Introduction
Colorectal cancer (CRC) is one of the leading causes
of cancer-associated mortality in the world and currently ranks
third in terms of prevalence in the USA (1). The incidence of CRC in developing
countries is increasing at an alarming rate (1). Despite the significant progress made in
the development of treatment options for CRC, the overall survival
rate of patients with CRC has not improved sufficiently (2). Identifying novel biomarkers associated
with CRC may therefore help prolonging the survival time of
patients with CRC (3,4).
Transgelin (TAGLN) has been previously implicated in
numerous diseases, including hypertension, asthma and cancer
(5–7). First discovered in 1998, TAGLN2 is a
member of the calponin family that was found to be aberrantly
expressed in various diseases, especially in cancers (8). The relationship between TAGLN2 and the
pathophysiology of cancer has been extensively studied, and it is
believed that TAGLN2 is an oncogene and can regulate various
biological processes in cancer cells, including proliferation,
differentiation and apoptosis (9,10). A
previous study reported that TAGLN2 knockdown can effectively
reduce cell proliferation, migration and invasion in bladder cancer
cells (11). TAGLN2 increased
expression was also demonstrated to be associated with poor
survival in patients with glioma, suggesting a potential role of
TAGLN2 in the prognosis of glioma (12). A previous study has reported that
TAGLN2 is significantly upregulated in CRC tissues compared with
normal mucosa, and that its high expression is positively corrected
with the poor prognostic of patients and high metastasis (13). It has been proposed that epigenetic
repression of the miR-1-133a cluster may play a critical role in
CRC metastasis by silencing TAGLN2 (14). Furthermore, a recent study
demonstrated that TAGLN2 is mainly expressed by the tumor cells and
that its level is higher in these cellsin comparison with adjacent
healthy cells, such as colorectal, bladder, lung and hepatocellular
cancers, and cervical squamous cell carcinoma (15). However, the effects of TAGLN2 on the
proliferation, invasion, migration and epithelial-mesenchymal
transition (EMT) of CRC cells and its underlying mechanisms remain
to be elucidated.
Annexin A2 (ANXA2), a calcium-dependent
phospholipid-binding protein, is expressed mainly in endothelial
cells, macrophages and tumor cells and is widely distributed in the
nucleus, cytoplasm and extracellular surfaces (16). High expression of ANXA2 has been
shown to be associated with the progression of invasion,
metastasis, and angiogenesis in cancer (17). Notably, multiple studies have
elucidated that ANXA2 is expressed at higher levels in CRC than in
normal colorectal tissues and is associated with malignancy in CRC,
as evidenced by alterations associated with cell proliferation,
motility progression, recurrence and survival (18,19),
suggesting a significant and non-negligible role for ANXA2 in tumor
treatment and prognosis.
Signal transducer and activator of transcription 3
(STAT3) is one of the most frequently activated transcription
factors during inflammatory processes related to cancer (20). STAT3 knockdown in various cell types,
including tumor cells and macrophages, can effectively suppress
carcinogenesis and tumor growth (21–23). In
particular, mice with STAT3 deficiency exhibit an admirable ability
of increasing anti-tumor immune responses (22). It is widely accepted that higher
expression level of STAT3 is considered as a marker of poor
prognosis in patients with CRC (24). STAT3, which is overexpressed in CRC,
has been shown to play a prominent and pathogenic role in CRC
initiation, progression and metastasis (25). Targeting STAT3 may therefore be
considered as a potential therapeutic target for the treatment of
various types of cancer.
The present study aimed to explore the potential
role of TAGLN2 in CRC and to determine whether STAT3 could be
involved in the pathogenesis mechanism of CRC. This may help the
identification of novel targets for the treatment of CRC.
Materials and methods
Cell culture
The human colon epithelial cell line NCM460 and the
CRC cell lines HCT116, SNU-C1, LoVo and SW480 were purchased from
The Cell Bank of Type Culture Collection of The Chinese Academy of
Sciences. Cells were incubated in DMEM (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) and placed at 37°C in a humidified incubator
containing 5% CO2. HO-3867 (Selleck Chemicals; 10
µmol/l), which is a STAT3 inhibitor, was used to inhibit the
expression of STAT3.
Search Tool for the Retrieval of
Interacting Genes/Proteins (STRING) database
The STRING database 11.0 (https://www.string-db.org/) was used to predict the
binding sites between STAT3 and ANXA2.
Cell transfection
The short hairpin (sh)RNAs against TAGLN2
(shRNA-TAGLN2-1 and shRNA-TAGLN2-2, 100 nM), the shRNAs against
annexin 2 (ANXA2; shRNA-ANXA2-1 and shRNA-ANXA2-2, 100 nM), a
scrambled sequence used as the negative control (shRNA-NC, 100 nM)
for shRNA, the overexpression plasmid of TAGLN2 (OV-TAGLN2, 100 nM)
and the empty vector plasmid (OV-NC, 100 nM) were purchased from
Shanghai GenePharma Co., Ltd. SW480 cells were cultured in 6-well
plates and transfections were performed using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. Cells
incubated by shRNA were used as subsequent experiments after 24 h
at 37°C. Cells untreated were used as the control group.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from the NCM460, HCT116,
SNU-C1, LoVo and SW480 cells using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) and RNA was reverse
transcribed into cDNA using the RevertAid First Strand cDNA
Synthesis kit (Thermo Fisher Scientific, Inc.) according to the
manufacturers' protocol. RT-qPCR was performed with 2 µg cDNA using
iTaq™ Universal SYBR® Green Supermix (Bio-Rad
Laboratories, Inc.). The reactions were conducted in an ABI PRISM
7500 Real-time PCR system (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The thermocycling conditions were as follows:
Initial denaturation at 94°C for 10 min, followed by 45 cycles at
95°C for 15 sec and 60°C for 30 sec. The sequences of the primers
were as follows (Sangon Biotech Co., Ltd.): TAGLN2, forward
5′-CTACCTGAAGCCGGTGTCC-3′, reverse 5′-ATCCCCAGAGAAGAGCCCAT-3′;
N-cadherin (N-cad), forward 5′-TCAGGCTGTGGACATAGAAACC-3′, reverse
5′-GCTGTAAACGACTCTGGCACT−3′; vimentin, forward
5′-GACGCCATCAACACCGAGTT-3′, reverse 5′-CTTTGTCGTTGGTTAGCTGGT−3′;
zinc finger E-box binding homeobox 2 (ZEB2), forward
5′-GGAGACGAGTCCAGCTAGTGT−3′, reverse 5′-CCACTCCACCCTCCCTTATTTC-3′;
E-cadherin (E-cad), forward 5′-ATTTTTCCCTCGACACCCGAT−3′, reverse
5′-TCCCAGGCGTAGACCAAGA-3′; ANXA2, forward
5′-GTGAAGCGGGCTTGGGATT-3′, reverse 5′-CAAGGGCTGGAAAGCAGTC-3′; and
GAPDH, forward 5′-GAGTCAACGGATTTGGTCGT-3′ and reverse
5′-TTGATTTTGGAGGGATCTCG-3′.
The relative expression levels were normalized to
the endogenous control GAPDH and were expressed as
2−ΔΔCq (26).
Cell counting kit-8 (CCK-8) assay
Cell viability was evaluated using a CCK-8 assay
(Shanghai YiSheng Biotechnology Co., Ltd.). Briefly, SW480 cells
after transfection were plated into 96-well plates
(3×104 cells/well). Following cell attachment, 10 µl
CCK-8 reagent was added into the 96-well plates and cells were
incubated for 2 h at 37°C before cell viability was measured using
a spectrophotometer (Thermo Fisher Scientific, Inc.). The optical
density was measured at 450 nm.
5-ethynyl-2′-deoxyuridine (EdU)
assay
Cell proliferation ability was monitored by means of
EdU staining. SW480 cells were plated into 96-well plates
(5×104 cells/well) and incubated overnight at 37°C. The
next day, 10 µM EdU (Thermo Fisher Scientific, Inc.) was added into
the plates and incubated for 24 h at 37°C. Cells were then fixed
with 4% paraformaldehyde for 15 min at room temperature and
permeabilized with 0.3% Triton X-100 for 15 min at room
temperature. After rinsing with PBS three times and incubation with
4′, 6-diamidino-2-phenylindole (DAPI), a fluorescence microscope
(Olympus Corporation; magnification, ×200) was used to observe and
capture proliferative cells (EdU-positive) in three random fields
of view per slide and the cell counting was performed using ImageJ
software 1.52r (National Institutes of Health).
Wound healing assay
Wound healing assay was used to determine cell
migration. SW480 cells (3×105 cells/well) were seeded
into 6-well plates. Upon reaching 80% confluence, a 200 µl pipette
tip was used to draw a straight line on the cell monolayer. After
washing the cell debris with PBS three times, cells were cultured
with serum-free DMEM for 24 h at 37°C. Images of migratory cells
were captured at 0 and 24 h under a light microscope
(magnification, ×100; Olympus Corporation). The relative cell
migration rate of each group (24 h scratch distance-initial
distance) was normalized according to the average migrated distance
of the control group (ImageJ software 1.52r, National Institutes of
Health).
Cell invasion assay
To detect the cell invasive ability, Transwell
plates (Corning, Inc.) with 8-µm pore insert coated with Matrigel
(BD Biosciences) was used. SW480 cells (5×104
cells/well) were seeded into the upper chamber with serum-free
medium, whereas DMEM supplemented with 10% FBS was added to the
lower chamber. After incubation for 24 h at 37°C, invasive cells
were fixed with 4% formaldehyde for 15 min at 37°C and stained with
0.1% crystal violet for 20 min at 37°C. Cells were then visualized
and counted under a light microscope (magnification, ×100; Olympus
Corporation) from five randomly selected fields. The relative cell
invasive rate was calculated by average invasive cells in the
corresponding group normalized to the average invasive cells of the
control group (ImageJ software 1.52r, National Institutes of
Health).
Western blotting
Proteins were extracted from cells using RIPA lysis
buffer on ice (Beyotime Institute of Biotechnology). Protein
concentration was measured using a bicinchoninic acid protein assay
kit (Sigma-Aldrich; Merck KGaA). Proteins (40 µg) were separated by
10% SDS-PAGE and transferred onto PVDF membranes (EMD Millipore).
After blocking with 5% skimmed milk for 1 h at room temperature,
the membranes were incubated with primary antibodies against TAGLN2
(cat. no. 62567S; 1:1,000; Cell Signaling Technology, Inc.), matrix
metalloproteinase (MMP)2 (cat. no. 40994S; 1:1,000; Cell Signaling
Technology, Inc.), MMP9 (cat. no. 13667T; 1:1,000; Cell Signaling
Technology, Inc.), E-cad (cat. no. 3195T; 1:1,000; Cell Signaling
Technology, Inc.), N-cad (cat. no. 13116T; 1:1,000; Cell Signaling
Technology, Inc.), vimentin (cat. no. 5741T; 1:1,000; Cell
Signaling Technology, Inc.), phosphorylated-STAT3 (p-STAT3; cat.
no. 9145T; 1:1,000; Cell Signaling Technology, Inc.), STAT3 (cat.
no. 8768T; 1:1,000; Cell Signaling Technology, Inc.), ANXA2 (cat.
no. 8235S; 1:1,000; Cell Signaling Technology, Inc.), GAPDH (cat.
no. 5174T; 1:1,000; Cell Signaling Technology, Inc.) and ZEB2 (cat.
no. sc-271984; 1:1,000; Santa Cruz Biotechnology, Inc.) at 4°C
overnight. After rinsing with PBS, the membranes were incubated
with the appropriate HRP-conjugated secondary antibodies (cat. no.
7074S; 1:3,000; Cell Signaling Technology, Inc. and cat. no.
sc271984; 1:1,000; Santa Cruz Biotechnology, Inc.) at room
temperature for 60 min. The bands were visualized using enhanced
chemiluminescence reagent (EMD Millipore). Protein bands were
quantified using ImageJ software (version 1.52r; National
Institutes of Health). The relative protein level was normalized to
GAPDH.
Statistical analysis
The data were presented as the means ± standard
deviation of three independent experiments. Statistical analyses
were performed using GraphPad Prism version 6.0 (GraphPad Software,
Inc.). Comparisons between multiple groups were conducted using
ANOVA followed by Tukey's post hoc test. P<0.05 was considered
to indicate a statistically significant difference.
Results
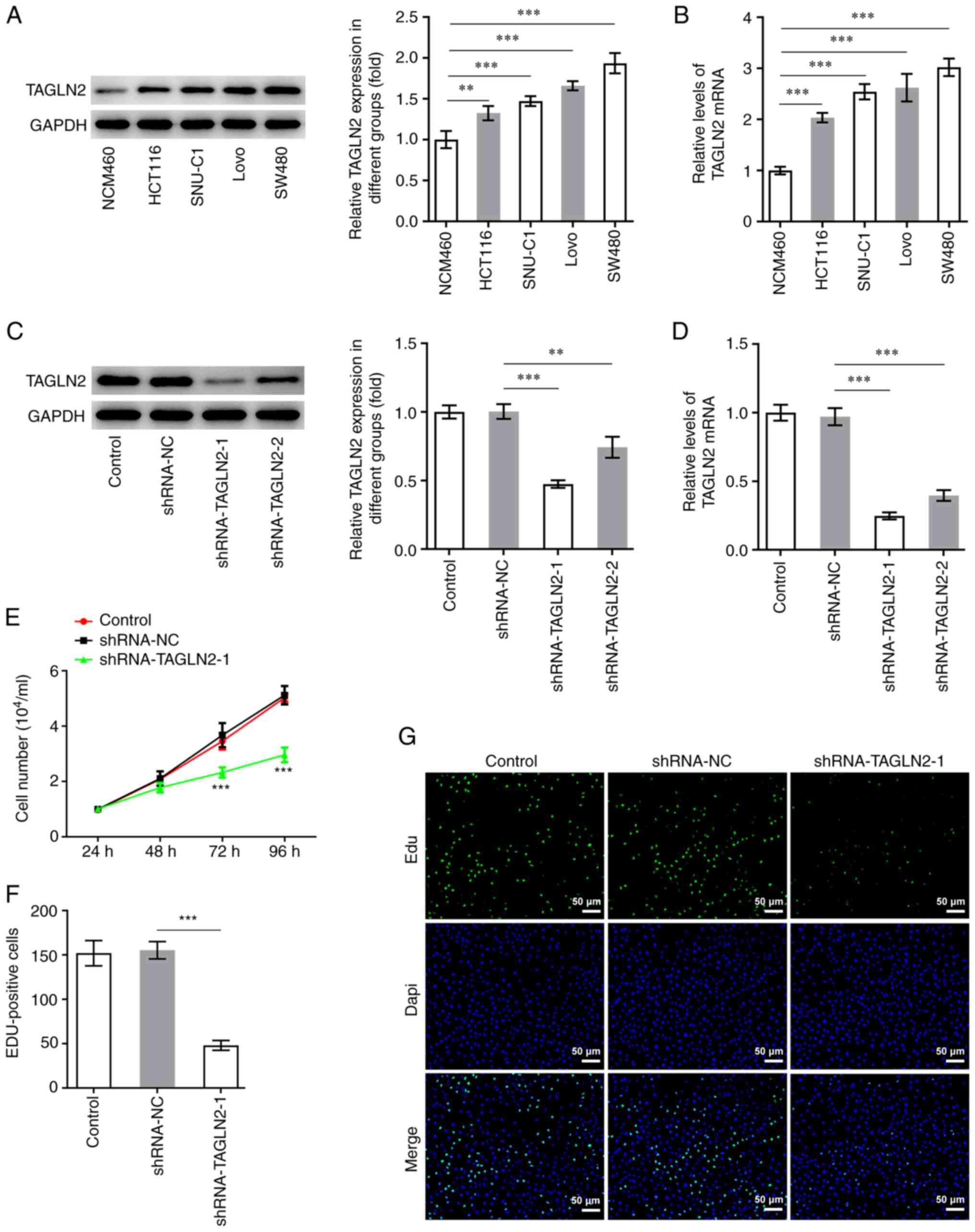
TAGLN2 expression is increased in CRC
cells
To study the specific role of TAGLN2 in CRC cells,
RT-qPCR and western blotting were conducted on CRC cell lines
(HCT116, SNU-C1, LoVo and SW480) and the NCM460 cell line. The
expression level of TAGLN2 was found to be the lowest in NCM460
cells, whereas TAGLN2 expression was significantly increased in all
CRC cells tested compared with NCM460 cells (Fig. 1A and B). Among all CRC cell lines
tested, TAGLN2 expression in SW480 cells was demonstrated to be the
highest. SW480 cell line was therefore chosen for subsequent
experiments.
TAGLN2 knockdown inhibits the
proliferation, migration, invasion, EMT and STAT3 phosphorylation
in CRC cells
To investigate the relationship between TAGLN2
expression and the physiology of CRC cells, TAGLN2 expression was
knocked down using shRNAs. The shRNA-TAGLN2-1, which significantly
decreased TAGLN2 expression, was used to knock down TAGLN2
expression in SW480 cells in subsequent experiments (Fig. 1C and D). An inhibitory effect of
TAGLN2 knockdown on the proliferation of SW480 cells compared with
the shRNA-NC group was observed in Fig.
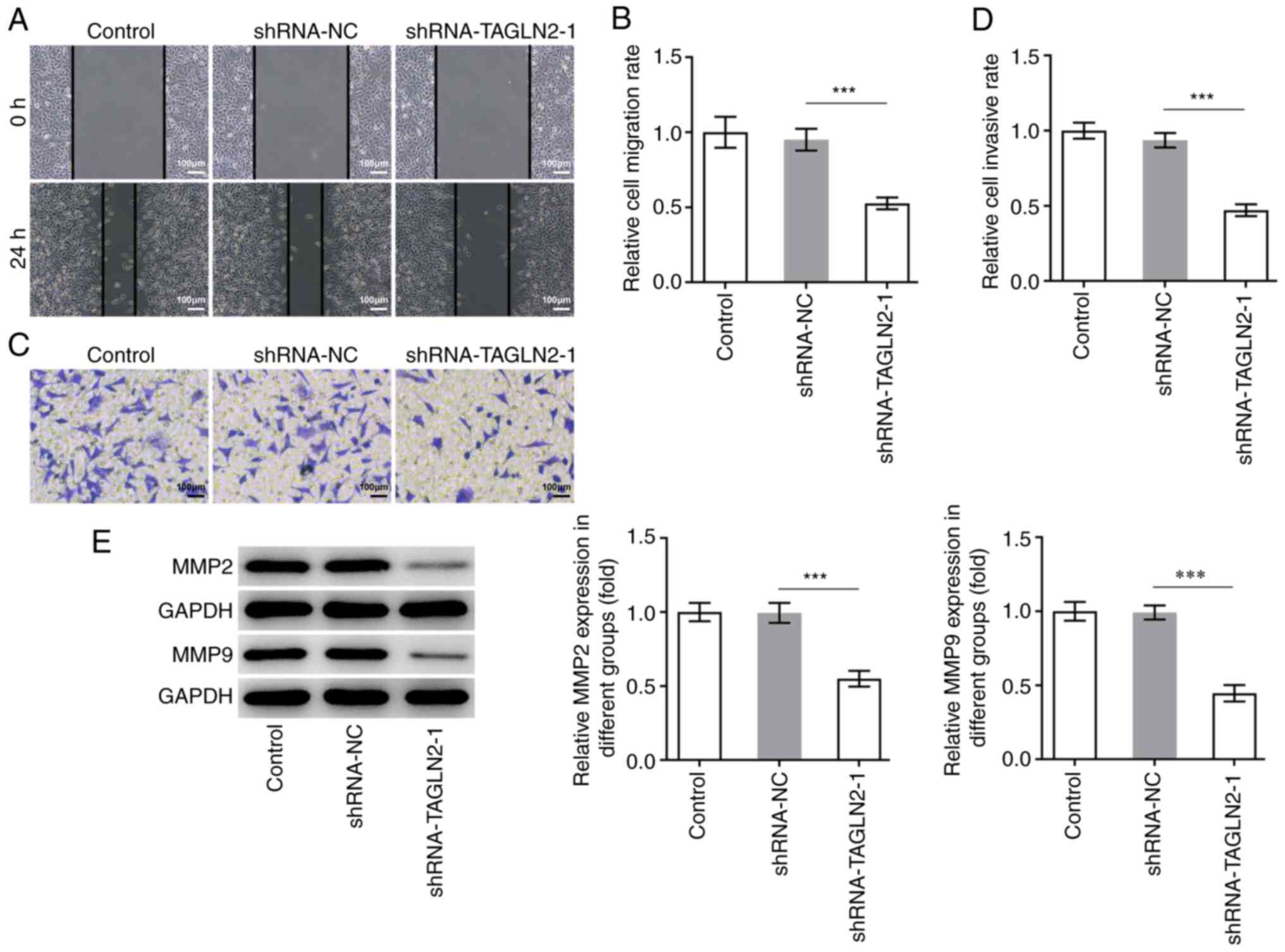
1E and F. In addition, TAGLN2 significantly decreased SW480
cell migration and invasion compared with the shRNA-NC group
(Fig. 2A-D). MMP2 and MMP9 are two
important migration-related proteins involved during tumorigenesis
(27,28). A significant decrease in MMP2 and
MMP9 expression was observed in the shRNA-TAGLN2-1 group compared
with the shRNA-NC group (Fig.
2E).
EMT is a process during which epithelial cells lose
their polarity and acquire a mesenchymal phenotype, which is an
important process for promoting tumor invasiveness and metastasis
(29). It is commonly characterized
by downregulation of E-cad, which is a key epithelial marker, along
with upregulation of N-cad, vimentin and ZEB2, which are crucial
mesenchymal marker genes (30,31). As
presented in Fig. 3A and B, the
protein expression of the mesenchymal markers N-cad, vimentin and
ZEB2 was significantly decreased, while E-cad expression was
significantly increased in SW480 cells transfected with
shRNA-TAGLN2-1 compared with the shRNA-NC group. Genes regulated by
canonical EMT have been reported to be downstream targets of STAT3.
Numerous cancer cell lines and primary tumors were reported to
possess constitutively active STAT3 (p-STAT3) that can lead to
cellular transformation and ultimately tumorigenesis (32). The expression of STAT3 and p-STAT3
was therefore determined in the present study. The results from
western blotting demonstrated that STAT3 phosphorylation was
significantly decreased in the shRNA-TAGLN2-1 group compared with
the shRNA-NC group (Fig. 3C). Taken
together, these findings suggested that TAGLN2 knockdown may
inhibit the proliferation, migration, invasion and EMT in CRC
cells, and decrease STAT3 phosphorylation.
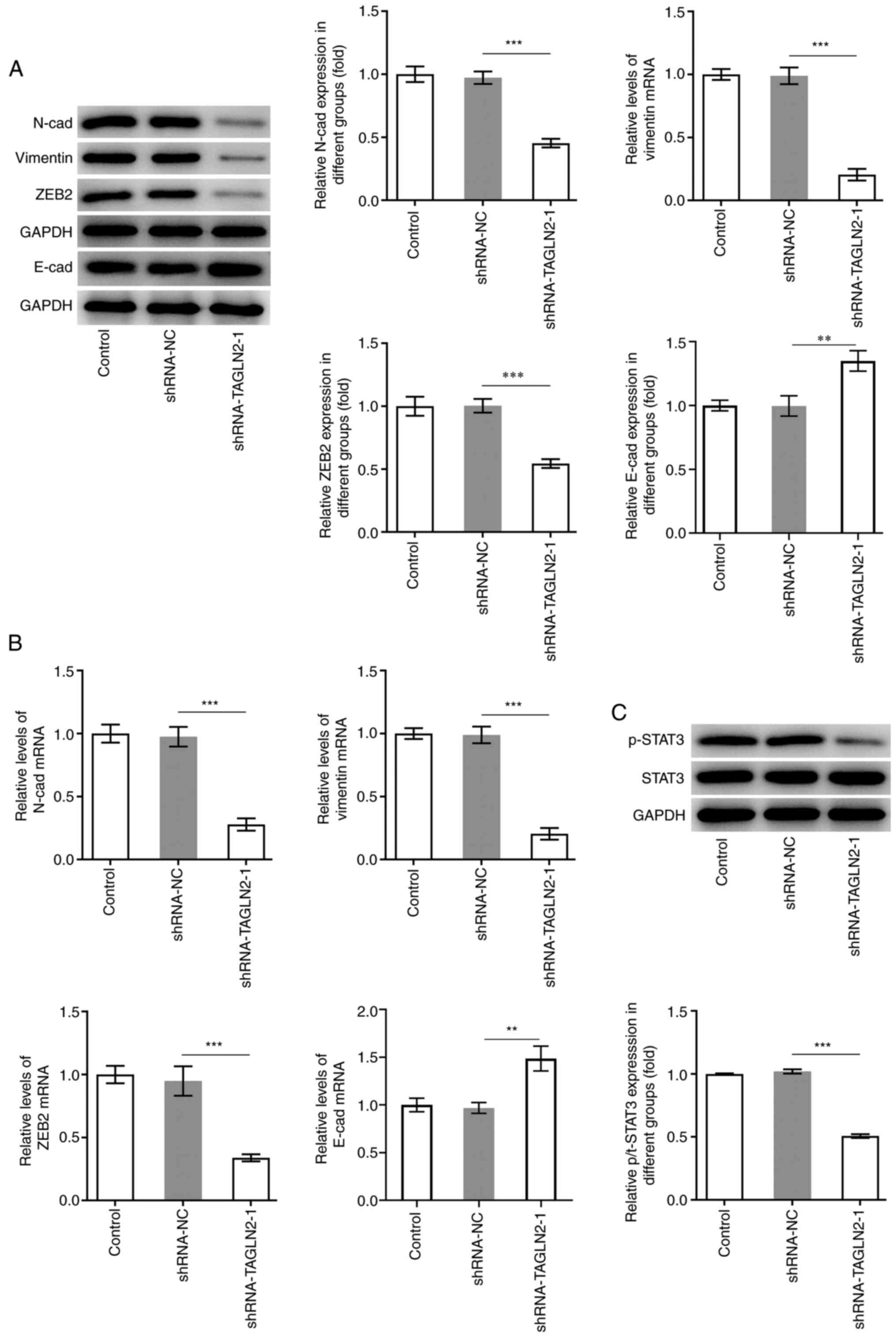 | Figure 3.Knockdown of transgelin 2 inhibits
EMT and STAT3 phosphorylation in colorectal cancer cells. (A and B)
Expression of proteins associated with EMT (N-cad, vimentin, ZEB2
and E-cad) was measured by reverse transcription-quantitative PCR
and western blotting. (C) Expression of STAT3 and p-STAT3 was
evaluated using western blotting. **P<0.01 and ***P<0.001.
EMT, epithelial-mesenchymal transition; STAT3, signal transducer
and activator of transcription 3; TAGLN2, transgelin 2; sh, short
hairpin; NC, negative control; N-cad, N-cadherin; E-cad,
E-cadherin; p, phosphorylated; ZEB2, zinc finger E-box binding
homeobox 2. |
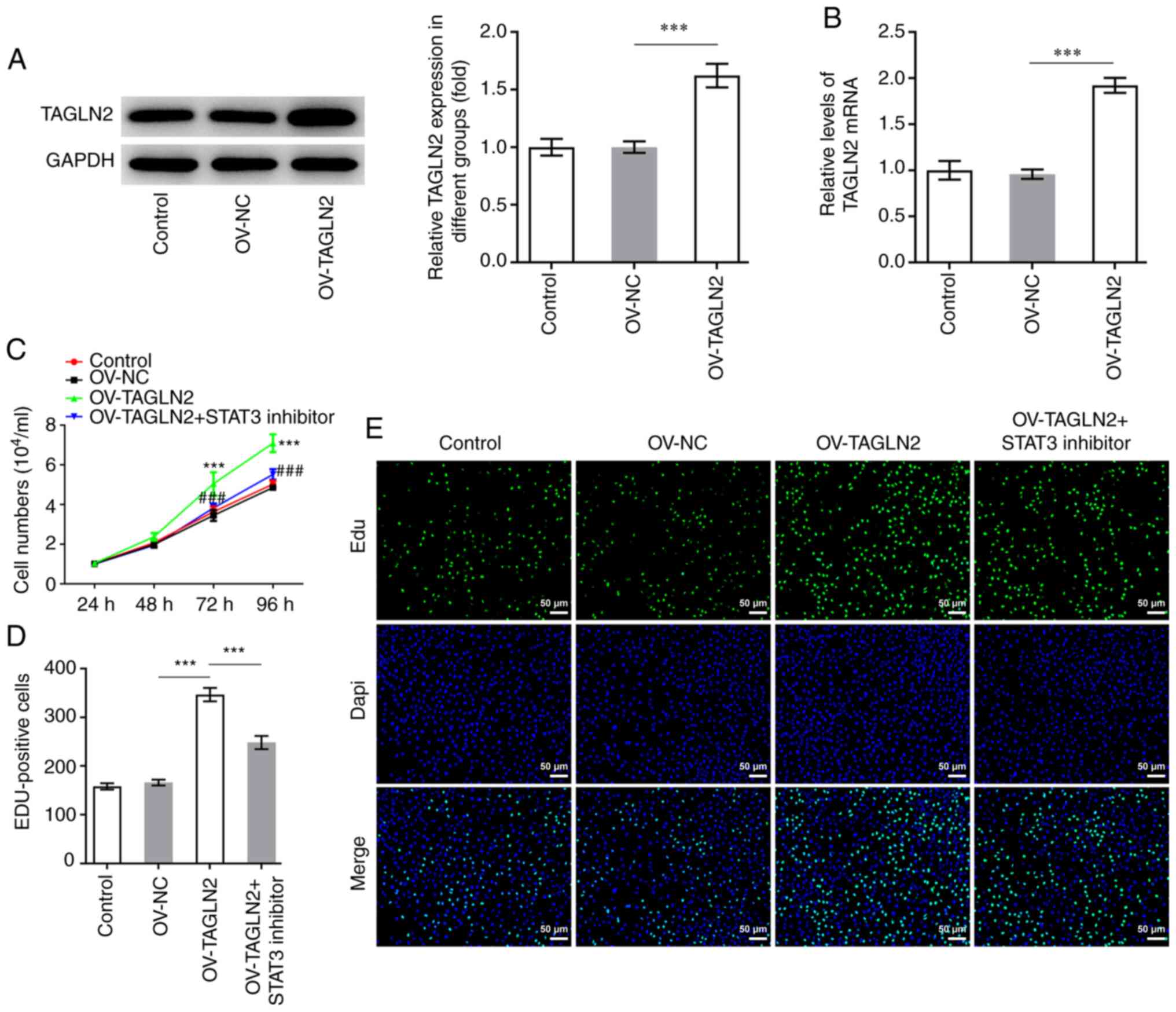
STAT3 inhibitor reverses the promoting
effects of TAGLN2 overexpression on the proliferation, migration,
invasion and EMT of CRC cells
To further investigate whether TAGLN2 could function
as an oncogenic gene by regulating STAT3 in CRC, TAGLN2 was
overexpressed. As presented in Fig. 4A
and B, the protein and mRNA expression of TAGLN2 was
significantly increased following transfection with TAGLN2 plasmid
compared with the OV-NC group. Subsequently, HO-3867 (10 µmol/l),
which is a STAT3 inhibitor, was used to inhibit the expression of
STAT3 in SW480 cells overexpressing TAGLN2. As presented in
Fig. 4C and D, overexpression of
TAGLN2 led to an enhanced proliferative ability of SW480 cells
according to CCK-8 and EdU staining compared with OV-NC group;
however, TAGLN2 overexpression combined with HO-3867 treatment
partially abolished the promoting effects of OV-TAGLN2.
Furthermore, TAGLN2 overexpression significantly increased the
migration and invasion of SW480 cells compared with the OV-NC
group, both of which were partially counteracted by treatment with
HO-3867 (Fig. 5A-D). The expression
profiles of MMP2 and MMP9 in the combined OV-TAGLN2 + HO-3867
treatment group exhibited similar trends as those observed for the
migration and invasion following OV-TAGLN2 transfection (Fig. 5E). Subsequently, significantly
upregulated N-cad, vimentin and ZEB2 expression coupled with
downregulated expression of E-cad were demonstrated following
TAGLN2 overexpression in SW480 cells compared with OV-NC group,
which were partially reversed by HO-3867 treatment (Fig. 5F and G). Taken together, these
findings suggested that TAGLN2 overexpression may promote the
proliferation, migration, invasion and EMT of SW480 cells at least
in part through STAT3 signaling.
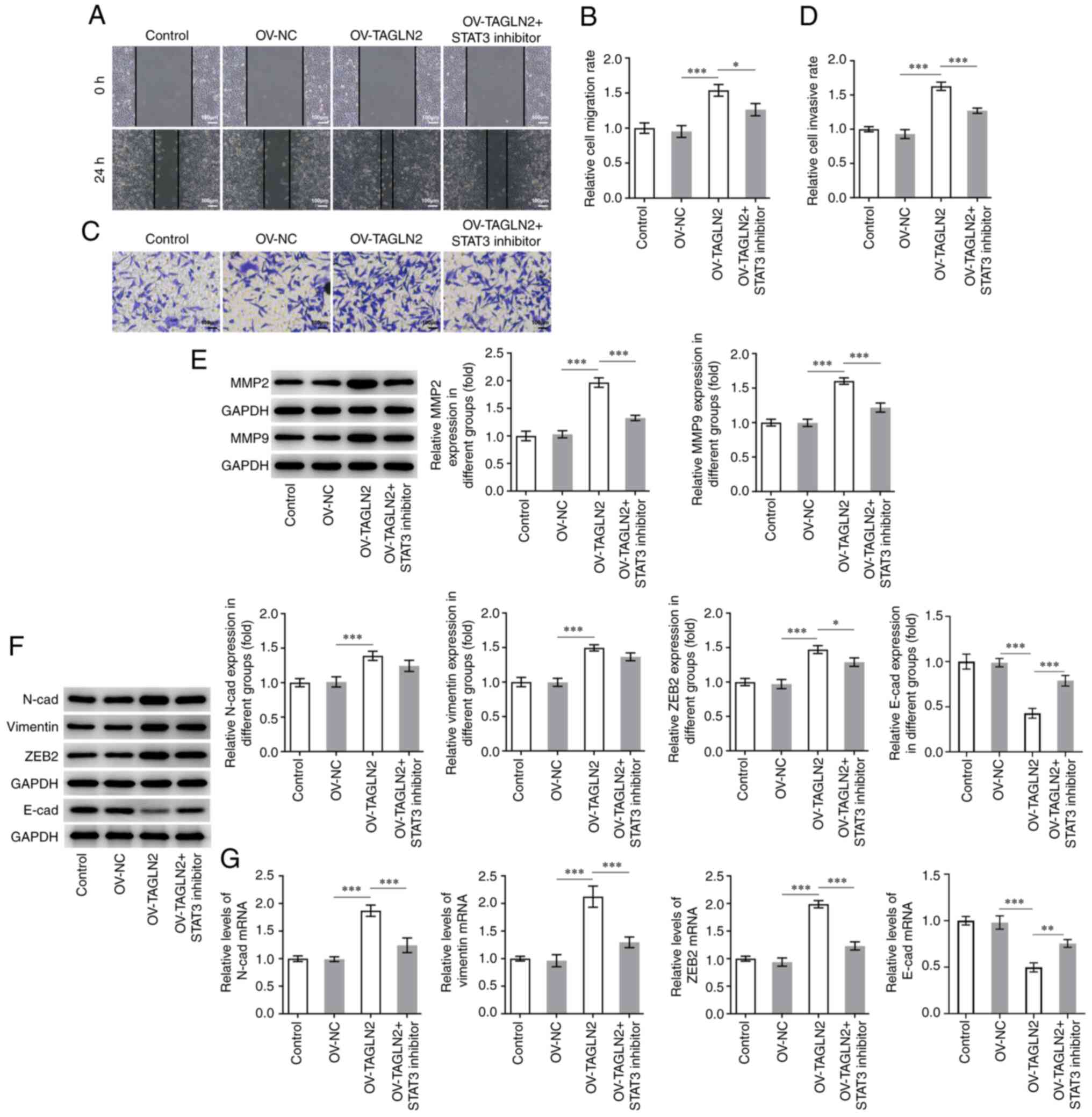 | Figure 5.STAT3 inhibitor HO-3867 reverses the
positive effects of TAGLN2 overexpression on the migration,
invasion and EMT of colorectal cancer cells. SW480 cell migration
and invasion were measured by (A and B) wound healing and (C and D)
Transwell assays, respectively (scale bar, 100 µm). (E) Expression
of MMP2 and MMP9 was measured by western blotting. (F and G)
Expression of proteins associated with EMT (N-cad, vimentin, ZEB2
and E-cad) was measured by reverse transcription-quantitative PCR
and western blotting. *P<0.05, **P<0.01 and ***P<0.001.
EMT, epithelial-mesenchymal transition; MMP, matrix
metalloproteinase; TAGLN2, transgelin 2; OV, overexpressing
plasmid; NC, negative control; STAT3, signal transducer and
activator of transcription 3; N-cad, N-cadherin; E-cad, E-cadherin;
ZEB2, zinc finger E-box binding homeobox 2. |
TAGLN2 overexpression promotes the
expression of STAT3 via ANXA2
It was previously reported that TAGLN2 is involved
in CRC progression by regulating ANXA2 (33). Further analysis using STRING database
predicted that ANXA2 may bind to STAT3 (Fig. 6A). The expression of ANXA2 was
therefore evaluated in SW480 cells and was found to be
significantly higher in SW480 cells overexpressing TAGLN2 compared
with SW480 cells transfected with OV-NC, suggesting the regulating
effect of TAGLN2 to ANXA2 (Fig. 6B and
C). ANXA2 expression was subsequently knocked down by
transfection with shRNA-ANXA2-1 and shRNA-ANXA2-2. Since
shRNA-ANXA2-1 exhibited a superior transfection efficiency,
shRNA-ANXA2-1 was chosen for following experiments (Fig. 6D and E). Overexpression of TAGLN2 led
to an increase in the levels of STAT3 phosphorylation, which was
reversed following shRNA-ANXA2 transfection (Fig. 6F). These findings suggested that
TAGLN2 overexpression may activate STAT3 signaling potentially via
ANXA2.
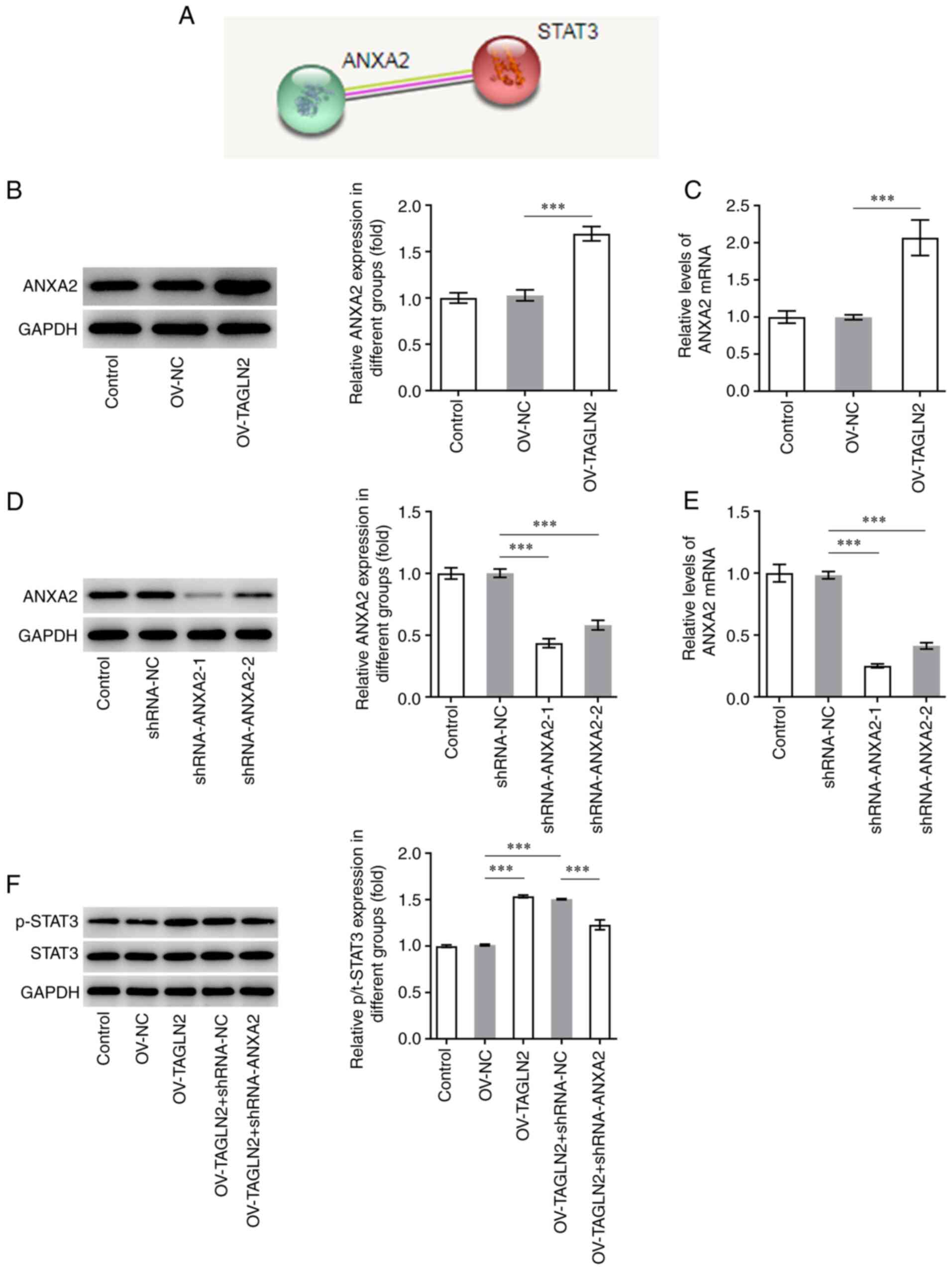 | Figure 6.Overexpression of TAGLN2 enhances the
expression of STAT3 downstream of ANXA2. (A) ANXA2 was predicted to
be associated with STAT3 according to the Search Tool for the
Retrieval of Interacting Genes/Proteins database. After
overexpressing TAGLN2, the expression of ANXA2 was measured using
(B) RT-qPCR and (C) western blotting. After knocking down ANXA2 in
TAGLN2-overexpressing cells, the expression of ANXA2 was detected
by (D) RT-qPCR and (E) western blotting. (F) Western blotting was
used to examine the expression of STAT3 and p-STAT3. ***P<0.001.
RT-qPCR, reverse transcription-quantitative PCR; TAGLN2, transgelin
2; ANXA2, annexin 2; STAT3, signal transducer and activator of
transcription 3; sh, short hairpin; NC, negative control; OV,
overexpressing plasmid; p, phosphorylated. |
Discussion
The calponin homolog domain contained within the
TAGLN2 protein structure implicates the possible involvement of
TAGLN2 in cytoskeletal reorganization, and TAGLN2 was shown to be
directly and indirectly involved in numerous cancer-related
processes, such as migration, proliferation, differentiation or
apoptosis (19,34). Previous studies reported a
relationship between TAGLN2 expression and cancer (33,35).
Higher expression levels of TAGLN2 were previously observed in
pancreatic cells (36). TAGLN2
expression was also found to be increased in CRC tissues, which was
associated with lymph node and distant metastases, suggesting a
potential role of TAGLN2 in the pathophysiology of CRC (35). However, the effects of TAGLN2 on the
proliferation, invasion, migration and EMT of CRC cells and its
potential mechanisms remain to be elucidated. The present study
demonstrated that TAGLN2 expression was significantly elevated in
CRC cells and that knocking down TAGLN2 expression significantly
inhibited CRC cell proliferation, migration, invasion and EMT. The
effects of TAGLN2 on the progression of CRC may be partly mediated
by STAT3, which is located downstream of the ANXA2 signaling
pathway.
A previous study reported that TAGLN2 is
significantly upregulated in CRC tissues compared with normal
mucosa, and that its high expression is positively corrected with
the poor prognostic of these patients and high metastasis (17). To the best of our knowledge, the
present study was the first to investigate the effects of TAGLN2 on
the progression of CRC. The results demonstrated that TAGLN2 was
significantly elevated in CRC cells and that TAGLN2 silencing
inhibited the proliferation, migration, invasion and EMT of CRC
cells. The indirect inactivation of STAT3 signaling using tyrosine
kinase inhibitors has already been applied clinically and was shown
to regulate cell cycle and apoptosis in tumor cells, which in turn
can control the progression of numerous types of cancer, including
medulloblastomas and renal cell carcinoma (37,38). A
previous study using an in vivo mice model has also
demonstrated that STAT3 inhibitors can serve as potent tumor
suppressors due to their anti-tumor activity (22). Another study also reported that STAT3
activation is a marker of poor prognosis in patients with CRC
(24). STAT3 overexpression has been
shown to play a crucial and pathogenic role in CRC initiation,
progression and metastasis (25).
Targeting STAT3 may therefore be promising for future CRC
therapies. EMT is a process that frequently occurs during fibrosis
and development of cancers (39).
STAT3 has been widely reported to be a transcriptional activator of
genes associated with EMT in gastrointestinal cancer (40). To verify whether TAGLN2 could
regulate STAT3 expression and subsequent CRC cell behavior, TAGLN2
was overexpressed in SW480 cells in the present study. The
proliferation, migration, invasion and EMT were all enhanced
following TAGLN2 overexpression. In addition, following treatment
with the STAT3 inhibitor HO-3867, the potentiated proliferation,
migration, invasion and EMT previously induced by TAGLN2
overexpression was partially reversed in SW480 cells. A previous
study similarly reported the preventive effect of STAT3 inhibition
on against the cellular alterations because of the EMT (41).
The findings from the present study suggested that
TAGLN2 may promote pathological cellular behaviors of CRC,
including proliferation and invasion, through STAT3. However, the
underlying mechanisms by which TAGLN2 could regulate STAT3 remain
to be fully elucidated. A previous study reported the involvement
of TAGLN2 in the metastasis and progression of hepatocellular
carcinoma via ANXA2 (33). ANXA2 is
a type of calcium-dependent phospholipid-binding proteins that is
highly expressed in various types of cancer cells, including breast
cancer, gastric cancer, lung cancer and CRC cells (42–46). It
has also been reported that the histological type, tumor size and
Tumor-Node-Metastasis stage are associated with the high expression
levels of ANXA2 (43). In addition,
the recurrence and survival rates of patients with CRC are closely
related to ANXA2 expression (47).
ANXA2 is mainly localized in the plasma membranes of cells in CRC
tissues and adjacent non-cancerous tissues, although its expression
is significantly higher in CRC tissues compared with adjacent
non-cancerous tissues (48). ANXA2
expression may therefore be used in the clinical diagnosis of
CRC.
ANXA2 has been demonstrated to activate STAT3 in
macrophages and to enhance the formation of breast cancer,
pancreatic cancer and hepatoma (49). ANXA2 overexpression serves a crucial
role in CRC invasiveness through Src/ANXA2/STAT3 pathway activation
(41). ANXA2 might regulate the
proliferation, invasion and migration of CRC cells in association
with STAT3 (50). Compared with the
aforementioned previous studies, the present study hypothesized
that STAT3 may serve an important role in the downstream signaling
pathway of TAGLN2 and ANXA2 during CRC pathogenesis. Although the
relationship between STAT3 and ANXA2 in CRC has been already
reported, the present study may be the first to report the
relationship between TAGLN2, STAT3 and ANXA2 in CRC. In the present
study, a positive relationship between TAGLN2 and ANXA2 expression
was found, which was supported by the fact that TAGLN2
overexpression also increased ANXA2 expression. Furthermore, ANXA2
knockdown led to decreased STAT3 phosphorylation in
TAGLN2-overexpressing cells, suggesting that TAGLN2 overexpression
could activate STAT3 downstream of ANXA2.
This study presented some limitations. Firstly, only
one CRC cell line was used to investigate the effects of TAGLN2 in
CRC and its regulatory effects on ANXA2/STAT3. Secondly, the
effects of TAGLN2/ANXA2/STAT3 axis on tumor growth and metastasis
of mice bearing tumor was not investigated.
In summary, the present study demonstrated for the
first time that TAGLN2 overexpression could promote CRC cell
proliferation, migration and invasion, in addition to facilitating
EMT by activating STAT3 downstream of ANXA2, which may be the
underlying mechanism by which TAGLN2 regulates the progression of
CRC. This potentially provides a therapeutic target for CRC, as
inhibition or knockdown of TAGLN2 reduces CRC cell tumor formation
by suppressing STAT3 signaling downstream of ANXA2.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Science and
Technology Fund of Tianjin Health Bureau (grant no. 2014KZ123).
Availability of data and materials
The analyzed data sets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZZ and LL were responsible for conceiving and
designing the study. LL and WL contributed to perform the
experiment and collect the data and review the manuscript. ZZ and
LL were responsible for analyzing the data and confirming the
authenticity of all the raw data. All authors read and approved the
final manuscript and confirm the authenticity of all the raw
data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Thanikachalam K and Khan G: Colorectal
cancer and nutrition. Nutrients. 11:1642019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Carrato A: Adjuvant treatment of
colorectal cancer. Gastrointest Cancer Res. 2 (Suppl 4):S42–S46.
2008.PubMed/NCBI
|
|
3
|
Tauriello DV, Calon A, Lonardo E and
Batlle E: Determinants of metastatic competency in colorectal
cancer. Mol Oncol. 11:97–119. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hafizi M, Kalanaky S, Moaiery H,
Khayamzadeh M, Noorian S, Kaveh V, Gharib B, Foudazi H, Razavi M,
Jenabian A, et al: A randomized, double-blind, placebo-controlled
investigation of BCc1 nanomedicine effect on survival and quality
of life in metastatic and non-metastatic gastric cancer patients. J
Nanobiotechnology. 17:522019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Huang L, Li L, Yang T, Li W, Song L, Meng
X, Gu Q, Xiong C and He J: Transgelin as a potential target in the
reversibility of pulmonary arterial hypertension secondary to
congenital heart disease. J Cell Mol Med. 22:6249–6261. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sun Y, Peng W, He W, Luo M, Chang G, Shen
J, Zhao X and Hu Y: Transgelin-2 is a novel target of KRAS-ERK
signaling involved in the development of pancreatic cancer. J Exp
Clin Cancer Res. 37:1662018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yin LM, Xu YD, Peng LL, Duan TT, Liu JY,
Xu Z, Wang WQ, Guan N, Han XJ, Li HY, et al: Transgelin-2 as a
therapeutic target for asthmatic pulmonary resistance. Sci Transl
Med. 10:eaam86042018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li M, Li S, Lou Z, Liao X, Zhao X, Meng Z,
Bartlam M and Rao Z: Crystal structure of human transgelin. J
Struct Biol. 162:229–236. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nohata N, Sone Y, Hanazawa T, Fuse M,
Kikkawa N, Yoshino H, Chiyomaru T, Kawakami K, Enokida H, Nakagawa
M, et al: miR-1 as a tumor suppressive microRNA targeting TAGLN2 in
head and neck squamous cell carcinoma. Oncotarget. 2:29–42. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Du YY, Zhao LM, Chen L, Sang MX, Li J, Ma
M and Liu JF: The tumor-suppressive function of miR-1 by targeting
LASP1 and TAGLN2 in esophageal squamous cell carcinoma. J
Gastroenterol Hepatol. 31:384–393. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yoshino H, Chiyomaru T, Enokida H,
Kawakami K, Tatarano S, Nishiyama K, Nohata N, Seki N and Nakagawa
M: The tumour-suppressive function of miR-1 and miR-133a targeting
TAGLN2 in bladder cancer. Br J Cancer. 104:808–818. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Han MZ, Xu R, Xu YY, Zhang X, Ni SL, Huang
B, Chen AJ, Wei YZ, Wang S, Li WJ, et al: TAGLN2 is a candidate
prognostic biomarker promoting tumorigenesis in human gliomas. J
Exp Clin Cancer Res. 36:1552017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mo X, Su Z, Yang B, Zeng Z, Lei S and Qiao
H: Identification of key genes involved in the development and
progression of early-onset colorectal cancer by co-expression
network analysis. Oncol Lett. 19:177–186. 2020.PubMed/NCBI
|
|
14
|
Chen WS, Leung CM, Pan HW, Hu LY, Li SC,
Ho MR and Tsai KW: Silencing of miR-1-1 and miR-133a-2 cluster
expression by DNA hypermethylation in colorectal cancer. Oncol Rep.
28:1069–1076. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dvorakova M, Nenutil R and Bouchal P:
Transgelins, cytoskeletal proteins implicated in different aspects
of cancer development. Expert Rev Proteomics. 11:149–165. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Deng S, Wang J, Hou L, Li J, Chen G, Jing
B, Zhang X and Yang Z: Annexin A1, A2, A4 and A5 play important
roles in breast cancer, pancreatic cancer and laryngeal carcinoma,
alone and/or synergistically. Oncol Lett. 5:107–112. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu XH, Pan W, Kang LH, Feng H and Song YQ:
Association of annexin A2 with cancer development (review). Oncol
Rep. 33:2121–2128. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang T, Peng H, Wang J, Yang J, Nice EC,
Xie K and Huang C: Prognostic and diagnostic significance of
annexin A2 in colorectal cancer. Colorectal Dis. 15:e373–e381.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
He H, Xiao L, Cheng S, Yang Q, Li J, Hou
Y, Song F, Su X, Jin H, Liu Z, et al: Annexin A2 enhances the
progression of colorectal cancer and hepatocarcinoma via
cytoskeleton structural rearrangements. Microsc Microanal.
25:950–960. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yu H, Kortylewski M and Pardoll D:
Crosstalk between cancer and immune cells: Role of STAT3 in the
tumour microenvironment. Nat Rev Immunol. 7:41–51. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chiarle R, Simmons WJ, Cai H, Dhall G,
Zamo A, Raz R, Karras JG, Levy DE and Inghirami G: Stat3 is
required for ALK-mediated lymphomagenesis and provides a possible
therapeutic target. Nat Med. 11:623–629. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kortylewski M, Kujawski M, Wang T, Wei S,
Zhang S, Pilon-Thomas S, Niu G, Kay H, Mulé J, Kerr WG, et al:
Inhibiting Stat3 signaling in the hematopoietic system elicits
multicomponent antitumor immunity. Nat Med. 11:1314–1321. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chan KS, Sano S, Kiguchi K, Anders J,
Komazawa N, Takeda J and DiGiovanni J: Disruption of Stat3 reveals
a critical role in both the initiation and the promotion stages of
epithelial carcinogenesis. J Clin Invest. 114:720–728. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kusaba T, Nakayama T, Yamazumi K, Yakata
Y, Yoshizaki A, Inoue K, Nagayasu T and Sekine I: Activation of
STAT3 is a marker of poor prognosis in human colorectal cancer.
Oncol Rep. 15:1445–1451. 2006.PubMed/NCBI
|
|
25
|
Chalikonda G, Lee H, Sheik A and Huh YS:
Targeting key transcriptional factor STAT3 in colorectal cancer.
Mol Cell Biochem. Apr 18–2021.doi: 10.1007/s11010-021-04156-8.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Su X, Gao C, Feng X and Jiang M: miR-613
suppresses migration and invasion in esophageal squamous cell
carcinoma via the targeting of G6PD. Exp Ther Med. 19:3081–3089.
2020.PubMed/NCBI
|
|
28
|
Wang W, Cheng X and Zhu J: Long non-coding
RNA OTUD6B-AS1 overexpression inhibits the proliferation, invasion
and migration of colorectal cancer cells via downregulation of
microRNA-3171. Oncol Lett. 21:1932021. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kaufhold S and Bonavida B: Central role of
Snail1 in the regulation of EMT and resistance in cancer: A target
for therapeutic intervention. J Exp Clin Cancer Res. 33:622014.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sreekumar R, Al-Saihati H, Emaduddin M,
Moutasim K, Mellone M, Patel A, Kilic S, Cetin M, Erdemir S, Navio
MS, et al: The ZEB2-dependent EMT transcriptional programme drives
therapy resistance by activating nucleotide excision repair genes
ERCC1 and ERCC4 in colorectal cancer. Mol Oncol. 8:2065–2083. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Feng J, Cen J, Li J, Zhao R, Zhu C, Wang
Z, Xie J and Tang W: Histone deacetylase inhibitor valproic acid
(VPA) promotes the epithelial mesenchymal transition of colorectal
cancer cells via up regulation of Snail. Celll Adhes Migr.
9:495–501. 2015. View Article : Google Scholar
|
|
32
|
Zhao C, Li H, Lin HJ, Yang S, Lin J and
Liang G: Feedback activation of STAT3 as a cancer drug-resistance
mechanism. Trends Pharmacol Sci. 37:47–61. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shi J, Ren M, She X, Zhang Z, Zhao Y, Han
Y, Lu D and Lyu L: Transgelin-2 contributes to proliferation and
progression of hepatocellular carcinoma via regulating Annexin A2.
Biochem Biophys Res Commun. 523:632–638. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Marchler-Bauer A, Anderson JB, Derbyshire
MK, DeWeese-Scott C, Gonzales NR, Gwadz M, Hao L, He S, Hurwitz DI,
Jackson JD, et al: CDD: A conserved domain database for interactive
domain family analysis. Nucleic Acids Res. 35((Database Issue)):
D237–D240. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang Y, Ye Y, Shen D, Jiang K, Zhang H,
Sun W, Zhang J, Xu F, Cui Z and Wang S: Identification of
transgelin-2 as a biomarker of colorectal cancer by laser capture
microdissection and quantitative proteome analysis. Cancer Sci.
101:523–529. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen R, Yi EC, Donohoe S, Pan S, Eng J,
Cooke K, Crispin DA, Lane Z, Goodlett DR, Bronner MP, et al:
Pancreatic cancer proteome: The proteins that underlie invasion,
metastasis, and immunologic escape. Gastroenterology.
129:1187–1197. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yang F, Van Meter TE, Buettner R, Hedvat
M, Liang W, Kowolik CM, Mepani N, Mirosevich J, Nam S, Chen MY, et
al: Sorafenib inhibits signal transducer and activator of
transcription 3 signaling associated with growth arrest and
apoptosis of medulloblastomas. Mol Cancer Ther. 7:3519–3526. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Xin H, Zhang C, Herrmann A, Du Y, Figlin R
and Yu H: Sunitinib inhibition of Stat3 induces renal cell
carcinoma tumor cell apoptosis and reduces immunosuppressive cells.
Cancer Res. 69:2506–2513. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li B and Huang C: Regulation of EMT by
STAT3 in gastrointestinal cancer (review). Int J Oncol. 50:753–767.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Rocha MR, Barcellos-de-Souza P,
Sousa-Squiavinato ACM, Fernandes PV, de Oliveira IM, Boroni M and
Morgado-Diaz JA: Annexin A2 overexpression associates with
colorectal cancer invasiveness and TGF-β induced epithelial
mesenchymal transition via Src/ANXA2/STAT3. Sci Rep. 8:112852018.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Gerke V: Annexins A2 and A8 in endothelial
cell exocytosis and the control of vascular homeostasis. Biol Chem.
397:995–1003. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Emoto K, Yamada Y, Sawada H, Fujimoto H,
Ueno M, Takayama T, Kamada K, Naito A, Hirao S and Nakajima Y:
Annexin II overexpression correlates with stromal tenascin-C
overexpression: A prognostic marker in colorectal carcinoma.
Cancer. 92:1419–1426. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sharma MR, Koltowski L, Ownbey RT,
Tuszynski GP and Sharma MC: Angiogenesis-associated protein annexin
II in breast cancer: Selective expression in invasive breast cancer
and contribution to tumor invasion and progression. Exp Mol Pathol.
81:146–156. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Emoto K, Sawada H, Yamada Y, Fujimoto H,
Takahama Y, Ueno M, Takayama T, Uchida H, Kamada K, Naito A, et al:
Annexin II overexpression is correlated with poor prognosis in
human gastric carcinoma. Anticancer Res. 21:1339–1345.
2001.PubMed/NCBI
|
|
46
|
Jia JW, Li KL, Wu JX and Guo SL: Clinical
significance of annexin II expression in human non-small cell lung
cancer. Tumour Biol. 34:1767–1771. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wang CY and Lin CF: Annexin A2: Its
molecular regulation and cellular expression in cancer development.
Dis Markers. 2014:3089762014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhai H, Acharya S, Gravanis I, Mehmood S,
Seidman RJ, Shroyer KR, Hajjar KA and Tsirka SE: Annexin A2
promotes glioma cell invasion and tumor progression. J Neurosci.
31:14346–14360. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Maji S, Chaudhary P, Akopova I, Nguyen PM,
Hare RJ, Gryczynski I and Vishwanatha JK: Exosomal annexin II
promotes angiogenesis and breast cancer metastasis. Mol Cancer Res.
15:93–105. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Xiu D, Liu L, Qiao F, Yang H, Cui L and
Liu G: Annexin A2 coordinates STAT3 to regulate the invasion and
migration of colorectal cancer cells in vitro. Gastroenterol Res
Pract. 2016:35214532016. View Article : Google Scholar : PubMed/NCBI
|