Introduction
The success of immune checkpoint inhibitors (ICIs)
targeting programmed cell death-1 (PD-1)/programmed cell death-1
ligands (PD-L1) and cytotoxic T lymphocyte-associated antigen-4
(CTLA-4) has highlighted the important role of antitumour immunity
in cancer treatment (1). In recent
years, there has been increased interest in studies on antitumour
immunity for effective cancer treatment (2). Clinically, a combination strategy of
ICIs and other cancer therapies that induce local and systemic
immune responses achieve an improved response rate compared with
ICIs alone (3,4).
Previous studies have suggested stimulation of local
and systemic immune responses after radiotherapy; for example,
radiotherapy promotes the induction of damage-associated molecular
patterns and tumour-associated antigens as well as the release of
exosomes containing DNA fragments that activate dendritic cells
(5–7). In addition, radiotherapy up-regulates
the expression of human leukocyte antigen, a central component of
specific immune stimulation (8).
Increased serum levels of interferon-β and early dynamic changes of
blood T cell clones have been observed in patients who received
radiotherapy and demonstrated greater systemic immune responses
(9), suggesting that radiotherapy
can strongly activate antitumour immunity. Furthermore, DNA damage
signalling after irradiation also stimulates innate immune
responses via cyclic GMP-AMP synthase (cGAS)/stimulator of IFN
genes (STING)- or the melanoma differentiation-associated gene
5/retinoic acid-inducible gene I (RIG-I)/mitochondrial
antiviral-signalling protein pathways. These signals promote the
release of interferons (10–13).
Thus, although radiotherapy can be a promising tool to stimulate
the antitumour immune response, positive stimulation may not fully
achieve antitumour effects due to the undesired immune-suppressive
signal, for example, PD-L1 up-regulation (14); especially if radiotherapy
stimulates the immune-suppressive signal. Thus, it is important to
understand the regulation of immune response in the context of its
ligand expression in each patient to effectively implement this
combination therapy.
To date, multiple pathways regarding the regulation
of PD-L1 expression in tumours have been suggested. Microsatellite
instability induced by defects in mismatch repair (15,16)
and PD-L1 expression (17) have
been proposed as predictive markers for the efficacy of
anti-PD-1/PD-L1 antibodies monotherapy. In the context of DNA
damage-dependent regulation of PD-L1 expression, our previous study
indicated that PD-L1 expression is up-regulated at the
transcriptional level (14). This
up-regulation was dependent on STAT1/3-IRF1 after the activation of
ataxia telangiectasia mutated/ataxia telangiectasia and
Rad3-related (ATR)/checkpoint kinase 1 (Chk1) (14). Our previous study also demonstrated
in U2OS cells that PD-L1 up-regulation is enhanced in cells that
are deficient in Ku80, a central regulator of non-homologous end
joining (NHEJ).
The present study examined the correlation of mRNA
expression between PD-L1 and NHEJ factors using The Cancer Genome
Atlas (TCGA) dataset analysis. Ku80 mRNA expression was
negatively correlated with PD-L1 mRNA expression in all the
selected tissues. Moreover, the current study aimed to examine the
relationship between Ku80 and PD-L1 expression in patients who
received radiotherapy.
Materials and methods
TCGA analysis
Normalised RNA sequences of Ku80 and
PD-L1 expression status data provided by the TCGA project
were downloaded from the Genomic Data Commons Data Portal
(https://portal.gdc.cancer.gov). Datasets
for colon adenocarcinoma (COAD), breast invasive carcinoma (BRCA),
skin cutaneous melanoma (SKCM), lung adenocarcinoma (LUAD), head
and neck squamous cell carcinoma (HNSC), uterine corpus endometrial
carcinoma (UCEC) and cervical squamous cell carcinoma and
endocervical adenocarcinoma (CESC) were analysed in the present
study (18–24). Partial correlations and statistical
significance presented as volcano plots were calculated using
non-parametric Spearman's formula, shown as the coefficient rho (r)
by GraphPad Prism v9.0 software (GraphPad Software, Inc.) and the
XLSTAT 2021.1.1 add-in feature in Microsoft Excel v16.46 (Microsoft
Corporation). P<0.05 was considered to indicate a statistically
significant difference. The total sample numbers for each dataset
were 480 (COAD), 109 (BRCA), 470 (SKCM), 1,086 (LUAD), 502 (HNSC),
1,607 (UCEC) and 306 (CESC).
Patients and tumour
characteristics
A total of 75 patients with cervical squamous cell
carcinoma (median age, 62 years; range, 32–87 years) who met the
following criteria were eligible for the present retrospective
study: i) Pathologically-confirmed newly diagnosed cervical cancer
by pathologists who were not associated with the present study; ii)
treated with definitive platinum-based chemoradiotherapy (CRT
group) or radiotherapy alone (RT group) at Gunma University
Hospital between August 2009 and November 2013; iii) staged as
IB1-IVA according to the FIGO classification 2008 (25); and iv) no previous exposure to
radiotherapy or cytotoxic agents. Patients with recurrence were
excluded from this study. Tumour samples from all patients before
radiotherapy (pre-RT) and after 10 Gy of radiotherapy (10 Gy-RT)
were used for pathological analysis and immunohistochemical
staining. The time between the radiotherapy start date and the
biopsy specimen date was 5–11 days. Most samples were collected on
days 8–9 (89% in the RT group and 95% in the CRT group). The time
between the 10 Gy-RT day and the specimen biopsy day ranged from
0–4 days. The majority of samples (75%) were biopsied on the same
day as that of receiving 10 Gy-RT. Patient characteristics were
recorded according to tumour stage [stages IB, II, III and IV
according to FIGO classification 2008 (25)] and age (Table I). The present study protocol was
approved by the Institutional Review Board for clinical trials of
Gunma University (Maebashi, Japan; approval no. HS2020-015). All
patients provided their informed consent to participate in the
study using the opt-out approach by public notice on the internet
site of the Institutional Review Board for clinical trials of Gunma
University.
 | Table I.Characteristics of patients (n=75)
with cervical squamous cell carcinoma. |
Table I.
Characteristics of patients (n=75)
with cervical squamous cell carcinoma.
|
Characteristics | Value |
|---|
| Observation period,
months |
|
| Median
(range) | 63 (8–120) |
| Age, years |
|
| Median
(range) | 62 (32–87) |
| Treatment, n
(%) |
|
| RT
alone | 27 (36%) |
|
Concurrent CRT | 48 (64%) |
| FIGO stage 2008, n
(%) |
|
| IB | 11 (15%) |
| II | 31 (41%) |
|
III | 31 (41%) |
|
IVA | 2 (3%) |
| Lymph node
metastasis in pelvis, n (%) |
|
|
Positive | 36 (48%) |
|
Negative | 39 (52%) |
| Para-aortic lymph
node metastasis, n (%) |
|
|
Positive | 6 (8%) |
|
Negative | 69 (92%) |
Treatment of patients
All patients received definitive radiotherapy in
combination with external body radiotherapy (EBRT) and intracavity
brachytherapy (ICBT). EBRT typically used 10 MV X-rays with 4-field
irradiation. The most common EBRT dose and fractionation was 50 Gy
in 25 fractions (2 Gy/fraction, once per day, five times weekly).
EBRT was performed using a combination of total pelvic irradiation
(20–40 Gy) followed by a central shielded irradiation at a 3-cm
width. The pelvic field was expanded to the metastatic area in
patients with paraaortic lymph node metastases. Patients with lymph
node metastases received an additional boost of 6–8 Gy in three to
four fractions. ICBT was performed once a week with concurrent
central shielding EBRT. EBRT was not performed on the day of ICBT
administration. Three-dimensional image-guided brachytherapy was
performed on all patients using a high-dose-rate 192Ir
remote after loading system (microSelectron® Digital;
Elekta Instrument AB). The prescription dose for each ICBT was
determined to cover 90% of the high-risk clinical target volume,
using a total dose of 6 Gy. Bulky and/or asymmetrical tumours were
treated using additional interstitial irradiation. ICBT was
typically performed four times. Concurrent chemotherapy was given
to 64.0% (48/75) of the patients. According to the Japan Society of
Gynecologic Oncology (JSGO) Guidelines 2017 for the Treatment of
Uterine Cervical Cancer, cisplatin was usually administered weekly
at a dose of 40 mg/m2 (26).
Immunohistochemical staining for PD-L1
and Ku80
PD-L1 expression in tumour cells and Ku80 expression
in the nuclei of biopsy specimens excised from the cervical
squamous cell carcinoma pre-RT and 10 Gy-RT were evaluated using
immunohistochemical staining. Biopsied samples were fixed in 10%
buffered formalin for 24 h at room temperature and then dehydrated,
degreased and paraffin-embedded. Paraffin embedded sections (4-µm
thick) were dewaxed in xylene at room temperature and rehydrated
using a graded ethanol series. Endogenous peroxidase activity was
blocked using a 10-min incubation in 0.3% hydrogen peroxide. After
blocking by 10% goat normal serum (cat. no. 5425; Cell Signalling
Technology, Inc.) for PD-L1 and Ku80 sections with a 20 min
incubation at room temperature, the sections were incubated
overnight with primary antibodies at 4°C. The sections were
incubated with a commercially available biotin-streptavidin
immunoperoxidase kit [Histofine; SAB-PO (rabbit); cat. no. 424032;
Nichirei Biosciences, Inc.; Nichirei Corporation] for 20 min at
room temperature. Then, the sections were incubated for 5 min with
diaminobenzidine at room temperature. The following antibodies were
used: Monoclonal anti-PD-L1 antibody (1:100; clone E1L3N rabbit
IgG; cat. no. 13684; Cell Signalling Technology, Inc.) and
monoclonal anti-Ku80 antibody (1:200; clone C48E7, and rabbit IgG;
cat. no. 2180T; Cell Signalling Technology, Inc.). The quality of
the tumour samples was carefully evaluated and validated
independently by two pathologists who are co-authors of the present
study.
Quantification of PD-L1 and Ku80 in
immunohistochemical images
All immunostaining images were obtained using the
light microscope Leica DM4000 B (Leica Microsystems, GmbH) equipped
with a ×20 objective lens. Expression of cytosolic PD-L1 was
measured using public domain software ImageJ v1.53a (National
Institutes of Health) as follows. In 32-bit colour images, which
are commonly used in computers, each pixel has a red, blue and
green signal intensity from 0 to 255. Brown, which is composed of a
higher red than blue signal, represents PD-L1 labelled by antibody
in the immunohistochemical image. The nucleus is shown in blue and
the background is shown in white, which consists of high red, blue
and green signals. Therefore, brown is the only colour that has
much higher red compared with blue signal in the microscopy images.
The captured images were split into red, blue and green channels.
The PD-L1 signal was enhanced by subtracting the blue signal from
the red signal, taking advantage of the fact that all the
subtracted values are returned as zero when the subtracted value is
zero or less. The mean of the signal intensities in three areas of
the tumour tissues were quantified. PD-L1 change was calculated as
PD-L1 intensity (10 Gy-RT) subtracted by PD-L1 intensity (pre-RT).
Furthermore, the percentages of tumour cells with cell-surface
staining for PD-L1 were recorded and expressed as a tumour
proportion score (TPS). If the TPS was 1–50%, the sample was
classified as PD-L1 1+, whereas samples with TPS >50% were
classified as PD-L1 2+. The population of Ku80-positive cells were
measured using an opened source software QuPath (v0.1.2; Queen's
University Belfast; Northern Ireland). The tumour tissue area was
targeted to detect Ku80-positive cells and the Ku80 positivity was
quantified.
Cell culture, small interfering
(si)RNA-knockdown, flow cytometry and immunoblotting
HeLa cells were obtained from the American Type
Culture Collection and cultured in Eagle's Minimum Essential Medium
(FUJIFILM Wako Pure Chemical Corporation) with 10% foetal calf
serum (Sigma Aldrich; Merck KGaA) at 37°C. siRNA transfection was
performed using DharmaFECT (GE Healthcare Dharmacon, Inc.). siRNA
was added to suspended HeLa cells after trypsinisation (the final
concentration of siRNA is 16 nM). Non-targeting siRNA was used as
negative control. The siRNA oligonucleotides used are listed in
Table SI. Cells were incubated
after transfection for 48 h at 37°C before 10 Gy of irradiation.
X-ray irradiation was performed using an MX-160Labo irradiator (160
kVp; 1.06 Gy/min; mediXtec Corporation).
For immunoblotting, cells were harvested at 48 h
after transfection with ×1 Sample Buffer [50-mM Tris, 2% sodium
dodecyl sulphate, 6% glycerol, 1% (w/w) 3-mercapto-1,2-propanediol
and 0.008% bromophenol blue] following PBS wash. Harvested samples
were boiled at 95°C for 5 min and sonicated using a Q55 Sonicator
Ultrasonic Homogeniser (QSonica LLC; power amplification 15% for 5
sec, twice) (27). The number of
cells each condition was confirmed to align the protein amount as
the previous study. Subsequently, samples (10 µl/lane) were loaded
onto 4–20% Mini-PROTEAN TGX Precast Protein gels (Bio-Rad
Laboratories, Inc.) and were run in Mini-PROTEAN Tetra Vertical
Electrophoresis Cell (Bio-Rad Laboratories, Inc.) at 200 V for 30
min. They were then transferred onto a BioTrace NT Nitrocellulose
Transfer Membrane (Pall Life Sciences) using a Mini-PROTEAN Tetra
Vertical Electrophoresis Cell (Bio-Rad Laboratories, Inc.) at 100 V
for 1 h. The membrane was incubated overnight at room temperature
with anti-Ku80 (Rabbit mAb; 1:2,000; cat. no. 2180; Cell Signaling
Technology, Inc.) and anti-Actin antibodies (mouse mAb, 1:5,000;
cat. no. 3700; Cell Signaling Technology, Inc.), which served as
the primary antibodies. After the membranes were washed trice with
×1 Tris-buffered saline (TBST; 0.05% Tween 20), they were incubated
with goat anti-rabbit IgG, horseradish peroxidase-conjugated
antibodies (1:4,000; cat. no. 7074; Cell Signalling Technology,
Inc.) or anti-mouse IgG, horse anti-mouse horseradish
peroxidase-conjugated antibodies (1:4,000; cat. no. 7076; Cell
Signalling Technology, Inc.) at room temperature for 1 h. After
being washed trice with ×1 TBST, the membrane was reacted with ECL
Western Blotting Detection Reagent (GE Healthcare), and chemical
luminescence was detected using a LAS-600 Bioimaging Analyser
System (Azure Biosystems, Inc.). The signal intensities of Ku80 and
Actin were measured using ImageJ. The signal intensity of Ku80 was
normalized to that of Actin; subsequently, the Ku80 signal
intensity ratio was normalized to the Ku80 signal intensity in HeLa
cells transfected with siControl.
Cells were exposed to 10 Gy of irradiation then
incubated for 48 h prior to flow cytometry analysis. Harvested
cells were washed and collected with ice-cold 1 mM
EDTA-phosphate-buffered saline and then stained with anti-PD-L1
antibodies (clone E1L3N rabbit IgG; cat. no. 13684; Cell Signalling
Technology, Inc.) for 20 min on ice. The fluorochrome used was APC.
Dead cells detected by propidium iodide (Sigma-Aldrich; Merck KGaA)
were excluded from the analysis. Flow cytometry analysis was
performed on an Attune NxT Flow Cytometer (Thermo Fisher
Scientific, Inc.). Mean fluorescence intensity (MFI) of the PD-L1
was calculated as: MFI (PD-L1)-MFI (isotype control).
Statistical analysis
The correlation between Ku80 positivity and PD-L1
signal intensity was evaluated using Spearman's rank correlation
coefficient. Clinical outcomes were calculated using the
Kaplan-Meier method, and statistically significant differences were
confirmed using the log-rank test. Mann-Whitney U test was used to
compare Ku80 positivity in pre-RT samples and classification of
PD-L1 expression. MFI in flow cytometry analysis was compared using
unpaired Student's two-tailed t-test. Results were shown as mean ±
error bars, which represent the standard error of three samples in
the experiment. P<0.05 was considered to indicate a
statistically significant difference. All statistical analyses were
performed using SPSS (v24; IBM Corp.).
Results
TCGA analysis demonstrated negative
correlation between Ku80 and PD-L1
TCGA datasets were used in the present study to
investigate the relationship of gene expression between PD-L1 and
DNA double-strand break (DSB) repair factors in tumour cells. Our
previous study reported that tumour specimens harbouring mutations
in NHEJ genes (Ku70/80) exhibit greater PD-L1 expression after
irradiation in several cancer types (14). To determine whether the mRNA
expression of NHEJ factors demonstrated a correlation with
PD-L1 mRNA expression, TCGA analyses between central NHEJ
factors [Ku80, Ku70, DNA-dependent protein kinase catalytic
subunit (DNA-PKcs), X-ray repair cross-complementing protein
4 (XRCC4) and DNA ligase 4] and PD-L1 were performed
(Table II). Fig. 1 presents the results of the seven
cancer tissues, COAD, BRCA, SKCM, LUAD, HNSC, UCEC and CESC.
Spearman's rho was <0 for all cancer types in Ku80
(Fig. 1; Table II). Because the high proliferative
conditions in cancer cells can induce an unrepairable amount of DNA
replication-associated genotoxic stress, including DSBs (28), it is hypothesized that the result
of TCGA analysis reflects the situation in cells that are exposed
to exogenous DNA damage, such as ionising radiation. Therefore, the
present study investigated the correlation between Ku80 and PD-L1
in clinical specimens that received radiotherapy.
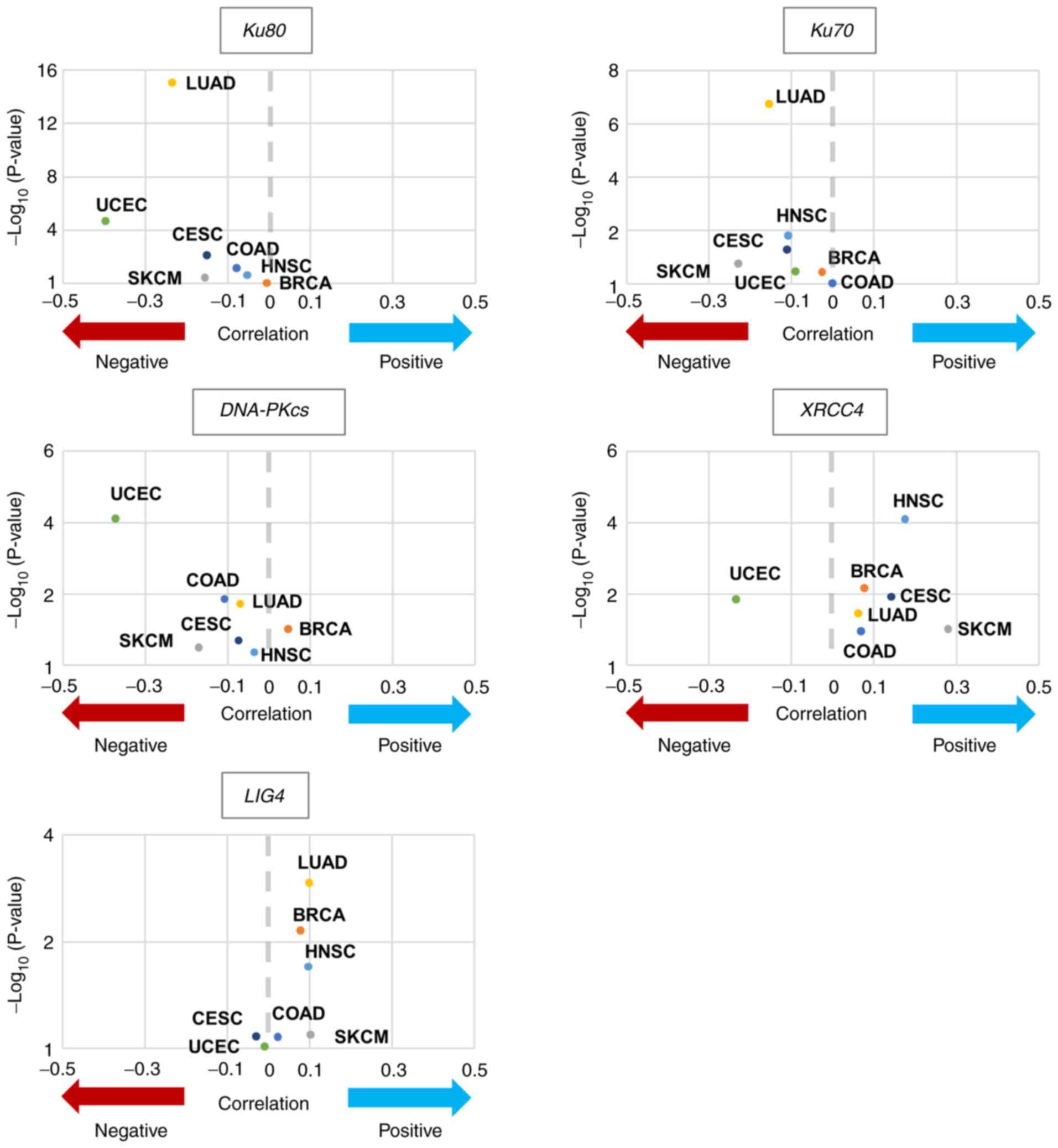 | Figure 1.Correlation between PD-L1 gene
expression and DNA double-strand break repair factors. Volcano
plots indicating the correlation between PD-L1 expression and
central non-homologous end joining factors expression levels.
Mutation statuses were provided by The Cancer Genome Atlas project
were downloaded from the Genomic Data Commons Data Portal. BRCA,
breast invasive carcinoma; CESC, cervical squamous cell carcinoma
and endocervical adenocarcinoma; COAD, colon adenocarcinoma; HNSC,
head and neck squamous cell carcinoma; LUAD, lung adenocarcinoma;
SKCM, skin cutaneous melanoma; UCEC, uterine corpus endometrial
carcinoma; DNA-PKcs, DNA-dependent protein kinase, catalytic
subunit; XRCC4, X-ray repair cross-complementing protein 4; LIG4,
DNA ligase 4. |
| Table II.TCGA analysis. |
Table II.
TCGA analysis.
| Gene | Study | Correlation | Approximation
formula | Spearman's rho
(r) | P-value |
|---|
| Ku80 | COAD | Negative |
y=−0.001536×+32430 | -0.079 | 0.070 |
|
| BRCA | Negative |
y=−0.003198×+34244 | -0.005 | 0.869 |
|
| SKCM | Negative |
y=−0.01340×+62900 | -0.157 | 0.360 |
|
| LUAD | Negative | y=−0.04393×
+181872 | -0.236 |
8.88×10−16 |
|
| HNSC | Negative |
y=−0.01328×+136161 | -0.053 | 0.232 |
|
| UCEC | Negative |
y=−0.009837×+30808 | -0.397 |
2.21×10−5 |
|
| CESC | Negative |
y=−0.0433×+177708 | -0.151 | 0.008 |
| Ku70 | COAD | Negative |
y=−0.003319×+38104 |
4.81×10−4 | 0.991 |
|
| BRCA | Positive |
y=0.0009455×+26915 | -0.025 | 0.374 |
|
| SKCM | Negative |
y=−0.01050×+71586 | -0.229 | 0.179 |
|
| LUAD | Positive |
y=0.001316×+118475 | -0.154 | 1.87
×10−7 |
|
| HNSC | Negative |
y=−0.01467×+165072 | -0.108 | 0.016 |
|
| UCEC | Positive | y=0.001272×
+10171 | -0.090 | 0.353 |
|
| CESC | Negative |
y=−0.008341×+141397 | -0.110 | 0.054 |
|
DNA-PKcs | COAD | Positive |
y=0.01038×+25588 | -0.108 | 0.014 |
|
| BRCA | Positive |
y=0.003123×+27447 | 0.047 | 0.098 |
|
| SKCM | Negative |
y=−0.01111×+45835 | -0.171 | 0.320 |
|
| LUAD | Positive |
y=0.01127×+117308 | -0.069 | 0.019 |
|
| HNSC | Negative |
y=−0.03491×+137903 | -0.035 | 0.432 |
|
| UCEC | Negative |
y=−0.008411×+16815 | -0.371 |
7.98×10−5 |
|
| CESC | Negative |
y=−0.1111×+163343 | -0.074 | 0.198 |
| XRCC4 | COAD | Positive |
y=0.1244×+19506 | 0.069 | 0.116 |
|
| BRCA | Positive |
y=0.03546×+26440 | 0.077 | 0.007 |
|
| SKCM | Positive |
y=0.5896×+763.2 | 0.280 | 0.098 |
|
| LUAD | Positive |
y=0.1271×+111566 | 0.062 | 0.036 |
|
| HNSC | Positive |
y=0.3040×+87953 | 0.175 |
8.04×10−5 |
|
| UCEC | Negative | y=−0.02123×
+14814 | -0.233 | 0.014 |
|
| CESC | Negative |
y=−0.1882×+137753 | 0.142 | 0.012 |
| LIG4 | COAD | Positive |
y=0.07266×+25129 | 0.022 | 0.615 |
|
| BRCA | Positive |
y=0.08249×+23007 | 0.078 | 0.006 |
|
| SKCM | Positive |
y=0.08564×+33814 | 0.102 | 0.552 |
|
| LUAD | Positive |
y=0.1574×+109242 | 0.099 | 0.001 |
|
| HNSC | Positive |
y=0.08293×+110697 | 0.097 | 0.029 |
|
| UCEC | Negative | y=−0.02302×
+14377 | -0.009 | 0.924 |
|
| CESC | Negative |
y=−0.07419×+122540 | -0.029 | 0.605 |
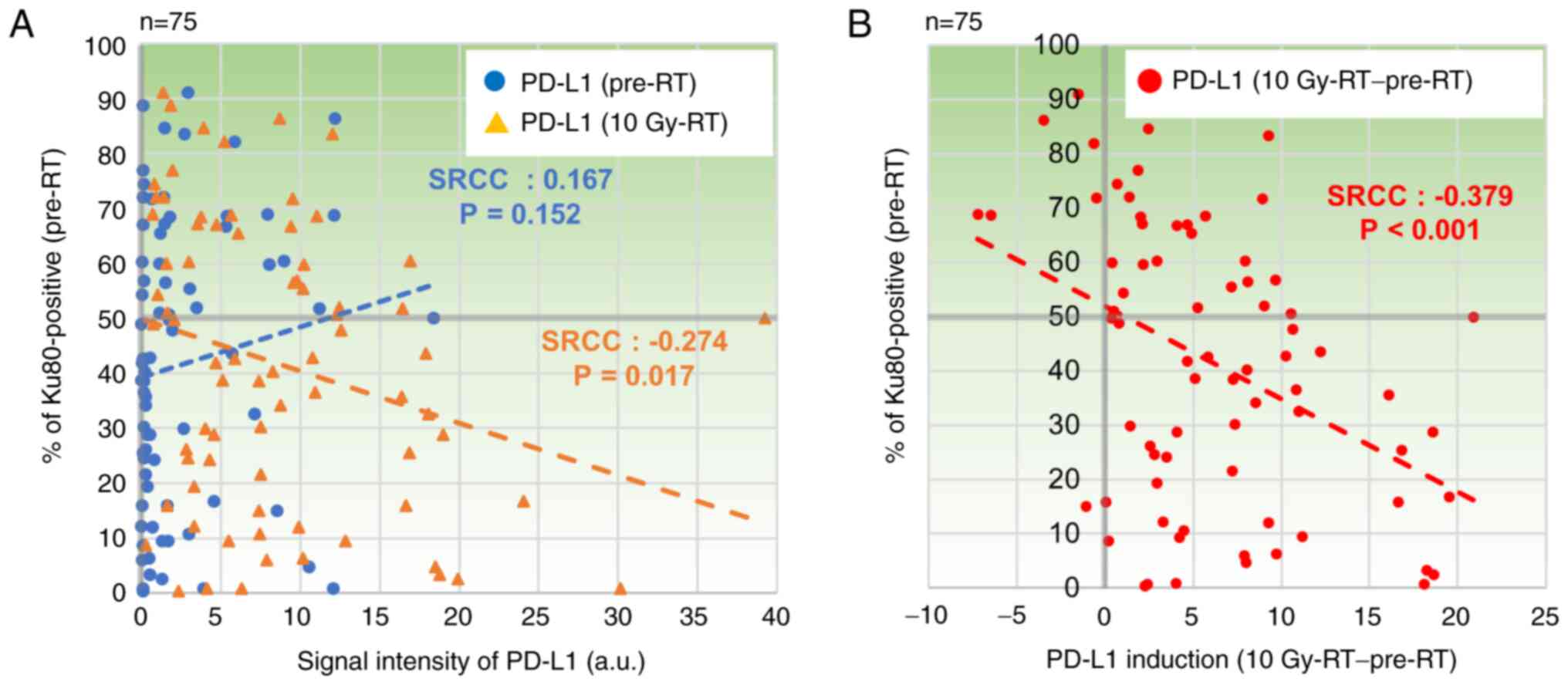
PD-L1 expression is negatively
correlated with Ku80 expression in cervical squamous cell carcinoma
samples treated with radiotherapy
Our previous study reported that PD-L1 expression is
up-regulated in cervical squamous cell carcinoma specimens after
being treated with 10 Gy of radiotherapy (29). The present study investigated
whether Ku80 expression was correlated with PD-L1 expression in the
clinical sample sets with or without radiotherapy. Ku80 and PD-L1
expression levels were quantitatively measured in
immunohistochemical stained specimens using ImageJ and QuPath
(Figs. S1 and S2). Although there was no obvious
correlation between Ku80 and PD-L1 in pre-RT specimens [Spearman's
rank correlation coefficient (SRCC)=0.167, P=0.152; Fig. 2A], PD-L1 in 10 Gy-RT exhibited a
weak negative correlation with Ku80 positivity in pre-RT specimens
(SRCC=−0.274, P=0.017; Fig. 2A).
Next, to investigate the impact of radiotherapy on PD-L1 expression
in the context of Ku80 positivity, PD-L1 changes in 10 Gy-RT were
plotted against Ku80 positivity of pre-RT specimens. Notably, a
significant negative correlation between Ku80 positivity of pre-RT
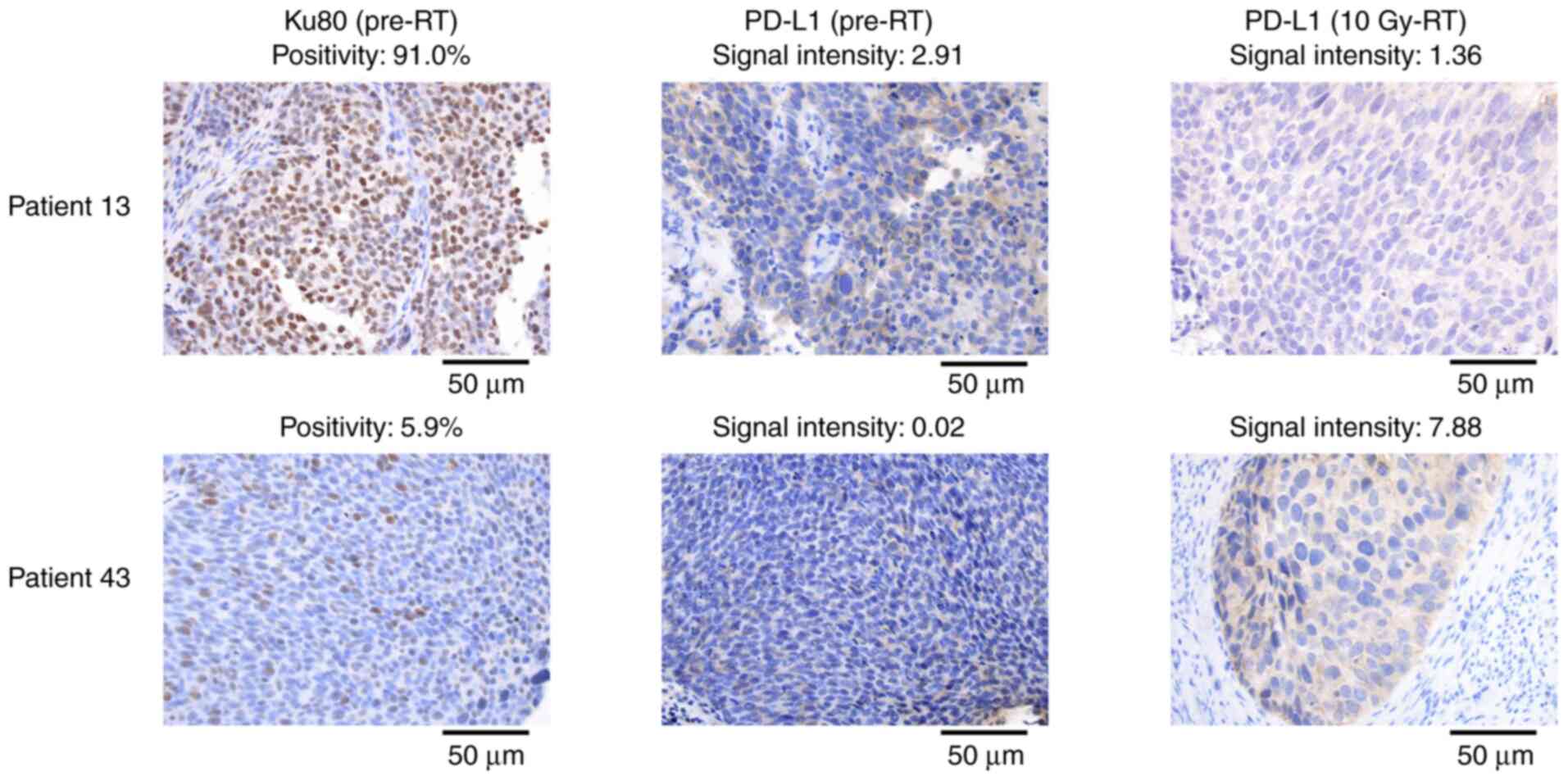
and PD-L1 changes was revealed (SRCC=−0.379; P<0.001; Fig. 2B). Representative images are shown
in Fig. 3. To validate the
computational quantification analysis, the same analysis scoring
was performed using a manual classification (TPS) in the same
specimens. Similar to the results presented in Fig. 2B, a negative correlation was
observed between Ku80 positivity of pre-RT and PD-L1 changes after
10 Gy-RT (Fig. S3). In addition,
the potential association between Ku80 positivity and clinical
outcomes was examined. Overall survival, progression-free survival
and local control did not exhibit significant differences with Ku80
expression (Fig. S4). These data
suggested that PD-L1 expression after 10 Gy-RT was negatively
correlated with Ku80, whereas there was no correlation between
PD-L1 expression and patients' outcome.
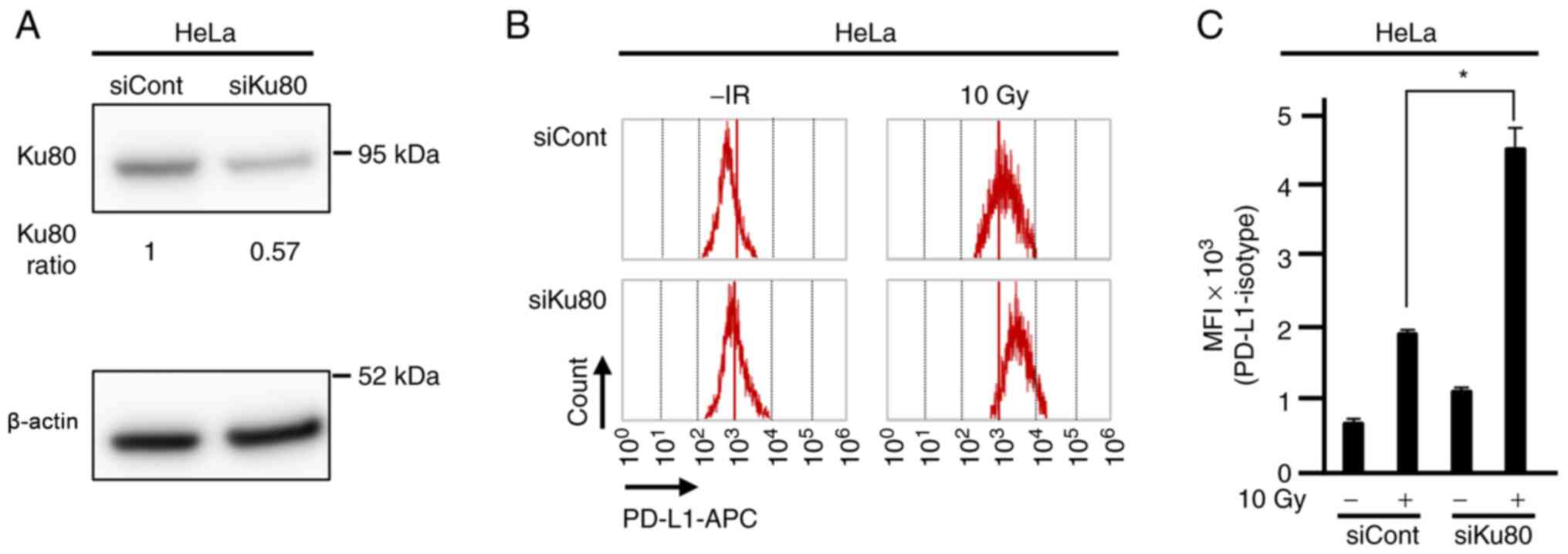
PD-L1 up-regulation after irradiation
is enhanced in Ku80-depleted HeLa cells
To confirm radiation-induced PD-L1 up-regulation in
a Ku80-depleted cervical cancer cell line in vitro, flow
cytometry analysis was performed in HeLa cells transfected with
Ku80 siRNA to examine the levels of cell-surface PD-L1 expression
after X-ray irradiation. Consistent with the immunohistochemistry
results seen in the tumour specimens, PD-L1 up-regulation induced
by 10 Gy X-ray irradiation was significantly increased in
Ku80-depleted cells compared with controls treated with X-ray
irradiation (Figs. 4, S5 and S6), suggesting that cell-surface PD-L1
expression was enhanced in Ku80-depleted condition.
Discussion
The present study revealed that Ku80 mRNA
expression in tumours was negatively correlated with PD-L1
expression using TCGA database analysis. Notably, in the clinical
sample sets from the present study, PD-L1 induction after 10 Gy-RT
was inversely correlated with Ku80 expression in pre-RT specimens.
In addition, significant up-regulation of PD-L1 expression after
irradiation was confirmed in Ku80-depleted HeLa cells compared with
irradiated controls, supporting supporting the hypothesis that low
Ku80 expression can affect PD-L1 up-regulation after
radiotherapy.
Our previous studies reported that DNA damage
signalling induced by irradiation or oxidative damage up-regulates
PD-L1 expression at the transcriptional level (14,30).
The increase in PD-L1 mRNA transcription consequently leads
to enhanced PD-L1 presentation on the cell surface. This
up-regulation is dependent on ATR/Chk1 (14). Since ATR/Chk1 activation is
associated with the progression of DSB end resection (high
resection causes greater ATR/Chk1 signalling), the magnitude of
resection is correlated with PD-L1 expression (31,32).
Consistent with this, our previous study revealed that Ku80
depletion, which causes greater DSB end resection, leads to greater
ATR/Chk1 signalling and PD-L1 expression in cancer cell lines such
as U2OS cells (14).
The present study revealed that Ku80-depleted HeLa
cells also expressed greater PD-L1 after irradiation. Together with
the findings of TCGA analysis, which demonstrated that low
Ku80 mRNA expression was correlated with high PD-L1
expression in all the cancer types analysed in this study (colon
adenocarcinoma, breast invasive carcinoma, skin cutaneous melanoma,
lung adenocarcinoma, head and neck squamous cell carcinoma, uterine
corpus endometrial carcinoma, cervical squamous cell carcinoma and
endocervical adenocarcinoma), Ku80 expression levels were
associated with PD-L1 expression, particularly when cells were
exposed to genotoxic stress. The results also suggested that
greater PD-L1 up-regulation can occur in patients who have received
radiotherapy if Ku80 expression is low.
To date, multiple pathways regarding PD-L1
expression after DNA damage have been suggested, such as cytosolic
sensors cGAS/RIG-I transducing immune-activating signal releasing
interferon stimulate genes following the recognition of cytosolic
DNA/RNA released from the nucleus (10–12).
Since radiotherapy can generate micronuclei that should contain
DNA/RNA (33), further studies are
required to examine the involvement of this pathway.
The present study quantified the signal intensity of
PD-L1 using software. Although the signals from inside the cell or
the cell surface were unable to be differentiated, which seems to
be important to detect functional PD-L1 on cell surface, the
software approach has advantages in measuring an interval scale of
their expression. PD-L1 expression was also examined using a manual
classification method scored by cell-surface staining for PD-L1.
Notably, the present study obtained similar results using both
methods. Thus, it was hypothesized that this computational
measurement could be a useful tool to quantify PD-L1 expression;
however, it must be carefully accessed depending on the type of
targets.
Limitations of the present study included the
limited number of clinical samples. As there are multiple
regulators of PD-L1 expression (34), other regulators may also affect the
PD-L1 expression after RT in clinical samples. Data variability may
be minimised by analysing the expression of PD-L1 in subgroups
classified by regulators of PD-L1 expression other than Ku80.
Future studies involving larger cohorts may help in further
elucidating the relationship between PD-L1 and Ku80 expression in
clinical settings. In addition, clinical trials are required to
analyse whether the increase in PD-L1 after RT in clinical tumours
with low Ku80 expression affects ICI treatment in the future.
Although overexpression of Ku80 was not examined in the present
study, it should be examined in future studies to determine the
magnitude of Ku80 expression; for example, low Ku80 expression
compared with normal levels was associated with PD-L1 up-regulation
after X-ray irradiation as indicated in the present study.
Recent radiotherapy technologies are highly
developed and enable radiation oncologists to target cancer using
high-precision radiotherapy, while also avoiding irradiation to the
surrounding normal tissue. These technological developments
increase the importance of controlling tumour cells outside the
irradiation field for long-term survival of patients (35). For several decades, radiation has
been considered a potent inducer of immune activation (36); however, the clinical outcomes of
patients treated with combination therapy differ per case, and the
mechanisms underlying this difference need to be investigated.
Therefore, the radiotherapy-dependent immune stimuli are still
unknown.
Notably, the situation of radiotherapy, such as dose
fractionation and radiation quality, affects immune responses. For
example, hypofractionated radiation at 8–12 Gy per fraction
activates the cGAS/STING pathway more effectively compared with a
higher single dose of ≥20 Gy (11). Also, our recent study reported that
PD-L1 up-regulation is more significantly induced by high linear
energy transfer carbon-ion irradiation compared with X-rays
(27). Hence, the reaction of the
immune system after radiotherapy is comprehensive. To identify the
optimal modality for radiation/ICI combination therapy, further
research regarding optimal dose/fractionation, field settings,
combination timing and type of irradiation are required. By
contrast, recent studies have demonstrated that radiotherapy also
up-regulates PD-L1 in tumours (34,37–39);
therefore, combination therapy with ICIs is promising. Indeed,
phase III clinical trials have revealed that combination therapy
improves clinical outcomes in patients with locally advanced
non-small cell lung cancer or metastatic castration-resistant
prostate cancer (40–43). However, this may not be applicable
to all patients with cancer from the viewpoint of precision
medicine; for example, PD-L1 is not effectively up-regulated in
some patients with certain genetic backgrounds, such as those
without Ku80 mutation, as indicated in the present study.
The present study demonstrated that PD-L1 expression
is induced by radiotherapy and can be affected by the expression
levels of Ku80 prior to radiotherapy. The present study did not
reveal any differences in Ku80 positivity with patient outcomes,
which may be because of the multiple functions of Ku80 that are
involved in both tumour immunity and DNA repair (14,44).
Furthermore, PD-L1 expression is not significantly correlated with
clinical outcomes in the same cohort in our previous study
(29). Although whether PD-L1
expression can be a predictive marker of ICIs is still debatable,
the findings of the present study suggest that Ku80 may be a
potential biomarker for radiation/ICI combination therapy. Further
studies into this type of approach will provide important data to
determine the best strategy for radiation/ICI combination
therapy.
Supplementary Material
Supporting Data
Acknowledgements
The authors thank Dr Yuya Yoshimoto (Fukushima
Medical University, Fukushima; Gunma University, Maebashi, Japan)
for his assistance in performing the immunohistochemical analysis.
The authors thank Mr Koji Isoda (Gunma University, Maebashi, Japan)
for his technical assistance in performing the immunohistochemical
analysis.
Funding
This study was supported by JSPS KAKENHI (grant nos. JP17H04713
and JP19K08195), the Takeda Science Foundation, the Uehara Memorial
Foundation, the Astellas Foundation for Research on Metabolic
Disorders, The Kanae Foundation for the Promotion of Medical
Science, the Yasuda Memorial Medicine Foundation and the Nakajima
Foundation. This study was also supported by the Program of the
Network-Type Joint Usage/Research Centre for Radiation Disaster
Medical Science of Hiroshima University, Nagasaki University and
Fukushima Medical University and the Grants-in-Aid from the
Ministry of Education, Culture, Sports, Science and Technology of
Japan for programmes for Leading Graduate Schools, Cultivating
Global Leaders in Heavy Ion Therapeutics and Engineering.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
TK, YM, HS, TBMP, YU, KO, SK, KS, HI, HY and AS
performed the experiments and analysed the data. TK, TBMP, HS and
AS wrote the manuscript. TK, YM, SN and KO coordinated the clinics,
carried out the treatment and participated in the patient
follow-up. SG, TN and TO contributed conception and design, and
reagents/materials/analysis tools and gave final approval for the
manuscript version to be published. TK and HS confirm the
authenticity of all the raw data. HS and TO supervised the study
with the support from AS. All authors have read and approved the
final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Review Board for clinical trials of Gunma University (approval no.
HS2020-015; Maebashi, Japan). The protocol is described on the
hospital website, and subjects were provided the opportunity to
opt-out; therefore, no additional consent was required from
patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
BRCA
|
breast invasive carcinoma
|
|
CESC
|
cervical squamous cell carcinoma and
endocervical adenocarcinoma
|
|
COAD
|
colon adenocarcinoma
|
|
DSB
|
double-strand break
|
|
HNSC
|
head and neck squamous cell
carcinoma
|
|
ICI
|
immune checkpoint inhibitors
|
|
LUAD
|
lung adenocarcinoma
|
|
MFI
|
mean fluorescence intensity
|
|
NHEJ
|
non-homologous end joining
|
|
SKCM
|
skin cutaneous melanoma
|
|
STING
|
stimulator of IFN genes
|
|
TCGA
|
The Cancer Genome Atlas
|
|
TPS
|
tumour proportion score
|
|
UCEC
|
uterine corpus endometrial
carcinoma
|
|
ATR
|
ataxia telangiectasia and
Rad3-related
|
|
Chk1
|
checkpoint kinase 1
|
|
DNA-PKcs
|
DNA-dependent protein kinase,
catalytic subunit
|
|
XRCC4
|
X-ray repair cross-complementing
protein 4
|
|
LIG4
|
DNA ligase 4
|
References
|
1
|
Sharma P and Allison JP: Immune checkpoint
targeting in cancer therapy: Toward combination strategies with
curative potential. Cell. 161:205–214. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Galon J and Bruni D: Approaches to treat
immune hot, altered and cold tumours with combination
immunotherapies. Nat Rev Drug Discov. 18:197–218. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yap TA, Parkes EE, Peng W, Moyers JT,
Curran MA and Tawbi HA: Development of immunotherapy combination
strategies in cancer. Cancer Discov. 11:1368–1397. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Guillem JG, Chessin DB, Cohen AM, Shia J,
Mazumdar M, Enker W, Paty PB, Weiser MR, Klimstra D, Saltz L, et
al: Long-term oncologic outcome following preoperative combined
modality therapy and total mesorectal excision of locally advanced
rectal cancer. Ann Surg. 241:829–836; discussion 836–838. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Golden EB, Frances D, Pellicciotta I,
Demaria S, Helen Barcellos-Hoff M and Formenti SC: Radiation
fosters dose-dependent and chemotherapy-induced immunogenic cell
death. Oncoimmunology. 3:e285182014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Diamond JM, Vanpouille-Box C, Spada S,
Rudqvist NP, Chapman JR, Ueberheide BM, Pilones KA, Sarfraz Y,
Formenti SC and Demaria S: Exosomes Shuttle TREX1-Sensitive
IFN-Stimulatory dsDNA from irradiated cancer cells to DCs. Cancer
Immunol Res. 6:910–920. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rodriguez-Ruiz ME, Vanpouille-Box C,
Melero I, Formenti SC and Demaria S: Immunological mechanisms
responsible for Radiation-Induced Abscopal effect. Trends Immunol.
39:644–655. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Reits EA, Hodge JW, Herberts CA, Groothuis
TA, Chakraborty M, Wansley EK, Camphausen K, Luiten RM, de Ru AH,
Neijssen J, et al: Radiation modulates the peptide repertoire,
enhances MHC class I expression, and induces successful antitumor
immunotherapy. J Exp Med. 203:1259–1271. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Formenti SC, Rudqvist NP, Golden E, Cooper
B, Wennerberg E, Lhuillier C, Vanpouille-Box C, Friedman K, Ferrari
de Andrade L, Wucherpfennig KW, et al: Radiotherapy induces
responses of lung cancer to CTLA-4 blockade. Nat Med. 24:1845–1851.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Harding SM, Benci JL, Irianto J, Discher
DE, Minn AJ and Greenberg RA: Mitotic progression following DNA
damage enables pattern recognition within micronuclei. Nature.
548:466–470. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Vanpouille-Box C, Alard A, Aryankalayil
MJ, Sarfraz Y, Diamond JM, Schneider RJ, Inghirami G, Coleman CN,
Formenti SC and Demaria S: DNA exonuclease Trex1 regulates
radiotherapy-induced tumour immunogenicity. Nat Commun.
8:156182017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yamazaki T, Kirchmair A, Sato A, Buqué A,
Rybstein M, Petroni G, Bloy N, Finotello F, Stafford L, Navarro
Manzano E, et al: Mitochondrial DNA drives abscopal responses to
radiation that are inhibited by autophagy. Nat Immunol.
21:1160–1171. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Feng X, Tubbs A, Zhang C, Tang M,
Sridharan S, Wang C, Jiang D, Su D, Zhang H, Chen Z, et al: ATR
inhibition potentiates ionizing radiation-induced interferon
response via cytosolic nucleic acid-sensing pathways. EMBO J.
39:e1040362020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sato H, Niimi A, Yasuhara T, Permata TBM,
Hagiwara Y, Isono M, Nuryadi E, Sekine R, Oike T, Kakoti S, et al:
DNA double-strand break repair pathway regulates PD-L1 expression
in cancer cells. Nat Commun. 8:17512017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Le DT, Durham JN, Smith KN, Wang H,
Bartlett BR, Aulakh LK, Lu S, Kemberling H, Wilt C, Luber BS, et
al: Mismatch repair deficiency predicts response of solid tumors to
PD-1 blockade. Science. 357:409–413. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Marabelle A, Le DT, Ascierto PA, Di
Giacomo AM, De Jesus-Acosta A, Delord JP, Geva R, Gottfried M,
Penel N, Hansen AR, et al: Efficacy of Pembrolizumab in patients
with noncolorectal high Microsatellite Instability/Mismatch
Repair-Deficient cancer: Results from the Phase II KEYNOTE-158
Study. J Clin Oncol. 38:1–10. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sunshine J and Taube JM: PD-1/PD-L1
inhibitors. Curr Opin Pharmacol. 23:32–38. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cancer Genome Atlas Network, .
Comprehensive molecular characterization of human colon and rectal
cancer. Nature. 487:330–337. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cancer Genome Atlas Network, .
Comprehensive molecular portraits of human breast tumours. Nature.
490:61–70. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cancer Genome Atlas Network, . Genomic
classification of cutaneous melanoma. Cell. 161:1681–1696. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cancer Genome Atlas Research Network, .
Comprehensive molecular profiling of lung adenocarcinoma. Nature.
511:543–550. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cancer Genome Atlas Network, .
Comprehensive genomic characterization of head and neck squamous
cell carcinomas. Nature. 517:576–582. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cancer Genome Atlas Research Network, .
Kandoth C, Schultz N, Cherniack AD, Akbani R, Liu Y, Shen H,
Robertson AG, Pashtan I, Shen R, et al: Integrated genomic
characterization of endometrial carcinoma. Nature. 497:67–73. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cancer Genome Atlas Research Network,
Albert Einstein College of Medicine Analytical Biological Services,
Barretos Cancer Hospital, Baylor College of Medicine, Beckman
Research Institute of City of Hope, Buck Institute for Research on
Aging, Canada's Michael Smith Genome Sciences Centre, Harvard
Medical School, Helen F. Graham Cancer Center & Research
Institute at Christiana Care Health Services, et al:
Integrated genomic and molecular characterization of cervical
cancer. Nature. 543:378–384. 2017.PubMed/NCBI
|
|
25
|
Pecorelli S: Revised FIGO staging for
carcinoma of the vulva, cervix, and endometrium. Int J Gynaecol
Obstet. 105:103–104. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ebina Y, Mikami M, Nagase S, Tabata T,
Kaneuchi M, Tashiro H, Mandai M, Enomoto T, Kobayashi Y, Katabuchi
H, et al: Japan Society of Gynecologic Oncology guidelines 2017 for
the treatment of uterine cervical cancer. Int J Clin Oncol.
24:1–19. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Permata TBM, Sato H, Gu W, Kakoti S,
Uchihara Y, Yoshimatsu Y, Sato I, Kato R, Yamauchi M, Suzuki K, et
al: High linear energy transfer carbon-ion irradiation upregulates
PD-L1 expression more significantly than X-rays in human
osteosarcoma U2OS cells. J Radiat Res. 62:773–781. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shrivastav M, De Haro LP and Nickoloff JA:
Regulation of DNA double-strand break repair pathway choice. Cell
Res. 18:134–147. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mori Y, Sato H, Kumazawa T, Permata TBM,
Yoshimoto Y, Murata K, Noda SE, Kaminuma T, Ando K, Oike T, et al:
Analysis of radiotherapy-induced alteration of CD8+ T
cells and PD-L1 expression in patients with uterine cervical
squamous cell carcinoma. Oncol Lett. 21:4462021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Permata TBM, Hagiwara Y, Sato H, Yasuhara
T, Oike T, Gondhowiardjo S, Held KD, Nakano T and Shibata A: Base
excision repair regulates PD-L1 expression in cancer cells.
Oncogene. 38:4452–4466. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jazayeri A, Falck J, Lukas C, Bartek J,
Smith GC, Lukas J and Jackson SP: ATM- and cell cycle-dependent
regulation of ATR in response to DNA double-strand breaks. Nat Cell
Biol. 8:37–45. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rhind N: Changing of the guard: How ATM
hands off DNA double-strand break signaling to ATR. Mol Cell.
33:672–674. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kobayashi D, Oike T, Murata K, Irie D,
Hirota Y, Sato H, Shibata A and Ohno T: Induction of micronuclei in
cervical cancer treated with radiotherapy. J Pers Med. 10:1102020.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sato H, Jeggo PA and Shibata A: Regulation
of programmed death-ligand 1 expression in response to DNA damage
in cancer cells: Implications for precision medicine. Cancer Sci.
110:3415–3423. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chetty IJ, Martel MK, Jaffray DA, Benedict
SH, Hahn SM, Berbeco R, Deye J, Jeraj R, Kavanagh B, Krishnan S, et
al: Technology for innovation in radiation oncology. Int J Radiat
Oncol Biol Phys. 93:485–492. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lhuillier C, Rudqvist NP, Elemento O,
Formenti SC and Demaria S: Radiation therapy and anti-tumor
immunity: Exposing immunogenic mutations to the immune system.
Genome Med. 11:402019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dovedi SJ, Adlard AL, Lipowska-Bhalla G,
McKenna C, Jones S, Cheadle EJ, Stratford IJ, Poon E, Morrow M,
Stewart R, et al: Acquired resistance to fractionated radiotherapy
can be overcome by concurrent PD-L1 blockade. Cancer Res.
74:5458–5468. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Patel KR, Martinez A, Stahl JM, Logan SJ,
Perricone AJ, Ferris MJ, Buchwald ZS, Chowdhary M, Delman KA,
Monson DK, et al: Increase in PD-L1 expression after pre-operative
radiotherapy for soft tissue sarcoma. Oncoimmunology.
7:e14421682018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lim SH, Hong M, Ahn S, Choi YL, Kim KM, Oh
D, Ahn YC, Jung SH, Ahn MJ, Park K, et al: Changes in tumour
expression of programmed death-ligand 1 after neoadjuvant
concurrent chemoradiotherapy in patients with squamous oesophageal
cancer. Eur J Cancer. 52:1–9. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Antonia SJ, Villegas A, Daniel D, Vicente
D, Murakami S, Hui R, Yokoi T, Chiappori A, Lee KH, de Wit M, et
al: Durvalumab after chemoradiotherapy in Stage III Non-Small-Cell
lung cancer. N Engl J Med. 377:1919–1929. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Antonia SJ, Villegas A, Daniel D, Vicente
D, Murakami S, Hui R, Kurata T, Chiappori A, Lee KH, de Wit M, et
al: Overall survival with Durvalumab after chemoradiotherapy in
Stage III NSCLC. N Engl J Med. 379:2342–2350. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kwon ED, Drake CG, Scher HI, Fizazi K,
Bossi A, van den Eertwegh AJ, Krainer M, Houede N, Santos R,
Mahammedi H, et al: Ipilimumab versus placebo after radiotherapy in
patients with metastatic castration-resistant prostate cancer that
had progressed after docetaxel chemotherapy (CA184-043): A
multicentre, randomised, double-blind, phase 3 trial. Lancet Oncol.
15:700–712. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ludmir EB, McCaw ZR, Wei LJ, Re: Karim
Fizazi, Charles G. Drake, Tomasz M. Beer, et al: Final Analysis of
the Ipilimumab Versus Placebo Following Radiotherapy Phase III
Trial in Postdocetaxel Metastatic Castration-resistant Prostate
Cancer Identifies an Excess of Long-term Survivors. Eur Urol. In
press. https://doi.org/10.1016/j.eururo.2020.07.032Interpreting
the effect of Ipilimumab following radiotherapy for patients with
Postdocetaxel metastatic Castration-resistant Prostate cancer. Eur
Urol, 79, e10-e11, 2021. PubMed/NCBI
|
|
44
|
Shibata A and Jeggo PA: DNA double-strand
break repair in a cellular context. Clin Oncol (R Coll Radiol).
26:243–249. 2014. View Article : Google Scholar : PubMed/NCBI
|