Introduction
Acute myelogenous leukemia (AML) is the most common
type of acute leukemia, accounting for ~80% of all cases worldwide
(1). AML develops from the bone
marrow and blood, and is characterized by rapid progression and a
highly aggressive nature (2).
Although AML is a severe malignancy, it is treatable and often
curable with chemotherapy in combination with targeted drugs
(3,4). However, chemoresistance frequently
occurs after long-term therapy, resulting in chemotherapeutic
failure and poor patient survival times (5,6).
Therefore, the development of novel therapeutic approaches to
further improve patient survival is urgently required.
Molecular-targeted therapies are emerging novel
approaches for AML treatment, which regulate the expression of
related genes (7–9). With an increasing understanding of the
molecular mechanisms of AML, certain molecular pathways, such as
PI3K/Akt/mTOR signaling and glutathione metabolism pathways, have
been indicated as potential targets for anti-AML therapy (10,11).
However, molecular-targeted therapy is still in the research
stages. Assessment of therapeutic safety, and the identification of
more effective drug targets, are being widely researched. As
covalently closed, single-strand RNA transcripts, circular RNAs
(circRNAs) are involved in various cancer types by regulating the
expression of cancer-related genes (12). Noncoding RNAs have been regarded as
novel regulators of cancer progression (13). microRNAs (miRNAs/miRs) and circRNAs
are two important types of noncoding RNA. MiR-34b is one of the
hallmark miRNAs that are associated with AML chemotherapy
resistance (14). Furthermore,
miR-34b regulates ubiquitin-specific protease 2a expression to
increase intracellular glutathione content and indirectly interfere
with the oxidative cascade, triggered by chemotherapeutic agents
(15). Circ-ATAD1 is a newly
identified circRNA contributing to gastric cancer cell progression
(16). However, to the best of our
knowledge, its role in other cancer types is yet to be reported.
Therefore, the present study was conducted to investigate the role
of circ-ATAD1 and miR-34b in AML.
Materials and methods
Research subjects
A total of 60 patients with AML (38 men and 22
women; age, 62.2+/-5.7 years) and 60 healthy controls (38 men and
22 women; age, 62.1+/-5.8 years) who were admitted to the Xing'an
League People's Hospital (Ulanhot, China) between May 2017 and May
2020, were enrolled in the present study. All healthy controls
showed normal physiological parameters in systemic physiological
examination, including bone marrow blasts <4%, and had no
history of AML. All patients were diagnosed using a bone marrow
test. All enrolled patients had a percentage of bone marrow blasts
>20% (with a mean of 21.5%) and no previous history of AML or
other malignancies. Patients with other clinical disorders (such as
metabolic disorders, chronic diseases and severe infection), and
who had initiated therapy for such disorders within 3 months prior
to admission, were excluded. The present study was approved by the
Ethics Committee of Xing'an League People's Hospital (approval no.
323SE), and all patients and control subjects provided written
informed consent.
Bone marrow mononuclear cells (BMMNCs)
and AML cell lines
Bone marrow was collected from all patients and
healthy controls by biopsy, and used to isolate BMMNCs using
lymphocyte separation medium (TBD Science; Tian Jin Hao Yang
Biological Manufacture Co., Ltd.). The isolation procedure was
conducted following the manufacturer's instructions. Briefly, 2 ml
bone marrow mixed with medium was centrifuged at 400 × g for 15
min. The second layer of the supernatant (containing the
lymphocytes) was then used for cell culture. The Beckman MoFlo
Astrios high-performance, live-cell sorting system (Beckman
Coulter, Inc.) was used to isolate BMMNC subpopulations, as
outlined in a previous study (17).
PE-conjugated mouse anti-human CXCR4 (cat. no. 60089PE.1; Stemcell
Technologies, Inc.) and APC-conjugated rat anti-human CD45 (cat.
no. 28145-1; Signalway Antibody LLC) were used to isolate the
BMMNCs, per the sorting identification strategy of a previous study
(17). The cells were resuspended in
minimal essential medium with Earle's salts (Gibco; Thermo Fisher
Scientific, Inc.) containing 10% fetal calf serum and 1%
antibiotics (penicillin/streptomycin; Gibco; Thermo Fisher
Scientific, Inc.); 1×107 resuspended bone marrow cells
were seeded into 100-mm cell culture dishes and incubated at 37°C
in a humidified incubator with 5% CO2. Kasumi-3 and
Kasumi-6 AML cell lines were purchased from the ATCC, and cultured
in RPMI-1640 medium (10% FBS) at 37°C in an incubator with 5%
CO2 and 95% humidity. Cells used in the subsequent
assays were collected at ~85% confluence.
Transient transfection
Overexpression of circ-ATAD1 and miR-34b was
achieved by transfecting 1×106 Kasumi-3 or Kasumi-6
cells with a circ-ATAD1 expression vector (1 µg; pcDNA3.1 backbone
vector; Invitrogen; Thermo Fisher Scientific, Inc.) or miR-34b
mimics (10 nM; Sigma-Aldrich; Merck KGaA) using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) (1:2). Untransfected and empty vector- or NC
miRNA-(Sigma-Aldrich; Merck KGaA) transfected cells were used as
the normal control and negative control (NC) cells, respectively.
The sequences of the miR-34b mimics and miR-NC are displayed in the
Table I. After incubation with the
transfection mixture for 6 h at 37°C, cells were immediately washed
with fresh medium, followed by culture in fresh medium for another
48 h prior to use.
 | Table I.Sequences of oligonucleotides. |
Table I.
Sequences of oligonucleotides.
| Oligonucleotide | Sequence (5′-3′) |
|---|
| Mimics NC
sense |
UUCUCCGAACGUGUCACGUU |
| Mimics NC
antisense |
AACGUGACACGUUCGGAGAA |
| miR-34b mimics
sense |
UAGGCAGUGUCAUUAGCUGAUUG |
| miR-34b mimics
antisense |
CAAUCAGCUAAUGACACUGCCUA |
RNA isolation and reverse
transcription-quantitative (RT-q)PCR
Total RNA was isolated from BMMNCs and in
vitro cultured cells using RNAzol reagent (Sigma-Aldrich; Merck
KGaA), and treated with DNA eraser (Takara Bio, Inc.) at 37°C until
the OD260 nm/280 nm ratio had reached ~2.0 (pure RNA). The
integrity of all RNA samples was determined by electrophoresis
using a 5% urea-PAGE gel. Total RNA was used for first-strand cDNA
synthesis with the Prime Script RT reagent kit (Takara Bio, Inc.)
per the manufacturer's instructions. Circ-ATAD1 expression was
determined using the SYBR® Green Quantitative RT-qPCR
Kit (Sigma-Aldrich; Merck KGaA) with GAPDH as the internal control.
Mature miR-34b expression was analyzed using the All-in-One™ miRNA
qRT-PCR Detection Kit (GeneCopoeia, Inc.) with U6 as the internal
control. All operations were completed following the manufacturers'
instructions. The qPCR thermocycling conditions for all genes,
cirRNA and miRNAs were as follows: 95°C for 1 min, followed by 40
cycles of 95°C for 10 sec and 60°C for 45 sec. PCR reactions were
performed using the CFX96 Touch Real-Time PCR Detection System
(Bio-Rad Laboratories, Inc.). Ct values of the targeted genes were
normalized to the corresponding internal controls based on the
2−ΔΔCq method, which was used to quantify gene
expression (18). The qPCR primers
are listed in Table II.
| Table II.Sequences of quantitative PCR
primers. |
Table II.
Sequences of quantitative PCR
primers.
| Gene | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| Circ-ATAD1 |
GTTTCCTTCCTGTGTGAGGC |
GGTCCGAGACGGTCCTTAAA |
| U6 |
CTCGCTTCGGCAGCACA |
AACGCTTCACGAATTTGCGT |
| miR-34b |
AGGCAGTGTCATTAGCTGATTGT |
ACAATCAGCTAATGACACTGCCT |
| GAPDH |
CCATTTGCAGTGGCAAAG |
CACCCTTTGTGTTAGTG |
Subcellular fractionation assay
Both the nuclear and cytoplasmic fractions of
Kasumi-3 and Kasumi-6 cells were prepared using the Nuclei
Isolation Kit: Nuclei EZ Prep (Sigma-Aldrich; Merck KGaA) according
to the manufacturer's instructions, and used for RNA isolation and
RT-qPCR to detect circ-ATAD1, with GAPDH as the internal
control.
Fluorescence in situ hybridization
(FISH) and immunofluorescence (IF)
IF staining with anti-histone H3 (1:300; cat. no.
ab6002; Abcam) was in reference to a previous study (19). The Blocking buffer was PBS with 1%
BSA (Thermo Fisher Scientific, Inc.). The Alexa fluor 555
anti-rabbit antibody (cat. no. ab150078; Abcam) was used as the
secondary antibody (1:500). FISH was performed using Kasumi-3 and
Kasumi-6 cells as described previously (19). The probe of cir-ATAD1
(TACCACAGCCTGGAGGCCCATAG) was synthesized and labeled with
digoxigenin (DIG-dUTP) by Sangon Biotech (Shanghai) Co., Ltd.
Specifically, the slices covered by cells were fixed in 4%
paraformaldehyde (MilliporeSigma) for 10 min at room temperature.
Before pre-hybridization, cells were permeabilized with cold 0.1%
Triton X-100 and pre-hybridized with a hybridization buffer at 37°C
for 1 h. The slides were incubated with a hybridization buffer
containing the FISH probe at 95°C for 5 min, and then at 37°C
overnight in the dark in a humid chamber. The samples were washed
with 2 × saline sodium citrate buffer (SSC) for 10 min at 37°C, 1 ×
SSC for 2 × 5 min at 37°C, and 0.5 × SSC for 10 min at room
temperature. The slides were then incubated with anti-DIG-488
(1:300; cat. no. ab150077; Abcam) at 37°C for 50 min, and the
nuclei were counterstained with DAPI at room temperature for 30
min. Finally, the slices were sealed in fluorescence
decay-resistant medium and images were obtained under a
fluorescence microscope (Nikon Corporation).
Methylation-specific PCR (MSP)
Genomic DNA isolation from transfected Kasumi-3 and
Kasumi-6 cells was performed using a routine method (20). All genomic DNA samples were processed
using the EZ DNA Methylation-Gold™ Kit (Zymo Research Corp.) per
the manufacturer's protocol. Then, qPCR and routine PCR were
performed to detect methylation of the miR-34b gene promoter using
PCR Master Mix ×2 (Invitrogen; Thermo Fisher Scientific, Inc.). The
MSP primers can be used to amplify methylated template, while the
primers for unmethylation-specific PCR (USP) do not amplify these
products. Both the MSP and USP conditions were as following: 95°C
for 5 min, followed by 40 cycles of 95°C for 30 sec, 55°C for 30
sec and 72°C for 50 sec, and then 72°C for 10 min. All primers used
for MSP and USP are listed in Table
III.
| Table III.Sequences of primers used for
MSP. |
Table III.
Sequences of primers used for
MSP.
| Gene | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| miR-34b USP |
TTTTTATTTGTTTTGTTTTGTGTTTGTTTTG |
CAACTACAACTCCCAAACAATCC |
| miR-34b MSP |
ATTCGTTTCGTTTCGCGTTCGTTTC |
CGACTACAACTCCCGAACGATCCG |
BrdU assay
A total of 3×103 transfected Kasumi-3 and
Kasumi-6 cells were transferred to each well of a 96-well plate in
0.1 ml medium, and cultured at 37°C for 48 h before the addition of
BrdU. The experiment was conducted using the BrdU Cell
Proliferation Assay (cat. no. QIA58; Sigma-Aldrich; Merck KGaA)
according to the manufacturer's protocol. Then, cells were cultured
with 20 µl/well diluted BrdU reagent (10 mM) for 6 h, fixed with
Fixing solution and incubated for 30 min. After fixation, the cells
were incubated for another 48 h with peroxidase-coupled
anti-BrdU-antibody (supplied by the kit), followed by washing twice
with ice-cold PBS. After incubation with peroxidase substrate for 3
h, OD values were measured at 450 nm. For the represent images, the
anti-Brdu (1:500; cat. no. ab6362; Abcam) was used as primary
antibody, and Goat anti-rabbit Alexa Fluor® 546
(1:2,000; cat. no. A11010; Invitrogen; Therno Fisher4 Scientific,
Inc.) was used as the secondary antibody. The detailed method has
been previously published (21).
Statistical analysis
AML and control groups were compared by unpaired
t-test. Comparisons among multiple independent cell transfection
groups were analyzed by one-way ANOVA followed by Tukey's test.
Correlations were analyzed by Pearson's correlation coefficient.
Data are presented as the mean ± standard deviation of three
independent replicates. P<0.05 was considered to indicate a
statistically significant difference.
Results
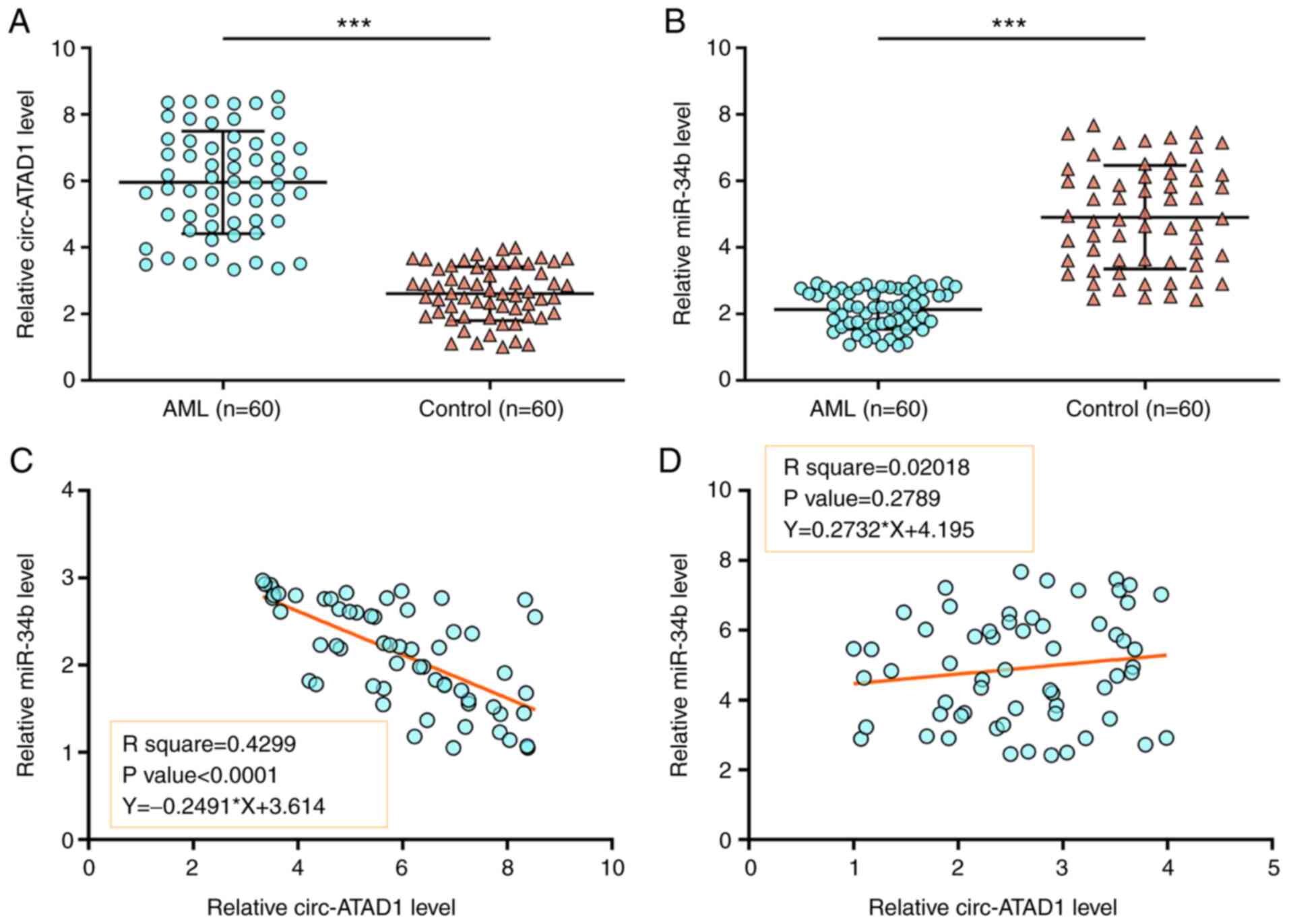
Patients with AML exhibit altered
expression of circ-ATAD1 and miR-34b
Samples of BMMNCs from both patients with AML (n=60)
and healthy controls (n=60) were subjected to RNA isolation and
RT-qPCR to determine the differential expression of circ-ATAD1 and
miR-34b. Compared with the controls, circ-ATAD1 was highly
expressed in AML (Fig. 1A,
P<0.01), while miR-34b expression was lower in AML (Fig. 1B, P<0.01), suggesting that
circ-ATAD1 upregulation and miR-34a downregulation may be involved
in AML. To study the crosstalk between circ-ATAD1 and miR-34b,
their correlations across both AML and control samples were
determined using Pearson's correlation analysis. The data showed
that circ-ATAD1 and miR-34b were closely and inversely correlated
across AML samples (Fig. 1C;
P<0.0001), but not across the control samples (Fig. 1D). Therefore, circ-ATAD1 and miR-34b
may interact with each other in AML.
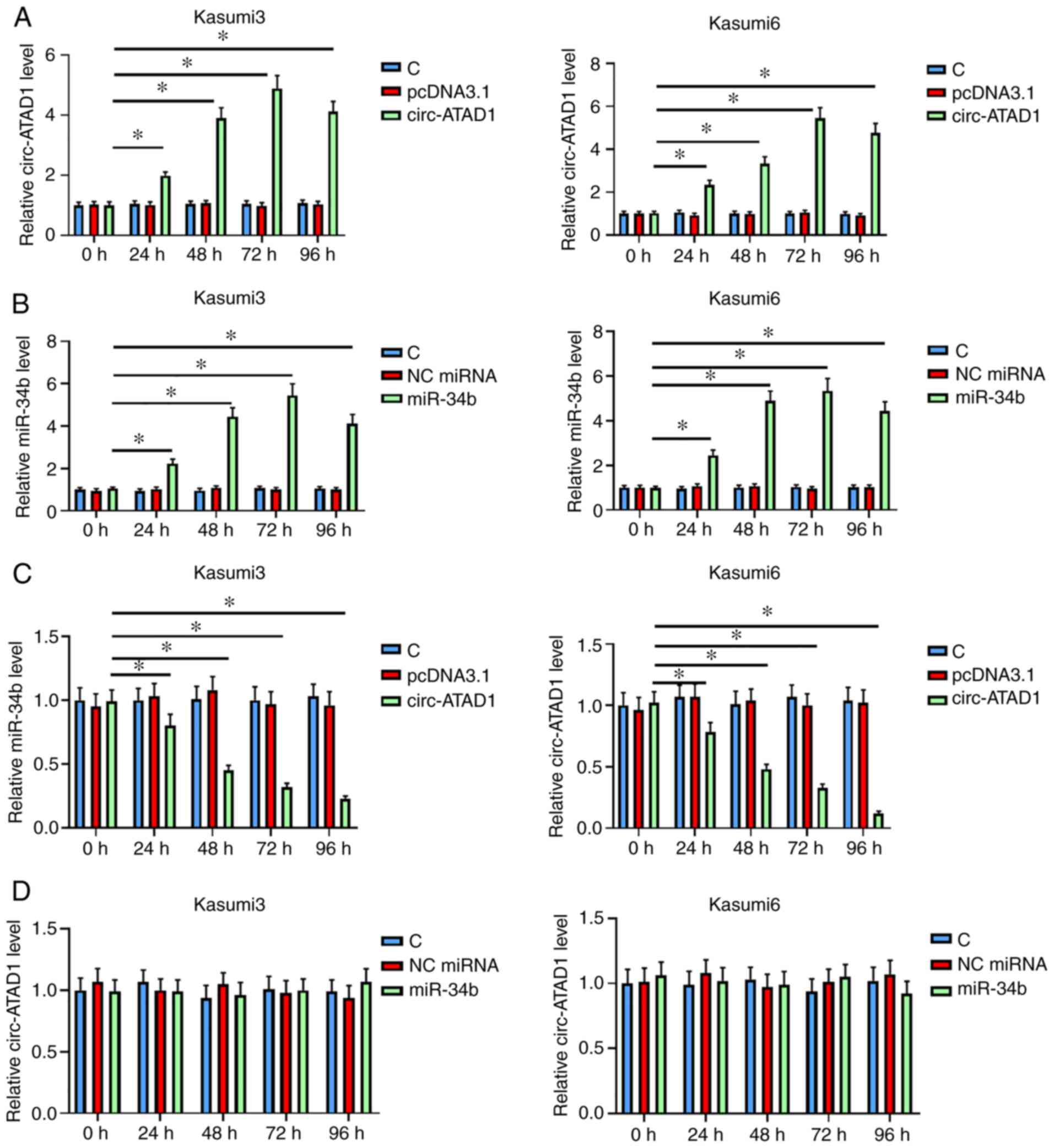
circ-ATAD1 overexpression decreases
miR-34b expression in AML cells
To further study the crosstalk between circ-ATAD1
and miR-34b, Kasumi-3 and Kasumi-6 cells were transfected with
either a circ-ATAD1 overexpression vector or miR-34b mimics,
followed by expression confirmation every 24 h until 96 h. It was
observed that both circ-ATAD1 and miR-34b were overexpressed
between 24 and 96 h post transfection (Fig. 2A and B, P<0.05). In addition,
circ-ATAD1 overexpression decreased that of miR-34b (Fig. 2C, P<0.05). By contrast, miR-34b
overexpression failed to significantly alter circ-ATAD1 expression
(Fig. 2D). Therefore, circ-ATAD1 may
downregulate miR-34b expression in AML cells.
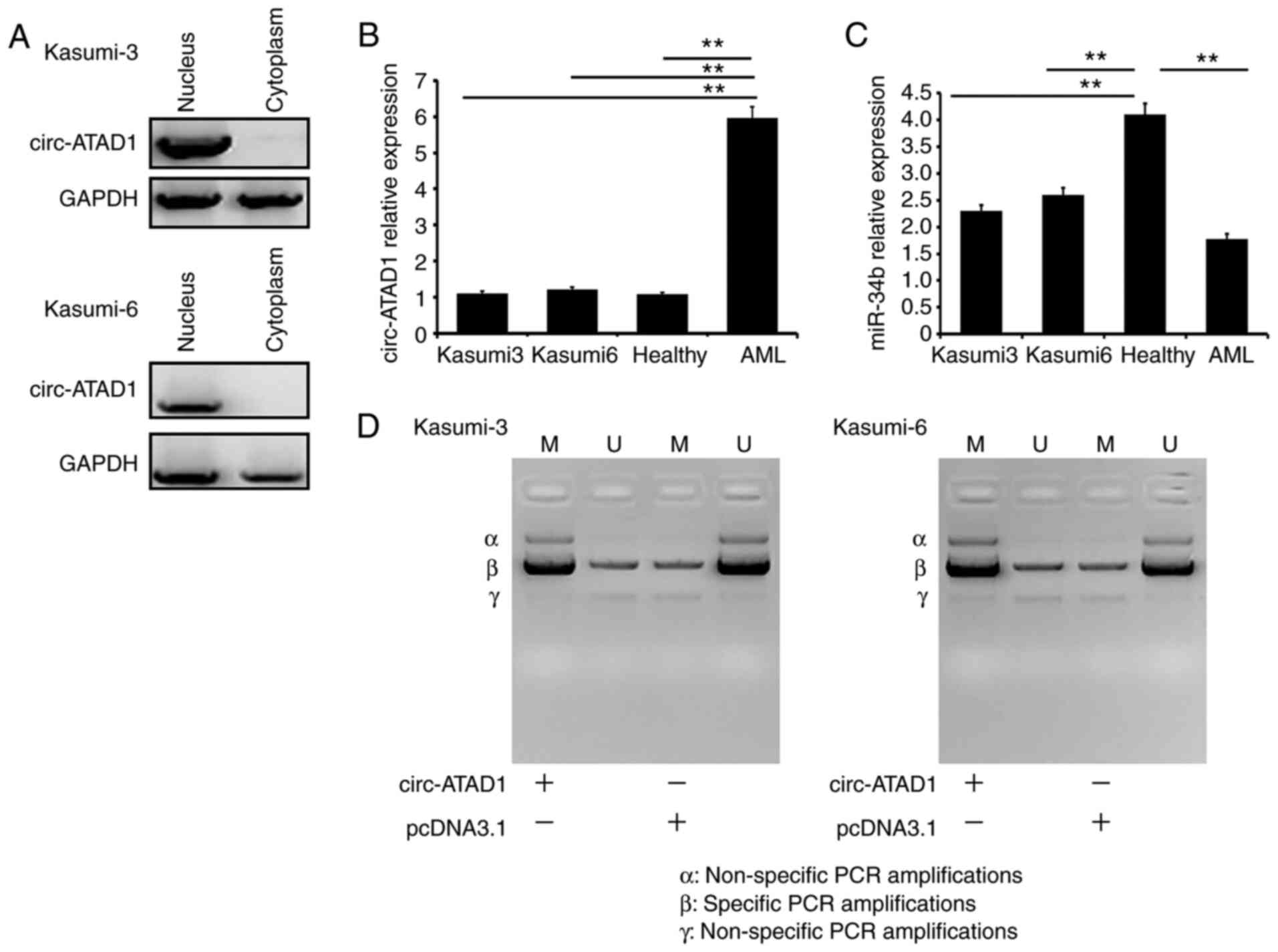
Circ-ATAD1 is a nucleus-specific
circRNA that increases miR-34b gene methylation in AML cells
A subcellular fractionation assay was used to
determine the subcellular location of circ-ATAD1 in both Kasumi-3
and Kasumi-6 cells, and circ-ATAD1 was only detected in the nuclear
fractions (Fig. 3A). The relative
expression of circ-ATAD1 and miR-34b in Kasumi-3 cells (nuclei), in
Kasumi-6 cells (nuclei), in the control and that in the AML patient
samples is also provided. The expression levels of circ-ATAD1 were
highest in AML samples (Fig. 3B;
P<0.01), and those of miR-34b were highest in the healthy
control samples (Fig. 3C;
P<0.01). FISH (Figs. S1 and
S2) also showed similar results. In
addition, the effects of circ-ATAD1 overexpression on miR-34b gene
methylation were analyzed by MSP. Compared with cells transfected
with empty pcDNA3.1 vector, cells transfected with the circ-ATAD1
expression vector showed increased methylation of miR34b (Fig. 3D). These results indicate the
significantly different expression of these two molecules in
healthy and tumor cells. In addition, the expression of miR-34b was
regulated by circ-ATAD1 through methylation-related mechanisms in
Kasumi-3 and Kasumi-6 cells.
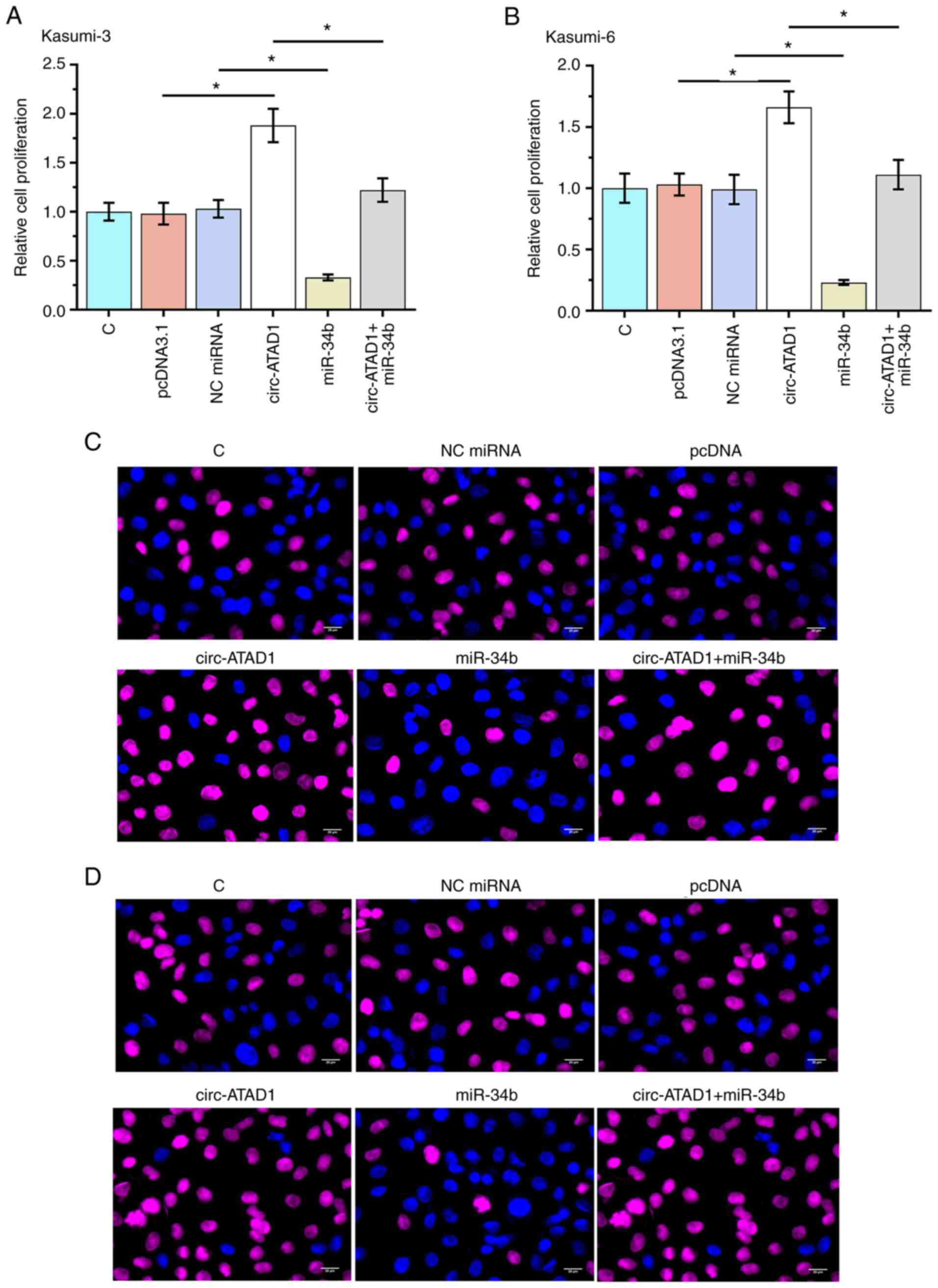
Circ-ATAD1 overexpression promotes the
proliferation of AML cells through miR-34b
The role of circ-ATAD1 and miR-34b in regulating
Kasumi-3 (Fig. 4A and C) and
Kasumi-6 (Fig. 4B and D) cell
proliferation was analyzed using a BrdU assay. Circ-ATAD1
overexpression increased cellular proliferation, while miR-34b
overexpression decreased cellular proliferation (both (P<0.05).
Moreover, co-transfection analysis showed that the effect of
circ-ATAD1 overexpression on AML cell proliferation was reduced by
miR-34b overexpression (P<0.05). The results indicated
Circ-ATAD1 overexpression promotes the proliferation of AML cells
through miR-34b. The Circ-ATAD1-miR-34b cell signaling axis, which
contributed the tumor cell proliferation, has been revealed.
However, methylation-related mechanisms were the major regulation
mechanism between Circ-ATAD1 and miR-34b.
Discussion
BMMNCs are located in hematopoietic niches and play
important roles in protecting leukemic cells from conventional
chemotherapy (19). Leukemia
initiating cells (LICs) interact with their surrounding bone marrow
microenvironment (22), and the
complex cell network surrounding LICs determines their fate
(23). Various cellular signaling
pathways and metabolic mechanisms regulate the BMMNC
microenvironment and contribute to AML development (24,25).
However, the detailed regulation mechanism for BMMNC malignant
transformation has not been clearly defined.
The involvement of circ-ATAD1 in AML, and its
potential crosstalk with miR-34b, a tumor suppressor in AML
(26), were investigated in the
present study. As such, circ-ATAD1 was found to be upregulated in
AML, which downregulated miR-34b expression through methylation,
promoting AML cell proliferation.
A previous study characterized circ-ATAD1 as an
oncogenic circRNA in gastric cancer (16), where it was reported to be
upregulated. Furthermore, circ-ATAD1 was found to sponge miR-140-3p
to upregulate YY1 expression. Additionally, YY1 activates the
transcription of phosphorylated CTD interacting factor 1 to promote
cancer cell proliferation, invasiveness and migration (16). Consistently, circ-ATAD1 was found to
be upregulated in AML in the present study, with enhancing effects
on cancer cell proliferation. Therefore, circ-ATAD1 is likely an
oncogenic circRNA in AML. However, as miRNA sponges, most circRNAs
are primarily localized in the cytoplasm (27); the present study revealed that
circ-ATAD1 was only detected in the nucleus, and not in the
cytoplasm. Therefore, we hypothesize that circ-ATAD1 may be
expressed in different subcellular locations in different cell
types, though further confirmative investigation is required.
miR-34b was recently confirmed to target heat shock
factor protein 1 in AML, which suppressed cell survival (26). Consistently, the present study showed
that miR-34b was downregulated in AML, and that its overexpression
suppressed cellular proliferation, further confirming the tumor
suppressive role of miR-34b in AML. However, to the best of our
knowledge, the upstream regulators of miR-34b in AML have not been
reported previously. In the current study, circ-ATAD1 was shown to
be a nucleus-specific circRNA. DNA methylation is known to occur in
the nucleus (28), and numerous
circRNAs potentially regulate the methylation of miRNAs in
association with cancer biology (29,30).
Notably, the present study revealed that circ-ATAD1 downregulated
miR-34b expression in AML by increasing methylation of the miR-34b
gene promoter. Furthermore, miR-34b mimics reversed circ-ATAD1
overexpression-induced cellular proliferation, indicating an
interaction between the two noncoding RNAs. However, a moderate
correlation was observed between circ-ATAD1 and miR-34b across AML,
but not the control samples. Therefore, certain AML-related factors
may be involved in mediating the interaction between circ-ATAD1 and
miR-34b. Collectively, the results of the present study revealed
the circ-ATAD1-miR-34b axis as a novel regulatory signaling pathway
specific to AML malignant transformation. However, its downstream
genes, and the specific role and contribution in different cell
types at different transformation phases, requires further
investigation.
In conclusion, circ-ATAD1 is upregulated in AML and
promotes AML cell proliferation by downregulating miR-34b via
promoter methylation.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This work was supported by the Natural Science
Foundation of Inner Mongolia (grant no. 2019MS08047).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YZ put forward the concept, designed the
experiments, provided general supervision, edited the manuscript
and was a guarantor of integrity of the entire study. YW, BG and XQ
acquired and analyzed the data. YW wrote the manuscript. LB, BL and
HB conducted the literature search and interpretated the data. XW
and XYW defined intellectual content, conducted the literature
search, were involved in interpreting the results and data
analysis, and revised the manuscript. YZ and YW confirm the
authenticity of all the raw data. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Xing'an League People's Hospital. All experiments were
performed in accordance with the 1964 Declaration of Helsinki and
its later amendments. Written informed consent to participate in
the study was obtained from patients and controls prior to sample
collection.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
AML
|
acute myelogenous leukemia
|
|
BMMNCs
|
bone marrow mononuclear cells
|
|
FISH
|
fluorescence in situ
hybridization
|
|
IF
|
immunofluorescence
|
|
MSP
|
methylation-specific PCR
|
|
USP
|
unmethylation-specific PCR
|
|
LICs
|
leukemia initiating cells
|
References
|
1
|
Rose-Inman H and Kuehl D: Acute leukemia.
Hematol Oncol Clin North Am. 31:1011–1028. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Parkin B, Ouillette P, Yildiz M,
Saiya-Cork K, Shedden K and Malek SN: Integrated genomic profiling,
therapy response, and survival in adult acute myelogenous leukemia.
Clin Cancer Res. 21:2045–2056. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Schiller GJ: High-risk acute myelogenous
leukemia: Treatment today and tomorrow. Hematology Am Soc Hematol
Educ Program. 2013:201–208. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gores GJ and Kaufmann SH: Selectively
targeting Mcl-1 for the treatment of acute myelogenous leukemia and
solid tumors. Genes Dev. 26:305–311. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jiang XJ, Huang KK, Yang M, Qiao L, Wang
Q, Ye JY, Zhou HS, Yi ZS, Wu FQ, Wang ZX, et al: Synergistic effect
of panobinostat and bortezomib on chemoresistant acute myelogenous
leukemia cells via AKT and NF-κB pathways. Cancer Lett.
326:135–142. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Piya S, Andreeff M and Borthakur G:
Targeting autophagy to overcome chemoresistance in acute
myleogenous leukemia. Autophagy. 13:214–215. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gill H, Leung AY and Kwong YL: Molecularly
targeted therapy in acute myeloid leukemia. Future Oncol.
12:827–838. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hatzimichael E, Georgiou G, Benetatos L
and Briasoulis E: Gene mutations and molecularly targeted therapies
in acute myeloid leukemia. Am J Blood Res. 3:29–51. 2013.PubMed/NCBI
|
|
9
|
Konig H and Levis M: Is targeted therapy
feasible in acute myelogenous leukemia? Curr Hematol Malig Rep.
9:118–127. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Martelli AM, Evangelisti C, Chiarini F and
McCubrey JA: The phosphatidylinositol 3-kinase/Akt/mTOR signaling
network as a therapeutic target in acute myelogenous leukemia
patients. Oncotarget. 1:89–103. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pei S, Minhajuddin M, Callahan KP, Balys
M, Ashton JM, Neering SJ, Lagadinou ED, Corbett C, Ye H, Liesveld
JL, et al: Targeting aberrant glutathione metabolism to eradicate
human acute myelogenous leukemia cells. J Biol Chem.
288:33542–33558. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Vo JN, Cieslik M, Zhang Y, Shukla S, Xiao
L, Zhang Y, Wu YM, Dhanasekaran SM, Engelke CG, Cao X, et al: The
landscape of circular RNA in cancer. Cell. 176:869–881.e13. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fang Y and Fullwood MJ: Roles, functions,
and mechanisms of long non-coding RNAs in cancer. Genomics
Proteomics Bioinformatics. 14:42–54. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rücker FG, Russ AC, Cocciardi S, Kett H,
Schlenk RF, Botzenhardt U, Langer C, Krauter J, Fröhling S,
Schlegelberger B, et al: Altered miRNA and gene expression in acute
myeloid leukemia with complex karyotype identify networks of
prognostic relevance. Leukemia. 27:353–361. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Benassi B, Marani M, Loda M and Blandino
G: USP2a alters chemotherapeutic response by modulating redox. Cell
Death Dis. 4:e8122013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang L, Chang X, Zhai T, Yu J, Wang W, Du
A and Liu N: A novel circular RNA, circ-ATAD1, contributes to
gastric cancer cell progression by targeting miR-140-3p/YY1/PCIF1
signaling axis. Biochem Biophys Res Commun. 525:841–849. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang J, Liu X, Lu H, Jiang C, Cui X, Yu L,
Fu X, Li Q and Wang J: CXCR4(+)CD45(−) BMMNC subpopulation is
superior to unfractionated BMMNCs for protection after ischemic
stroke in mice. Brain Behav Immun. 45:98–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rao X, Huang X, Zhou Z and Lin X: An
improvement of the 2(-delta delta CT) method for quantitative
real-time polymerase chain reaction data analysis. Biostat
Bioinforma Biomath. 3:71–85. 2013.PubMed/NCBI
|
|
19
|
Pavani RS and Elias MC: Following
trypanosoma cruzi RPA-DNA interaction using fluorescent in situ
hybridization coupled with immunofluorescence (FISH/IF). Methods
Mol Biol. 2281:209–215. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yuan Y, Wang Q, Ma SL, Xu LQ, Liu MY, Han
B, Du N, Sun XL, Yin XL and Cao FF: lncRNA PCAT-1 interacting with
FZD6 contributes to the malignancy of acute myeloid leukemia cells
through activating Wnt/β-catenin signaling pathway. Am J Transl
Res. 11:7104–7114. 2019.PubMed/NCBI
|
|
21
|
Mehdipour M, Etienne J, Chen CC, Gathwala
R, Rehman M, Kato C, Liu C, Liu Y, Zuo Y, Conboy MJ and Conboy IM:
Rejuvenation of brain, liver and muscle by simultaneous
pharmacological modulation of two signaling determinants, that
change in opposite directions with age. Aging (Albany NY).
11:5628–5645. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ishikawa F, Yoshida S, Saito Y, Hijikata
A, Kitamura H, Tanaka S, Nakamura R, Tanaka T, Tomiyama H, Saito N,
et al: Chemotherapy-resistant human AML stem cells home to and
engraft within the bone-marrow endosteal region. Nat Biotechnol.
25:1315–1321. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nakamura-Ishizu A, Takubo K, Kobayashi H,
Suzuki-Inoue K and Suda T: CLEC-2 in megakaryocytes is critical for
maintenance of hematopoietic stem cells in the bone marrow. J Exp
Med. 212:2133–2146. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Stevens AM, Xiang M, Heppler LN, Tošić I,
Jiang K, Munoz JO, Gaikwad AS, Horton TM, Long X, Narayanan P, et
al: Atovaquone is active against AML by upregulating the integrated
stress pathway and suppressing oxidative phosphorylation. Blood
Adv. 3:4215–4227. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Vignon C, Debeissat C, Bourgeais J, Gallay
N, Kouzi F, Anginot A, Picou F, Guardiola P, Ducrocq E, Foucault A,
et al: Involvement of GPx-3 in the reciprocal control of redox
metabolism in the leukemic niche. Int J Mol Sci. 21:85842020.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li G, Song Y, Zhang Y, Wang H and Xie J:
MiR-34b Targets HSF1 to suppress cell survival in acute myeloid
leukemia. Oncol Res. 24:109–116. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hansen TB, Jensen TI, Clausen BH, Bramsen
JB, Finsen B, Damgaard CK and Kjems J: Natural RNA circles function
as efficient microRNA sponges. Nature. 495:384–388. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Massart R, Barnea R, Dikshtein Y, Suderman
M, Meir O, Hallett M, Kennedy P, Nestler EJ, Szyf M and Yadid G:
Role of DNA methylation in the nucleus accumbens in incubation of
cocaine craving. J Neurosci. 35:8042–8058. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu Z, Yu Y, Huang Z, Kong Y, Hu X, Xiao
W, Quan J and Fan X: CircRNA-5692 inhibits the progression of
hepatocellular carcinoma by sponging miR-328-5p to enhance DAB2IP
expression. Cell Death Dis. 10:9002019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gu Y, Ci C, Zhang X, Su M, Lv W, Chen C,
Liu H, Zhang D, Zhang S and Zhang Y: Prediction of circRNAs based
on the DNA methylation-mediated feature sponge function in breast
cancer. Front Bioeng Biotechnol. 7:3652019. View Article : Google Scholar : PubMed/NCBI
|