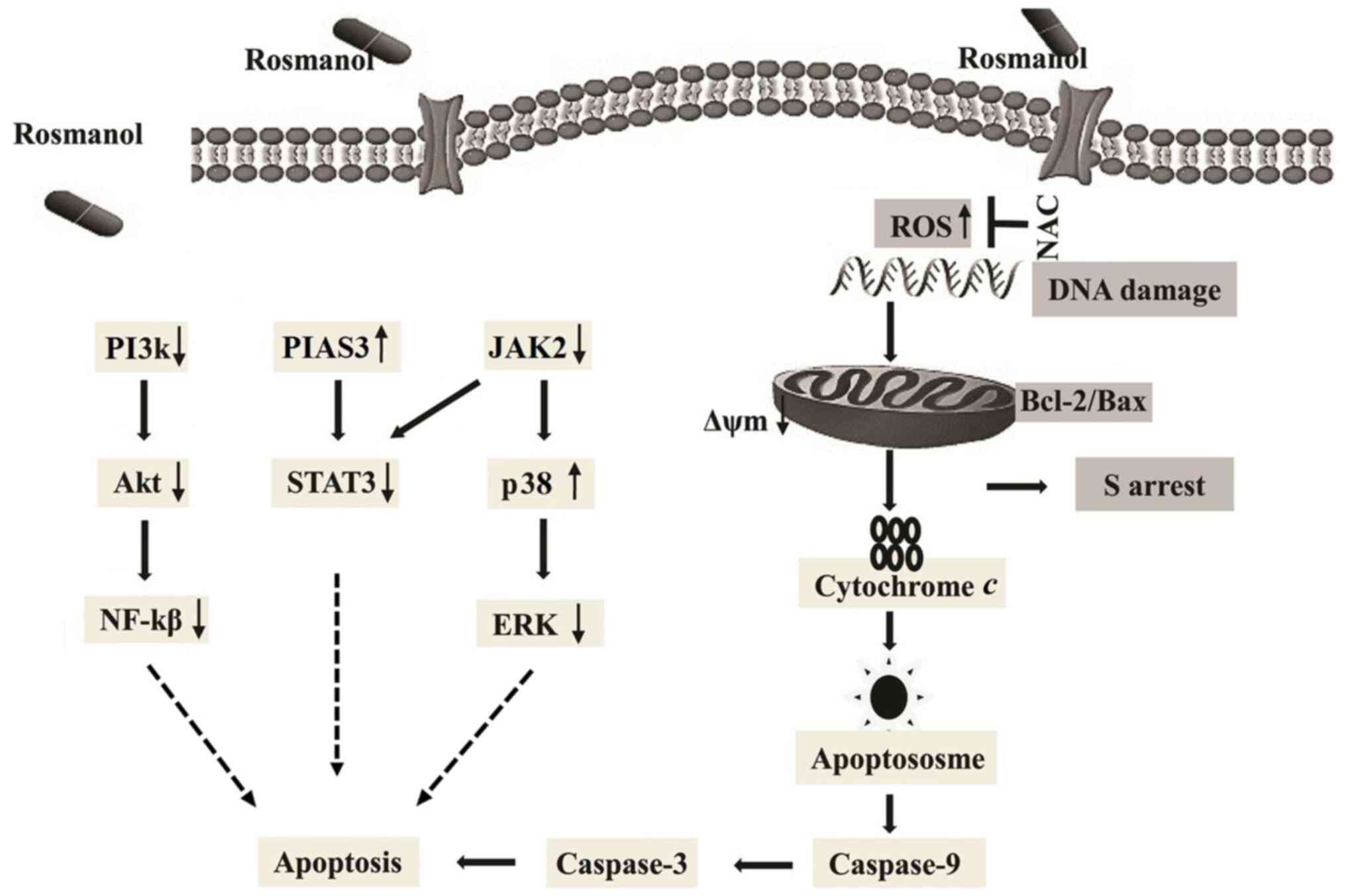
The overall findings of the present study are
summarized in Fig. 7, demonstrating
that Rosmanol triggers oxidative stress causes ROS-induced DNA
damage, and serves an anti-proliferative role with a mitochondrial
apoptotic effect in MCF-7 and MDA-MB 231 cells. Rosmanol causes
cancer cell arrest at the S phase in the cell cycle. Furthermore,
it was demonstrated that Rosmanol has effective antiproliferative
and anticancer effects via the inhibition of JAK2/STAT3 and
PI3K/AKT signals.
Acknowledgements
Not applicable.
Funding
This study was supported by the Ministry of Science
and Technology (grant no. 2021YFE0108000 and 2016YFE0128500), the
National Natural Science Foundation of China (grant no. 31870758),
the Jilin Provincial Science & Technology Department (grant no.
20200201025JC).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
DJ performed the cell and protein studies, analyzed
the data, and drafted the manuscript. JX and SL participated in the
cell and protein studies. MIN, XZ and XTL participated in analyzing
the data, WW and TM performed cell cycle studies in MCF7 cells and
cell apoptosis experiments by immunofluorescence in MDA-MB-231
cells after the article was submitted. JL and XML conceived the
idea, designed the experiment, and drafted the manuscript. DJ and
MIN confirmed the authenticity of all the raw data. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Draper L: Breast cancer: Trends, risks,
treatments, and effects. AAOHN J. 54:445–453. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bleicher RJ: Timing and delays in breast
cancer evaluation and treatment. Ann Surg Oncol. 25:2829–2838.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Castaneda SA and Strasser J: Updates in
the treatment of breast cancer with radiotherapy. Surg Oncol Clin N
Am. 26:371–382. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Quintela-Fandino M, Soberon N, Lluch A,
Manso L, Calvo I, Cortes J, Moreno-Antón F, Gil-Gil M,
Martinez-Jánez N, Gonzalez-Martin A, et al: Critically short
telomeres and toxicity of chemotherapy in early breast cancer.
Oncotarget. 8:21472–21482. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kuete V, Mbaveng AT, Nono EC, Simo CC,
Zeino M, Nkengfack AE and Efferth T: Cytotoxicity of seven
naturally occurring phenolic compounds towards multi-factorial
drug-resistant cancer cells. Phytomedicine. 23:856–863. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lucas L, Russell A and Keast R: Molecular
mechanisms of inflammation. Anti-inflammatory benefits of virgin
olive oil and the phenolic compound oleocanthal. Curr Pharm Des.
17:754–768. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Petiwala SM and Johnson JJ: Diterpenes
from rosemary (Rosmarinus officinalis): Defining their
potential for anti-cancer activity. Cancer Lett. 367:93–102. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lai CS, Lee JH, Ho CT, Liu CB, Wang JM,
Wang YJ and Pan MH: Rosmanol potently inhibits
lipopolysaccharide-induced iNOS and COX-2 expression through
downregulating MAPK, NF-kappaB, STAT3 and C/EBP signaling pathways.
J Agric Food Chem. 57:10990–10998. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cheng AC, Lee MF, Tsai ML, Lai CS, Lee JH,
Ho CT and Pan MH: Rosmanol potently induces apoptosis through both
the mitochondrial apoptotic pathway and death receptor pathway in
human colon adenocarcinoma COLO 205 cells. Food Chem Toxicol.
49:485–493. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tabata K, Kim M, Makino M, Satoh M, Satoh
Y and Suzuki T: Phenolic diterpenes derived from Hyptis incana
induce apoptosis and G(2)/M arrest of neuroblastoma cells.
Anticancer Res. 32:4781–4789. 2012.PubMed/NCBI
|
|
11
|
Cheng M, Michalski S and Kommagani R: Role
for growth regulation by estrogen in breast cancer 1 (GREB1) in
hormone-dependent cancers. Int J Mol Sci. 19:25432018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mishra AK, Abrahamsson A and Dabrosin C:
Fulvestrant inhibits growth of triple negative breast cancer and
synergizes with tamoxifen in ERα positive breast cancer by
up-regulation of ERβ. Oncotarget. 7:56876–56888. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sharma VR, Gupta GK and Sharma AK, Batra
N, Sharma DK, Joshi A and Sharma AK: PI3K/Akt/mTOR intracellular
pathway and breast cancer: Factors, mechanism and regulation. Curr
Pharm Des. 23:1633–1638. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu M, Qi B, Xiaoxiang W, Xu J and Liu X:
Baicalein increases cisplatin sensitivity of A549 lung
adenocarcinoma cells via PI3K/Akt/NF-κB pathway. Biomed
Pharmacother. 90:677–685. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Voskas D, Ling LS and Woodgett JR: Signals
controlling un-differentiated states in embryonic stem and cancer
cells: Role of the phosphatidylinositol 3′kinase pathway. J Cell
Physiol. 229:1312–1322. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huth HW, Albarnaz JD, Torres AA, Bonjardim
CA and Ropert C: MEK2 controls the activation of MKK3/MKK6-p38 axis
involved in the MDA-MB-231 breast cancer cell survival: Correlation
with cyclin D1 expression. Cell Signal. 28:1283–1291. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Heckler MM, Thakor H, Schafer CC and
Riggins RB: ERK/MAPK regulates ERRγ expression, transcriptional
activity and receptor-mediated tamoxifen resistance in ER+ breast
cancer. FEBS J. 281:2431–2442. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Harrison DA, Binari R, Nahreini TS, Gilman
M and Perrimon N: Activation of a Drosophila Janus kinase (JAK)
causes hematopoietic neoplasia and developmental defects. EMBO J.
14:2857–2865. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang Z, Wang F, Du C, Guo H, Ma L, Liu X,
Kornmann M, Tian X and Yang Y: BRM/SMARCA2 promotes the
proliferation and chemoresistance of pancreatic cancer cells by
targeting JAK2/STAT3 signaling. Cancer Lett. 402:213–224. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ho JM, Nguyen MH, Dierov JK, Badger KM,
Beattie BK, Tartaro P, Haq R, Zanke BW, Carroll MP and Barber DL:
TEL-JAK2 constitutively activates the extracellular
signal-regulated kinase (ERK), stress-activated protein/Jun kinase
(SAPK/JNK), and p38 signaling pathways. Blood. 100:1438–1448.
2002.PubMed/NCBI
|
|
21
|
Nasser MI, Masood M, Wei W and Li X, Zhou
Y, Liu B, Li J and Li X: Cordycepin induces apoptosis in SGC7901
cells through mitochondrial extrinsic phosphorylation of PI3K/Akt
by generating ROS. Int J Oncol. 50:911–919. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jiang C, Masood M, Rasul A, Wei W, Wang Y,
Ali M, Mustaqeem M, Li J and Li X: Altholactone inhibits NF-κB and
STAT3 activation and induces reactive oxygen species-mediated
apoptosis in prostate cancer DU145 Cells. Molecules. 22:2402017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fan TJ, Han LH, Cong RS and Liang J:
Caspase family proteases and apoptosis. Acta Biochim Biophys Sin
(Shanghai). 37:719–727. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shi Y: Mechanisms of caspase activation
and inhibition during apoptosis. Mol Cell. 9:459–470. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Marinello PC, da Silva TN, Panis C, Neves
AF, Machado KL, Borges FH, Guarnier FA, Bernardes SS,
de-Freitas-Junior JC, Morgado-Díaz JA, et al: Mechanism of
metformin action in MCF-7 and MDA-MB-231 human breast cancer cells
involves oxidative stress generation, DNA damage, and transforming
growth factor β1 induction. Tumour Biol. 37:5337–5346. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Vrhovac Madunić I, Madunić J, Antunović M,
Paradžik M, Garaj-Vrhovac V, Breljak D, Marijanović I and Gajski G:
Apigenin, a dietary flavonoid, induces apoptosis, DNA damage, and
oxidative stress in human breast cancer MCF-7 and MDA MB-231 cells.
Naunyn Schmiedebergs Arch Pharmacol. 391:537–550. 2018. View Article : Google Scholar
|
|
27
|
Kello M, Takac P, Kubatka P, Kuruc T,
Petrova K and Mojzis J: Oxidative stress-induced DNA damage and
apoptosis in clove buds-treated MCF-7 cells. Biomolecules.
10:1392020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bakhtiari E, Hosseini A and Mousavi SH:
The role of ROS and NF-κB pathway in olmesartan induced-toxicity in
HeLa and mcf-7 cell lines. Biomed Pharmacother. 93:429–434. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fumarola C, Caffarra C, La Monica S,
Galetti M, Alfieri RR, Cavazzoni A, Galvani E, Generali D,
Petronini PG and Bonelli MA: Effects of sorafenib on energy
metabolism in breast cancer cells: Role of AMPK-mTORC1 signaling.
Breast Cancer Res Treat. 141:67–78. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Haldosén LA, Zhao C and Dahlman-Wright K:
Estrogen receptor beta in breast cancer. Mol Cell Endocrinol.
382:665–672. 2014. View Article : Google Scholar
|
|
31
|
Sun Y, Liu WZ, Liu T, Feng X, Yang N and
Zhou HF: Signaling pathway of MAPK/ERK in cell proliferation,
differentiation, migration, senescence and apoptosis. J Recept
Signal Transduct Res. 35:600–604. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cakir M and Grossman AB: Targeting MAPK
(Ras/ERK) and PI3K/Akt pathways in pituitary tumorigenesis. Expert
Opin Ther Targets. 13:1121–1134. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lee S, Rauch J and Kolch W: Targeting MAPK
signaling in cancer: Mechanisms of drug resistance and sensitivity.
Int J Mol Sci. 21:11022020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Loesch M and Chen G: The p38 MAPK stress
pathway as a tumor suppressor or more? Front Biosci. 13:3581–3593.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang J, Liu Z, Feng X, Gao S, Xu S and Liu
P: Tumor suppressor gene ING3 induces cardiomyocyte hypertrophy via
inhibition of AMPK and activation of p38 MAPK signaling. Arch
Biochem Biophys. 562:22–30. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Martínez-Limón A, Joaquin M, Caballero M,
Posas F and de Nadal E: The p38 pathway: From biology to cancer
therapy. Int J Mol Sci. 21:19132020. View Article : Google Scholar
|
|
37
|
Yu Z, Du Y, Li H, Huang J, Jiang D, Fan J,
Shen Y, Zhang L, Yu X, Xu N and Ke Q: miR-642 serves as a tumor
suppressor in hepatocellular carcinoma by regulating SEMA4C and p38
MAPK signaling pathway. Oncol Lett. 20:742020.PubMed/NCBI
|
|
38
|
Hong B, Li H, Zhang M, Xu J, Lu Y, Zheng
Y, Qian J, Chang JT, Yang J and Yi Q: p38 MAPK inhibits breast
cancer metastasis through regulation of stromal expansion. Int J
Cancer. 136:34–43. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dabir S, Kluge A, Kresak A, Yang M, Fu P,
Groner B, Wildey G and Dowlati A: Low PIAS3 expression in malignant
mesothelioma is associated with increased STAT3 activation and poor
patient survival. Clin Cancer Res. 20:5124–5132. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Aghazadeh S and Yazdanparast R:
Mycophenolic acid potentiates HER2-overexpressing SKBR3 breast
cancer cell line to induce apoptosis: Involvement of AKT/FOXO1 and
JAK2/STAT3 pathways. Apoptosis. 21:1302–1314. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Liang W, Guan H, He X, Ke W, Xu L, Liu L,
Xiao H and Li Y: Down-regulation of SOSTDC1 promotes thyroid cancer
cell proliferation via regulating cyclin A2 and cyclin E2.
Oncotarget. 6:31780–31791. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Geng Y, Michowski W, Chick JM, Wang YE,
Jecrois ME, Sweeney KE, Liu L, Han RC, Ke N, Zagozdzon A, et al:
Kinase-independent function of E-type cyclins in liver cancer. Proc
Natl Acad Sci USA. 115:1015–1020. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yam CH, Fung TK and Poon RY: Cyclin A in
cell cycle control and cancer. Cell Mol Life Sci. 59:1317–1326.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Mehdad A, Brumana G, Souza AA, Barbosa J,
Ventura MM and de Freitas SM: A Bowman-Birk inhibitor induces
apoptosis in human breast adenocarcinoma through mitochondrial
impairment and oxidative damage following proteasome 20S
inhibition. Cell Death Discov. 2:150672016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lim S and Kaldis P: Cdks, cyclins and
CKIs: Roles beyond cell cycle regulation. Development.
140:3079–3093. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Timofeev O, Cizmecioglu O, Settele F,
Kempf T and Hoffmann I: Cdc25 phosphatases are required for timely
assembly of CDK1-cyclin B at the G2/M transition. J Biol Chem.
285:16978–16990. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Moulager M, Corellou F, Vergé V, Escande
ML and Bouget FY: Integration of light signals by the
retinoblastoma pathway in the control of S phase entry in the
picophytoplanktonic cell Ostreococcus. PLoS Genet. 56:e10009572010.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wang M, Shu ZJ, Wang Y and Peng W:
Stachydrine hydrochloride inhibits proliferation and induces
apoptosis of breast cancer cells via inhibition of Akt and ERK
pathways. Am J Transl Res. 9:1834–1844. 2017.PubMed/NCBI
|
|
49
|
Asati V, Mahapatra DK and Bharti SK:
PI3K/Akt/mTOR and Ras/Raf/MEK/ERK signaling pathways inhibitors as
anticancer agents: Structural and pharmacological perspectives. Eur
J Med Chem. 109:314–341. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Sun H, Zheng X, Wang Q, Yan J, Li D, Zhou
Y, Lin Y, Zhang L and Wang X: Concurrent blockade of NF-κB and Akt
pathways potentiates cisplatin's antitumor activity in vivo.
Anticancer Drugs. 23:1039–1046. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Pastuszak-Lewandoska D,
Domańska-Senderowska D, Kordiak J, Antczak A, Czarnecka KH,
Migdalska-Sęk M, Nawrot E, Kiszałkiewicz JM and Brzeziańska-Lasota
E: Immunoexpression analysis of selected JAK/STAT pathway molecules
in patients with non-small-cell lung cancer. Pol Arch Intern Med.
127:758–764. 2017.PubMed/NCBI
|
|
52
|
Yagil Z, Nechushtan H, Kay G, Yang CM,
Kemeny DM and Razin E: The enigma of the role of protein inhibitor
of activated STAT3 (PIAS3) in the immune response. Trends Immunol.
31:199–204. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Yang X, Tang Z, Zhang P and Zhang L:
Research advances of JAK/STAT signaling pathway in lung cancer.
Zhongguo Fei Ai Za Zhi. 22:45–51. 2019.(In Chinese). PubMed/NCBI
|
|
54
|
Lu Y, Zhou J, Xu C, Lin H, Xiao J, Wang Z
and Yang B: JAK/STAT and PI3K/AKT pathways form a mutual
transactivation loop and afford resistance to oxidative
stress-induced apoptosis in cardiomyocytes. Cell Physiol Biochem.
21:305–314. 2008. View Article : Google Scholar : PubMed/NCBI
|