Introduction
Lung cancer remains the leading cause of
tumor-associated death in the world. Globally, 2.09 million newly
diagnosed cases (accounting for 11.6% of the global cancer burden)
and 1.76 million deaths (18.4% of total cancer deaths) were
estimated in 2018 (1). Non-small
cell lung cancer (NSCLC) represents a heterogeneous group of tumors
that affects >80% of all patients with lung cancer. The majority
of patients with advanced lung cancer are treated with chemotherapy
(2). Platinum-based drugs,
especially cisplatin, are the standard first-line chemotherapy for
numerous types of cancer, including NSCLC (3). However, cancer cells commonly acquire
resistance against cisplatin treatment resulting in high recurrence
rates (4–6). Thus, there is a need to clarify the
molecular mechanisms involved in cisplatin resistance to develop
more effective chemotherapy for NSCLC and other types of
cancer.
Regulated upon Activation Normal T cell Expressed
and Secreted (RANTES), also known as C-C motif chemokine ligand 5
(CCL5), which is now widely recognized as an inflammatory
chemokine, modulates cancer cell migration, metastasis and
chemotherapy resistance in human malignancy (7,8).
Increasing evidence has demonstrated that altered CCL5 expression
is associated with disease progression, aggressiveness, survival,
prognosis and cisplatin resistance in patients with melanoma,
breast, lung and pancreatic cancer (9–11). The
CCL5/C-C motif chemokine receptor 5 (CCR5) axis promotes tumor
progression through a variety of mechanisms: i) CCL5 promotes tumor
growth in an autocrine or paracrine manner (12); ii) CCL5 facilitates cancer cell
migration and invasion by remodeling the tumor microenvironment
(TME) through collagen degradation, integrin activation and actin
polarization (13); iii) CCL5
enhances chemotherapy resistance of cancer cells by increasing
repair responses to DNA damage (14); and iv) CCL5/CCR5 interaction promotes
cancer stem cell expansion (15).
Additionally, CCL5 facilitates cisplatin (DDP) resistance of cancer
cells in various human malignancies, including cervical, breast and
oral cancer (10,16,17).
CCL5 can be produced and secreted by tumors themselves or stromal
cells in the TME (15).
Tumor progression depends on tumor-stromal crosstalk
(18). Cancer-associated fibroblasts
(CAFs) are one of the vital components in the TME. CAFs can remodel
and reprogram the tumoral extracellular matrix (ECM) to facilitate
the growth, metastasis and invasion of the neoplastic lesion,
generating a permissive niche for the invasive cancer cells
(19). CAFs secrete elevated levels
of ECM proteins, such as fibronectin and type I collagen, express
higher level of caveolin-1, which can remodel fibronectin matrix,
and increase the expression and activation of matrix
metallopeptidases (20). These
processes contribute to remodeling the ECM biochemically and
mechanically. Among all CAF functions, the current study focused on
CAFs as important regulators of chemotherapy resistance in
NSCLC.
Long non-coding RNAs (lncRNAs) are described as
mRNA-like, non-protein coding RNA >200 nucleotides in length
(21). lncRNAs regulate diverse
processes, including chromatin dynamics, genomic reprogramming
(22), gene imprinting (23), transcription and post-transcriptional
processing (24,25). Increasing evidence has revealed that
lncRNAs serve a crucial role in the processes of tumor cell
proliferation, differentiation, apoptosis and metastasis (26,27). HOX
transcript antisense RNA (HOTAIR) is one of the few most
deeply-studied lncRNAs (28,29). HOTAIR is aberrantly expressed in
numerous types of cancer, including lung cancer, and has been
recognized as an oncogene in cancer development (28). Higher HOTAIR expression has been
associated with tumor metastasis and poor prognosis in lung cancer
(30–32). Recently, HOTAIR has been reported to
be involved in cisplatin resistance of cancer cells (33,34).
However, it is currently unclear whether HOTAIR serves a part in
cisplatin resistance in NSCLC and its respective mechanism.
The present study aimed to investigate whether CAFs
isolated from patients with NSCLC created a supportive TME that
affected the sensitivity of NSCLC cells to cisplatin.
Materials and methods
Isolation of primary fibroblasts
The present study was approved by the medical
ethical committee of Hanchuan People's Hospital (Hanchuan, China).
Written informed consent was obtained from patients. Human CAFs
were isolated from primary tumor tissues, and normal fibroblasts
(NFs) were isolated from adjacent normal tissues (5 cm away from
the tumor tissues), respectively, from two patients with NSCLC at
pT2N0M0 (female, aged 61 years) and pT1N0M0 (male, aged 63 years)
stage undergoing surgical resection at Hanchuan People's Hospital
between February 2019 and October 2019 with informed consent from
the patients. Primary fibroblasts were isolated within 2 h after
excision according to a previously published study (35). Briefly, the fresh tissues were cut
into small pieces ~1×1×1 mm in size and were digested with 1.5
mg/ml collagenase IV (Roche Applied Science) and 20 mg/ml trypsin
(Sigma-Aldrich; Merck KGaA) at 37°C for 1 h. Subsequently, the
mixture was filtered through a 40-µm mesh (Falcon; Corning Life
Sciences) and the cells were maintained in DMEM/F12 medium
containing 10% FBS (both Gibco; Thermo Fisher Scientific, Inc.),
100 U/ml penicillin and 100 µg/ml streptomycin at 37°C in 5%
CO2. The first passage was conducted when cells reached
~80% confluency. The cells (after 2–3 passages) were analyzed by
immunofluorescence, quantitative (q)PCR and western blotting for
α-smooth muscle actin (α-SMA), fibroblast activation protein (FAP)
and fibroblast specific protein 1 (FSP1), which are highly
expressed in fibroblasts, to confirm the homology of
fibroblasts.
To prepare conditioned medium of CAFs (CAF-CM) and
NFs (NF-CM) for subsequent in vitro experiments, CAFs and
NFs at a density of 2×105 were plated into a
25-cm2 culture flask in 5 ml RPMI-1640 medium (Gibco;
Thermo Fisher Scientific, Inc.) supplemented with 10% FBS at 37°C
in 5% CO2 and cultured for 24 h. Subsequently, the
medium was replaced with RPMI-1640 with 0.5% FBS for another 24 h,
after which the culture medium was collected and centrifuged at
3,000 × g at 4°C for 10 min. The supernatant was collected as CM
and stored at −80°C until further use.
Cell lines and cultivation
NSCLC A549 (lung adenocarcinoma) and H1299 (lung
large cell carcinoma) cell lines were purchased from the China
Center for Type Culture Collection, and cultivated in RPMI-1640
medium supplemented with 10% FBS, 100 U/ml penicillin and 100 µg/ml
streptomycin in a humidified incubator with 5% CO2 at
37°C. Cells in the logarithmic growth phase were used for all
experiments.
For cell treatment, cancer cells were incubated with
CAF-CM or NF-CM in combination with either anti-CCL5 antibody (0.1
µg/ml; cat. no. MAB678-SP; R&D Systems, Inc.), CCR5 antagonist
(Met-RANTES; 0.1 µg/ml; cat. no. 335-RM-025; R&D Systems, Inc.)
or recombinant human CCL5 (3 ng/ml; cat. no. 300-06; PeproTech,
Inc.) for 6 h, followed by treatment with 50 µM DDP (Sigma-Aldrich;
Merck KGaA) in the presence of CM for another 48 h.
Cell transfection
The small interfering RNA (siRNA) against HOTAIR
(siHOTAIR) and non-targeting control siRNA (siNC) were purchased
from Shanghai GenePharma Co., Ltd.. The sequences of the two siRNAs
were as follows: siHOTAIR forward, 5′-AUUGAUUAGCUGUUUGUUCCC-3′ and
reverse, 5′-AAAGUCUAGACAAUAGAUGGC-3′; siNC forward,
5′-CUAUUGUCUAGACUUUUAUCU-3′ and reverse,
5′-GAAAUCUGGUACAAAGGAAAG-3′. Cells were seeded into 6-well plates
to 40–60% confluence and then transfected with siHOTAIR or siNC at
a concentration of 60 nM using Lipofectamine 2000®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) and Opti-MEM I
Reduced Serum Medium (Thermo Fisher Scientific, Inc.) following the
manufacturer's protocol. At 36 h after transfection, cells were
collected for subsequent experiments.
Cell viability analysis
Cell viability was determined using the MTT assay.
Briefly, A549 or H1299 cells were grown in 96-well plates at a
density of 5×103 cells/well and treated with 50 µM DDP
for 48 h. Subsequently, 20 µl MTT (5 µg/µl) was added to each well
at 37°C for 4 h. The reaction was stopped by the addition of DMSO,
and the optical density (OD) was detected at 490 nm by Multiscan
Spectrum (Bio-Tek Instruments, Inc.; Agilent Technologies, Inc.).
All experiments were repeated three times, and the average OD for
each experiment was calculated.
Analysis of apoptosis by flow
cytometry
Apoptosis was determined using an Annexin V-FITC/PI
apoptosis detection kit (BD Pharmingen; BD Biosciences) according
to the manufacturer's protocol. Briefly, 6×105 cells
were seeded into 6-well plates and incubated with DDP (50 µM) for
24 h. Subsequently, cells were harvested and incubated with
FITC-Annexin V and propidium iodide (PI) at room temperature for 15
min in the dark. The apoptotic rate was analyzed using BD FACScan
flow cytometer (Becton-Dickinson and Company). The data were
analyzed using the CellQuest software (version 5.1;
Becton-Dickinson and Company) Cells positive for Annexin V-FITC
alone (early apoptosis) and Annexin V-FITC/PI (late apoptosis) were
calculated. All samples were examined in triplicate.
ELISA
The quantity of CCL5 in CM of CAFs, NFs, A549 and
H1299 cells was determined using a commercial ELISA kit. Briefly,
~1×106 cells in 3 ml serum-free RPMI 1640 medium were
seeded in a 25-cm2 flask for 48 h. Subsequently, CM was
collected and CCL5 quantity was assessed using human RANTES ELISA
Kit (CCL5) following the manufacturer's instructions (cat. no.
ab174446; Abcam). CCL5 quantities were normalized to the cell
number.
Western blotting
The cells were lysed with RIPA buffer containing 1
mM PMSF and protease inhibitor cocktail according to the
manufacturer's instructions (Beyotime Institute of Biotechnology).
Protein concentrations were quantified using a BCA kit (Beyotime
Institute of Biotechnology). Total protein (30 µg/lane) was
separated by 10% SDS-PAGE and transferred to polyvinylidene
difluoride membranes. After blocking with 5% skimmed milk in TBS
containing 0.5% Tween-20 (TBST) for 1 h at room temperature, the
membranes were incubated with the following primary antibodies
overnight at 4°C: β-actin (1:1,000; cat. no. ab8226), E-cadherin
(1:5,000; cat. no. ab40772), Vimentin (1:1,000; cat. no. ab92547),
FAP (1:500; cat. no. ab53066), FSP1 (1:1,000; cat. no. ab197896)
(all Abcam), α-SMA (1:1,000; cat. no. 19245), Bcl-2 (1:1,000; cat.
no. 15071), cleaved-poly (ADP-ribose) polymerase (PARP; 1:1,000;
cat. no. 5625), PARP (1:1,000, cat. no. 9532), cleaved-caspase-3
(1:1,000, cat. no. 9661) and caspase-3 (1:1,000; cat. no. 9662)
(all Cell Signaling Technology, Inc.). After washing three times
for 5 min each with TBST, the membranes were incubated with the
appropriate HRP-conjugated secondary antibodies (1:2,000; cat. nos.
7076 and 7074, Cell Signaling Technology, Inc.) for 1 h at room
temperature. The signal was visualized using chemiluminescent
reagents according to the manufacturer's protocols
(MilliporeSigma), and the protein bands were quantified using
ImageJ software (Version 1.51; National Institutes of Health).
Reverse transcription (RT)-qPCR
For the RNA expression analysis, TRIzol®
(Invitrogen; Thermo Fisher Scientific, Inc.) was used to extract
total RNA in A549 cells, and the RNA level of each group was
examined. Subsequently, 5 µg RNA was reverse transcribed to cDNA
using an RT kit (Promega Corporation) following the manufacturer's
protocol. qPCR was performed with the GoTaq qPCR Master Mix
(Promega Corporation) on an Mx3005P Real-Time PCR System
(Stratagene; Agilent Technologies, Inc.). The thermocycling
conditions were as follows: 95°C for 10 min, followed by 40 cycles
of 95°C for 15 sec, 60°C for 30 sec and 72°C for 30 sec. The primer
sequences were as follows: HOTAIR forward,
5′-GATCAGATGCCTGGGTCGAA-3′ and reverse, 5′-AATGATTCTTGCTGGGGGCA-3′;
α-SMA forward, 5-TGCCTGCTCTCTGATGTTGG-3 and reverse,
5-GCTACCGAGCCCTGAGTTAC-3; FAP forward, 5-ATGAGCTTCCTCGTCCAATTCA-3
and reverse, 5-AGACCACCAGAGAGCATATTTTG-3; FSP1 forward,
5-GATGAGCAACTTGGACAGCAA-3 and reverse, 5-CTGGGCTGCTTATCTGGGAAG-3;
CCL5 forward, 5-CCAGCAGTCGTCTTTGTCAC-3 and reverse,
5-CTCTGGGTTGGCACACACTT-3; CCR5 forward, 5-TTCTGGGCTCCCTACAACATT-3
and reverse, 5-TTGGTCCAACCTGTTAGAGCTA-3 and GAPDH forward,
5′-GGAGCGAGATCCCTCCAAAAT-3′ and reverse,
5′-GGCTGTTGTCATACTTCTCATGG-3′. Each experiment was performed at
least three times. The gene expression levels were calculated using
the 2−ΔΔCq method (36)
GAPDH was used as an internal reference.
Immunofluorescence staining
Cells were washed three times in cold PBS, and then
fixed with 4% paraformaldehyde for 10 min at room temperature.
After permeabilization with PBS supplemented with 0.25% of Triton
X-100 for 10 min at room temperature, the cells were washed three
times in PBS and blocked with 1% BSA in PBS for 60 min at room
temperature. The following primary antibodies were used for
immunofluorescence staining, overnight at 4°C: anti-E-cadherin
(1:100, cat. no. 14472), anti-Vimentin (1:100, cat. no. 5741) and
anti-α-SMA (1:200, cat. no. 48938) (all from Cell Signaling
Technology, Inc.). The cells were washed again with PBS and
incubated with Alexa Fluor® 488- or Alexa
Fluor® 594-conjugated secondary antibodies (1:500, cat.
no. 4408, and 1:500 cat. no. 8890 (respectively), Cell Signaling
Technology, Inc.). DAPI (1:1,000, Beyotime Institute of
Biotechnology) was used for counterstaining. The images were
captured by a fluorescence microscope (Olympus Corporation).
Statistical analysis
Each experiment was performed at least three times
independently. All data were presented as the mean ± SD. SPSS 20.0
software (IBM Corp.) was used for data analysis. Comparisons
between two groups were performed using unpaired Student's t-test,
and one-way ANOVA followed by Bonferroni's post-hoc test was used
to examine differences among ≥3 groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
Characterization of primary NFs and
CAFs
Human CAFs were isolated from primary tumor tissues,
and normal fibroblasts (NFs) were isolated from adjacent normal
tissues, respectively, from two patients with NSCLC. However, the
specimen from the patient at stage pT1N0M0 was contaminated due to
improper operation during isolation of the NFs and CAFs. Therefore,
the NFs and CAFs used in subsequent experiments were all from the
other patient at stage pT2N0M0. The mesenchymal marker vimentin and
the epithelial marker E-cadherin were used to identify homogeneity
of isolated cells. Isolated CAFs highly expressed vimentin, but
expressed lower levels of E-cadherin compared with NFs, as measured
by western blot analysis (Fig. 1A).
Additionally, the expression levels of α-SMA, FAP and FSP1 in CAFs
were 1.47-, 2.84- and 2.60-fold higher than those in NFs (Fig. 1A). It is commonly agreed that CAFs
are activated fibroblasts that express mesenchyme-specific proteins
α-SMA, FAP and FSP1 (37,38). RT-qPCR analysis confirmed that the
expression levels of α-SMA, FAP and FSP1 were significantly higher
in CAFs than in NFs (Fig. 1B).
Furthermore, immunocytochemistry staining confirmed that CAFs and
NFs both expressed vimentin, and that α-SMA expression was higher
in CAFs than in NFs (Fig. 1C).
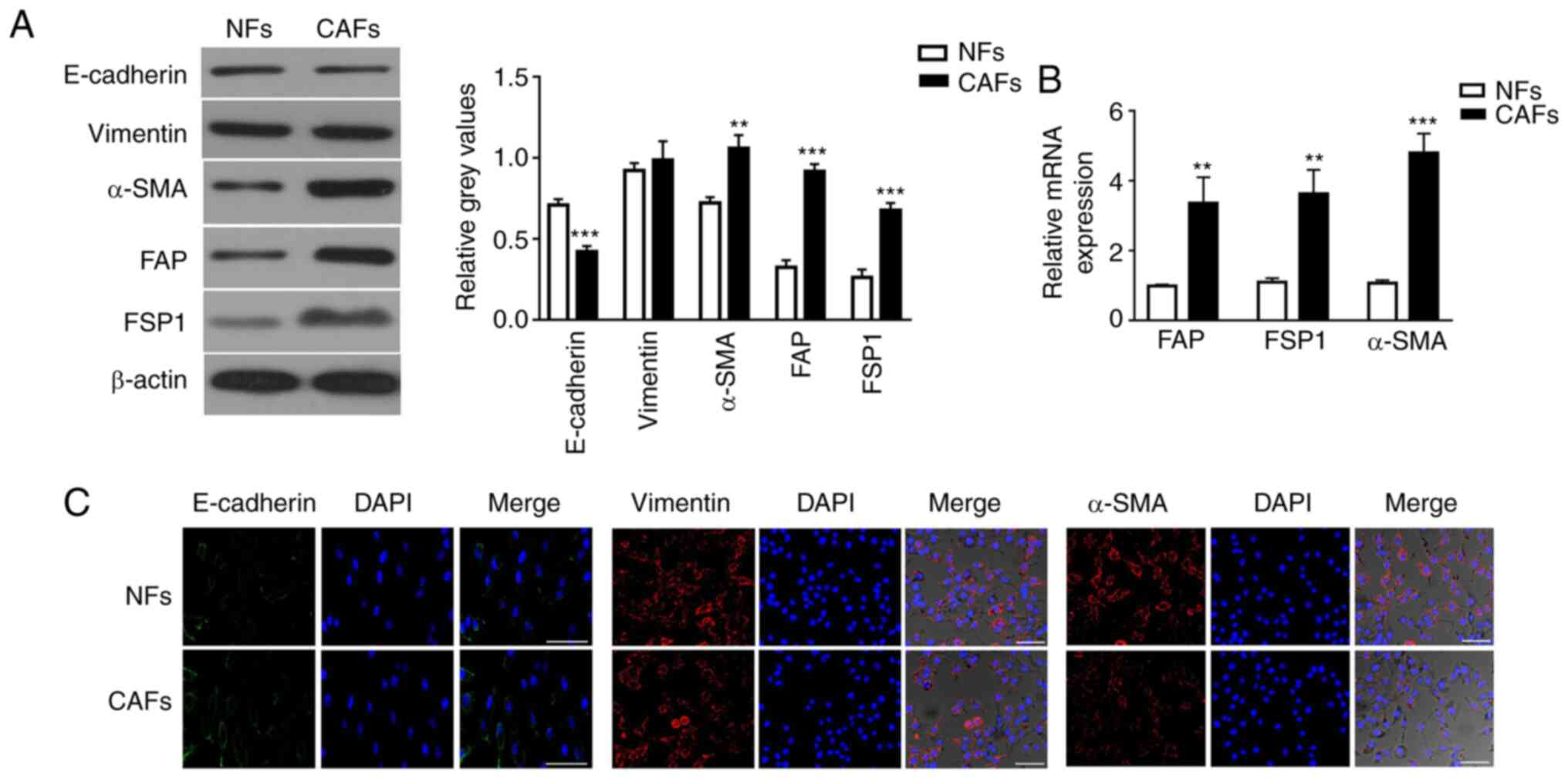 | Figure 1.Characterization of primary isolated
and cultivated NFs and CAFs. (A) Protein expression levels of
E-cadherin, vimentin, α-SMA, FAP and FSP1 in NFs and CAFs were
detected by western blotting. The intensity of each experimental
band was normalized to that of the loading control (β-actin). Each
bar represents the mean ± SD of three relative intensity arbitrary
units. (B) mRNA expression levels of CAF-specific genes, including
α-SMA, FAP and FSP1, in NFs and CAFs were detected by reverse
transcription-quantitative PCR using GAPDH gene as the internal
control. Results were expressed as the mean ± SD, and the means
were calculated from ≥3 independent experiments. (C)
Immunofluorescence staining showed the subcellular location and the
expression of E-cadherin, vimentin and α-SMA in NFs and CAFs. Scale
bar, 10 µm. **P<0.01 and ***P<0.001 vs. NFs. CAF,
cancer-associated fibroblast; NF, normal fibroblast; α-SMA,
α-smooth muscle actin; FAP, fibroblast activation protein; FSP1,
fibroblast specific protein 1. |
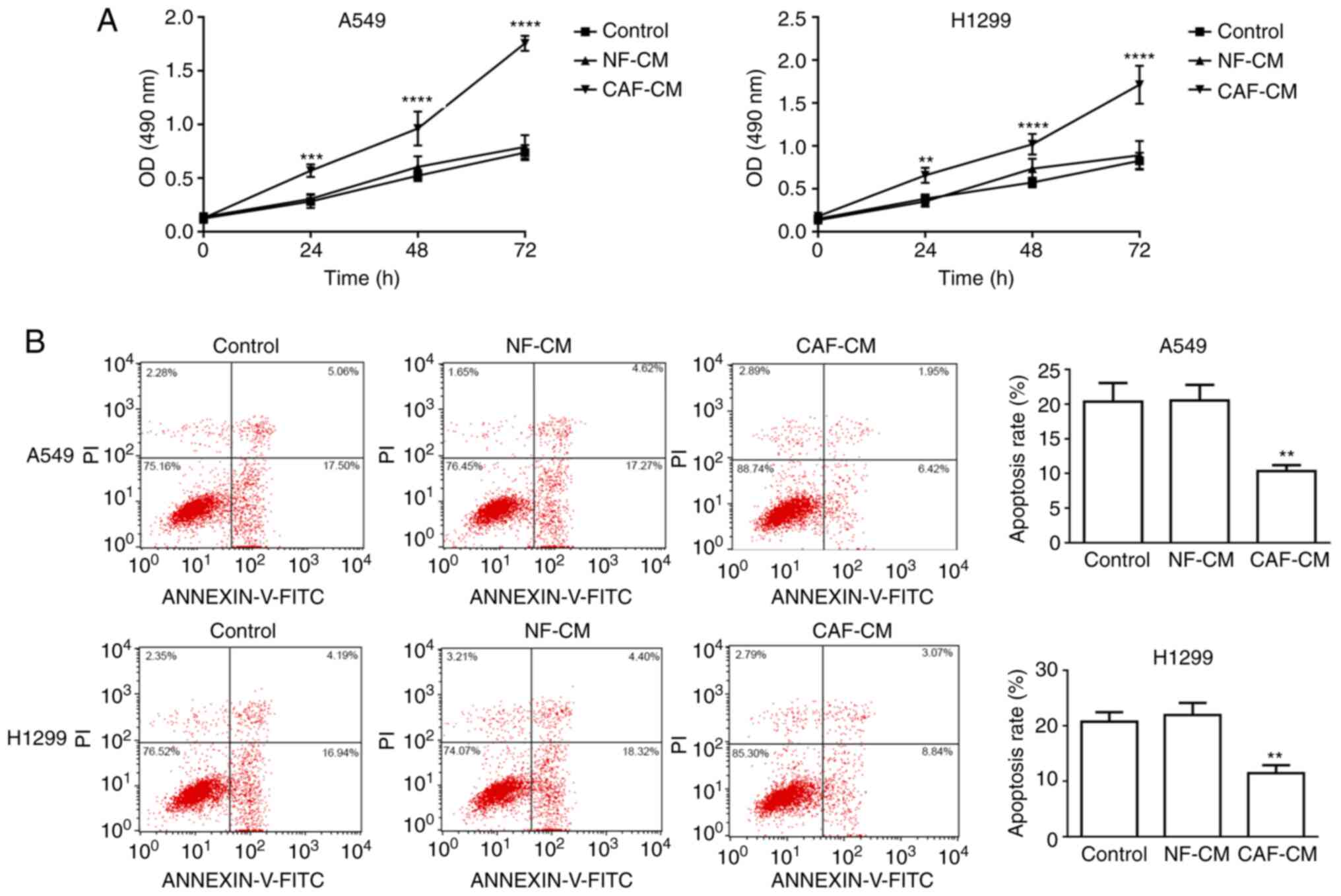
CAF-CM promotes NSCLC cell resistance
to cisplatin
Considering the crucial role of the tumor stroma on
tumor growth and metastasis, the present study assessed whether
factors secreted by CAFs could affect the viability and apoptosis
of A549 cells. First, the role of CAF-CM on the viability and
apoptosis of A549 cells was analyzed. A549 and H1299 cells were
pre-incubated with CAF-CM or NF-CM for 24 h, and then treated with
DDP in the presence of CM for another 48 h. MTT assay revealed that
A549 and H1299 cells exhibited significantly higher cell viability
with CAF-CM treatment than with NF-CM treatment (Fig. 2A). To demonstrate the effect of
CAF-CM on apoptosis, A549 and H1299 cells were pre-treated with
CAF-CM or NF-CM for 24 h before the addition of DDP in the presence
of CM for another 48 h, and the percentage of apoptotic cells was
analyzed using Annexin V-FITC/PI apoptosis staining. The apoptotic
rates of A549 cells treated with control, NF-CM and CAF-CM were
20.35±2.71, 20.52±2.27 and 10.35±0.85%, respectively, while the
apoptotic rates of H1299 cells treated with control, NF-CM and
CAF-CM were 20.71±1.72, 21.93±2.19 and 11.43±1.48%, respectively,
revealing that apoptosis was significantly decreased by CAF-CM
(Fig. 2B). Overall, these data
indicated that CAF-CM attenuated the pro-apoptotic effect induced
by chemotherapy in NSCLC cells.
Paracrine effect of CCL5 in
CAF-CM
In order to determine whether CAF-secreted CCL5 may
induce cisplatin resistance in NSCLC cells, RT-qPCR was performed
to assess CCL5 mRNA expression in NFs, CAFs and the two NSCLC cell
lines, A549 and H1299, revealing that CCL5 expression was
significantly higher in CAFs compared with in all other tested
cells (Fig. 3A). Additionally, ELISA
results confirmed significantly higher levels of CCL5 in CAF-CM
compared with in NF-CM (359.8±23.47 vs. 276±11.77 pg/ml,
respectively) and in A549 and H1299 cells (Fig. 3B). In addition, incubation with
CAF-CM induced resistance to DDP in NSCLC cells, which was blocked
by the anti-CCL5 antibody (Fig.
3C).
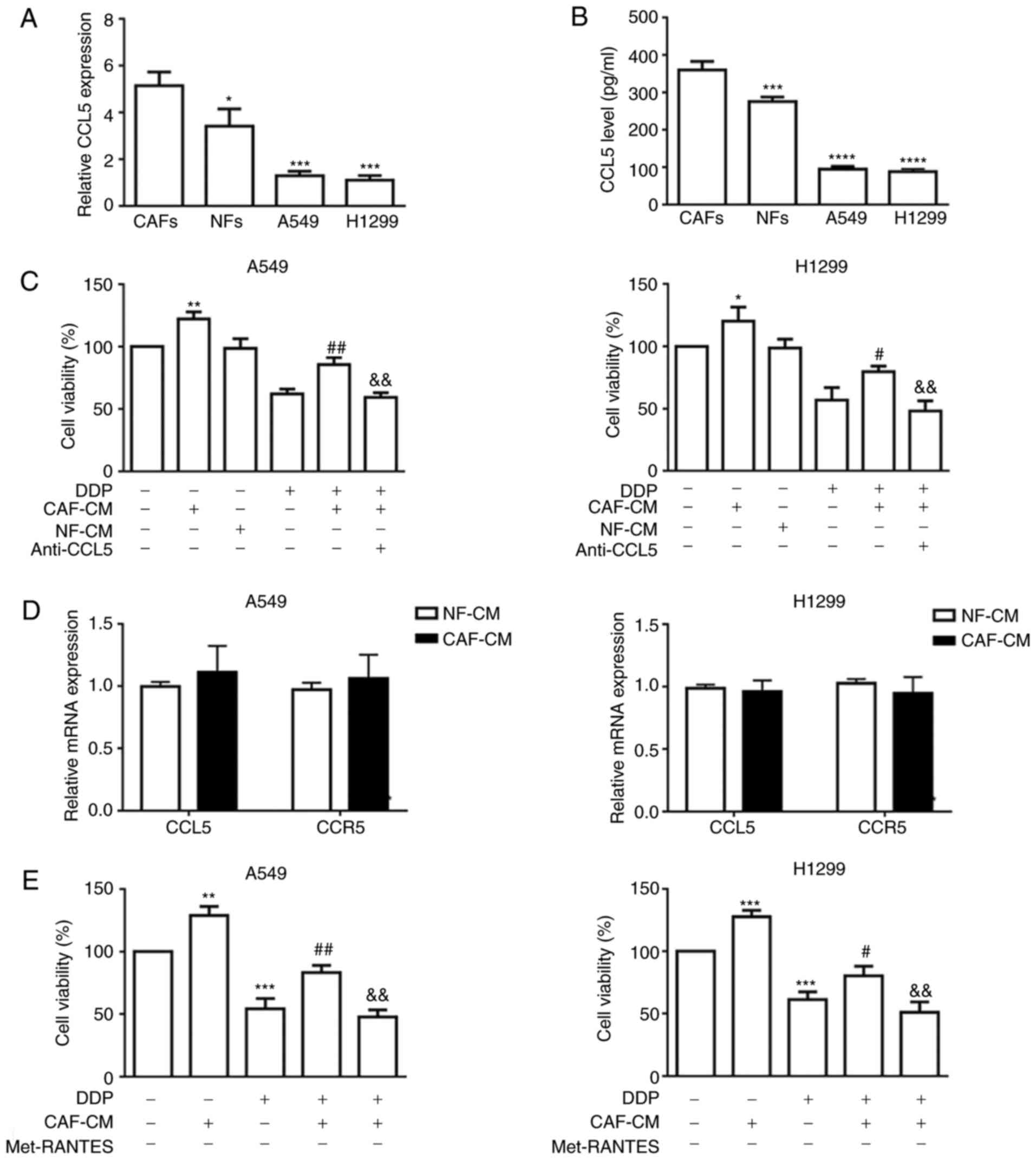 | Figure 3.CAFs secrete elevated CCL5 to promote
cell viability in NSCLC cancer cells via paracrine activation. (A)
mRNA expression levels of CCL5 in CAFs, NFs and two NSCLC cell
lines by RT-qPCR using GAPDH gene as the internal control. (B) CCL5
in conditioned medium secreted by NFs and CAFs were quantified by
ELISA. *P<0.05, ***P<0.001 and ****P<0.0001 vs. CAFs. (C)
A549 and H1299 cancer cells were incubated with CAF-CM or NF-CM in
combination with anti-CCL5 antibody (0.1 µg/ml) for 6 h followed by
treatment with DDP for 48 h. Cell viability was determined by the
MTT assay. (D) mRNA expression levels of CCL5 and CCR5 in NSCLC
cell lines cultured with CAF-CM were assessed by RT-qPCR using
GAPDH gene as the normalization control. (E) A549 and H1299 cancer
cells were incubated with CAF-CM or NF-CM in combination with CCR5
inhibitor (Met-RANTES; 0.1 µg/ml) for 6 h followed by treatment
with DDP for 48 h. Cell viability was determined by the MTT assay.
*P<0.05, **P<0.01 and ***P<0.001 vs. control;
#P<0.05 and ##P<0.01 vs. DDP;
&&P<0.01 vs. CAF-CM and DDP. Results were
expressed as the mean ± SD, and the means were calculated from ≥3
independent experiments. RT-qPCR, reverse
transcription-quantitative PCR; CAF, cancer-associated fibroblast;
NF, normal fibroblast; CM, conditioned medium; DDP, cisplatin;
CCL5, C-C motif chemokine ligand 5; CCR5, C-C motif chemokine
receptor 5; NSCLC, non-small cell lung cancer. |
To estimate whether the activation of CCL5 signaling
in NSCLC cells was mainly caused by paracrine action of CCL5
secreted by CAFs, the expression levels of CCL5 and CCR5 in A549
and H1299 cells, in the presence or absence of CAF-CM, were
examined by RT-qPCR. As shown in Fig.
3D, neither CCL5 nor CCR5 expression was markedly altered in
the CAF-CM group compared with in the NF-CM group, indicating that
CCL5 pathway activation triggered by CAF-CM was not the direct
consequence of autocrine or reverse-paracrine mechanisms. Paracrine
activation of CCL5 by CAF-CM was further verified by the CCL5
blocking assays using a neutralizing anti-CCL5 antibody or a CCR5
antagonist (Met-RANTES). Cell viability results indicated that
either blockade of CCL5 binding to its cognate receptor (Fig. 3C) or inhibition of CCR5 (Fig. 3E) alleviated the CAF-CM-induced
cisplatin resistance, indicating that CCL5 alone may be sufficient
to facilitate cisplatin resistance in NSCLC cell lines.
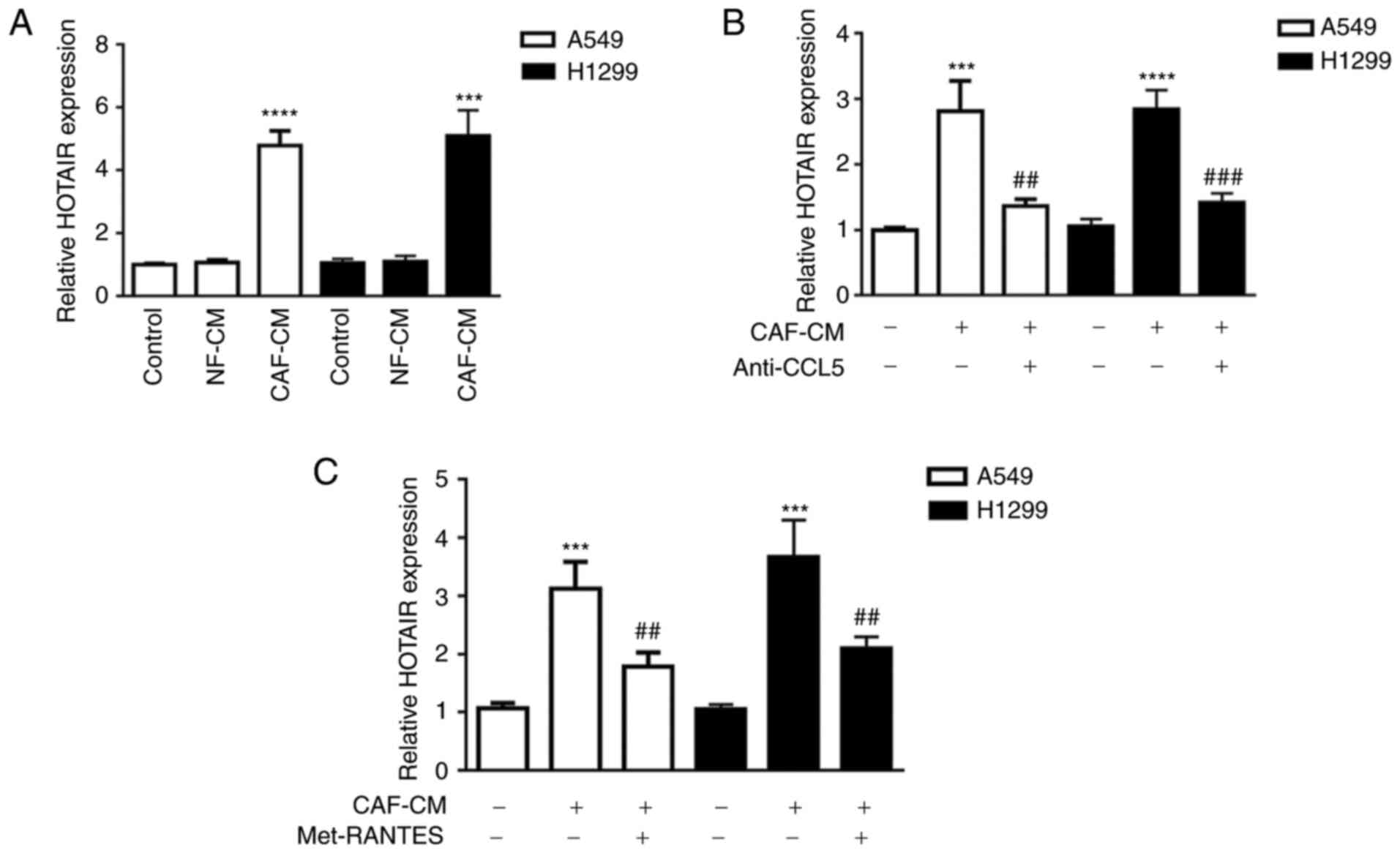
CAF-CM-derived CCL5 suppresses DDP
sensitivity of NSCLC cells partially by upregulating lncRNA HOTAIR
expression
Previous data have shown that HOTAIR is expression
is markedly increased in human DDP-resistant NSCLC tissues and
cells. Silencing of HOTAIR partially reverses the acquired
cisplatin resistance in DDP-resistant NSCLC cells, both in
vitro and in vivo (39).
Thus, it was hypothesized that CCL5 may induce tumor cell
resistance to cisplatin via regulation of lncRNA HOTAIR expression.
To verify this, the expression levels of lncRNA HOTAIR in A549 and
H1299 cell lines were examined by RT-qPCR. As shown in Fig. 4A, HOTAIR expression in tumor cells
was significantly increased when cells were incubated with CAF-CM
compared with NF-CM stimulation. Additionally, anti-CCL5 antibody
treatment significantly decreased HOTAIR expression by 2.06 and
2.00-fold in A549 and H1299 cells, respectively, compared with
CAF-CM treatment alone (Fig. 4B).
Similarly, the CCR5 antagonist (Met-RANTES) significantly
downregulated HOTAIR expression by 1.74 and 1.75-fold in A549 and
H1299 cells, respectively, compared with CAF-CM treatment alone
(Fig. 4C). These results
demonstrated that CCL5 was at least partially involved in promoting
lncRNA HOTAIR expression in both A549 and H1299 cells.
Recently, lncRNA HOTAIR has been found to serve a
role in tumor cell proliferation, migration and invasion (30,40,41).
Moreover, based on the association of increased lncRNA HOTAIR
expression with NSCLC advanced pathological stage, lymph-node
metastasis and poor prognosis (42),
the present study hypothesized that HOTAIR may serve a role in
NSCLC cell resistance to cisplatin. Therefore, its expression was
silenced using RNA interference to estimate the effect of HOTAIR on
tumor cell sensitivity to cisplatin. qPCR analysis revealed that
HOTAIR mRNA expression was significantly downregulated in
siHOTAIR-transfected A549 cells compared with in control cells
transfected with siNC (Fig. 5A).
Additionally, cisplatin treatment significantly decreased HOTAIR
expression in siNC-transfected cells compared with siNC-transfected
cells without cisplatin treatment (Fig.
5A). Furthermore, the effects of HOTAIR on NSCLC cell
sensitivity to cisplatin were assessed by MTT assay. Consistent
with the aforementioned hypothesis, silencing HOTAIR in A549 cells
significantly sensitized these cells to cisplatin compared with
tumor cells transfected with siNC (Fig.
5B). Additionally, the apoptotic rate of A549 cells transfected
with siHOTAIR was significantly higher than that in A549 cells
transfected with siNC with CAF-CM (21.87±2.70 vs. 12.07±2.45%,
respectively), as detected by flow cytometric analysis (Fig. 5C). These results indicated that
silencing of lncRNA HOTAIR inhibited NSCLC cell viability and
increased apoptosis in vitro.
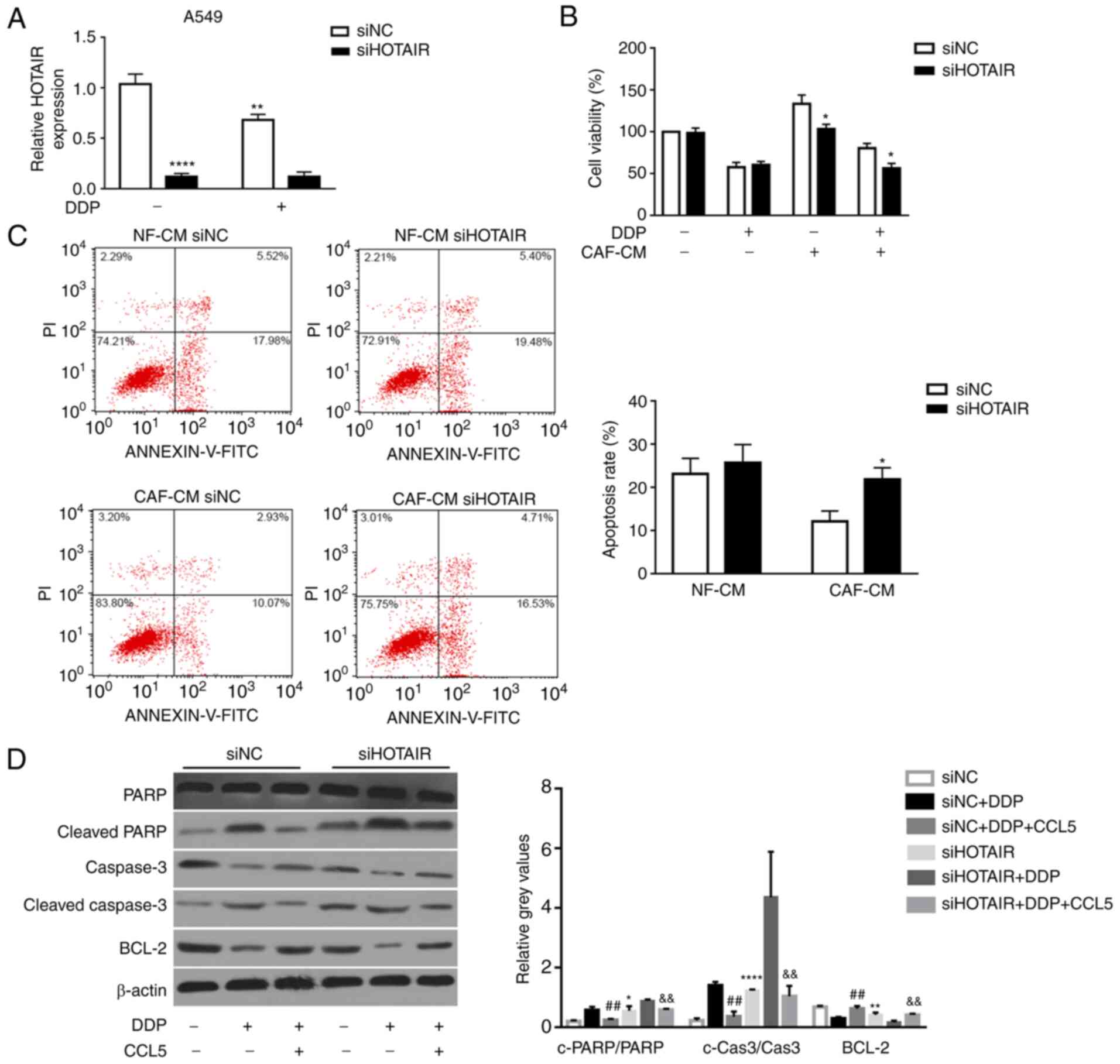 | Figure 5.HOTAIR-knockdown abrogates
CCL5-induced cisplatin resistance via activation of
caspase-3-dependent apoptosis. (A) HOTAIR expression in A549 cells
transfected with either siNC or siHOTAIR was determined by
quantitative PCR. (B) Cell viability and (C) apoptosis of DDP- and
CAF-CM-treated A549 cells transfected with either siNC or siHOTAIR
was determined by MTT and flow cytometric assays, respectively.
*P<0.05, **P<0.01 and ****P<0.0001 vs. siNC. (D) Protein
expression levels of PARP, cleaved-PARP, Bcl-2, cleaved caspase-3
and caspase-3 in siNC- or siHOTAIR-transfected A549 cell lines with
or without treatment with exogenous CCL5 (100 ng/ml) were
determined by western blotting. The intensity of each experimental
band was normalized to that of the loading control (β-actin). Each
bar represents the mean ± SD of three relative intensity arbitrary
units. **P<0.01 vs. siNC; ##P<0.01 vs. siNC+DDP;
&&P<0.01 vs. siHOTAIR+DDP. CAF,
cancer-associated fibroblast; NF, normal fibroblast; CM,
conditioned medium; DDP, cisplatin; CCL5, C-C motif chemokine
ligand 5; HOTAIR, HOX transcript antisense RNA; si, small
interfering RNA; NC, negative control; PARP, poly (ADP-ribose)
polymerase. |
HOTAIR-knockdown suppresses
CCL5-induced cisplatin resistance through activation of the
PARP-dependent apoptotic signaling pathway
Some studies (43,44) have
revealed that lncRNA HOTAIR is involved in enhancing cancer cell
proliferation by suppressing apoptosis (45). Therefore, the present study
investigated whether silencing HOTAIR could abrogate the
CCL5-induced cisplatin resistance in NSCLC cell lines through the
apoptotic signaling pathway. The expression levels of
apoptosis-associated proteins, including cleaved PARP and cleaved
caspase-3, in siNC- or siHOTAIR-transfected NSCLC cells upon CCL5
treatment (100 ng/ml) were examined by western blotting. The
results revealed that cleaved PARP and cleaved caspase-3 expression
was significantly upregulated in A549 cells transfected with
siHOTAIR compared with in cells with siNC transfection (Fig. 5D). Moreover, the expression levels of
cleaved PARP and cleaved caspase-3 were significantly decreased in
siNC transfected-A549 cells, which were pre-treated with CCL5 for
24 h before the treatment of DDP, compared with siNC
transfected-A549 cells with DDP treatment alone (Fig. 5D). By contrast, the expression levels
of the anti-apoptotic protein Bcl-2 were significantly upregulated
in the siNC+DDP+CCL5 groups compared with that in the siNC+DDP
group (Fig. 5D). Additionally, Bcl-2
expression was significantly reduced in siHOTAIR transfected-A549
cells compared with that in siNC transfected-A549 cells (Fig. 5D). These results suggested that
HOTAIR-knockdown promoted NSCLC cell sensitivity to DDP via the
apoptotic signaling pathway.
Overall, the current findings indicated that
CAF-derived CCL5 may serve a role in tumor cells by paracrine
action, upregulating lncRNA HOTAIR expression in tumor cells and
thus upregulating the expression levels of the anti-apoptotic
protein Bcl-2, ultimately leading to NSCLC cell resistance to DDP.
Therefore, the present study may provide a novel therapeutic
strategy for patients with NSCLC with acquired DDP resistance.
Discussion
Cisplatin-based chemotherapy schedule is a
first-line treatment against NSCLC, but cisplatin resistance among
patients with NSCLC has become increasingly severe, contributing to
chemotherapy failure and high recurrence and mortality rates
(46). The TME serves a crucial role
in tumor progression, metastasis and recurrence. CAFs, a dominant
component of the TME, serve a prominent functional role in
promoting cancer initiation, progression and therapeutic resistance
(47). In the present study, it was
demonstrated that CAFs from primary NSCLC tumor tissues had typical
myofibroblast characteristics, with high expression levels of
α-SMA, FAP and FSP1 (48,49), which were very low or undetectable in
NFs. Additionally, it was revealed that CAF-CM could increase the
viability of the NSCLC A549 and H1299 cell lines, and inhibit the
induction of apoptosis by cisplatin in these cells. However, the
specimen from one patient was contaminated during isolation of the
NFs and CAFs. The lack of additional CAFs/NFs cell lines to support
the findings of the present study is a limitation of the present
study. Further studies are necessary to isolate NFs and CAFs from
more patients with NSCLC and repeat some key experiments of the
present study. A previous study has indicated that the interaction
between CAFs and tumor cells affects chemoresistance by inducing
the secretion of survival factors (50,51). For
instance, elevated secretion of IL-6 from CAFs mediates
chemoresistance in NSCLC by promoting epithelial-mesenchymal
transition (51). In the present
study, higher levels of CCL5 were detected in the CAF-CM. However,
CCL5 also has some anticancer properties, since it promotes
antitumor immunity by inducing the recruitment of anticancer
tumor-infiltrating lymphocytes (52), natural killer cells (53), dendritic cells (54), T helper cell type 1 and type 1
cytotoxic cells (55) to the TME,
and therefore enhances the infiltration of the tumor by different
types of immune cells. Usually, the elevated expression of a given
chemokine can improve the outcome for one type of cancer (56), but promote the progression of other
types of cancer. There is no single CC chemokine that facilitates
or suppresses the progression of all types of cancer. Thus, a
thorough understanding of the pro-cancer and anticancer features of
individual chemokines may contribute to a prediction of the
outcomes to improve the efficacies of anticancer therapies.
Although the role of CCL5 in cancer has been extensively studied,
to the best of our knowledge, no such studies have been conducted
to date to investigate the role of CAF-secreted CCL5 in
chemoresistance of NSCLC. The present study described that CAFs
isolated from NSCLC tumor tissues expressed and secreted higher
amounts of CCL5 than NFs, and CAF-derived CCL5 enhanced tumor cell
resistance to cisplatin via paracrine activation in NSCLC A549 and
H1299 cell lines. Notably, CCL5 was substantially expressed by both
CAFs and NFs (at least twice higher than A549 cells). However,
NF-CM did not induce HOTAIR expression in A549 or H1299 cells. A
possible explanation may be that CCL5 is secreted from CAFs and
transferred to cancer cells mainly via exosomes rather than via a
soluble form, and CAF-derived exosomal CCL5 may be much higher than
CCL5 in NF-derived exosomes; thus, NF-CM seemed to have no effects
on HOTAIR expression in A549 or H1299 cells in the present study.
However, this speculation needs to be confirmed by further
research. Li et al (57) has
observed a significant upregulation of TGF-β1 in CAF-derived
exosomes compared with in normal omentum fibroblasts-derived
exosomes, which subsequently activated SMAD2/3 signaling to promote
an EMT phenotype in ovarian cancer cells. However, whether
CAF-derived CCL5 exerts its effect on A549 or H1299 cells via
exosomes requires to be further investigated. Notably, treatment
with a neutralizing CCL5 antibody or CCR5 inhibitor, Met-RANTES,
reversed CAF-CM-induced cancer cell chemoresistance. No significant
upregulation of CCL5 or CCR5 expression was detected in cancer
cells under CAF-CM treatment compared with that under NF-CM
treatment, further confirming that CCL5 secreted by CAFs is
necessary to induce NSCLC cancer cell chemoresistance. Another
limitation of the present study is the lack of clinical evidence to
confirm the influence of CCL5 expression on the survival and
disease-free outcome of patients with NSCLC. Further studies are
required to conduct in silico studies evaluating public
datasets to evaluate the influence of CCL5 expression on the
outcome of patients with NSCLC as evidence to corroborate the
current findings.
Emerging evidence has revealed that numerous lncRNAs
exert crucial functions in tumorigenesis, indicating that they
could provide new insights into the biology of this disease. Among
these lncRNAs, lncRNA HOTAIR is one of the well-studied lncRNAs in
various types of cancer, including lung cancer, and is reported to
be dysregulated in various types of cancer. Upregulation of HOTAIR
expression has been found in cervical cancer tissues and has been
associated with lymph node metastasis and decreased overall
survival (58). In diffuse large
B-cell lymphoma, high HOTAIR expression is positively associated
with a poor prognosis (59). In
breast cancer, high expression levels of HOTAIR promote cancer
development and progression (60).
However, the expression profile of lncRNA HOTAIR in NSCLC and its
association with cisplatin resistance are not fully understood. The
present study indicated that HOTAIR expression was significantly
increased in two NSCLC A549 and H1299 cell lines stimulated with
CAF-CM when compared with that in these cells stimulated with
NF-CM. Furthermore, silencing of HOTAIR in NSCLC cells partly
blocked CCL5-induced cisplatin resistance. Additionally,
HOTAIR-knockdown suppressed cell viability and enhanced apoptosis.
These data indicated that HOTAIR served a crucial role in cell
proliferation and that its high expression was positively
associated with cisplatin resistance. Numerous studies (61–63) have
indicated that lncRNAs serve a crucial role in the occurrence and
progression of several types of cancer, including NSCLC, acting as
a reservoir for microRNAs and regulating their downstream target
genes (64). It has been reported
that HOTAIR targets polycomb repressive complex 2 and regulates
H3K27 methylation and gene expression, which ultimately promotes
tumor progression and metastasis (65,66). The
present study revealed that knockdown of HOTAIR abrogated
CCL5-induced cisplatin resistance in NSCLC cells by upregulating
cleaved caspase-3 and cleaved PARP levels and suppressing Bcl-2
expression.
In conclusion, the current study demonstrated an
association between CAF-derived CCL5, HOTAIR and chemoresistance.
CAF-derived CCL5 inhibited cisplatin-induced apoptosis and induced
tumor cell cisplatin resistance. Moreover, CCL5 enhanced tumor cell
chemoresistance by upregulating lncRNA HOTAIR expression and
resulting in increased anti-apoptotic protein Bcl-2 expression in
cancer cells. The present results supported CAF-secreted CCL5 as a
crucial player in cisplatin-based chemoresistance. Therefore,
targeting the CAF-derived CCL5 axis may be of clinical significance
for reversing cisplatin resistance in patients with NSCLC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XS and ZC made substantial contributions to
conception and design, acquisition of data, or analysis and
interpretation of data, took part in drafting the article or
revising it critically for important intellectual content, and
agreed to submit to the current journal. Both authors have read and
approved the final manuscript, agree to be accountable for all
aspects of the work and confirm the authenticity of all the raw
data.
Ethics approval and consent to
participate
The present study was approved by the medical
ethical committee of Hanchuan People's Hospital (Hanchuan, China).
Written informed consent was obtained from patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bade BC and Dela Cruz CS: Lung cancer
2020: Epidemiology, etiology, and prevention. Clin Chest Med.
41:1–24. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, DeSantis C, Virgo K, Stein K,
Mariotto A, Smith T, Cooper D, Gansler T, Lerro C, Fedewa S, et al:
Cancer treatment and survivorship statistics, 2012. CA Cancer J
Clin. 62:220–241. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rossi A and Di Maio M: Platinum-based
chemotherapy in advanced non-small-cell lung cancer: Optimal number
of treatment cycles. Expert Rev Anticancer Ther. 16:653–660. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Olaussen KA, Dunant A, Fouret P, Brambilla
E, André F, Haddad V, Taranchon E, Filipits M, Pirker R, Popper HH,
et al: DNA repair by ERCC1 in non-small-cell lung cancer and
cisplatin-based adjuvant chemotherapy. N Engl J Med. 355:983–991.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mitsudomi T, Morita S, Yatabe Y, Negoro S,
Okamoto I, Tsurutani J, Seto T, Satouchi M, Tada H, Hirashima T, et
al: Gefitinib versus cisplatin plus docetaxel in patients with
non-small-cell lung cancer harbouring mutations of the epidermal
growth factor receptor (WJTOG3405): An open label, randomised phase
3 trial. Lancet Oncol. 11:121–128. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Scagliotti GV, Parikh P, von Pawel J,
Biesma B, Vansteenkiste J, Manegold C, Serwatowski P, Gatzemeier U,
Digumarti R, Zukin M, et al: Phase III study comparing cisplatin
plus gemcitabine with cisplatin plus pemetrexed in
chemotherapy-naive patients with advanced-stage non-small-cell lung
cancer. J Clin Oncol. 26:3543–3551. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ben-Baruch A: Inflammation-associated
immune suppression in cancer: The roles played by cytokines,
chemokines and additional mediators. Semin Cancer Biol. 16:38–52.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Karnoub AE, Dash AB, Vo AP, Sullivan A,
Brooks MW, Bell GW, Richardson AL, Polyak K, Tubo R and Weinberg
RA: Mesenchymal stem cells within tumour stroma promote breast
cancer metastasis. Nature. 449:557–563. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Luboshits G, Shina S, Kaplan O, Engelberg
S, Nass D, Lifshitz-Mercer B, Chaitchik S, Keydar I and Ben-Baruch
A: Elevated expression of the CC chemokine regulated on activation,
normal T cell expressed and secreted (RANTES) in advanced breast
carcinoma. Cancer Res. 59:4681–4687. 1999.PubMed/NCBI
|
|
10
|
Niwa Y, Akamatsu H, Niwa H, Sumi H, Ozaki
Y and Abe A: Correlation of tissue and plasma RANTES levels with
disease course in patients with breast or cervical cancer. Clin
Cancer Res. 7:285–289. 2001.PubMed/NCBI
|
|
11
|
Yaal-Hahoshen N, Shina S, Leider-Trejo L,
Barnea I, Shabtai EL, Azenshtein E, Greenberg I, Keydar I and
Ben-Baruch A: The chemokine CCL5 as a potential prognostic factor
predicting disease progression in stage II breast cancer patients.
Clin Cancer Res. 12:4474–4480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Aldinucci D, Borghese C and Casagrande N:
The CCL5/CCR5 axis in cancer progression. Cancers (Basel).
12:17652020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Aldinucci D and Casagrande N: Inhibition
of the CCL5/CCR5 axis against the progression of gastric cancer.
Int J Mol Sci. 19:14772018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhou B, Sun C, Li N, han W, Lu H, Guo L,
Guo E, Xia M, Weng D, Meng L, et al: Cisplatin-induced CCL5
secretion from CAFs promotes cisplatin-resistance in ovarian cancer
via regulation of the STAT3 and PI3K/Akt signaling pathways. Int J
Oncol. 48:2087–2097. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huang R, Wang S, Wang N, Zheng Y, Zhou J,
Yang B, Wang X, Zhang J, Guo L, Wang S, et al: CCL5 derived from
tumor-associated macrophages promotes prostate cancer stem cells
and metastasis via activating β-catenin/STAT3 signaling. Cell Death
Dis. 11:2342020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chuang JY, Yang WH, Chen HT, Huang CY, Tan
TW, Lin YT, Hsu CJ, Fong YC and Tang CH: CCL5/CCR5 axis promotes
the motility of human oral cancer cells. J Cell Physiol.
220:418–426. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Velasco-Velázquez M, Jiao X, De La Fuente
M, Pestell TG, Ertel A, Lisanti MP and Pestell RG: CCR5 antagonist
blocks metastasis of basal breast cancer cells. Cancer Res.
72:3839–3850. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Koontongkaew S: The tumor microenvironment
contribution to development, growth, invasion and metastasis of
head and neck squamous cell carcinomas. J Cancer. 4:66–83. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kalluri R and Zeisberg M: Fibroblasts in
cancer. Nat Rev Cancer. 6:392–401. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Erdogan B and Webb DJ: Cancer-associated
fibroblasts modulate growth factor signaling and extracellular
matrix remodeling to regulate tumor metastasis. Biochem Soc Trans.
45:229–236. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ernst C and Morton CC: Identification and
function of long non-coding RNA. Front Cell Neurosci. 7:1682013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Loewer S, Cabili MN, Guttman M, Loh YH,
Thomas K, Park IH, Garber M, Curran M, Onder T, Agarwal S, et al:
Large intergenic non-coding RNA-RoR modulates reprogramming of
human induced pluripotent stem cells. Nat Genet. 42:1113–1117.
2010. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gregg C, Zhang J, Weissbourd B, Luo S,
Schroth GP, Haig D and Dulac C: High-resolution analysis of
parent-of-origin allelic expression in the mouse brain. Science.
329:643–648. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Guttman M and Rinn JL: Modular regulatory
principles of large non-coding RNAs. Nature. 482:339–346. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Batista PJ and Chang HY: Long noncoding
RNAs: Cellular address codes in development and disease. Cell.
152:1298–1307. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhao W, Geng D, Li S, Chen Z and Sun M:
LncRNA HOTAIR influences cell growth, migration, invasion, and
apoptosis via the miR-20a-5p/HMGA2 axis in breast cancer. Cancer
Med. 7:842–855. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shao TR, Zheng ZN, Chen YC, Wu QQ, Huang
GZ, Li F, Zeng WS and Lv XZ: LncRNA AC007271.3 promotes cell
proliferation, invasion, migration and inhibits cell apoptosis of
OSCC via the Wnt/β-catenin signaling pathway. Life Sci.
239:1170872019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang J, Zhang P, Wang L, Piao HL and Ma
L: Long non-coding RNA HOTAIR in carcinogenesis and metastasis.
Acta Biochim Biophys Sin (Shanghai). 46:1–5. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wan Y and Chang HY: HOTAIR: Flight of
noncoding RNAs in cancer metastasis. Cell Cycle. 9:3391–3392. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhao W, An Y, Liang Y and Xie XW: Role of
HOTAIR long noncoding RNA in metastatic progression of lung cancer.
Eur Rev Med Pharmacol Sci. 18:1930–1936. 2014.PubMed/NCBI
|
|
31
|
Nakagawa T, Endo H, Yokoyama M, Abe J,
Tamai K, Tanaka N, Sato I, Takahashi S, Kondo T and Satoh K: Large
noncoding RNA HOTAIR enhances aggressive biological behavior and is
associated with short disease-free survival in human non-small cell
lung cancer. Biochem Biophys Res Commun. 436:319–324. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu MY, Li XQ, Gao TH, Cui Y, Ma N, Zhou Y
and Zhang GJ: Elevated HOTAIR expression associated with cisplatin
resistance in non-small cell lung cancer patients. J Thorac Dis.
8:3314–3322. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Guo J, Dou D, Zhang T and Wang B: HOTAIR
promotes cisplatin resistance of osteosarcoma cells by regulating
cell proliferation, invasion, and apoptosis via miR-106a-5p/STAT3
axis. Cell Transplant. Aug 6–2020.(Epub ahead of print). doi:
10.1177/0963689720948447. View Article : Google Scholar
|
|
34
|
Zhang Y, Ai H, Fan X, Chen S, Wang Y and
Liu L: Knockdown of long non-coding RNA HOTAIR reverses cisplatin
resistance of ovarian cancer cells through inhibiting
miR-138-5p-regulated EZH2 and SIRT1. Biol Res. 53:182020.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li J, Guan J, Long X, Wang Y and Xiang X:
mir-1-mediated paracrine effect of cancer-associated fibroblasts on
lung cancer cell proliferation and chemoresistance. Oncol Rep.
35:3523–3531. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Augsten M: Cancer-associated fibroblasts
as another polarized cell type of the tumor microenvironment. Front
Oncol. 4:622014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yi Y, Zeng S, Wang Z, Wu M, Ma Y, Ye X,
Zhang B and Liu H: Cancer-associated fibroblasts promote
epithelial-mesenchymal transition and EGFR-TKI resistance of
non-small cell lung cancers via HGF/IGF-1/ANXA2 signaling. Biochim
Biophys Acta Mol Basis Dis. 1864:793–803. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhan Y, Abuduwaili K, Wang X, Shen Y,
Nuerlan S and Liu C: Knockdown of long non-coding RNA HOTAIR
suppresses cisplatin resistance, cell proliferation, migration and
invasion of DDP-resistant NSCLC Cells by targeting
miR-149-5p/doublecortin-like kinase 1 axis. Cancer Manag Res.
12:7725–7737. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Huang J, Ke P, Guo L, Wang W, Tan H, Liang
Y and Yao S: Lentivirus-mediated RNA interference targeting the
long noncoding RNA HOTAIR inhibits proliferation and invasion of
endometrial carcinoma cells in vitro and in vivo. Int J Gynecol
Cancer. 24:635–642. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Pei CS, Wu HY, Fan FT, Wu Y, Shen CS and
Pan LQ: Influence of curcumin on HOTAIR-mediated migration of human
renal cell carcinoma cells. Asian Pac J Cancer Prev. 15:4239–4243.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liu XH, Liu ZL, Sun M, Liu J, Wang ZX and
De W: The long non-coding RNA HOTAIR indicates a poor prognosis and
promotes metastasis in non-small cell lung cancer. BMC Cancer.
13:4642013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Qiu JJ, Wang Y, Ding JX, Jin HY, Yang G
and Hua KQ: The long non-coding RNA HOTAIR promotes the
proliferation of serous ovarian cancer cells through the regulation
of cell cycle arrest and apoptosis. Exp Cell Res. 333:238–248.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li D, Chai L, Yu X, Song Y, Zhu X, Fan S,
Jiang W, Qiao T, Tong J, Liu S, et al: The HOTAIRM1/miR-107/TDG
axis regulates papillary thyroid cancer cell proliferation and
invasion. Cell Death Dis. 11:2272020. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li D, Feng J, Wu T, Wang Y, Sun Y, Ren J
and Liu M: Long intergenic noncoding RNA HOTAIR is overexpressed
and regulates PTEN methylation in laryngeal squamous cell
carcinoma. Am J Pathol. 182:64–70. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Galluzzi L, Vitale I, Michels J, Brenner
C, Szabadkai G, Harel-Bellan A, Castedo M and Kroemer G: Systems
biology of cisplatin resistance: Past, present and future. Cell
Death Dis. 5:e12572014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kalluri R: The biology and function of
fibroblasts in cancer. Nat Rev Cancer. 16:582–598. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Chen WJ, Ho CC, Chang YL, Chen HY, Lin CA,
Ling TY, Yu SL, Yuan SS, Chen YJ, Lin CY, et al: Cancer-associated
fibroblasts regulate the plasticity of lung cancer stemness via
paracrine signalling. Nat Commun. 5:34722014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wintzell M, Hjerpe E, Åvall Lundqvist E
and Shoshan M: Protein markers of cancer-associated fibroblasts and
tumor-initiating cells reveal subpopulations in freshly isolated
ovarian cancer ascites. BMC Cancer. 12:3592012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Tao L, Huang G, Wang R, Pan Y, He Z, Chu
X, Song H and Chen L: Cancer-associated fibroblasts treated with
cisplatin facilitates chemoresistance of lung adenocarcinoma
through IL-11/IL-11R/STAT3 signaling pathway. Sci Rep. 6:384082016.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Shintani Y, Fujiwara A, Kimura T, Kawamura
T, Funaki S, Minami M and Okumura M: IL-6 secreted from
cancer-associated fibroblasts mediates chemoresistance in NSCLC by
increasing epithelial-mesenchymal transition signaling. J Thorac
Oncol. 11:1482–1492. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Lapteva N, Aldrich M, Weksberg D, Rollins
L, Goltsova T, Chen SY and Huang XF: Targeting the intratumoral
dendritic cells by the oncolytic adenoviral vaccine expressing
RANTES elicits potent antitumor immunity. J Immunother. 32:145–156.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Taub DD, Sayers TJ, Carter CR and Ortaldo
JR: Alpha and beta chemokines induce NK cell migration and enhance
NK-mediated cytolysis. J Immunol. 155:3877–3888. 1995.PubMed/NCBI
|
|
54
|
Böttcher JP, Bonavita E, Chakravarty P,
Blees H, Cabeza-Cabrerizo M, Sammicheli S, Rogers NC, Sahai E,
Zelenay S and Reis e Sousa C: NK cells stimulate recruitment of
cDC1 into the tumor microenvironment promoting cancer immune
control. Cell. 172:1022–1037.e14. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Simon PS, Bardhan K, Chen MR, Paschall AV,
Lu C, Bollag RJ, Kong FC, Jin J, Kong FM, Waller JL, et al: NF-κB
functions as a molecular link between tumor cells and Th1/Tc1 T
cells in the tumor microenvironment to exert radiation-mediated
tumor suppression. Oncotarget. 7:23395–23415. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Araujo JM, Gomez AC, Aguilar A, Salgado R,
Balko JM, Bravo L, Doimi F, Bretel D, Morante Z, Flores C, et al:
Effect of CCL5 expression in the recruitment of immune cells in
triple negative breast cancer. Sci Rep. 8:48992018. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Li W, Zhang X, Wang J, Li M, Cao C, Tan J,
Ma D and Gao Q: TGFβ1 in fibroblasts-derived exosomes promotes
epithelial-mesenchymal transition of ovarian cancer cells.
Oncotarget. 8:96035–96047. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Kim HJ, Lee DW, Yim GW, Nam EJ, Kim S, Kim
SW and Kim YT: Long non-coding RNA HOTAIR is associated with human
cervical cancer progression. Int J Oncol. 46:521–530. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Oh EJ, Kim SH, Yang WI, Ko YH and Yoon SO:
Long non-coding RNA HOTAIR expression in diffuse large B-cell
lymphoma: In relation to polycomb repressive complex pathway
proteins and H3K27 trimethylation. J Pathol Transl Med. 50:369–376.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yang X, Luo E, Liu X, Han B, Yu X and Peng
X: Delphinidin-3-glucoside suppresses breast carcinogenesis by
inactivating the Akt/HOTAIR signaling pathway. BMC Cancer.
16:4232016. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Ni W, Yao S, Zhou Y, Liu Y, Huang P, Zhou
A, Liu J, Che L and Li J: Long noncoding RNA GAS5 inhibits
progression of colorectal cancer by interacting with and triggering
YAP phosphorylation and degradation and is negatively regulated by
the m6A reader YTHDF3. Mol Cancer. 18:1432019.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Liu J, Tian W, Zhang W, Jia Y, Yang X,
Wang Y and Zhang J: MicroRNA-142-3p/MALAT1 inhibits lung cancer
progression through repressing β-catenin expression. Biomed
Pharmacother. 114:1088472019. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Lan T, Li H, Zhang D, Xu L, Liu H, Hao X,
Yan X, Liao H, Chen X, Xie K, et al: KIAA1429 contributes to liver
cancer progression through N6-methyladenosine-dependent
post-transcriptional modification of GATA3. Mol Cancer. 18:1862019.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Yang G, Lu X and Yuan L: LncRNA: A link
between RNA and cancer. Biochim Biophys Acta. 1839:1097–1109. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Gupta RA, Shah N, Wang KC, Kim J, Horlings
HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, et al: Long
non-coding RNA HOTAIR reprograms chromatin state to promote cancer
metastasis. Nature. 464:1071–1076. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Yao Y, Li J and Wang L: Large intervening
non-coding RNA HOTAIR is an indicator of poor prognosis and a
therapeutic target in human cancers. Int J Mol Sci. 15:18985–18999.
2014. View Article : Google Scholar : PubMed/NCBI
|