Introduction
Cholangiocarcinoma (CCA) is a lethal malignancy originating from the biliary epithelium; it accounted for 10–15% of all primary liver cancers and 3% of gastrointestinal cancers between 1996 and 2016 in European countries (1,2), and its incidence is increasing (2,3). The incidence of CCA is high in East and Southeast Asia, where liver-fluke infection is endemic, the primary cause of CCA in the region. In western countries, CCA is primarily associated with primary sclerosing cholangitis, hepatolithiasis and choledochal cysts. CCA has no specific symptoms and is undetectable at the early stages. Most patients with CCA are thus diagnosed at an advanced stage with local or distant metastasis (2,4). Curative surgery is commonly used to treat patients with CCA, but most cases are inoperable (5–7). Gemcitabine, cisplatin and 5-Fluorouracil (5-FU) are the chemotherapy regimens commonly used to improve the length of survival, with median survival times of 2–12 months recorded between 2014 and 2016 (8,9).
Recently, several groups have reported the genetic basis of CCA using advanced sequencing protocols. Numerous known cancer-related genes have been detected in CCA, including TP53, KRAS and SMAD4 (10). Other newly implicated genes related to Wnt signalling have been detected in 13% of patients with CCA, including APC, RNF43 and CTNNB1 (11). Several Wnt signalling-related genes are upregulated in CCA [Frizzled (FZD) receptor and ligand Wnt3a, Wnt5a, Wnt7b and Wnt10a] (12,13), while some are downregulated (such as RNF43). The Wnt-signalling pathway in CCA is implicated in both scenarios.
The ring finger protein 43 (RNF43), an integral membrane E3 ubiquitin ligase, negatively regulates the Wnt/β-catenin signalling pathway. It does this in two ways: i) By ubiquitinating the cytoplasmic site on the Frizzled receptor and lysosome degradation (14); or ii) by sequestering T-cell factor 4 (TCF4) to the nuclear membrane, thereby inhibiting Wnt-mediated transcription (15). The RNF43 loss-of-function mutation is associated with the progression of several cancers via an increase in Wnt signalling by promoting FZD receptor expression in pancreatic ductal adenocarcinoma (16), enhancing stemness properties and impairing the DNA damage response in gastric cancer (17,18). The loss of endogenous RNF43 expression also enhances proliferation and the invasive capacity of gastric and colorectal cancer cells (18). Inactivating the mutation and downregulating RNF43 are associated with a less favourable survival of patients with CCA (10,19). Since a functional analysis of RNF43 in CCA has not yet been performed, this present aimed to determine the functional role of RNF43 in CCA and its underlying mechanisms. Overexpression of RNF43 was achieved in the CCA cell line KKU-213B. The regulation of the Wnt-signalling pathway via RNF43 overexpression was determined by measuring key downstream proteins (β-catenin, TCF-4 and the transcriptional level of known Wnt-targeted genes). The current study also sought to determine the biological effect of RNF43 on cell proliferation and migration, as well as the sensitivity of CCA to chemotherapeutic drugs.
Materials and methods
Cell lines and cell culture conditions
Four human CCA cell lines (KKU-055, KKU-100, KKU213A and KKU-213B) were used in the present study. KKU-055 and KKU-100 were obtained from the Japanese Collection of Research Bioresources Cell Bank on June 22, 2015 (20). Two CCA cell lines -KKU-213A and KKU-213B were provided by Dr Sopit Wongkham after being authenticated in ref. 21 where Dr Sopit Wongkham is a co-author. According to the report of Sripa et al, KKU-213A and KKU-213B were identified as two different clones from a patient with CCA (ID-M213) (21). KKU-213A and KKU-213B share similar short tandem repeats (STR) but exhibited several distinct phenotypes (21). Cell growth and cell migration were verified before continuation of the study. All the cell lines were cultured in DMEM Nutrient Mixture (cat. no. 12100-046; Gibco; Thermo Fisher Scientific, Inc.) supplemented with 1% penicillin-streptomycin (cat. no. 15140-122; Gibco; Thermo Fisher Scientific, Inc.) and 10% FBS (cat. no. 10270-098; Gibco; Thermo Fisher Scientific, Inc.). Cells were incubated at 37°C in a humidified atmosphere containing 5% CO2. Cells with 70-80% confluence at 24 h were trypsinised with 0.25% trypsin-EDTA for 3 min at 37°C and sub-cultured in the same media.
Overexpression of RNF43
KKU-213B cells were cultured in a 6-well plate (4×105 cells/well) overnight. Cells were transfected with 2.5 µg of pCMV6-RNF43-FLAG tag-construct and control expression vector (cat. no. PS100001; OriGene Technologies, Inc.) using Lipofectamine® 3000 reagent (cat. no. 3000-008; Invitrogen; Thermo Fisher Scientific, Inc.) according to the manufacturer's instructions. The transfection complex in Opti-MEM (cat. no. 22600-050; Gibco; Thermo Fisher Scientific, Inc.) was removed after 24 h and replaced with DMEM supplement with 10% FBS for another 24 h at 37°C in a humidified atmosphere containing 5% CO2. After 24 or 48 h of transfection, cells were used for subsequent experiments.
Cell proliferation
Transfected KKU-213B cells with pCMV6-RNF43 construct or control expression vector were seeded at 1×104 cells/well into a 96-well plate and incubated for 0, 24, 48 and 72 h at 37°C in a humidified atmosphere containing 5% CO2. The numbers of viable cells were measured at each time point using Sulforhodamine B (SRB) assay as described by Talabnin et al (22). The experiment was performed independently three times.
Drug treatment
For drug treatments, a stock concentration of 384.4 mM of 5-fluorouracil (5-FU, cat. no. 1C 229/39; Boryung Pharmaceutical Co., Ltd.) was prepared in sterile water and stored at room temperature. Various concentrations of 5-FU were diluted in cell culture media for the drug sensitivity assay (0, 3.125, 6.25, 12.5; 25 and 50 µM). KKU-213B cells transfected with pCMV6-RNF43 construct or control expression vector were seeded at 1×104 cells/well into a 96-well plate. After 24 h seeding, both types of transfected cells were treated with the range of 5-FU concentrations for 72 h. Cell viability was measured using the SRB assay (22). Cell viability (%) = (A564 in treatment wells) ÷ (A564 in control wells) ×100. The half-maximal inhibitory concentration (IC50) values were calculated using GraphPad Prism 5 (GraphPad Software, Inc.). The experiment was performed independently three times.
Colony formation assay
Transfected cells were seeded at a density of 2×103 cells/well into a 6-well plate. After 24 h seeding, the transfected cells were treated with various concentrations of 5-FU (0, 3.125, 6.25, 12.5, 25 and 50 µM). After 48 h of drug treatment, the 5-FU was removed, replaced with DMEM supplemented with 10% FBS and incubated for 14 days. Crystal violet staining was used to visualise the colonies formed. Next, the colonies were washed thrice with 1X PBS. Absolute methanol was used to fix the colonies for 2 h at −20°C, followed by staining with 0.5% (w/v) crystal violet in 100% methanol for 1 h at room temperature. Finally, colonies were washed with tap water and air-dried. A colony was defined as consisting of at least 50 cells, and colony area was captured and calculated from three independent experiments using ImageJ software (version 1.53a; National Institutes of Health).
Wound scratching
KKU-213B was seeded at 4×105 cells/well into a 6-well plate and transfected with pCMV6-RNF43 construct or control expression vector as described. After 24 h of transfection with 90% cell confluence in serum-free Opti-MEM, a vertical wound was scratched through the cell monolayer using a 200 µl pipette tip. The debris was removed and replaced with DMEM supplemented with 10% FBS. The wound width at 0 h was measured before incubation. After incubation at 37°C in 5% CO2 for another 16, 20 and 24 h, wound closure was measured and the wound size was calculated using ImageJ software version 1.53a (National Institute of Health). In addition, the relative wound healing rate (relative to 0 h) was calculated.
Transwell migration assay
The Boyden chamber assay was used to track migratory cells using Transwell cell culture inserts (8.0-µm pore size, Corning, Inc.) in a 24-well plate. After 24 h of transfection, transfected cells were seeded at a density of 3×104 cells/well in 200 µl of serum-free DMEM into the upper compartment of the chamber. Next, DMEM (600 µl) supplemented with 10% FBS was placed into the lower chamber. After incubation at 37°C for 16 and 24 h, the migratory cells were stained using the SRB assay as described by Talabnin et al (22). After SRB staining, the number of migrating cells was visualized using an Olympus inverted microscope with ×100 magnification. Then absorbance at 564 nm was used as a proxy for the number of migratory cells.
Reverse transcription-quantitative (RT-q)PCR
Total RNA of transfected KKU-213B cells with pCMV6-RNF43 construct or control cells was extracted using TRIzol® Reagent (cat. no. 15596026; Invitrogen; Thermo Fisher Scientific, Inc.). According to the manufacturer's instructions, the first-strand cDNA was synthesised using a SensiFAST™ cDNA Synthesis kit (cat. no. BIO-65053; Bioline; Meridian Bioscience). qPCR was performed using a LightCycler® 480 SYBR-Green I Master (cat. no. 04707516001; Roche Molecular Systems, Inc). The sequences of primers for all the Wnt targeted genes are listed in Table I. Gene amplification was performed with slight modifications: Initial denaturation at 95°C for 5 min; followed by denaturation at 95°C for 10 sec, annealing at 58°C or 60°C for 10 sec and extension at 72°C for 10 sec, for 45 cycles (22). The annealing temperature was 58°C for RNF43, survivin (BIRC5), CHD1, c-MYC and ATP-binding cassette sub-family B member 1 (ABCB1), and 60°C for AXIN2, CDH2, CCND1, MMP-2, MMP-7, MM7-9 and ABCC1 (MRP1). The gene expression level was normalised against the housekeeping gene β-actin. Relative gene expression was calculated by 2−ΔΔCq (23).
 |
Table I.
Primer list of Wnt target genes and epithelial-mesenchymal transition genes.
|
Table I.
Primer list of Wnt target genes and epithelial-mesenchymal transition genes.
| Target gene |
Forward |
Reverse |
| RNF43 |
5′-TCTGTCTGGAGGAGTTCTCT-3′ |
5′-CTCCCTCTGTGATGTTGAAC-3′ |
| AXIN2 |
5′-CAGATCCGAGAGGATGAAGAGA-3′ |
5′-AGTATCGTCTGCGGGTCTTC-3′ |
| BIRC5 |
5′-CTGCTGTGGACCCTACTG-3′ |
5′-AACTGCGTCTCTGCCAGGAC-3′ |
| CDH1 |
5′-CACCACGGGCTTGGATTTTG-3′ |
5′-TGGGGGCTTCATTCACATCC-3′ |
| CDH2 |
5′-ATCCTGCTTATCCTTGTGCTG-3′ |
5′-GTCCTGGTCTTCTTCTCCTCC-3′ |
| c-MYC |
5′-TGAGGAGACACCGCCCAC-3′ |
5′-CAACATCGATTTCTTCCTCATCTTC-3′ |
| MMP-7 |
5′-CAGATGTGGAGTGCCAGATG-3′ |
5′-TGTCAGCAGTTCCCCATACA-3′ |
| MMP-2 |
5′-TGACCAAGGGTACAGCCTGT-3′ |
5′-AGAGGCCCCATAGAGCTCCT-3′ |
| MMP-9 |
5′-CTCTGGAGGTTCGACGTGAA-3′ |
5′-GGCTTTCTCTCGGTACTGGA-3′ |
| CCND1 |
5′-AACTACCTGGACCGCTTCCT-3′ |
5′-CCACTTGAGCTTGTTCACCA-3′ |
| ABCB1 (MDR1) |
5′-CTCAGACAGGATGTGAGTTGGT-3′ |
5′-ACAGCAAGCCTGGAACCTAT-3′ |
| ABCC1 (MRP1) |
5′-CTGGGCTTATTTCGGATCAA-3′ |
5′-TGAATGGGTCCAGGTTCATT-3′ |
| ACTB |
5′-GATCAGCAAGCAGGAGTATGACG-3′ |
5′-AAGGGTGTAACGCAACTAAGTCATAG-3′ |
Protein collection
After 48 h of transfection, cells were washed with 1X PBS then lysis buffer was added [150 mM NaCl; 50 mM Tris-HCl pH 7.4; 1% Sodium deoxycholate; 0.1% SDS; and 1X Protease inhibitor cocktail (Roche Diagnostics)]. The whole-cell lysate was harvested (19). A nuclear extraction kit (cat. no. 2900; Chemicon®; Merck Millipore) was used to prepare the nuclear and cytoplasmic fractions according to the manufacturer's protocols. The protein concentration of all protein samples was tested using a Pierce® BCA protein assay kit (cat no. 23225; Thermo Fisher Scientific, Inc.).
SDS-PAGE and western blot analysis
Protein samples (30 µg/lane) were subjected to protein separation using 10% SDS-PAGE. The separated proteins were further electro-transferred to Amersham Protran 0.45 NC nitrocellulose western blotting membranes (cat. no. 10600003; GE Healthcare) (22). Immunodetection of the proteins of interest was performed using the following specific monoclonal or polyclonal antibodies at 4°C overnight: Anti-RNF43 (cat. no. HPA008079; 1:1,000) from Sigma-Aldrich; Merck KGaA, anti-β-actin (cat. no. sc-47778; 1:2,000) and anti-β-catenin (cat. no. sc-7963; 1:1,000) from Santa Cruz Biotechnology, Inc., anti TCF4 (cat. no. 2569; 1:500), anti-Lamin B1 (cat. no. 12586; 1:500) and caspase-3 (cat. no. 9662: 1:1,000) from Cell Signaling Technology, Inc., and PARP1 (cat. no. 13371-1, 1:5,000), Bcl-2 (cat. no. 12789-1; 1:500) and Bax (cat. no. 50599-2; 1:1,000) from Proteintech. The membranes were then incubated with HRP-conjugated secondary antibodies (cat. no. NXA931 and NA934; GE Healthcare) at a dilution of 1:2,000 for 1 h at room temperature. Immunoreactivity of the target proteins was detected using an Immobilon Forte Western HRP substrate (cat. no. WBLUF0100; Merck Millipore) and illuminated on Amersham™ Hyperfilm™ (cat. no. 28906838; GE Healthcare). ImageJ software (version 1.53a; National Institutes of Health) was used to determine the density of each target protein then the relative intensity of the protein band was transformed into a quantitative value (i.e., normalised to β-actin). The respective ratio of each protein to β-actin and nuclear β-catenin to cytoplasmic β-catenin was calculated.
Immunofluorescence analysis
After 24 h of transfection, the cells were seeded onto 8-well cell culture slides at a density of 3×104 cells. After 24 h of seeding, the cells were fixed with 4% paraformaldehyde in PBS on ice for 15 min and permeabilized with 0.1% Triton X-100 in PBS for 10 min on ice. After washing with PBS, the samples were incubated with blocking solution (2% bovine serum albumin in 0.1% PBST) for 30 min on ice and with anti-Flag (cat. no. 8146S; 1:100; Cell Signaling Technology, Inc.) and anti-TCF4 (cat. no. 2569; 1:50; Cell Signaling Technology, Inc.) overnight at 4°C. After washing with PBS, anti-rabbit IgG-Alexa Fluor® 488 conjugate (cat. no. 4412S; 1:500; Cell Signaling Technology, Inc.) and anti-mouse IgG-Alexa™ 633 conjugate (cat. no. A21050; 1:500; Invitrogen; Thermo Fisher Scientific, Inc.) were used as secondary antibodies and incubated for 30 min in dark at room temperature. Cells were then stained with 300 nM DAPI solution for 3 min at room temperature. The samples were mounted with 50% glycerol in PBS and visualized using a Nikon confocal microscope (Nikon Corporation) with original magnification at ×200 and ×600.
Statistical analysis
Statistical analyses were performed using GraphPad Prism 5.0 (GraphPad Software, Inc.). The results (presented as the mean ± SEM) were calculated from three independent experiments. The Student's t-test was used for between-group statistical analyses. The one-way and two-way ANOVAs, followed by Bonferroni's correction, were used to compare multiple datasets. A value of P<0.05 was considered to indicate a statistically significant difference.
Results
RNF43 overexpression decreases β-catenin nuclear translocation and transcriptional level of Wnt target genes
The endogenous expression of RNF43 was determined in 4 CCA cell lines (KKU-055, KKU-100, KKU-213A and KKU-213B) to demonstrate its functional role. At baseline, KKU-213A and KKU213B had a higher RNF43 mRNA expression than KKU-055 and KKU-100. However, western blot analysis did not reveal any differences among cell lines at the protein level (Fig. 1A). Therefore, KKU-213B cells were used to determine the effect of RNF43 overexpression in a subsequent experiment because of their higher efficacy for transfection and lower transfection toxicity compared with KKU-055 and KKU-100 cells (data not shown). The exogenous expression of RNF43 in KKU-213B cells was detected at both mRNA and protein levels, but most RNF43 expression was in the nuclear fraction (Fig. 1B). Furthermore, a reduction in β-catenin nuclear translocation and nuclear TCF4 expression (Fig. 2A and B) and the low expression of six known WNT target genes [AXIN2, BIRC5, CCND1, c-MYC, MMP-7 and ABCB1 (MDR1)] was detected in RNF43-overexpressing KKU-213B cells (Fig. 2C). In order to verify the regulation of the Wnt/β-catenin pathway through the interaction between RNF43 and TCF4, double immunofluorescence staining of RNF43 and TCF4 was performed. Co-localization of RNF43 and TCF4 was observed at the nuclear rim in the RNF43-overexpressing KKU-213B cells. Moreover, TCF4 expression in the nuclear fraction was decreased after overexpression of RNF43 compared with the control (Fig. 2D). The results suggest that RNF43 has a role in inhibiting the Wnt signalling pathway via a reduction in TCF4 expression.
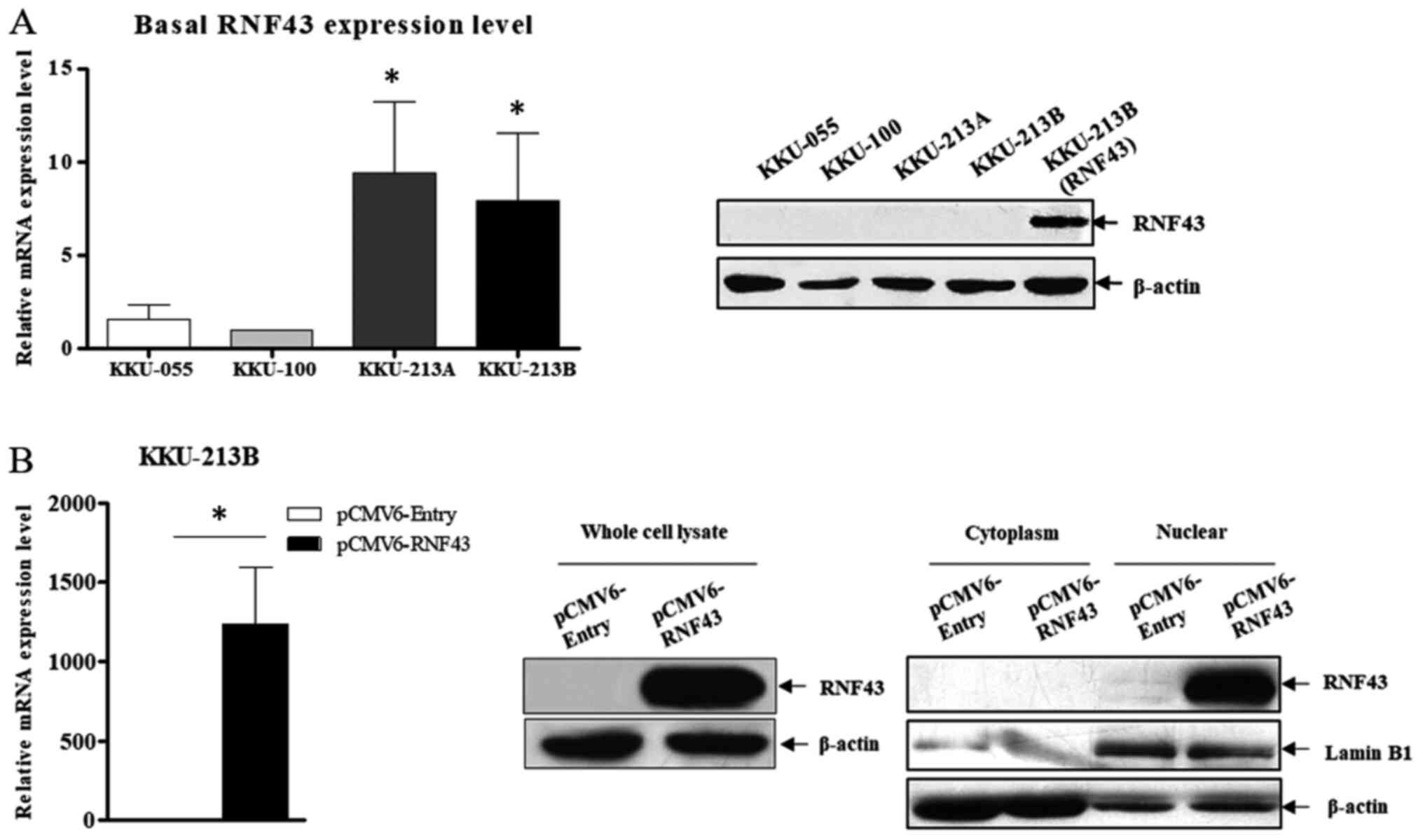 |
Figure 1.
RNF43 expression in CCA cell lines. (A) Basal expression of RNF43 in various CCA cell lines was determined using RT-qPCR and western blot analyses. RNF43 protein expression of the overexpressing KKU-213B cell line was used as the positive control. *P<0.05 vs. KKU-055. (B) The efficiency of RNF43 overexpression in KKU-213B after 24 h transfection was determined using RT-qPCR and western blot analyses. The mRNA expression level of RNF43 was normalised to β-actin as the reference genes and calculated using the 2−ΔΔCq method. Values are expressed as the mean ± SEM of three independent experiments. *P<0.05 vs. KKU-213B control. RNF43, ring finger protein 43; CCA, cholangiocarcinoma; RT-qPCR, reverse transcription-quantitative PCR.
|
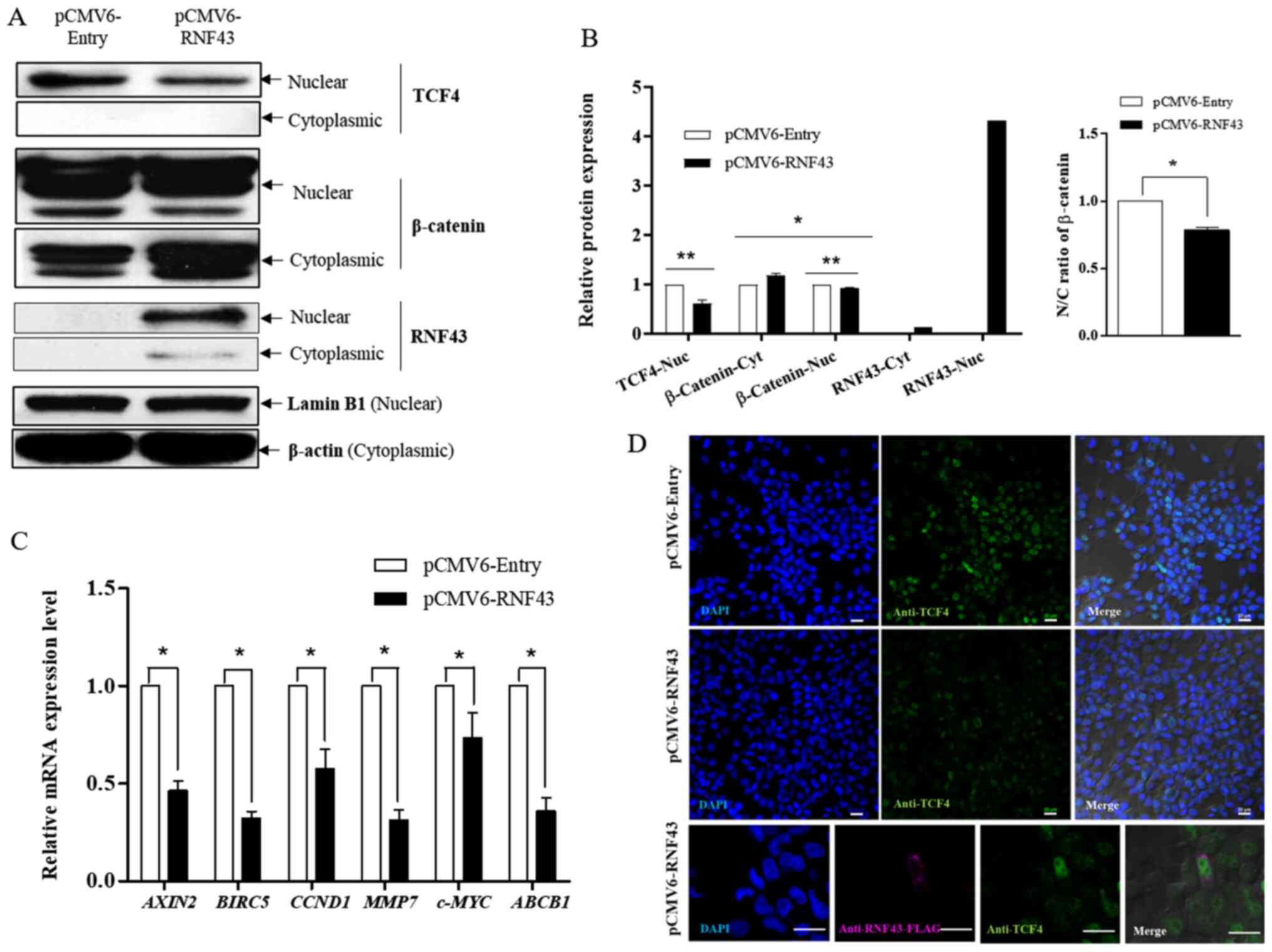 |
Figure 2.
RNF43 overexpression suppresses the Wnt/β-catenin-signaling pathway. KKU-213B cells were transfected with pCMV6-RNF43 or the control vector for 24 h. (A) The expression level of RNF43, β-catenin and TCF4 proteins in cytoplasmic and nuclear fractions were determined using western blot analysis and compared between the RNF43 overexpressing KKU-213B and control. (B) The ratio of N/C β-catenin and relative protein expression of RNF43, β-catenin and TCF4 were determined using ImageJ software and calculated by GraphPad Prism 5. (C) The relative mRNA expression of AXIN2, BIRC5, CCND1, MMP-7, c-MYC and ABCB1 were determined using reverse transcription-quantitative PCR and compared with the RNF43-overexpressing KKU-213B and control. (D) Immunofluorescent staining of RNF43-FLAG (magnification, ×600) and TCF4 (magnification, ×200) in RNF43-overexpressing KKU-213B cells. Scale bar, 20 µm. Values are expressed as the mean ± SEM of 3 independent experiments. *P<0.05; **P<0.001 vs. KKU-213B control. RNF43, ring finger protein 43; N, nuclear; C, cytoplasmic; TCF4, T-cell factor 4; BIRC5, baculoviral IAP Repeat Containing 5/surviving; Nuc, nuclear; cyt, cytoplasmic.
|
RNF43 overexpression increases 5-FU sensitivity
The WNT/β-catenin signalling pathway is now well-recognised as a critical pathway in tumour progression, including tumour growth, metastasis, and drug resistance (24,25). According to the results in Fig. 2C, RNF43 promoted the downregulation of known Wnt target genes [CCND1, c-MYC and ABCB1 (MDR1)] that are involved in cell proliferation and drug resistance. So as to determine whether RNF43 overexpression influences cell proliferation and drug responsiveness in KKU-213B cells, cell growth was assessed in the KKU-213B control and RNF43-overexpressing KKU-213B cells at 24, 48, and 72 h. The SRB assay demonstrated that RNF43-overexpressing KKU-213B cells have lower cell viability at 48 and 72 h compared with the control cells, suggesting that RNF43 overexpression reduces cell proliferation by decreasing cell proliferation markers (Figs. 2C and 3A).
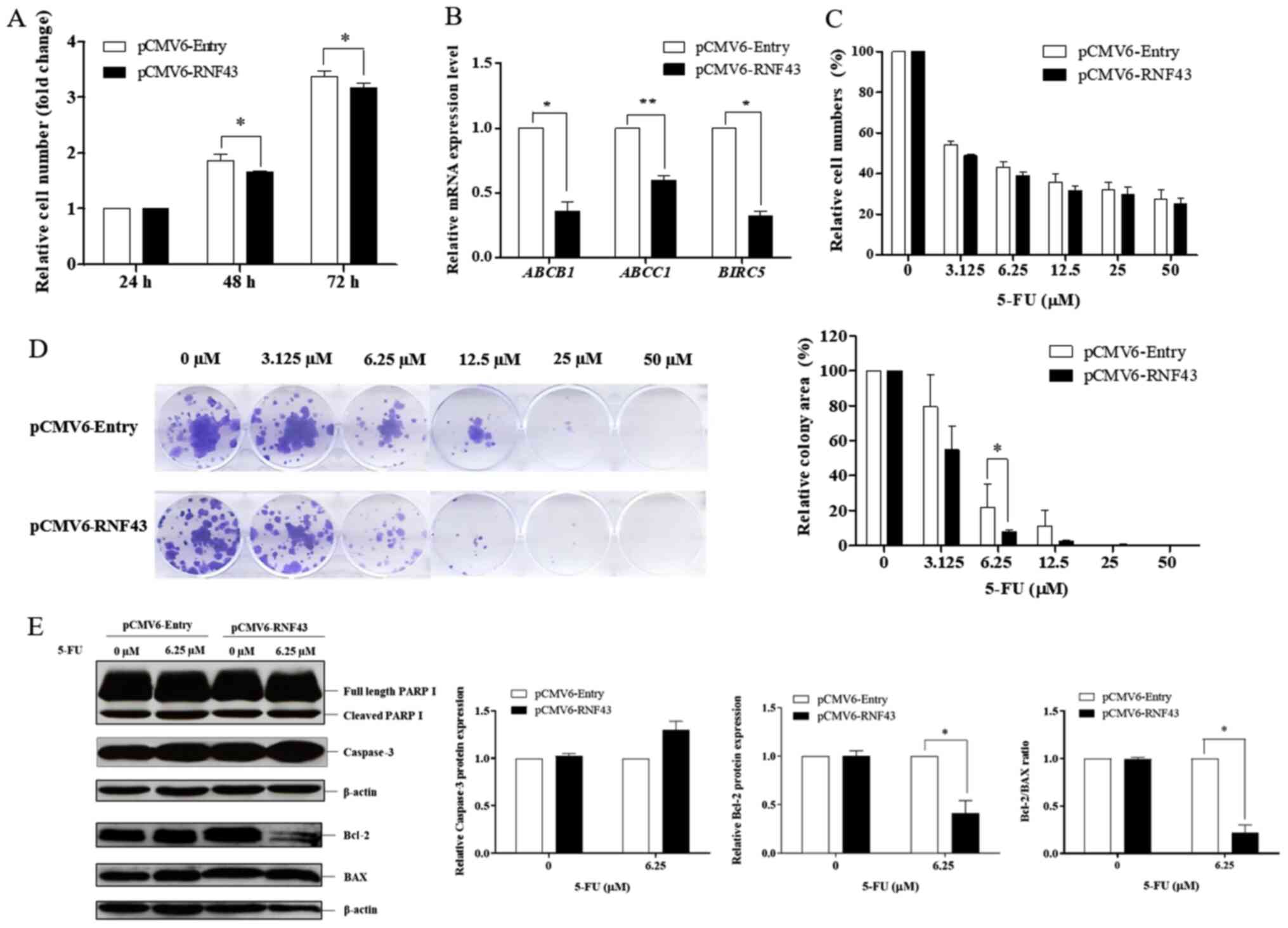 |
Figure 3.
RNF43 overexpression increases the sensitivity of cholangiocarcinoma cells to 5-FU. KKU-213B cells were transfected with pCMV6-RNF43 or the control vector for 24 h to determine cell growth and drug sensitivity. (A) The proliferative effect was determined at 24, 48 and 72 h. (B) The relative mRNA expression of BIRC5, ABCB1 and ABCC1 was determined using reverse transcription-quantitative PCR and compared between the RNF43-overexpressing KKU-213B cells and the control group. The RNF43-overexpressing KKU-213B cells and the control group were treated with 5-FU at various concentrations (0, 3.125, 6.25, 12.5, 25 and 50 µM), and (C) cell viability was determined at 72 h. (D) Colony formation was determined at 14 days. Cell viability was measured using sulforhodamine B assays, while colonies were stained using crystal violet. The intensity of the colonies was measured using ImageJ software, and quantification was performed using GraphPad Prism 5. (E) Apoptotic and anti-apoptotic proteins were determined using western blot analysis while the relative protein levels of cleaved PARP, caspase-3, Bcl-2 and Bax, and the Bcl-2/Bax ratio were determined using ImageJ software and quantified by GraphPad Prism 5. Values are expressed as the mean ± SEM of 3 independent experiments. *P<0.05; **P<0.001 vs. KKU-213B control. RNF43, ring finger protein 43; BIRC5, baculoviral IAP Repeat Containing 5/survivin; 5-FU, 5-fluorouracil.
|
Since downregulation of the ABC transporter superfamily (ABCB1 and ABCC1) and the anti-apoptotic gene BIRC5 were observed in the RNF43-overexpressing KKU-213B cells (Fig 3B), the chemotherapeutic response following RNF43 overexpression was explored. The RNF43-overexpressing KKU-213B cells showed a higher response to 5-FU in a dose-dependent manner compared with the control cells (Fig. 3C). There was, however, no change in the drug response during the cisplatin treatment (data not shown). The colony formation assay demonstrated that RNF43 overexpression significantly increased the sensitivity of 5-FU in the KKU-213B cells, especially at a concentration of 6.25 µM (Fig. 3D). The response to treatment with 5-FU at 6.25 µM in the RNF43-expressing KKU-213B cells was detectable via western blot analysis. The observation confirmed upregulation of caspase-3 and the downregulation of the anti-apoptotic protein, Bcl-2, at 5-FU concentration of 6.25 µM. A low ratio of Bcl-2/Bax was also observed in the RNF43-expressing KKU-213B cells treated with 5-FU (Fig. 3E). Taken together, these results demonstrate that RNF43 plays a role in chemotherapeutic drug susceptibility through the inhibition of the Wnt/β-catenin signalling pathway.
RNF43 overexpression inhibits cell migration
It is well accepted that dysregulation of the Wnt/β-catenin signalling pathway plays a critical role in tumour invasion and metastasis (26). Several metastatic-related genes, especially MMP-7, have been identified as targets of the WNT/β-catenin pathway (26). In the present study, suppression of MMP-7 expression was observed in the RNF43-expressing KKU-213B cells (Fig. 2C). The migration ability was investigated using the wound scratching assay, which indicated that RNF43-overexpressing KKU-213B cells have less ability to close a wound at 16, 20 and 24 h compared with the control cells (Fig. 4A and B). Similarly, the Transwell migration assay revealed that the RNF43-expressing KKU-213B cells have less migration capability at both 16 and 24 h (Fig. 4C). The effect of RNF43 overexpression on tumour metastasis was demonstrated by upregulation of E-cadherin (CDH1) and downregulation of N-cadherin (CDH2), MMP-2 and MMP-9 (Fig. 4D). These results confirm that RNF43 plays a role in inhibiting epithelial-mesenchymal transition by limiting the activity of Wnt/β-catenin signalling.
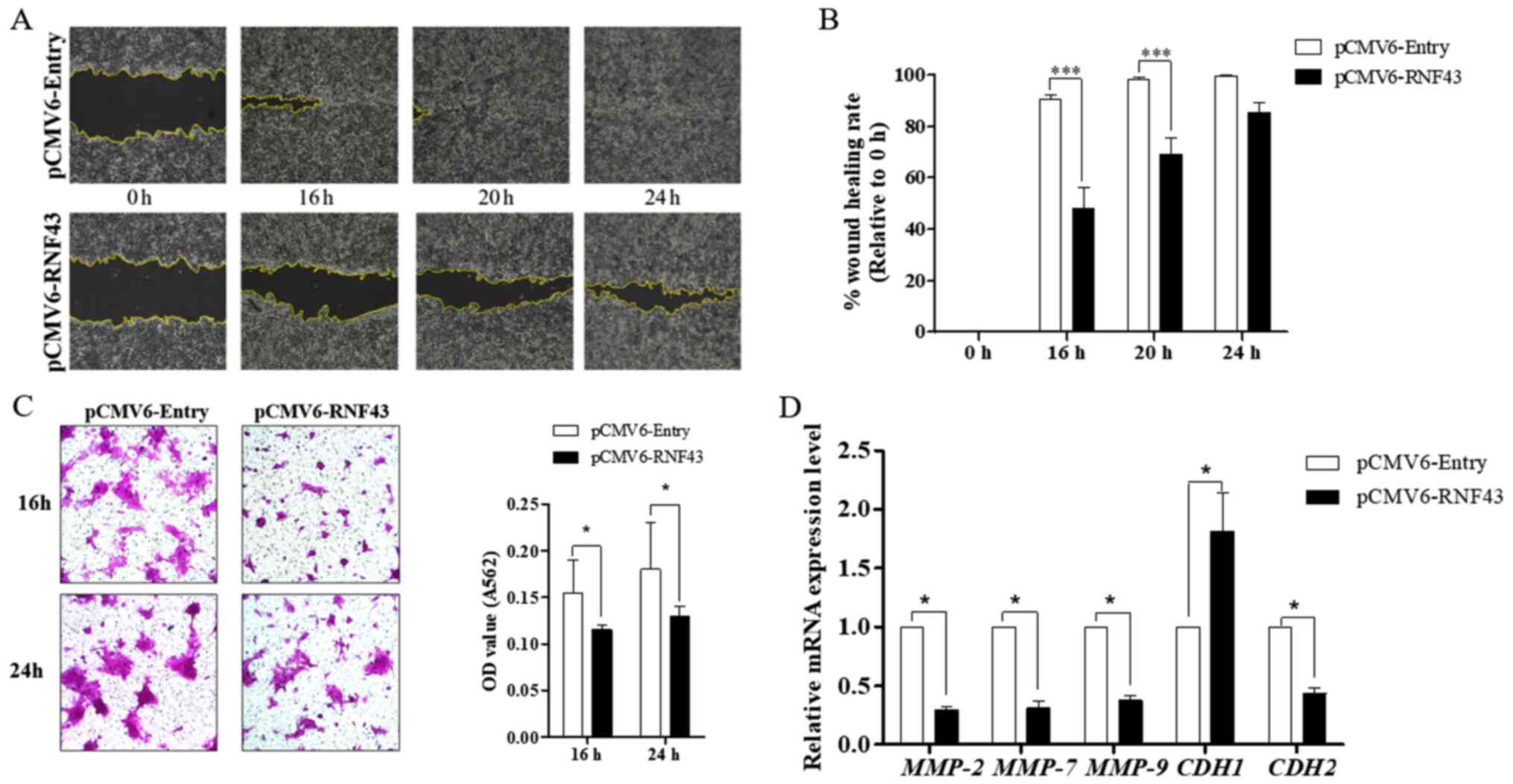 |
Figure 4.
RNF43 overexpression inhibits cholangiocarcinoma cell migration. KKU-213B cells were transfected with pCMV6-RNF43 or the control vector for 24 h. (A) The migratory effect by wound scratching was measured at 0, 16, 20 and 24 h (magnification, ×100). The wound closure area was measured using ImageJ and (B) quantification using GraphPad Prism 5. (C) The Transwell migration assay was measured at 16 and 24 h and the number of migratory cells (magnification, ×100) measured by SRB assay (A564). (D) The relative mRNA expression of MMP-2, −7 and −9, CDH1 and CDH2 were determined using reverse transcription-quantitative PCR and calculated using the 2−ΔΔCq method. Values are expressed as the mean ± SEM of 3 independent experiments. *P<0.05 and ***P<0.0001 vs. KKU-213B control. RNF43, ring finger protein 43;
|
Discussion
Inactivating mutation of RNF43 is reported in several types of cancers, including CCA. Patients with CCA with the inactivating RNF43 mutation and/or downregulation of RNF43 expression exhibit less favourable survival outcomes, suggesting that RNF43 may function as a tumour suppressor gene (10,19). Despite its potential role as a tumour suppressor gene in various cancer types, the biological function of RNF43 in CCA has never been thoroughly explored. Our previous study on CCA tissues reported that low RNF43 expression was localised in the nucleus (19). The current study confirmed a low expression of RNF43 at both the mRNA and protein levels in all CCA cell lines. This observation may have been influenced by the single nucleotide polymorphism in RNF43, particularly SNP rs2257205. A significant association has been observed between SNP rs2257205 in RNF43 genes and reduced RNF43 mRNA expression. Therefore, the RNF43 somatic mutation and epigenetic alteration by miR-550a-5p have been observed to reduce RNF43 expression (19). Exogenous expression of RNF43 in CCA cell lines also predominates in the nuclear fraction, and two studies have demonstrated that RNF43 functions as an inhibitor of the Wnt/β-catenin pathway by reducing nuclear TCF expression and sequestering TCF4 to the nuclear membrane (15,17). The current observations of CCA cell lines confirms that RNF43 overexpression attenuates β-catenin nuclear translocation, thereby reducing nuclear TCF4 expression.
Additionally, the localization of TCF4 was observed at the nuclear membrane, where it colocalised with RNF43. These findings support the hypothesis that exogenous RNF43 negatively regulates the Wnt/β-catenin signalling pathway through the repression of TCF4. The inhibitory effects of RNF43 on the Wnt/β-catenin signalling pathway also suppresses the transcriptional activity of six known Wnt target genes (including AXIN2, BIRC5, CCND1, MMP-7, c-MYC and ABCB1). Downregulation of the essential Wnt/β-catenin signalling target gene AXIN2 further corroborates the involvement of RNF43 in inhibiting the Wnt/β-catenin signalling. AXIN2 is a negative regulator of Wnt/β-catenin signalling, typically induced after vigorous transduction of the Wnt-signalling cascade (27,28). Although a significant decline in the c-MYC level has been observed following RNF43 overexpression, the downregulation of c-MYC was less evident than other known Wnt target genes. Thus, it was concluded that c-MYC is not specifically controlled by Wnt/β-catenin signalling, and is potentially regulated by different signalling mechanisms and transcription factors, especially the nuclear factor from activated T cells (NFAT) (29). Other than the six known Wnt target genes, the upregulation of CDH1 and downregulation of MMP2, MMP9 and CDH2 were observed after RNF43 overexpression. A possible explanation is that Twist and Snail are master regulators of EMT that repress the expression of CDH1 (E-cadherin) and activate expression of the entire suite of EMT-related genes, including CDH2, MMP2 and MMP9 (30). The present findings suggest that RNF43 has a role in inhibiting the Wnt signalling pathway, which may also suppress the transcription of other Wnt target genes such as the transcription factors Twist and Snail.
Given that the six known Wnt target genes are involved in tumour growth, metastasis and drug resistance, the functional role of RNF43 in mediating CCA tumour progression was investigated. Functional studies demonstrated that RNF43 overexpression reduced cell growth and 5-FU resistance and was correlated with downregulation of both the cell cycle genes (CCND1), and the chemotherapy resistance genes (ABCB1, ABCC1 and BIRC5), and upregulation of caspase-3. The present findings are consistent with studies on gastric cancer and colorectal cancer. The loss of RNF43 functions contributes to stemness properties (including drug resistance and tumorigenicity) by dampening the activation of the DNA damage response (17,18,31). RNF43 has an important role in the DNA damage response by ubiquitinating γH2AX after exposure to DNA damage-inducing chemotherapy. The increase of its expression may confer susceptibility to the DNA damage-induced apoptosis (31). Notwithstanding, changes in PARP expression were not observed in the present study perhaps because of an insufficient concentration of 5-FU that would otherwise promote a high degree of DNA damage-induced PARP activation in the CCA cells, or PARP was involved in the cellular response to single- and double-strand break(s) rather than DNA misincorporation. Therefore, the effect of RNF43 on cell proliferation and chemotherapeutic drug susceptibility through stemness properties and DNA damage response in CCA requires further study in the future.
CCA is a Wnt-high tumour in which the Wnt/β-catenin signalling pathway serves a critical role in both CCA growth and metastasis (11,13,32). RNF43 has also been reported to inhibit tumour migration and invasion in colorectal cancer (33). In the present study, RNF43 overexpression negatively regulated the Wnt/β-catenin signalling pathway, thereby suppressing the transcriptional level of MMP-7. MMP-7 has been identified as a target of the Wnt/β-catenin pathway and is highly expressed in metastatic tumours (26,27). The present study demonstrated that RNF43 overexpression significantly inhibited the migration capability of CCA cell lines through attenuation of epithelial-mesenchymal transition features via the Wnt/β-catenin signalling pathway. This finding highlights the importance of RNF43 as having a potentially significant role in tumour invasion and migration in CCA. Taken together, the current findings indicate that RNF43 has a tumour suppressive function in CCA, and these functions are implicated in all steps of tumour development and the progression of CCA.
Dysregulation of Wnt/β-catenin and PI3K/AKT signalling pathways are common in many cancer types, including CCA. The cross-talk between the Wnt/β-catenin and PI3K/AKT pathways was documented to drive tumor development and progression and resistance to cancer therapy (34,35). In the present study, whether RNF43 could influence PI3K/AKT signaling pathway was investigated. The expression level of phosphorylated Akt was examined in RNF43-overexpressing cells comparing to control cells. There was no difference in the expression of phosphorylated Akt following RNF43 overexpression (data not shown).
It should be noted that the RNF43 attenuation of the Wnt/β-catenin signalling pathway resulting in decreases in the progression of CCA was demonstrated using an overexpression model. Demonstrating the reverse effect using RNF43 knockdown would provide further supporting evidence. It should be noted that basal mRNA expression of RNF43 in all CCA cell lines used was detected at the cycle threshold value (Cq) >33 by RT-qPCR, and the basal protein expression of RNF43 was also undetectable. The implication is that demonstrating the reverse effect of RNF43 suppression may be problematic due to the low/absent RNF43 mRNA and protein expression in the control and RNF43-suppressing cells. Therefore, further investigations using rescue experiment will be important to confirm the effect of RNF43 in CCA.
The limitations of the present study still need to be addressed. Firstly, KKU-213A cell lines have unique metastatic characteristics and are more aggressive than KKU-213B. Therefore, KKU-213B cells were used to demonstrate the various effects of RNF43 overexpression in CCA. However, the verification of the results in other cell lines and in vivo experiments are required in further investigations. Secondly, gemcitabine is one of the most common drugs in CCA treatment. Future studies on gemcitabine sensitivity will be important to provide the significant role of RNF43 on chemotherapeutic sensitivity in CCA. Thirdly, the presence of RNF43 enhanced 5-FU-induced CCA death via the increased expression of caspase-3 protein. However, the possible induction of apoptotic process remains unclear, due to the lack of a direct apoptosis assay, such as TUNEL staining. Fourthly, the cell-free area was created after serum starvation and the cell migration was directly monitored within 24 h under 10% FBS as previously described (36,37). However, the standard wound healing assay is performed under serum-starved media with 0–1% FBS to avoid possible cell proliferation intervention (38). Nevertheless, the doubling time of KKU-213B is 24.5±0.2 h (21) and the result in Fig. 3A demonstrated no significant difference in cell proliferation within 24 h between RNF43-overexpressing KKU-213B cells and the control cells. Those findings suggest, cell proliferation has less effect on the wound healing assay under 10% FBS, albeit the standard wound healing assay with serum-reduced media is required to confirm the present findings.
In summary, the present study demonstrated the significant roles of RNF43 in mediating the inhibition of the Wnt/β-catenin signalling pathway in CCA. Firstly, RNF43 blocks β-catenin translocation followed by downregulation of the Wnt/β-catenin target genes, which are involved in the progression of CCA. Subsequently, overexpression of RNF43 attenuated CCA progression by decreasing CCA cell growth, migration and drug resistance. The expression of RNF43 is thus essential to preventing further aberrant activation of Wnt/β-catenin signalling that contributes to aggressive CCA phenotypes.
Acknowledgements
The authors would like to thank Professor Sopit Wongkham (Khon Kaen University) for providing the CCA cell lines and Mr Bryan Roderick Hamman for assistance with the English language presentation of the manuscript.
Funding
The present study was supported by the Young Research Grants, Thailand Research Fund (grant no. TRG5680041), Vithedbandit scholarship and SUT Research Unit grants.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Authors' contributions
CT conceived and designed the present study. NSP and CT performed the experiments. CT, NSP, KT, PC and JK analysed the data. All authors read and approved the final manuscript. CT and NSP confirm the authenticity of all the raw data.
Ethics approval and consent to participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
|
1
|
Bergquist A and von Seth E: Epidemiology of cholangiocarcinoma. Best Pract Res Clin Gastroenterol. 29:221–232. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Banales JM, Marin JJ, Lamarca A, Rodrigues PM, Khan SA, Roberts LR, Cardinale V, Carpino G, Andersen JB, Braconi C, et al: Cholangiocarcinoma 2020: The next horizon in mechanisms and management. Nat Rev Gastroenterol Hepatol. 17:557–588. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bertuccio P, Malvezzi M, Carioli G, Hashim D, Boffetta P, El-Serag HB, La Vecchia C and Negri E: Global trends in mortality from intrahepatic and extrahepatic cholangiocarcinoma. J Hepatol. 71:104–114. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pratap P, Raza ST and Pratap S: Cholangiocarcinoma: Etiology, pathogenesis, diagnosis, and management. Bioactive Components, Diet and Medical Treatment in Cancer Prevention. 201–213. 2018. View Article : Google Scholar
|
|
5
|
Patel T: Cholangiocarcinoma - controversies and challenges. Nat Rev Gastroenterol Hepatol. 8:189–200. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Martinez-Becerra P, Vaquero J, Romero MR, Lozano E, Anadon C, Macias RI, Serrano MA, Grañé-Boladeras N, Muñoz-Bellvis L, Alvarez L, et al: No correlation between the expression of FXR and genes involved in multidrug resistance phenotype of primary liver tumors. Mol Pharm. 9:1693–1704. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Razumilava N and Gores GJ: Cholangiocarcinoma. Lancet. 383:2168–2179. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chong DQ and Zhu AX: The landscape of targeted therapies for cholangiocarcinoma: Current status and emerging targets. Oncotarget. 7:46750–46767. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yao D, Kunam VK and Li X: A review of the clinical diagnosis and therapy of cholangiocarcinoma. J Int Med Res. 42:3–16. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ong CK, Subimerb C, Pairojkul C, Wongkham S, Cutcutache I, Yu W, McPherson JR, Allen GE, Ng CC, Wong BH, et al: Exome sequencing of liver fluke-associated cholangiocarcinoma. Nat Genet. 44:690–693. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jusakul A, Cutcutache I, Yong CH, Lim JQ, Huang MN, Padmanabhan N, Nellore V, Kongpetch S, Ng AW, Ng LM, et al: Whole-genome and epigenomic landscapes of etiologically distinct subtypes of cholangiocarcinoma. Cancer Discov. 7:1116–1135. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Loilome W, Bungkanjana P, Techasen A, Namwat N, Yongvanit P, Puapairoj A, Khuntikeo N and Riggins GJ: Activated macrophages promote Wnt/β-catenin signaling in cholangiocarcinoma cells. Tumour Biol. 35:5357–5367. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Boulter L, Guest RV, Kendall TJ, Wilson DH, Wojtacha D, Robson AJ, Ridgway RA, Samuel K, Van Rooijen N, Barry ST, et al: WNT signaling drives cholangiocarcinoma growth and can be pharmacologically inhibited. J Clin Invest. 125:1269–1285. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tsukiyama T, Fukui A, Terai S, Fujioka Y, Shinada K, Takahashi H, Yamaguchi TP, Ohba Y and Hatakeyama S: Molecular role of RNF43 in canonical and noncanonical Wnt signaling. Mol Cell Biol. 35:2007–2023. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Loregger A, Grandl M, Mejías-Luque R, Allgäuer M, Degenhart K, Haselmann V, Oikonomou C, Hatzis P, Janssen KP, Nitsche U, et al: The E3 ligase RNF43 inhibits Wnt signaling downstream of mutated β-catenin by sequestering TCF4 to the nuclear membrane. Sci Signal. 8:ra902015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jiang X, Hao HX, Growney JD, Woolfenden S, Bottiglio C, Ng N, Lu B, Hsieh MH, Bagdasarian L, Meyer R, et al: Inactivating mutations of RNF43 confer Wnt dependency in pancreatic ductal adenocarcinoma. Proc Natl Acad Sci USA. 110:12649–12654. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gao Y, Cai A, Xi H, Li J, Xu W, Zhang Y, Zhang K, Cui J, Wu X, Wei B, et al: Ring finger protein 43 associates with gastric cancer progression and attenuates the stemness of gastric cancer stem-like cells via the Wnt-β/catenin signaling pathway. Stem Cell Res Ther. 8:982017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Neumeyer V, Brutau-Abia A, Allgäuer M, Pfarr N, Weichert W, Falkeis-Veits C, Kremmer E, Vieth M, Gerhard M and Mejías-Luque R: Loss of RNF43 function contributes to gastric carcinogenesis by impairing DNA damage response. Cell Mol Gastroenterol Hepatol. 11:1071–1094. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Talabnin C, Janthavon P, Thongsom S, Suginta W, Talabnin K and Wongkham S: Ring finger protein 43 expression is associated with genetic alteration status and poor prognosis among patients with intrahepatic cholangiocarcinoma. Hum Pathol. 52:47–54. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sripa B, Leungwattanawanit S, Nitta T, Wongkham C, Bhudhisawasdi V, Puapairoj A, Sripa C and Miwa M: Establishment and characterization of an opisthorchiasis-associated cholangiocarcinoma cell line (KKU-100). World J Gastroenterol. 11:3392–3397. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sripa B, Seubwai W, Vaeteewoottacharn K, Sawanyawisuth K, Silsirivanit A, Kaewkong W, Muisuk K, Dana P, Phoomak C, Lert-Itthiporn W, et al: Functional and genetic characterization of three cell lines derived from a single tumor of an opisthorchis viverrini-associated cholangiocarcinoma patient. Hum Cell. 33:695–708. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Talabnin C, Talabnin K and Wongkham S: Enhancement of piperlongumine chemosensitivity by silencing heme oxygenase-1 expression in cholangiocarcinoma cell lines. Oncol Lett. 20:2483–2492. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shen DY, Zhang W, Zeng X and Liu CQ: Inhibition of Wnt/β-catenin signaling downregulates P-glycoprotein and reverses multi-drug resistance of cholangiocarcinoma. Cancer Sci. 104:1303–1308. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang Y and Wang X: Targeting the Wnt/β-catenin signaling pathway in cancer. J Hematol Oncol. 13:1652020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dey N, Young B, Abramovitz M, Bouzyk M, Barwick B, De P and Leyland-Jones B: Differential activation of Wnt-β-catenin pathway in triple negative breast cancer increases MMP7 in a PTEN dependent manner. Burchell JM: PLoS One. 8:e774252013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wu ZQ, Brabletz T, Fearon E, Willis AL, Hu CY, Li XY and Weiss SJ: Canonical Wnt suppressor, Axin2, promotes colon carcinoma oncogenic activity. Proc Natl Acad Sci USA. 109:11312–11317. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Takahashi N, Yamaguchi K, Ikenoue T, Fujii T and Furukawa Y: Identification of two Wnt-responsive elements in the intron of RING finger protein 43 (RNF43) gene. PLoS One. 9:e865822014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mognol GP, de Araujo-Souza PS, Robbs BK, Teixeira LK and Viola JP: Transcriptional regulation of the c-Myc promoter by NFAT1 involves negative and positive NFAT-responsive elements. Cell Cycle. 11:1014–1028. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ghahhari NM and Babashah S: Interplay between microRNAs and WNT/β-catenin signalling pathway regulates epithelial-mesenchymal transition in cancer. Eur J Cancer. 51:1638–1649. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Neumeyer V, Grandl M, Dietl A, Brutau-Abia A, Allgäuer M, Kalali B, Zhang Y, Pan KF, Steiger K, Vieth M, et al: Loss of endogenous RNF43 function enhances proliferation and tumour growth of intestinal and gastric cells. Carcinogenesis. 40:551–559. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Noll AT, Cramer T, Olde Damink SW and Schaap FG: Cholangiocarcinoma, gone without the Wnt? World J Hepatol. 8:1093–1096. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang G, Fu Y, Yang X, Luo X, Wang J, Gong J and Hu J: Brg-1 targeting of novel miR550a-5p/RNF43/Wnt signaling axis regulates colorectal cancer metastasis. Oncogene. 35:651–661. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Perry JM, Tao F, Roy A, Lin T, He XC, Chen S, Lu X, Nemechek J, Ruan L, Yu X, et al: Overcoming Wnt-β-catenin dependent anticancer therapy resistance in leukaemia stem cells. Nat Cell Biol. 22:689–700. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Toh TB, Lim JJ, Hooi L, Rashid MBMA and Chow EK: Targeting Jak/Stat pathway as a therapeutic strategy against SP/CD44+ tumorigenic cells in Akt/β-catenin-driven hepatocellular carcinoma. J Hepatol. 72:104–118. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sribenja S, Sawanyawisuth K, Kraiklang R, Wongkham C, Vaeteewoottacharn K, Obchoei S, Yao Q, Wongkham S and Chen C: Suppression of thymosin β10 increases cell migration and metastasis of cholangiocarcinoma. BMC Cancer. 13:4302013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Di Maira G, Gentilini A, Pastore M, Caligiuri A, Piombanti B, Raggi C, Rovida E, Lewinska M, Andersen JB, Borgo C, et al: The protein kinase CK2 contributes to the malignant phenotype of cholangiocarcinoma cells. Oncogenesis. 8:612019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Park DD, Phoomak C, Xu G, Olney LP, Tran KA, Park SS, Haigh NE, Luxardi G, Lert-Itthiporn W, Shimoda M, et al: Metastasis of cholangiocarcinoma is promoted by extended high-mannose glycans. Proc Natl Acad Sci USA. 117:7633–7644. 2020. View Article : Google Scholar : PubMed/NCBI
|














