Introduction
Liver cancer has the third highest rate of
cancer-associated death worldwide (1). Hepatitis B virus infection is
associated with most cases of liver cancer in China, accounting for
~85% of cases (2). Currently, the
treatment of liver cancer mainly involves surgery, radiotherapy and
chemotherapy. Although sorafenib has been established as an
effective drug for advanced liver cancer, renal cell carcinoma and
thyroid cancer, the number of patients with liver cancer exhibiting
a complete response to sorafenib is small (3). In view of the emerging crisis of
sorafenib resistance in hepatocellular carcinoma, further research
on drug resistance is urgently required.
Autophagy is a cellular metabolic process in which
cells degrade their own components through lysosomes or vacuoles to
maintain normal physiological activities and homeostasis (4). In the process of tumor development,
autophagy can allow the growth requirements of tumor cells to be
met by degrading organelles and proteins (4). Altering the autophagy level in tumor
cells is a new proposed approach for tumor therapy (5). Recently, a type of selective autophagy
called lipophagy, through which lipids are selectively recognized
and degraded, has been identified (6). Lipophagy serves an important role in
regulating lipid metabolism and maintaining intracellular lipid
homeostasis (6). Lipophagy is
directly or indirectly regulated by genes, enzymes, transcription
regulators and other factors (7,8). Several
studies support the idea that de novo lipogenesis in cancer cells
is associated with chemoresistance at multiple levels (6,7,9). The ability of lipophagy to respond to
changes in nutrient supply allows the cell to alter lipid droplet
(LD) metabolism to meet the cell's energy demands (10). Activation of lipophagy combined with
standard chemotherapy can effectively decrease the LDs leading to
improved chemotherapy efficiency (11). Consequently, targeting lipophagy to
mediate LDs degradation may be a novel strategy to treat tumors and
overcome drug resistance.
Non-coding RNAs (ncRNAs) have recently become
recognized as important regulators in signaling pathways associated
with drug resistance in liver cancer (12). Therefore, pharmacological targeting
of these ncRNAs may constitute a novel strategy for reversing drug
resistance (13). As an important
protein involved in the nutrient-sensing pathway, along with the
mammalian target of rapamycin protein and AMP-activated kinase in
mammals, silent information regulator 2 homolog 1 (SIRT1) directly
induces autophagy by deacetylating autophagy-related gene (ATG)5,
ATG7 and LC3 (14). SIRT1 can also
deacetylate FOXO to regulate the expression of autophagy regulatory
molecules and modulate lipophagy (15,16).
However, the role of ncRNAs in regulating lipophagy in the context
of drug resistance has not been reported. MicroRNA (miR/miRNA)-425
is an oncogene that serves an important role in tumor progression
and development (13); however, it
has no clear mechanism in HCC drug resistance. The present study
aimed to explore the mechanism by which miR-425 regulates lipophagy
and the effect of miR-425 on sorafenib resistance.
Materials and methods
Chemicals and materials
miR-425 mimics and miR-425 inhibitor were purchased
from Shanghai GenePharma Co., Ltd. Anti-LC3 (cat. no. 13394; Cell
Signaling Technology, Inc.; 1:1,000) and anti-β-actin antibodies
were purchased from Abcam (cat. no. ab8226; 1;1,000). Bafilomycin A
(BafA1; 25 nM; Beyotime Institute of Biotechnology) was used for
treatment for 6 h at 37°C.
Cell culture
HepG2 (CoBioer Biosciences Co., Ltd.; cat. no.
CBP60199), Hep3B (CoBioer Biosciences Co., Ltd.; cat. no.
CBP60197), PLC (CoBioer Biosciences Co., Ltd.; cat. no. CBP60223),
Huh7 (CoBioer Biosciences Co., Ltd.; cat. no. CBP60202) and MIHA
(Hunan Fenghui Biotechnology Co., Ltd.; cat. no. CL0469) cells were
cultured in DMEM (cat. no. 12430104; Invitrogen; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS (cat. no. 10091148;
HyClone; Cytiva) and 1% penicillin-streptomycin solution (cat. no.
15140163; Gibco; Thermo Fisher Scientific, Inc.; ratio 9:1:1) at
37°C and 5% CO2. Cross-contamination of the cell lines
was excluded by short tandem repeat profiling (17,18).
BODIPY staining
Cells were fixed with 4% paraformaldehyde for 10 min
at room temperature and stained with 1 µg/ml BODIPY (cat. no.
493/503; Sigma-Aldrich; Merck KGaA) in PBS for 10 min at room
temperature (11). After staining,
coverslips were washed with PBS and mounted in a slide with Prolong
Gold Antifade Reagent (Invitrogen; Thermo Fisher Scientific, Inc.).
BODIPY stained cells were examined under an inverted confocal
fluorescence microscope (Zeiss AG; magnification ×400).
Cell transfection
miR-425 mimics (5′-GUCAAAAAUGUCGAUAUGUCAUU-3′; 25
µM) and miR-425 inhibitor (5′-GGGGAGTTAGGATTAGGTC-3′; 50 µM) were
purchased from Shanghai GenePharma Co., Ltd. Liver cancer cells
were seeded in 6-well plates at 5×105 cells/well. When
the cells were 70% confluent, Lipofectamine® 3000 (cat.
no. L3000015; Invitrogen; Thermo Fisher Scientific, Inc.) was used
for transfection with the miR-425 mimics, miR-425 inhibitor or the
corresponding negative control (miR-NC 5′-CTCGCTTCGGCAGCACA-3′ and
si-NC: 5′-AACGCTTCACGAATTTGCGT-3′) at 37°C and 5% CO2.
The culture medium was changed after 6 h, and cells were harvested
for PCR analysis after 48 h, while western blotting was performed
after 72 h.
Immunofluorescence
Liver cancer cells were stained with anti-LC3
antibody (abcam; cat. no. ab225383) for 48 h and were then treated
with miR-425 inhibitors and stained with 1 µg/ml BODIPY (cat. no.
493/503; Sigma-Aldrich; Merck KGaA) in PBS for 10 min at room
temperature. After induction of autophagy by miR-425 inhibitors,
the number of mCherry-positive cells was increased compared with
that of untreated cells. Red fluorescent protein expression was
quantitatively analyzed by ImageJ (v1.52) software. Cells were
grown overnight on cover glasses, fixed with 4% paraformaldehyde
(37°C and 5% CO2) and sealed with PBS containing 1%
bovine serum albumin (BSA; cat. no. ST023; Beyotime Institute of
Biotechnology) for 1.5 h at room temperature. Stained samples were
visualized using a Zeiss-LSM 510 fluorescence microscope (Zeiss AG;
magnification, ×400).
Reverse transcription-quantitative
(RT-q)PCR
All detection of miRNA expression was performed by
RT-qPCR. TRIzol® reagent (cat. no. 15596026; Invitrogen;
Thermo Fisher Scientific, Inc.) was used to lyse cells for
extraction of total RNA. Total RNA (2 µg) was reverse transcribed
using a microRNA kit according to the manufacturer's protocol (cat.
no. 4366596; Invitrogen; Thermo Fisher Scientific, Inc.). A 20-µl
final reaction mixture was used for qPCR amplification using SYBR
Green PCR Master Mix (cat. no. 4309155; Thermo Fisher Scientific,
Inc.). Pre-denaturation at 95°C for 3 min, denaturation at 95°C for
30 sec, annealing at 55°C for 30 sec, extension at 72°C for 60 sec,
for a total of 30 cycles, and after completion of the cycle,
extension at 72°C for 5 min. The target mRNA expression levels were
normalized to the levels of β-actin, U6 was used as an reaction
internal reference. All samples were repeated for 3 wells. RNA
expression levels were calculated using the 2−ΔΔCq
method (19). The primer sequences
used were as follows: MiR-425 forward, 5′-TGCGCTCAGCAAACATTTATTG-3′
and reverse, 5′-CCAGTGCAGGGTCCGAGGTATT-3′; U6 forward,
5′-CTCGCTTCGGCAGCACA-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′;
SIRT1 forward, 5′-GTGCAGGTAGTTCCTCGGTG-3′ and reverse,
5′-CACAACTCACAGCATGCACAA-3′; and β-actin forward,
5′-AGCGAGCATCCCCCAAAGTT-3′ and reverse, 5′-GGGCACGAAGGCTCATCATT-3′;
ATGL forward, 5′-GTGTCAGACGGCGAGAATG-3′, reverse,
5′-TGGAGGGAGGGAGGGATG-3′
Western blot analysis
Cells were lysed in RIPA buffer (cat. no. P0013B;
Beyotime Institute of Biotechnology) at 4°C for 2 h. Supernatants
were collected and the protein concentrations were quantified by
bicinchoninic acid (BCA) via using a BCA protein assay kit
(Beyotime Institute of Biotechnology). Proteins (30 µg/lane) were
separated via 10% SDS-PAGE and were then transferred onto PVDF
membranes (cat. no. 88585; Thermo Fisher Scientific, Inc.).
Subsequently, membranes were blocked in 5% BSA for 2 h at 37°C and
incubated with anti-LC3 (1:1,000), anti-ATGL (1:1,000) (Abcam; cat.
no. ab207799) or anti-β-actin (1:1,000; cat. no. ab8226; Abcam)
primary antibodies overnight at 4°C. After primary antibody
incubation, membranes were incubated with HRP-conjugated secondary
antibodies (cat. no. sc-2357; 1:5,000; Santa Cruz Biotechnology,
Inc). The proteins were visualized using an enhanced
chemiluminescence detection kit (Amersham; Cytiva). The gray values
of the protein bands were measured using ImageJ (v1.52) software
(National Institutes of Health).
Luciferase reporter assay
The SIRT1 3′UTR containing the specific binding site
for miR-425 was amplified by PCR. The 3′UTR sequence containing a
mutation in the specific binding site was also amplified, and then
the sequences were inserted into the pre-miR-RB-REPORT vector and
transfection into cells (Thermo Fisher Scientific, Inc.).
Lipofectamine 3000 was used to transfection
(Lipofectamine® 3000; cat. no. L3000015; Invitrogen;
Thermo Fisher Scientific, Inc.). After 24 h of transfection with
mimic-NC and miR-425 mimics, a luciferase assay system (cat. no.
16170; Thermo Fisher Scientific, Inc.) was used to perform the
assay, and Renilla luciferase activity was used for
normalization.
Cell Counting Kit-8 (CCK-8) assay
Cells (2,000 cells/well) were seeded in 96-well
plates (Corning, Inc.) in DMEM with 10% FBS (cat. no. 10091148;
HyClone; Cytiva). Cells were then treated with sorafenib (10 µM,
37°C and 5% CO2). After culturing the cells for 48 h,
CCK-8 solution (cat. no. C0037; Beyotime Institute of
Biotechnology) was added to the cells for 30 min at room
temperature. The absorbance was measured with an ELISA plate reader
(Varioskan Flash; Thermo Fisher Scientific, Inc.) at 450 nm.
Bioinformatic analysis
The Cancer Genome Atlas (TCGA) liver cancer datasets
were obtained from https://portal.gdc.cancer.gov/. Gene Expression
Omnibus (GEO) datasets (dataset nos. GSE93595 and GSE94550)
(20,21) were obtained from https://www.ncbi.nlm.nih.gov/geo/. The gene
expression matrix for the TCGA datasets was analyzed using R
software v4.0.2 (limma package v3.44.3) (22,23)
using log fold-change (FC) >0.6 or logFC <-0.6 and P<0.05
as the cut-off values, and the correlation between miR-425 and
SIRT1 expression was evaluated using Pearson correlation analysis
with SPSS (22.0; IBM, Corp.). GEO datasets were analyzed with GEO2R
(v3.2.3) (Table SI) (24,25), and
Venn diagrams were generated with VennDiagram (v1.2.20) (24,25). The
site for miR-425 binding with SIRT1 was predicted using StarBase
2.0 (http://starbase.sysu.edu.cn/).
Invasion and migration assays
Cells (2×104 cells/chamber) were seeded
on the upper of the transwell chambers (Thermo Fisher Scientific,
Inc.), DMEM (cat. no. 12430104; Invitrogen; Thermo Fisher
Scientific, Inc.) and FBS (10%) were plated in the lower chamber,
and invasion assays were added matrix (4°C, duration of precoating:
30 min), Matrigel was used to distinguish between migration and
invasion. After 48 h, cells were fixed with 4% paraformaldehyde
(37°C and 5% CO2) for 20 min and then washed using PBS
three times. Subsequently, crystal violet (0.05% for 30 min at room
temperature) was used to stain cells. A light microscope was used
to observe migratory cells in the lower chamber (Olympus
Corporation).
Statistical analysis
All data were analyzed using SPSS 22.0 (IBM, Corp.)
and are presented as the mean ± SD (experiments were repeated in
triplicate). One-way ANOVA and Bonferroni's post hoc test was used
to compare differences between multiple groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-425 promotes sorafenib resistance
in liver cancer
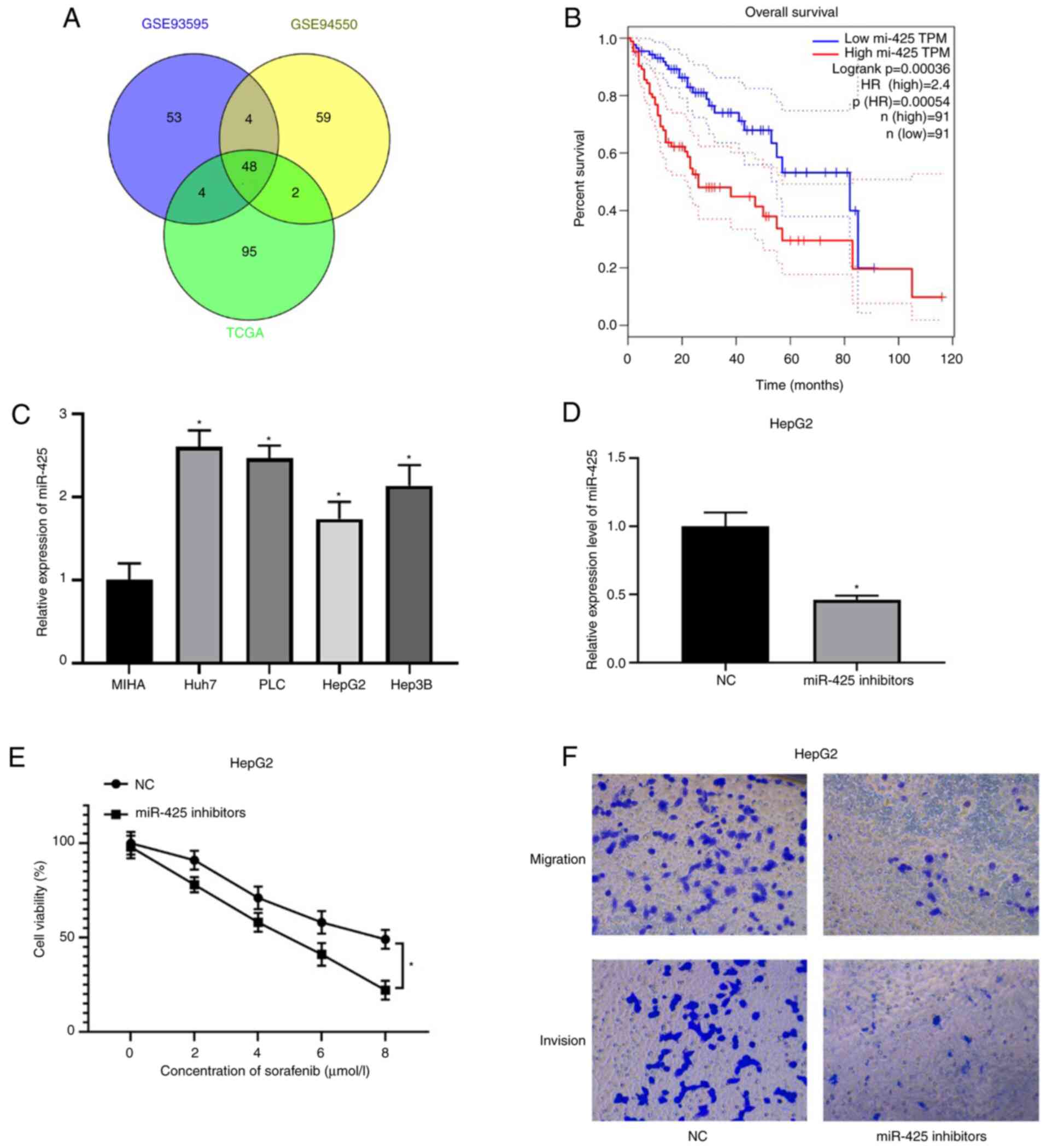
To explore the role of miRNAs in drug resistance in
liver cancer, TCGA database was screened for highly expressed
miRNAs, and their enrichment was analyzed in sorafenib-resistant
samples from GEO (GSE93595 and GSE94550), there were 48 different
overlapping miRNAs (Fig. 1A and
Table SI). Among the results,
miR-425 was identified to be highly expressed in tumor tissues. In
addition, miR-425 has been found to promote liver cancer cell
proliferation and migration (4). To
explore the association between miR-425 and clinical prognosis, the
miRBase database (http://www.mirbase.org/) (26) was used for analysis, revealing that
miR-425 was closely associated with poor survival and prognosis in
patients with liver cancer (Fig.
1B). Therefore, miR-425 was chosen for further study. miR-425
expression in liver cancer cells was detected by qPCR, and MIHA
cells were used as the negative control. The results indicated that
miR-425 was abnormally expressed in liver cancer cell lines
compared with in MIHA cells (P<0.05; Fig. 1C). miR-425-knockdown cell lines were
generated and miR-425 expression was detected after transfection
with miR-425 inhibitors (Fig. 1D).
To detect the effect of miR-425 on sorafenib sensitivity, miR-425
inhibitors plasmids were transfected in HepG2 cells, revealing that
knockdown of miR-425 increased the sensitivity of liver cancer
cells to sorafenib (Fig. 1E).
Further experiments indicated that miR-425-knockdown decreased the
migration and invasion of liver cancer cells (Fig. 1F).
Inhibition of miR-425 promotes
lipophagy by regulating autophagy in liver cancer
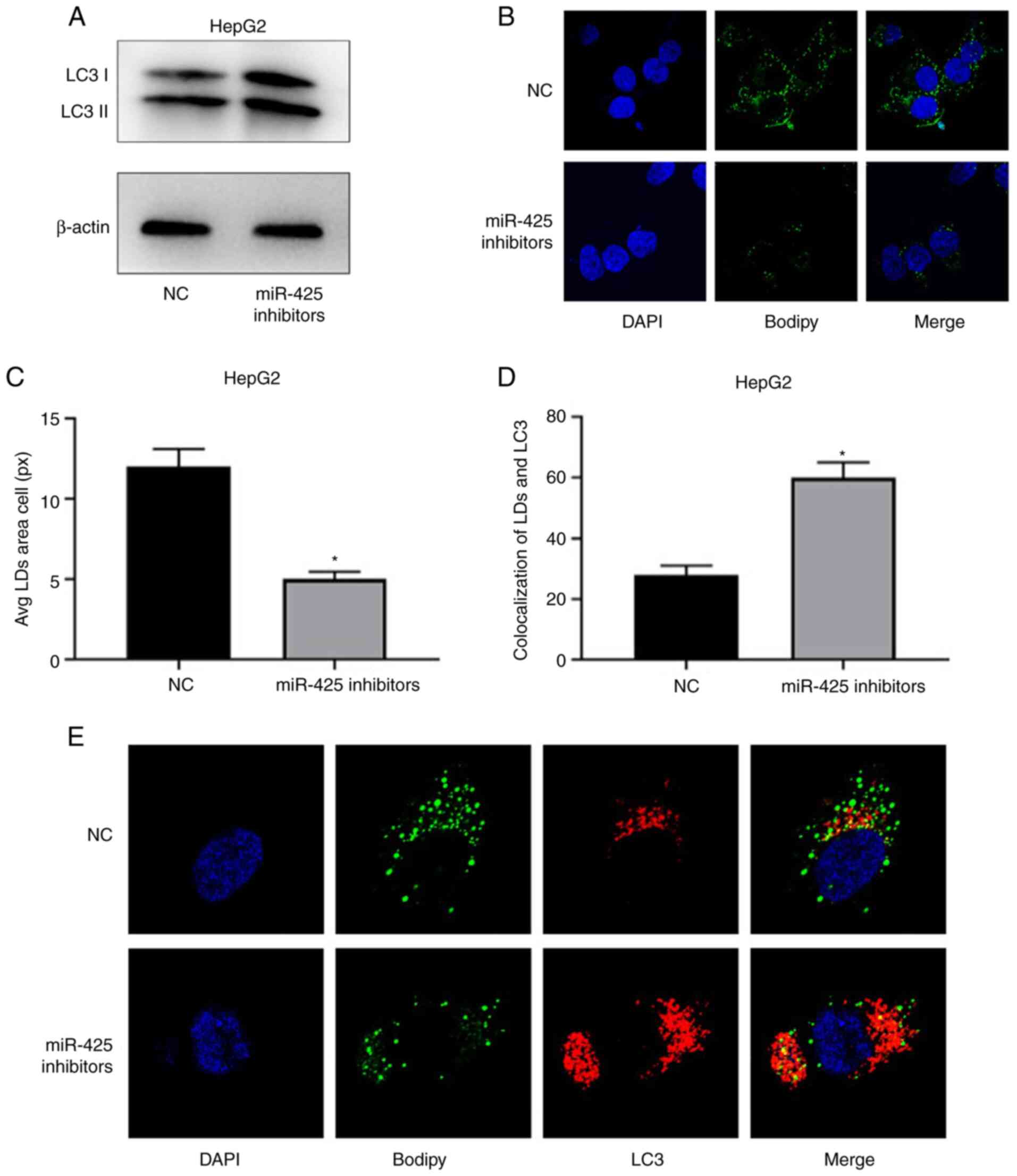
Since an increase in LDs is associated with
chemotherapeutic resistance (27),
the present study evaluated whether inhibition of miR-425 could
induce fat phagocytosis by decreasing the number of LDs. The HepG2
cell line was chosen to exclude the impact of p53 mutations, since
it is p53-wild-type, because p53 has reported to impact LDs
(28). The results indicated
upregulation of LC3 II (a marker of autophagy induction) after
inhibition of miR-425 in HepG2 cells (Fig. 2A). In addition, inhibition of miR-425
significantly decreased the number of LDs (Fig. 2B and C), indicating that miR-425
could regulate lipophagy. Consistent with this observation,
immunofluorescence analysis revealed that inhibition of miR-425
significantly induced the colocalization of LDs and LC3 (si-NC was
the negative control and miR-425 inhibitors is the inhibitors of
miR-425; Fig. 2D and E).
Inhibition of miR-425-induces
lipophagy reverses sorafenib resistance
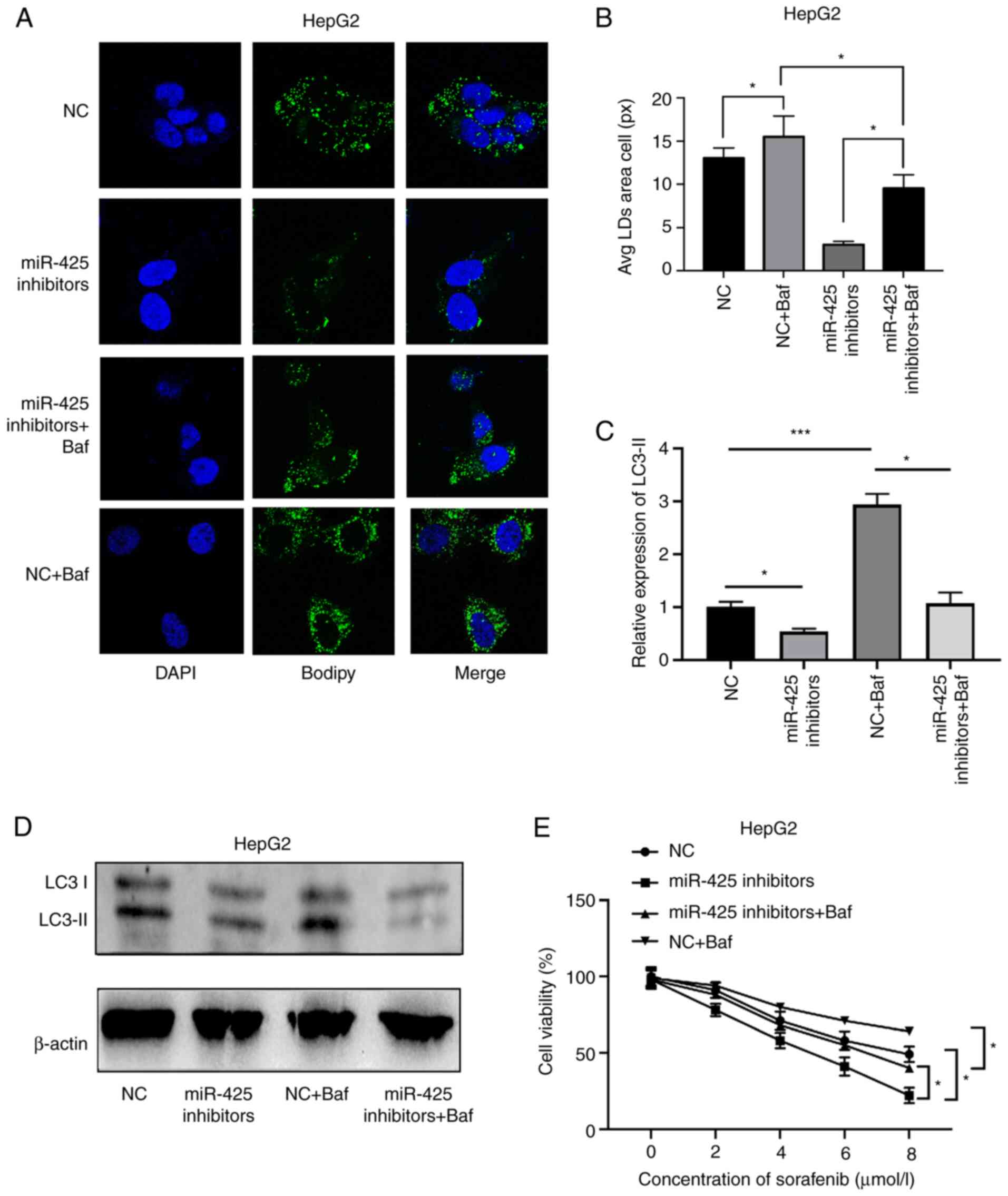
Cancer cells rich in LDs are resistant to
chemotherapeutic drugs (27).
Therefore, to understand whether lipophagy induced by miR-425
inhibitors is necessary for making liver cancer cells become
sensitive to sorafenib-induced cytotoxicity, liver cancer cells
were pretreated with BafA1 for 6 h and cytotoxicity was evaluated.
The results revealed that inhibition of autophagy by BafA1
attenuated the synergistic cytotoxicity of miR-425 inhibitors in
sorafenib-resistant HepG2 cells (Fig. 3A
and B). These results suggested that autophagy induced by
miR-425 inhibitors may promote the synergistic effects of standard
chemotherapeutic drugs in drug-resistant cells. To further confirm
the role of miR-425 in chemoresistance, miR-425 was transiently
knocked down in liver cancer cells, and it was confirmed that the
synergistic effect between miR-425 inhibitors and sorafenib
observed in resistant cells was dependent on autophagy (Fig. 3C and D) (11). The current results indicated that
pretreatment with 25 nM BafA1 for 6 h resulted in increased
resistance to sorafenib in liver cancer cells treated with miR-425
inhibitors compared with the control (Fig. 3E). These results clearly indicate
that autophagy induced by miR-425 inhibitors make liver cancer
cells sensitive to drug-induced cytotoxicity.
miR-425-knockdown promotes SIRT1
expression
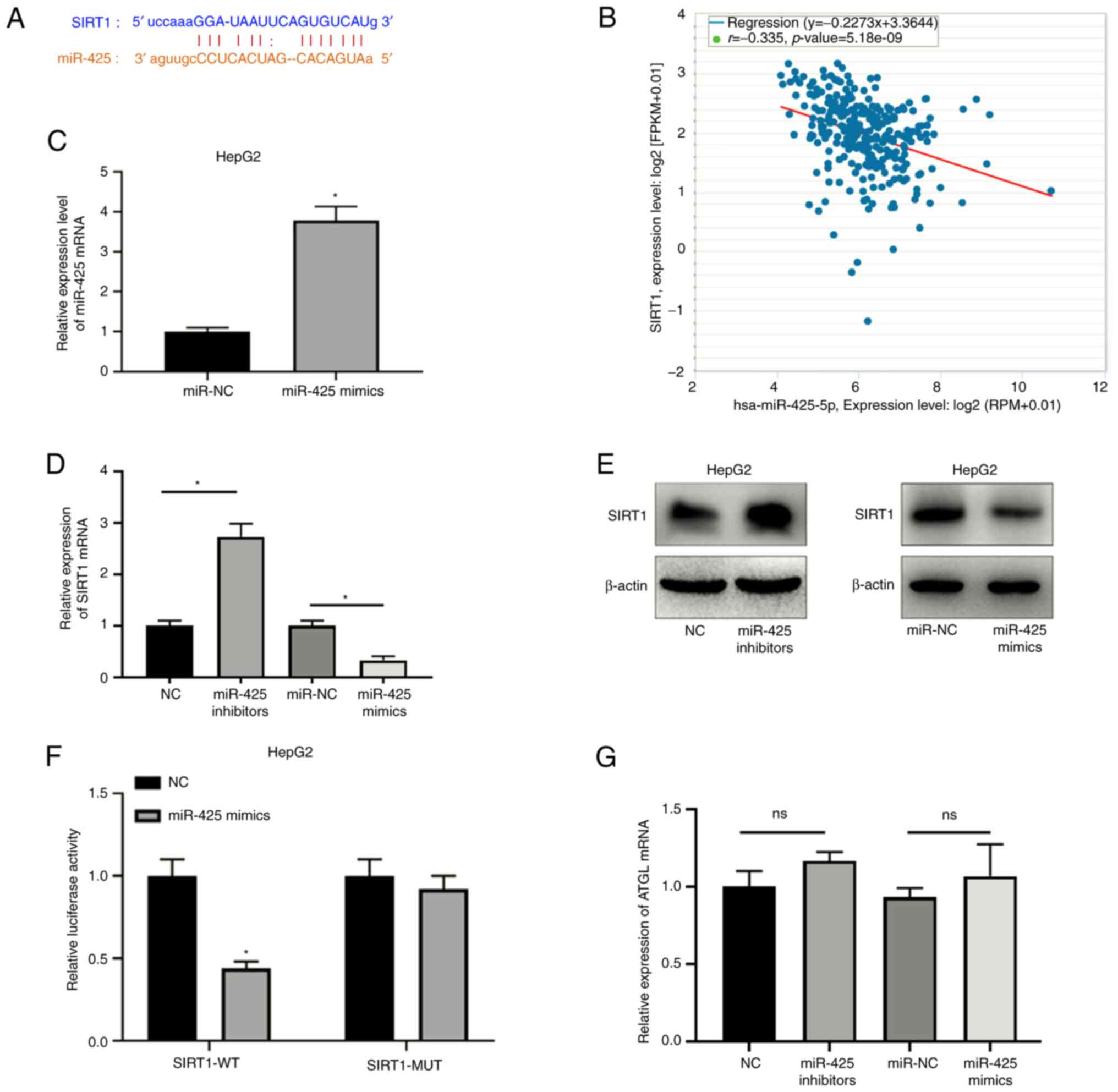
To further explore the targets of miR-425,
bioinformatic analysis using StarBase was performed. SIRT1 was
identified as a potential direct target of miR-425 (Fig. 4A). The correlation between SIRT1 and
miR-425 expression in liver cancer was also analyzed using the
StarBase database, revealing a negative correlation between SIRT1
and miR-425 expression (Fig.
4B).
To verify whether miR-425 regulated SIRT1
expression, miR-425 mimics were used to overexpress miR-425
(Fig. 4C). The RT-qPCR results
indicated that SIRT1 expression was significantly increased after
miR-425-knockdown and significantly decreased after miR-425
overexpression (Fig. 4D), which was
consistent with the western blotting results (Fig. 4E). Subsequently, a luciferase
reporter assay was used to verify whether miR-425 directly targeted
SIRT1. The results revealed that miR-425 significantly inhibited
luciferase activity in cells transfected with the wild-type
SIRT1-3′UTR plasmid, but had no effect on luciferase activity in
cells transfected with the mutant SIRT1-3′ UTR plasmid (Fig. 4F). A previous study has reported that
adipose triglyceride lipase (ATGL) can regulate SIRT1 to mediate
lipophagy (16). Therefore, qPCR was
performed in the present study to evaluate ATGL mRNA expression
after transfection with miR-425 inhibitors or mimics, revealing
that miR-425 modulation did not impact ATGL mRNA expression
(Fig. 4G). The current results
suggested that miR-425 regulated lipophagy independently of ATGL.
Therefore, combining the bioinformatic and experimental results, it
was hypothesized that SIRT1 may be a direct target of miR-425.
miR-425 regulates lipophagy via
SIRT1
To confirm whether miR-425 regulated lipophagy
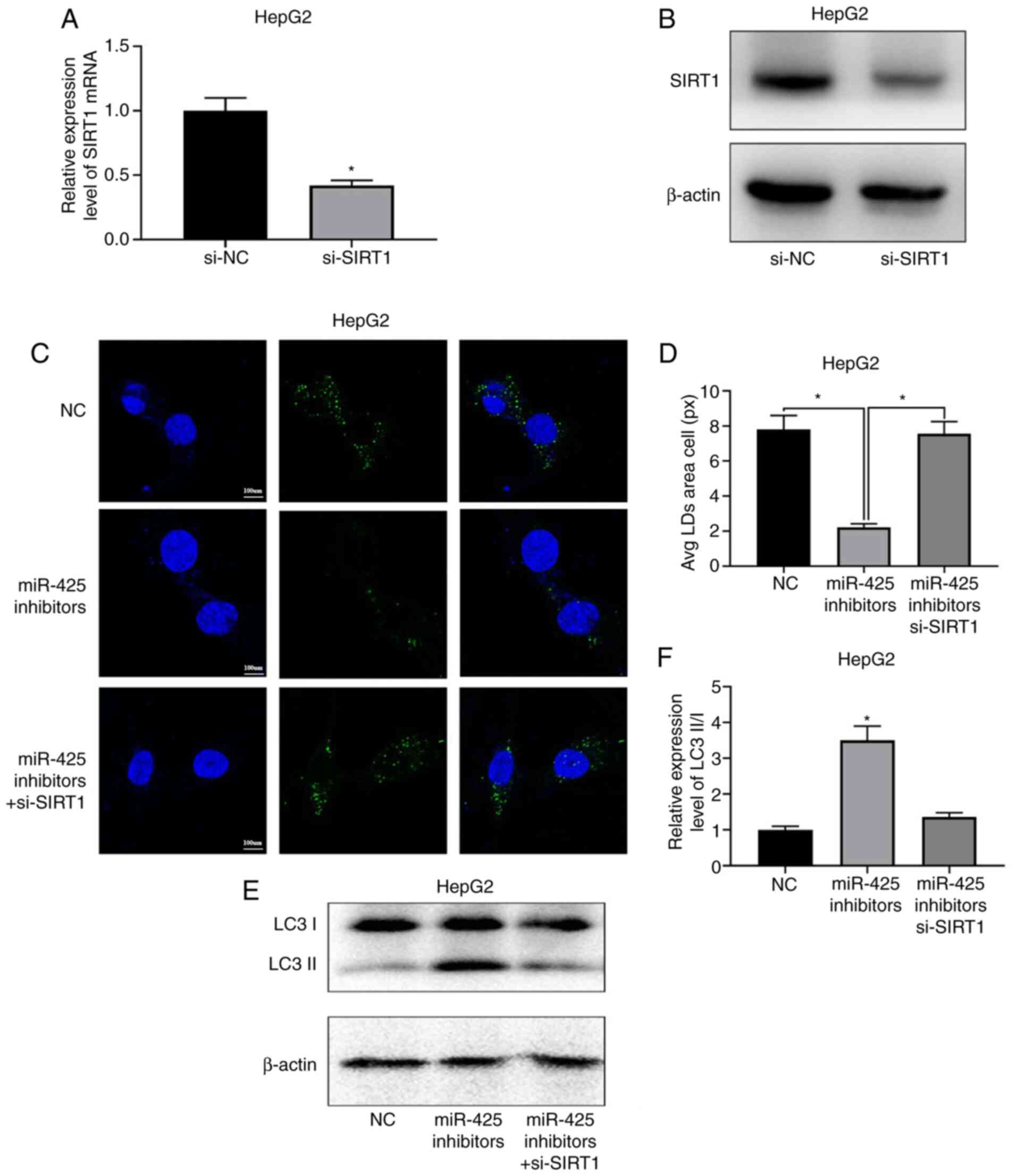
through SIRT1, si-SIRT1 was used to knock down SIRT1 expression,
and SIRT1-knockdown was verified by RT-qPCR and western blot
analysis (Fig. 5A and B).
Subsequently, the miR-425- and SIRT1-knockdown plasmids were
co-transfected into cells, revealing that autophagy and lipophagy
were decreased. The level of LDs was significantly increased in
co-transfected cells compared with that in cells with only
miR-425-knockdown, indicating that LDs were not degraded by
autophagy (Fig. 5C and D). Western
blot analysis of LC3 indicated that autophagy was inhibited in the
co-transfected group compared with the miR-425-knockdown group,
which confirmed the inhibition of lipophagy (Fig. 5E and F).
Discussion
Currently, the treatment of liver cancer mainly
includes surgical resection, radiotherapy and chemotherapy, but the
effects are unsatisfactory (1,2).
Although sorafenib is an effective drug for the treatment of
advanced liver cancer, the benefit that patients receive from
sorafenib treatment is limited due to the development of drug
resistance (3). Therefore, there is
an urgent need for further research on drug resistance. LDs are
spherical organelles with phospholipid monolayer membrane
structures that were initially identified as the main components
dynamically responding to the needs of cell energy metabolism and
regulating the storage of lipid molecules, triglycerides,
cholesterol esters and small amounts of vitamins (27). In recent years, with the in-depth
understanding of cellular lipid metabolism and LD function, the
role of LDs in drug resistance has been increasingly recognized
(11). Studies have demonstrated
that stearoyl-CoA desaturase is highly expressed in various types
of tumors and is associated with cell membrane fluidity and
chemotherapeutic resistance (29–32). In
fact, an increase in LDs is associated with drug resistance in
tumor cells, and this association has been confirmed in breast,
prostate and ovarian cancer (11).
In addition, the expression of components of cancer stem cell
marker signaling pathways is associated with LDs in several types
of cancer, such as glioma, colorectal cancer and HCC (33).
Autophagy serves an important role in malignant
biological behaviors, such as tumor proliferation, invasion,
metastasis and drug resistance (4).
An increasing number of studies have shown that autophagy is
closely associated with the development of tumors (5,34).
Interfering with the autophagy level in tumor cells is a new idea
for tumor therapy (5). For example,
Atg7 inhibitors and docetaxel can be used in combination to treat
breast cancer (34). In liver
cancer, SIRT1 is the only overexpressed member of the SIRT family
and is considered to be essential for all stages of liver cancer
tumorigenesis (35). In addition,
SIRT1 can modulate lipophagy (16),
which is the process of autophagic degradation of LDs (35). By decomposing intracellular lipids
from LDs into fatty acids, lipid phagocytosis provides energy for
cells as well as regulates other cellular processes, such as
activation of carcinogenic signaling pathways and drug resistance
(11). Considering the important
role of LDs and autophagy in liver cancer, lipophagy may be a
factor promoting the process of drug resistance in liver
cancer.
In the present study, mRNA sequencing data for
patients with hepatocellular carcinoma in TCGA database and for
sorafenib-resistant samples in the GEO database were compared to
identify miR-425, and it was revealed that miR-425 was associated
with a poor prognosis. Therefore, a cell line with low miR-425
expression was generated through transfection and the biological
function of miR-425 was analyzed. It has been previously found that
miR-425 regulated the proliferation, invasion and metastasis of
hepatocellular carcinoma cells (36). These results are consistent with the
previous findings that miR-425 can improve cell survival and is
associated with a poor outcome (37,38).
Overexpression of miR-425 promotes cell proliferation and migration
via targeting RNF11 in HCC and it can also bind with FOXD3 to
promote cell migration and invasion (36,37).
These results suggested that miR-425 is an oncogene to promote
tumor progression. However, relatively few studies have
investigated lipophagy and sofafenib resistance in HCC. In the
present study, western blot analysis and immunofluorescence
experiments were used to evaluate the level of lipophagy. The
results revealed that the level of lipophagy decreased after
miR-425 inhibition. Additionally, it was found that miR-425
regulated the drug resistance of liver cancer cells through
lipophagy. By further studying the mechanism by which miR-425
regulated drug resistance, it was revealed that SIRT1 expression, a
key protein in the process of lipophagy, was inhibited after
miR-425 inhibition, and rescue experiments demonstrated that
miR-425 regulated lipophagy through SIRT1. A previous study
demonstrated that SIRT1 expression was high in HCC cell lines,
including HepG2 and Hep3B cells, and its upregulation may result in
increased cell proliferation (39).
The current study identified a specific mechanism of miR-425 on
SIRT1-associated lipophagy, miR-425 inhibitors can increase SIRT1
expression to induce lipophagy which is consistent with previous
findings (16). The aforementioned
results indicated the important role and related mechanism of
miR-425 in lipophagy and drug resistance in liver cancer, and
suggested that miR-425 may become a new target for tumor
therapy.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81871983 and
82003040), the Natural Science Foundation of Liaoning Province
(grant no. LQNK201719) and the Natural Science Foundation of
Liaoning Province (grant no. 2020-BS-103).
Availability of data and materials
The datasets generated and/or analyzed during the
current study are available in the Gene Expression Omnibus
(https://ncbi.nlm.nih.gov/geo)
repository.
Authors' contributions
GS and LY designed the study and performed the
experiments. HJ, SW and BL analyzed the biological data. LY revised
the manuscript. HL conceived the study and participated in
manuscript writing and revisions. GS and LY were responsible for
confirming the authenticity of the raw data. All authors have read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ncRNA
|
non-coding RNA
|
|
SIRT1
|
silent information regulator 2 homolog
1
|
|
NC
|
negative control
|
|
ATG
|
autophagy-related gene
|
References
|
1
|
Chen Z, Xie H, Hu M, Huang T, Hu Y, Sang N
and Zhao Y: Recent progress in treatment of hepatocellular
carcinoma. Am J Cancer Res. 10:2993–3036. 2020.PubMed/NCBI
|
|
2
|
Zhu ZX, Huang JW, Liao MH and Zeng Y:
Treatment strategy for hepatocellular carcinoma in China:
Radiofrequency ablation versus liver resection. Jpn J Clin Oncol.
46:1075–1080. 2016.PubMed/NCBI
|
|
3
|
Perera S, Kelly D and O'Kane GM:
Non-immunotherapy options for the first-line management of
hepatocellular carcinoma: Exploring the evolving role of sorafenib
and lenvatinib in advanced disease. Curr Oncol. 27 (Suppl
3):S165–S172. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
White E, Mehnert JM and Chan CS:
Autophagy, metabolism, and cancer. Clin Cancer Res. 21:5037–5046.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li YJ, Lei YH, Yao N, Wang CR, Hu N, Ye
WC, Zhang DM and Chen ZS: Autophagy and multidrug resistance in
cancer. Chin J Cancer. 36:522017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang Z, Yao Z, Chen Y, Qian L, Jiang S,
Zhou J, Shao J, Chen A, Zhang F and Zheng S: Lipophagy and liver
disease: New perspectives to better understanding and therapy.
Biomed Pharmacother. 97:339–348. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Maan M, Peters JM, Dutta M and Patterson
AD: Lipid metabolism and lipophagy in cancer. Biochem Biophys Res
Commun. 504:582–589. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Madrigal-Matute J and Cuervo AM:
Regulation of liver metabolism by autophagy. Gastroenterology.
150:328–339. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Beloribi-Djefaflia S, Vasseur S and
Guillaumond F: Lipid metabolic reprogramming in cancer cells.
Oncogenesis. 5:e1892016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dong H and Czaja MJ: Regulation of lipid
droplets by autophagy. Trends Endocrinol Metab. 22:234–240. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mondal S, Roy D, Sarkar Bhattacharya S,
Jin L, Jung D, Zhang S, Kalogera E, Staub J, Wang Y, Xuyang W, et
al: Therapeutic targeting of PFKFB3 with a novel glycolytic
inhibitor PFK158 promotes lipophagy and chemosensitivity in
gynecologic cancers. Int J Cancer. 144:178–189. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li X, Zhou Y, Yang L, Ma Y, Peng X, Yang
S, Li H and Liu J: LncRNA NEAT1 promotes autophagy via regulating
miR-204/ATG3 and enhanced cell resistance to sorafenib in
hepatocellular carcinoma. J Cell Physiol. 235:3402–3413. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang S, Wang M, Yang L, Li Y, Ma Y, Peng
X, Li X, Li B, Jin H and Li H: MicroRNA-375 Targets ATG14 to
inhibit autophagy and sensitize hepatocellular carcinoma cells to
sorafenib. Onco Targets Ther. 13:3557–3570. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lapierre LR, Kumsta C, Sandri M, Ballabio
A and Hansen M: Transcriptional and epigenetic regulation of
autophagy in aging. Autophagy. 11:867–880. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Maiese K: Targeting the core of
neurodegeneration: FoxO, mTOR, and SIRT1. Neural Regen Res.
16:448–455. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sathyanarayan A, Mashek MT and Mashek DG:
ATGL promotes autophagy/lipophagy via SIRT1 to control hepatic
lipid droplet catabolism. Cell Rep. 19:1–9. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Masters JR, Thomson JA, Daly-Burns B, Reid
YA, Dirks WG, Packer P, Toji LH, Ohno T, Tanabe H, Arlett CF, et
al: Short tandem repeat profiling provides an international
reference standard for human cell lines. Proc Natl Acad Sci USA.
98:8012–8017. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li HR, Shagisultanova EI, Yamashita K,
Piao Z, Perucho M and Malkhosyan SR: Hypersensitivity of tumor cell
lines with microsatellite instability to DNA double strand break
producing chemotherapeutic agent bleomycin. Cancer Res.
64:4760–4767. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using realtime quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ohata Y, Shimada S, Akiyama Y, Mogushi K,
Nakao K, Matsumura S, Aihara A, Mitsunori Y, Ban D, Ochiai T, et
al: Acquired resistance with epigenetic alterations under Long-Term
antiangiogenic therapy for hepatocellular carcinoma. Mol Cancer
Ther. 16:1155–1165. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Regan-Fendt K, Li D, Reyes R, Yu L, Wani
NA, Hu P, Jacob ST, Ghoshal K, Payne PRO and Motiwala T:
Transcriptomics-Based drug repurposing approach identifies novel
drugs against Sorafenib-Resistant hepatocellular carcinoma. Cancers
(Basel). 12:27302020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pu Z, Zhu Y, Wang X, Zhong Y, Peng F and
Zhang Y: Identification of prognostic biomarkers and correlation
with immune infiltrates in hepatocellular carcinoma based on a
competing endogenous RNA network. Front Genet. 12:5916232021.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fang Q, Li T, Chen P, Wu Y, Wang T, Mo L,
Ou J and Nandakumar KS: Comparative analysis on abnormal methylome
of differentially expressed genes and disease pathways in the
immune cells of RA and SLE. Front Immunol. 12:6680072021.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shi Z, Kong X, Li C, Liu H, Aliagan AI,
Liu L, Shi Y, Shi X, Ma B, Jin R, et al: Bioinformatic analysis of
differentially expressed genes as prognostic markers in
pheochromocytoma and paraganglioma tumors. Genes Genet Syst.
96:55–69. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lu M, Fan X, Liao W, Li Y, Ma L, Yuan M,
Gu R, Wei Z, Wang C and Zhang H: Identification of significant
genes as prognostic markers and potential tumor suppressors in lung
adenocarcinoma via bioinformatical analysis. BMC Cancer.
21:6162021. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kozomara A, Birgaoanu M and
Griffiths-Jones S: MiRBase: From microRNA sequences to function.
Nucleic Acids Res. 47:D155–D162. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Martinez-Lopez N and Singh R: Autophagy
and lipid droplets in the liver. Annu Rev Nutr. 35:215–237. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Thai SF, Jones CP, Robinette BL, Ren H,
Vallanat B, Fisher AA and Kitchin KT: Effects of silver
nanoparticles and silver nitrate on mRNA and microRNA expression in
human hepatocellular carcinoma cells (HepG2). J Nanosci
Nanotechnol. 21:5414–5428. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Noto A, De Vitis C, Pisanu ME, Roscilli G,
Ricci G, Catizone A, Sorrentino G, Chianese G, Taglialatela-Scafati
O, Trisciuoglio D, et al: Stearoyl-CoA-desaturase 1 regulates lung
cancer stemness via stabilization and nuclear localization of
YAP/TAZ. Oncogene. 36:4573–4584. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kikuchi K and Tsukamoto H: Stearoyl-CoA
desaturase and tumorigenesis. Chem Biol Interact. 316:1089172020.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tesfay L, Paul BT, Konstorum A, Deng Z,
Cox AO, Lee J, Furdui CM, Hegde P, Torti FM and Torti SV:
Stearoyl-CoA Desaturase 1 protects ovarian cancer cells from
ferroptotic cell death. Cancer Res. 79:5355–5366. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pisanu ME, Noto A, De Vitis C, Morrone S,
Scognamiglio G, Botti G, Venuta F, Diso D, Jakopin Z, Padula F, et
al: Blockade of Stearoyl-CoA-desaturase 1 activity reverts
resistance to cisplatin in lung cancer stem cells. Cancer Lett.
406:93–104. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yi M, Li J, Chen S, Cai J, Ban Y, Peng Q,
Zhou Y, Zeng Z, Peng S, Li X, et al: Emerging role of lipid
metabolism alterations in cancer stem cells. J Exp Clin Cancer Res.
37:1182018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gong C, Hu C, Gu F, Xia Q, Yao C, Zhang L,
Qiang L, Gao S and Gao Y: Co-delivery of autophagy inhibitor ATG7
siRNA and docetaxel for breast cancer treatment. J Control Release.
266:272–286. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xiong H, Ni Z, He J, Jiang S, Li X, He J,
Gong W, Zheng L, Chen S, Li B, et al: LncRNA HULC triggers
autophagy via stabilizing Sirt1 and attenuates the chemosensitivity
of HCC cells. Oncogene. 36:3528–3540. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wu H, Shang J, Zhan W, Liu J, Ning H and
Chen N: MiR4255p promotes cell proliferation, migration and
invasion by directly targeting FOXD3 in hepatocellular carcinoma
cells. Mol Med Rep. 20:1883–1892. 2019.PubMed/NCBI
|
|
37
|
Rao D, Guan S, Huang J, Chang Q and Duan
S: MiR-425-5p acts as a molecular marker and promoted
proliferation, migration by targeting RNF11 in hepatocellular
carcinoma. Biomed Res Int. 2020:65309732020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Vaira V, Roncalli M, Carnaghi C, Faversani
A, Maggioni M, Augello C, Rimassa L, Pressiani T, Spagnuolo G, Di
Tommaso L, et al: MicroRNA-425-3p predicts response to sorafenib
therapy in patients with hepatocellular carcinoma. Liver Int.
35:1077–1086. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Portmann S, Fahrner R, Lechleiter A, Keogh
A, Overney S, Laemmle A, Mikami K, Montani M, Tschan MP, Candinas D
and Stroka D: Antitumor effect of SIRT1 inhibition in human HCC
tumor models in vitro and in vivo. Mol Cancer Ther. 12:499–508.
2013. View Article : Google Scholar : PubMed/NCBI
|